|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
山杏叶片光合生理参数对土壤水分和光照强度的阈值效应
植物生态学报
2011, 35 (3):
322-329.
DOI: 10.3724/SP.J.1258.2011.00322
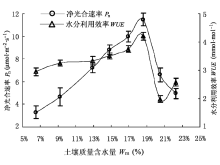
以半干旱黄土丘陵区主要灌木树种山杏(Prunus sibirica)为试验材料, 应用CIRAS-2型光合作用仪测定不同土壤质量含水量(Wm)下山杏叶片净光合速率(Pn)、蒸腾速率(Tr)及水分利用效率(WUE)的光响应过程, 探讨山杏光合特性对土壤水分和光照条件的适应性。结果表明: Pn、Tr及WUE对Wm和光照强度的变化有明显的阈值响应。随着Wm (6.5%-18.6%)的递增, 光补偿点降低, 光饱和点、表观量子效率和最大净光合速率均升高; 在Wm为18.6%时, 山杏利用弱光和强光的能力最强, 光照生态幅最宽。随着Wm (9.2%-18.6%)的递增, Pn、Tr有明显升高的趋势, 水分过高或过低, 两者均呈现下降趋势; 山杏对光照环境的适应性较强, 在光合有效辐射为800-1 200 µmol∙m-2∙s-1时, Pn和WUE都具有较高水平, 饱和光强在983-1 365 µmol∙m -2∙s-1之间。以光合生理参数为指标对山杏土壤水分有效性及生产力进行分级与评价, 确定Wm < 9.2%或Wm > 22.3%时为“低产中效水”; Wm在20.5%-22.3%和9.2%-12.9%时, 分别为“中产低效水”和“中产中效水”; Wm在12.9%-20.5%时为“高产高效水”。其中Wm为18.6%时为“最佳产效水”, 对应光强为1 365 µmol∙m-2∙s-1。
表1
不同土壤水分条件下山杏叶片的光合生理参数(平均值±标准误差)
正文中引用本图/表的段落
在受控温室内选用3年生山杏苗木, 2009年3月4日进行苗木盆栽, 用环刀法测得盆栽试验土壤的田间持水量为27.5%, 土壤容重为1.21 g?cm-3, pH值为6.5-7.5, 肥力中等, 有机质、有效氮、有效磷、有效钾含量分别为1.12%、40.23、11.20、70.15 mg?kg-1。在整个试验期间, 受控温室内光强为外界自然光强的90%左右, 温度在18-30 ℃之间, 空气相对湿度在41%-65%之间, CO2浓度在345-385 μmol?mol-1之间。采用田间环境条件下人为给水和自然耗水相结合的方法, 获取盆栽土壤不同的水分梯度, 盆钵直径30 cm、深40 cm。具体做法是: 选取生长健壮较一致的山杏盆栽苗6株(每盆一株), 试验观测1天前(7月10日)浇水, 使土壤水分饱和, 利用MI2X型土壤水分探头(Delta-T Cambridge, UK)监测土壤含水量的变化, 并以烘干法进行补充完善。1天后获得初期土壤水分, 土壤质量含水量(Wm)为22.3%, 相对含水量(Wr)为81.8%, 进行第1次光合生理参数的测定。以后通过自然耗水, 每隔1-2天获取一个水分梯度, 7月25日测定结束, 共获得8个水分梯度(表1), 分别进行光合生理参数的光响应测定。
式中, Pn为净光合速率, α、β、γ是3个系数, 其中α为I = 0时光响应曲线的初始斜率, 可作为表观量子效率, 即AQY; β为修正系数; I为光合有效辐射, 即PAR; LCP为光补偿点; γ = α/Pnmax, Pnmax为最大净光合速率。依据该模拟方程, 利用统计分析软件(SPSS 12.0)进行非线性回归分析, 并通过求导换算(叶子飘, 2007), 可求出如下参数: 光补偿点(LCP; μmol?m-2?s-1)、光饱和点 (LSP; μmol?m-2?s-1)、暗呼吸速率(Rd)等参数(表1)。采用SPSS 12.0进行单因素方差分析和LSD对各处理间的差异显著性在5%水平上进行检验; 同时对土壤水分点进行聚类分析。
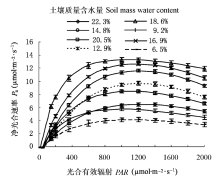
由表1可知, 山杏叶片AQY在水分胁迫与适宜水分条件下差异显著(p < 0.05), 并且在Wm为18.6%时, AQY达到最高, 表现出较强的光能利用潜力; Rd随着水分条件的不同差异显著(p < 0.05)。在水分胁迫下, 山杏幼苗有通过减少呼吸作用对光合产物的消耗, 以积累干物质的生理适应特性。随着Wm (12.9%-18.6%)的增加, LCP有降低趋势; 而在水分过高或者过低时LCP有增大趋势, 从而降低对弱光的利用效率, 可见山杏随着土壤水分的不同, 对弱光也表现出一定的适应性和可塑性。随着Wm的增加, LSP和Pnmax有增加趋势, Wm为18.6%时均达到最高, 随后LSP和Pnmax表现为降低趋势, 表明水分过高或者过低都不利于山杏对强光的利用, 其叶片最大光合能力也受到一定限制。分析可知, 维持较高Pn的Wm为12.9%-18.6%, 此水分范围内适宜的光强为800-1 400 μmol?m-2?s-1, 其中Pn最大值出现在Wm为18.6%左右, 对应光强为1 365 μmol?m-2?s-1。
在适宜的生长条件下实测的AQY, 一般植物在0.03-0.06之间(李合生, 2002), 山杏叶片在中等土壤肥力的适宜水分条件下, 最高可达0.083, 高于一般植物的AQY, 可见山杏在弱光下利用光能的潜力较大; 但水分亏缺和水分过多都会导致表观量子效率降低(表1), 可见适宜的水分条件在很大程度上能够提高植物的AQY。一般阳生植物LSP在540 μmol?m-2?s-1以上, LCP在13-36 μmol?m-2?s-1之间; 而阴生植物的LSP一般在90-l80 μmol?m-2?s-1之间, LCP在10 μmol?m-2?s-1以下(孟繁静, 2000)。可见山杏具有典型阳性植物的LSP和LCP, 在Wm为18.6%时, LCP最低, LSP最高, 表明在此中等土壤肥力的水分条件下, 山杏利用弱光和强光的能力较强, 光照生态幅最宽, 有利于有机物质的积累。土壤水分亏缺明显影响山杏叶片的LCP、LSP和AQY, 因而在相同光强下容易降低叶片光能利用率。
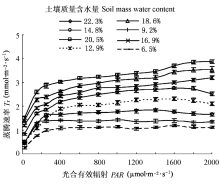
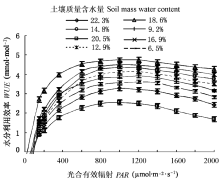
Pn和Tr在高水分点时, 对Wm表现出不同的响应规律, 随着Wm的递增Pn逐渐下降, 而Tr并不随Pn的降低而变化, 保持较高的Tr (图1, 图2); 同时Tr随着PAR上升其增幅程度相差很大, Tr的规律性变化势必会影响到WUE。造成山杏叶片Pn和Tr随Wm和PAR表现出不同变化趋势的原因, 可能是其自身体制的自我调节能力不同造成的, 也可能是受所处环境条件的影响; 而蒸腾作用的强弱主要取决于土壤中可利用的水分、所必需的能量以及叶片内外间存在的水势梯度, 也受植物体内部结构和生理状况的调节(张淑勇等, 2009)。表明山杏随着土壤湿度和光照强度的不同, 有规律地调节自身的生理活动, 以实现对水分和光照资源的充分利用。
本文的其它图/表
|