|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
大叶藻对氮磷营养盐的吸收动力学特征
植物生态学报
2020, 44 (7):
772-781.
DOI: 10.17521/cjpe.2019.0335
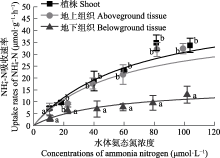
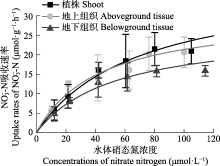
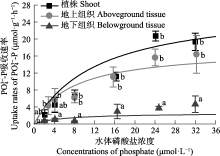
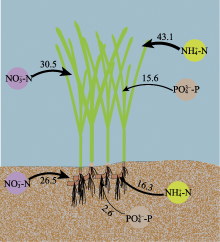
营养盐是影响海草生长的关键因子, 目前有关海草不同组织对不同形式氮和磷的吸收特征尚不明确。该研究通过利用海草地上和地下组织分隔培养装置, 设置不同的氨态氮、硝态氮和磷酸盐浓度, 探究了大叶藻(Zostera marina)植株及其地上和地下组织对氮磷营养盐的吸收动力学特征。结果显示: (1)大叶藻对氮磷的吸收符合饱和吸收动力学特征, 吸收速率和水体氮磷浓度可用米式方程描述; (2)大叶藻植株对NH4+-N的最大吸收速率(Vmax, 52 μmol·g-1·h-1)显著高于其对NO3--N的Vmax (39 μmol·g-1·h-1); (3)大叶藻地上组织和地下组织均可吸收氮磷, 但地上组织对氨态氮、硝态氮、磷酸盐的Vmax分别为43.1、30.5和15.6 μmol·g-1·h-1, 为地下组织的2.6、1.2和6倍。结果表明, 大叶藻对氨态氮的吸收能力高于硝态氮, 且对氮磷的吸收可能主要依赖地上组织(叶片)。结果为查明大叶藻对氮磷的吸收利用机制及评估大叶藻的海洋生态效应提供了理论依据。
表3
海草和藻类对NH4+-N、NO3--N和PO43--P的吸收动力学参数
正文中引用本图/表的段落
大叶藻在寡营养盐条件下具有明显的竞争优势, 减少氮营养盐的输入则可以明显促进退化海草床的自我恢复(Greening & Janicki, 2006; Sandoval-Gil et al., 2019)。富营养化会对大叶藻产生显著的负效应(Campbell et al., 2018; Li et al., 2019), 如van Katwijk等(1997)研究发现, 北半球富营养化水域大叶藻海草床的退化主要归咎于水体氨态氮的毒性。高氮环境下, 大叶藻植株的存活、生长和碳贮存均受到显著抑制(Villazán et al., 2015; Darnell & Dunton, 2017)。与藻类相比, 大叶藻对NH4+-N和NO3--N吸收的Vmax值较低, 而对PO43--P的Vmax较高(表3)。这种低氮高磷的吸收特征表明, 大型海藻的引入可能会导致大叶藻海草床的退化, 大型藻类能够抢先利用氮进行自身的快速繁殖, 进而造成大叶藻的氮营养盐限制, 因此在大叶藻海草床的修复过程中, 应警惕藻类的爆发, 一旦藻类大量繁殖, 不仅会减弱水下光照, 影响海草的光合作用, 还会掠夺海水中的营养盐, 限制大叶藻对氮营养盐的利用(Phillips et al., 1978; Orth & van Montfrans, 1984; Haglund & Pedersén, 1993)。
此外, 植株的吸收动力学特征受到环境光照、温度和盐度等环境条件的影响。光照的影响主要体现在植株的光合作用和酶活性等方面, 如有研究表明, 水生植物在光照条件下对营养盐的吸收速率远大于其在黑暗条件下的吸收速率, 光照还能激活水生植物体内的碱性磷酸酶活性, 为植物吸收利用磷酸盐提供条件(Weich & Granéli, 1989; 黄邦钦等, 1999; 刘长发等, 2001)。温度则主要通过影响植株的呼吸作用及光合作用暗反应的酶活性来影响植株对氮的吸收(Lomas & Glibert, 1999)。研究发现, 温度升高时, 龟裂泰来草(Thalassia testudinum)对NH4+-N和NO3--N的Vmax值也随之提高(Lee & Dunton, 1999)。同时, 光照和温度的交互作用也会对水生植物的营养盐吸收过程产生影响, 如程丽巍等(2010)研究表明, 在适温范围时, 水生植物对NO3--N的吸收速率随光强的增加而增加, 但在高温条件下, 光强的升高并不能促进营养盐的吸收。盐度也是调节水生植物营养盐吸收过程的关键因子, 低盐度条件下植物对NH4+-N的吸收速率较高(刘静雯和董双林, 2001)。
Km, 半饱和常数; Vmax, 最大吸收速率. ... Regulation of nitrate uptake by the seagrassZostera marina during upwelling. 1 2019 ... 大叶藻在寡营养盐条件下具有明显的竞争优势, 减少氮营养盐的输入则可以明显促进退化海草床的自我恢复( Extinction risk assessment of the world’s seagrass species 2 2011 ... 海草是一类海洋高等单子叶开花植物, 具有极高的初级生产力, 在我国主要分布于温带的山东、辽宁、河北, 亚热带、热带的广东、广西和海南等沿岸海域(
Km, 半饱和常数; Vmax, 最大吸收速率. ... Accelerating loss of seagrasses across the globe threatens coastal ecosystems 1 2009 ... 海草床生态系统较脆弱, 受全球变化和人类活动影响, 目前已是地球生物圈退化速率最快的生态系统, 且退化速率仍在逐年增加( Extracellular alkaline phosphatase activity in Ulva lactuca L 1 1989 ... 此外, 植株的吸收动力学特征受到环境光照、温度和盐度等环境条件的影响.光照的影响主要体现在植株的光合作用和酶活性等方面, 如有研究表明, 水生植物在光照条件下对营养盐的吸收速率远大于其在黑暗条件下的吸收速率, 光照还能激活水生植物体内的碱性磷酸酶活性, 为植物吸收利用磷酸盐提供条件( 真江蓠对氨氮去除效率与吸收动力学研究 1 2008 ... Uptake kinetics of NH4+-N, NO3--N and PO43--P by seagrasses and seaweeds
本文的其它图/表
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||