
|
|
||
|
三种藓类植物固碳耗水节律调节作用
植物生态学报
2023, 47 (7):
988-997.
DOI: 10.17521/cjpe.2022.0153
藓类植物作为缺乏维管组织的非等水植物, 其光合作用和耗水量主要受到光照、温度和水分供应状况的影响; 这些环境条件随昼夜交替改变, 是否使得藓类植物形成了类似维管植物的内在昼夜节律调节机制, 目前还所知甚少; 理解其节律响应特点对于藓类植物的保育与利用具有重要意义。该研究对脆枝青藓(Brachythecium thraustum)、大灰藓(Hypnum plumaeforme)和长叶提灯藓(Mnium lycopodioides)进行12 h/12 h的昼夜光周期驯化后进行24 h/0 h的持续光照处理, 分别测定3种藓类在12 h/12 h和24 h/0 h光周期条件下的净光合速率(Pn)和蒸腾速率(Tr), 结果表明: 排除光照和叶-气水汽压差(VPD)等环境影响之后, 3种藓类的这两个指标均观察到明显的节律变化, 节律调节作用能够解释其Pn和Tr日变化的23.4%和30.2%; 且维管植物中少见的Tr节律作用出现在所有受试的藓类植物上; Pn和Tr的节律响应存在明显的种间差异, 该研究中节律性最强的物种为脆枝青藓。以脆枝青藓为例进一步分析其全光照下非结构性碳水化合物(NSC)的节律响应时序转录组, 结果表明: 35.1%的淀粉、糖类合成代谢相关基因的表达具有节律性, 且与NSC含量的昼夜节律变化相关; 脱落酸信号和气孔调节相关基因的表达变化与Tr的节律调节相关; 光合电子传递和暗反应相关蛋白的转录水平变化与光合作用节律保持一致。藓类植物虽然丢失了生物钟核心反馈环CCA1/LHY基因, 但是核心生物钟功能集中于PRRs家族, 通过级联调节使得光合作用生物学过程和保卫细胞水分/离子运输过程等相关基因表达呈现显著的节律性, 最终导致固碳耗水的节律响应高度保守。 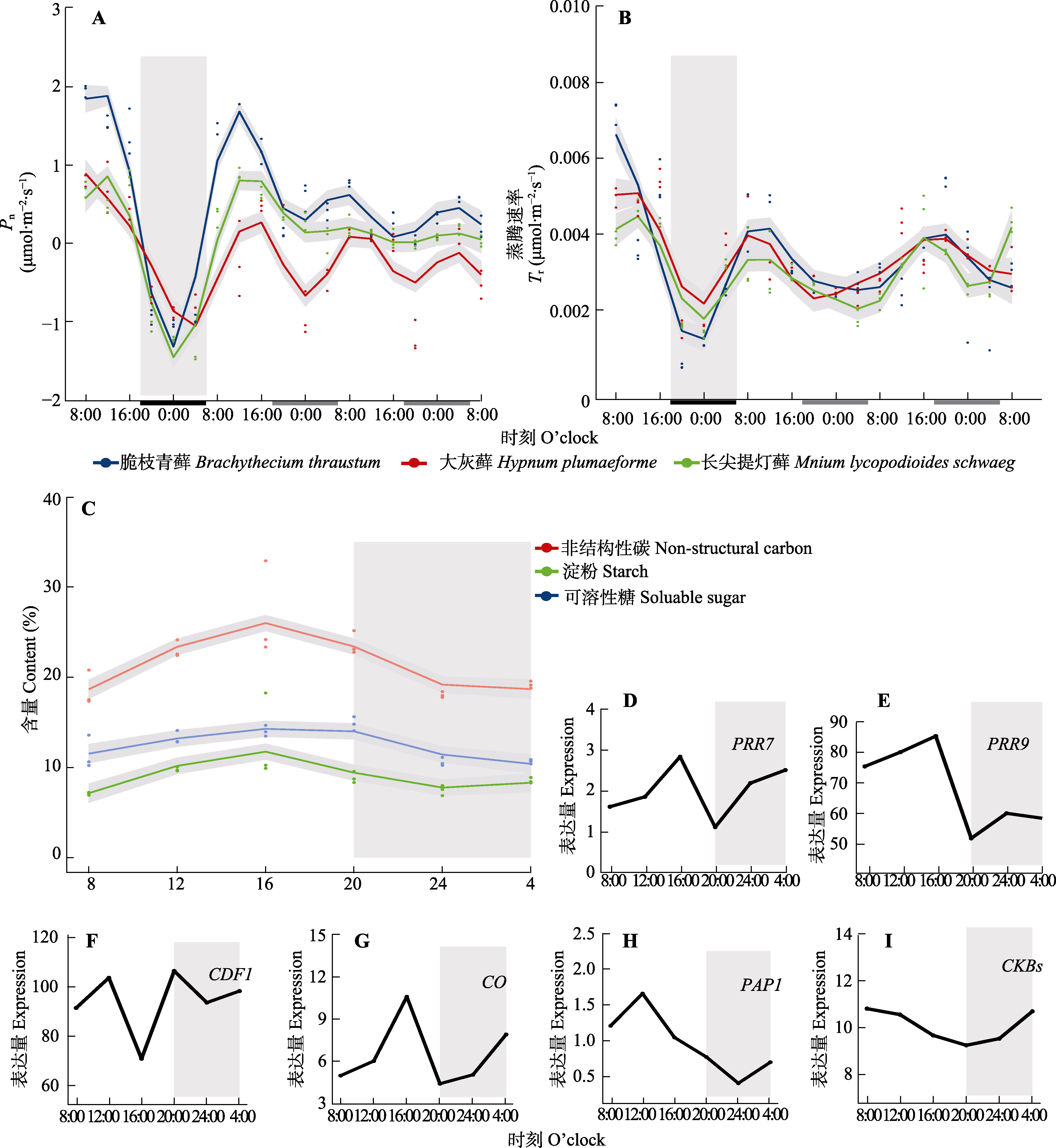 View image in article
图1
使用广义相加模型(GAM)拟合脆枝青藓、大灰藓和长叶提灯藓的净光合同化速率(Pn)、蒸腾速率(Tr)、非结构性碳水化合物和核心生物钟基因及节律调控相关基因昼夜节律。A、B中灰色部分对应于15 d正常光周期的最后一个夜晚。横坐标上的黑色和白色分别代表植物自然条件下的夜晚和白天, 灰色部分代表植物自然条件下黑暗但给予光照。C-I中灰色部分表示全光照条件下第二天(LL2d)外界条件下的夜晚。D-I分别为脆枝青藓生物钟调控相关基因的表达量。CDF1, CYCLING DOF FACTOR1; CKB, CASEIN KINASE II BETA CHAIN; CO, Constans; PAP1, PRODUCTION OF ANTHOCYANIN PIGMENT 1; PRR7, PSEUDO RESPONSE REGULATOR 7; PRR9, PSEUDO RESPONSE REGULATOR。
正文中引用本图/表的段落
3种藓类经过正常光周期(12 h/12 h)适应15天后的Pn (夜间无光照情况下表现为呼吸速率)均出现昼夜节律振荡(图1A), 但是种间差异明显, 其中脆枝青藓振幅最强而长叶提灯藓最弱。通过GAM拟合估算正常光周期条件下脆枝青藓的Pn从夜间最低-1.29 μmol·m?2·s?1到白天最高1.89 μmol·m?2·s?1。在恒定光照条件下(LL, 24 h/0 h)第1天(LL1d)脆枝青藓的Pn振荡范围为0.32-1.69 μmol·m?2·s?1, 是正常光周期的43.1%, 第2天(LL2d)脆枝青藓的Pn振幅占正常光周期的16.7%。同样, 大灰藓LL1d的振幅占正常光周期的48.2%, LL2d占30%。长叶提灯藓LL1d的振幅占正常光周期的1/3, 而LL2d昼夜节律振幅无显著变化。脆枝青藓正常光周期时Pn在正午/午夜时达到最高和最低峰值, LL1d延续这一趋势, 但是LL2d峰值前移4 h。而Tr的峰值与Pn相比延后4 h (图1B), 同样受到昼夜节律调控。脆枝青藓的Tr在LL1d/LL2d条件下分别占正常光周期的29.6%和25.9% (r = 0.92); 大灰藓分别占正常光周期的58.6%和34.4%; 长叶提灯藓在LL1d振幅占正常光周期的56.0%。
以拟南芥20个核心生物钟基因(CCA1、LHY、TOC1、PRR3、5、7、9、RVE1-8、LUX、ELF3/4, GI和ZTL)确定脆枝青藓的生物钟的直系同源基因及其表达特点。脆枝青藓具有4个核心生物钟基因家族的直系同源基因: 早间循环PRRs家族, 晚间循环ELF3/4、RVEs家族; 但缺少核心反馈环CCA1/LHY、TOC1的直系同源基因, 以及对TOC1等调节的GI、ZTL基因。如图1D、1E所示, 脆枝青藓核心生物钟基因PRRs在外界白天表达水平达到峰值, 具有显著的昼夜节律。而生物钟调控相关基因CYCLING DOF FACTOR 1 (CDF1)、Constans (CO)、PRODUCTION OF ANTHOCYANIN PIGMENT 1 (PAP1)和CASEIN KINASE II BETA CHAIN (CKB)同样具有显著的昼夜节律表达(图1F-1I)。
多数被子植物对光合作用具有节律响应, 蒸腾耗水作用缺乏节律性(Resco de Dios & Gessler, 2018); 然而本研究发现3种藓类植物均具有光合作用及蒸腾耗水节律。进一步采用脆枝青藓时序转录组分析, 发现超过一半的植物气孔运动相关的基因家族成员的表达具有较强节律性(图2A); 且这些基因表达与蒸腾耗水速率强相关, 表明蒸腾作用的节律性可以由气孔运动相关基因的节律调控进行解释。藓类植物是第一批脱离水生环境进入到陆地环境的陆生植物, 缺乏维管组织运输水分, 仅有1层细胞(极少情况下超过2层)的拟叶(即藓类植物的叶片)完成水分运输(Glime et al., 2007)。拟叶依靠气孔和细胞壁吸取水分并通过前者吸收CO2, 但缺少角质层保护, 因此对外界环境条件反应灵敏。除此前早已发现栖居环境中水分条件、光照对藓类的调节作用以外, 本研究表明藓类植物对于气孔周围组织的水分运输同样具有节律调节作用(图1B)。而且藓类植物气体交换指标(Pn和Tr)速率的节律调节振幅较强, 高于维管植物15%-25% (Resco de Dios & Gessler, 2018)的昼夜节律振幅。
本研究不仅观察到光合作用的节律变化, 光合电子传递链和暗反应相关蛋白的转录也具有严格的节律调节, 与拟南芥的全光照下时序转录组(Noordally et al., 2013)相似; 其中部分光合膜相关蛋白在凌晨出现表达峰值的内在机制值得进一步研究。固碳过程的节律性必然导致其最终产物NSC呈现严格的节律变化(图1C); 同时本研究表明淀粉合成和分解代谢相关基因的节律变化刚好相反(图2C), 与NSC变化完全一致。可溶性糖的变化同样具有节律性; Chia等(2004)验证了DPE2是夜间淀粉转化为蔗糖途径关键组分, 本研究观察到脆枝青藓DPE2同系物在夜间表达量升高。
本文的其它图/表
|

{kind=link}