
|
|
||
|
三种藓类植物固碳耗水节律调节作用
植物生态学报
2023, 47 (7):
988-997.
DOI: 10.17521/cjpe.2022.0153
藓类植物作为缺乏维管组织的非等水植物, 其光合作用和耗水量主要受到光照、温度和水分供应状况的影响; 这些环境条件随昼夜交替改变, 是否使得藓类植物形成了类似维管植物的内在昼夜节律调节机制, 目前还所知甚少; 理解其节律响应特点对于藓类植物的保育与利用具有重要意义。该研究对脆枝青藓(Brachythecium thraustum)、大灰藓(Hypnum plumaeforme)和长叶提灯藓(Mnium lycopodioides)进行12 h/12 h的昼夜光周期驯化后进行24 h/0 h的持续光照处理, 分别测定3种藓类在12 h/12 h和24 h/0 h光周期条件下的净光合速率(Pn)和蒸腾速率(Tr), 结果表明: 排除光照和叶-气水汽压差(VPD)等环境影响之后, 3种藓类的这两个指标均观察到明显的节律变化, 节律调节作用能够解释其Pn和Tr日变化的23.4%和30.2%; 且维管植物中少见的Tr节律作用出现在所有受试的藓类植物上; Pn和Tr的节律响应存在明显的种间差异, 该研究中节律性最强的物种为脆枝青藓。以脆枝青藓为例进一步分析其全光照下非结构性碳水化合物(NSC)的节律响应时序转录组, 结果表明: 35.1%的淀粉、糖类合成代谢相关基因的表达具有节律性, 且与NSC含量的昼夜节律变化相关; 脱落酸信号和气孔调节相关基因的表达变化与Tr的节律调节相关; 光合电子传递和暗反应相关蛋白的转录水平变化与光合作用节律保持一致。藓类植物虽然丢失了生物钟核心反馈环CCA1/LHY基因, 但是核心生物钟功能集中于PRRs家族, 通过级联调节使得光合作用生物学过程和保卫细胞水分/离子运输过程等相关基因表达呈现显著的节律性, 最终导致固碳耗水的节律响应高度保守。 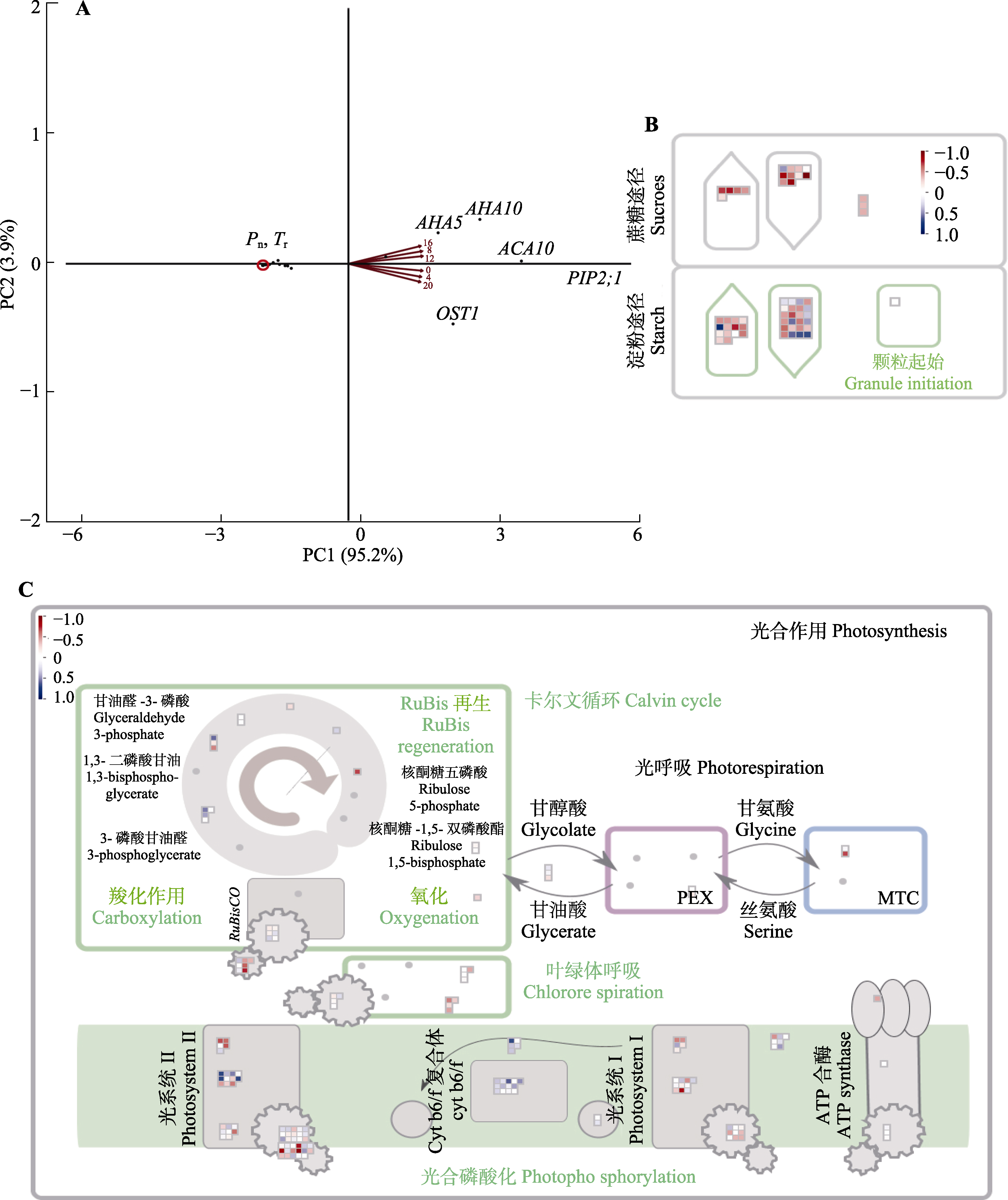 View image in article
图3
脆枝青藓相关基因的相关性分析。A, 15个显著的气孔相关节律性基因主成分(PC)分析图。B, 57个显著的碳固定相关节律性基因相关性图, 红色表示正相关, 蓝色表示负相关。C, 159个显著的光合相关节律性基因相关性图。热图采用了归一化表达量, 范围为-1到1。蓝色区域代表高表达量, 而红色区域则代表低表达量。ACA10, Ca2 +-ATP酶10; AHA5, 质膜H+-ATP酶; AHA10, H+-ATP酶10; MTC, PSII反应中心相关的膜内捕光复合物; OST1, 气孔开放因子1; PEX, 光系统II (PSII)外围蛋白质; PIP2;1, 水通道蛋白PIP2-1; Pn, 净光合速率; Tr, 蒸腾速率。
正文中引用本图/表的段落
气孔运动为水分耗散等气体交换提供条件; 研究气体交换作用与气孔相关基因的相关性, 对Pn、Tr和脆枝青藓气孔运动相关基因表达进行主成分分析(图3A)。PC1概括95.2%的变量, 因此PC1能够较为充分地反应指标间关系。PC1负向端概括Pn、Tr以及RCAR10、ABCB14、KUP6等气孔调节相关基因的表达变化; 而PC1正向端概括PIP2;1、ACA10、AHA5、AVP1等胞内水分运输分配相关基因及OST1激酶的表达变化。
通过MapMan注释脆枝青藓转录组, 发现具有节律性的光合相关基因25个(图2B), 其中17个基因的表达量从外界黎明开始上升到外界黄昏前后开始下降, 占光合相关基因的68.0%, 光系统II (PSII)相关基因占绝大多数; 而余下8个光合作用相关基因在4:00-8:00呈现较高的表达强度然后逐渐下降。对Pn与光合作用相关基因进行关联性分析(图3C), 其中只有15.7%的基因表达与脆枝青藓Pn具备相关性。通过相关性分析表明节律响应的光合相关基因主要集中于PSII相关蛋白, 尤其是LHC-II捕光复合体的多个蛋白含量与Pn呈正相关关系。
本文的其它图/表
|

{kind=link}