
|
|
||
|
长期放牧和氮添加对黄土高原典型草原长芒草种群生长的影响
植物生态学报
2024, 48 (3):
317-330.
DOI: 10.17521/cjpe.2023.0086
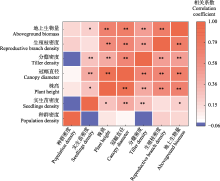
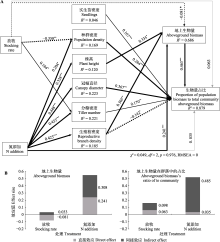
种群是草原生态系统结构、功能形成和发展的基础。然而, 长期放牧伴随着全球气候变化深刻影响着种群的生长和繁殖。长芒草(Stipa bungeana)是黄土高原典型草原的优势种, 具有较高的生态和经济价值。该研究依托黄土高原典型草原长期滩羊轮牧实验平台, 采用完全随机的裂区实验设计, 以放牧率(0、2.7、5.3、8.7 sheep·hm-2)作为主因子, 氮添加水平(0、5、10、20 g·m-2)作为副因子, 探究放牧、氮添加及其交互作用对长芒草的形态性状、地上生物量及其占群落生物量比例的作用。结果表明: 随放牧率的增加, 长芒草的株高、冠幅直径、分蘖密度、实生苗密度、地上生物量和生物量占比呈“单峰”曲线变化趋势, 种群密度降低。而氮添加增加长芒草的株高、冠幅直径、生殖枝密度、分蘖密度、地上生物量和生物量占比; 实生苗密度随氮添加的增加呈先增后减的趋势。相比氮添加, 放牧对长芒草地上生物量和生物量占比的总效应小, 放牧对地上生物量有直接的负效应, 并通过调控分蘖密度、种群密度及地上生物量影响其生物量占比; 氮添加不仅对地上生物量有直接的积极效应, 还通过株高、生殖枝密度对地上生物量有间接的正效应, 同时通过调控种群密度、冠幅直径、分蘖密度和生殖枝密度影响生物量占比。总体上, 氮添加增加长芒草的冠幅直径和生殖枝密度(相比株高和分蘖密度), 放牧增加实生苗密度(相比种群密度), 而放牧和氮添加的交互作用显著影响生殖枝密度。放牧率为4.10和5.29 sheep·hm-2时长芒草具有最大地上生物量及其群落占比。上述结果表明放牧和氮添加通过影响长芒草种群的形态特征来调控其地上生物量和群落地位。 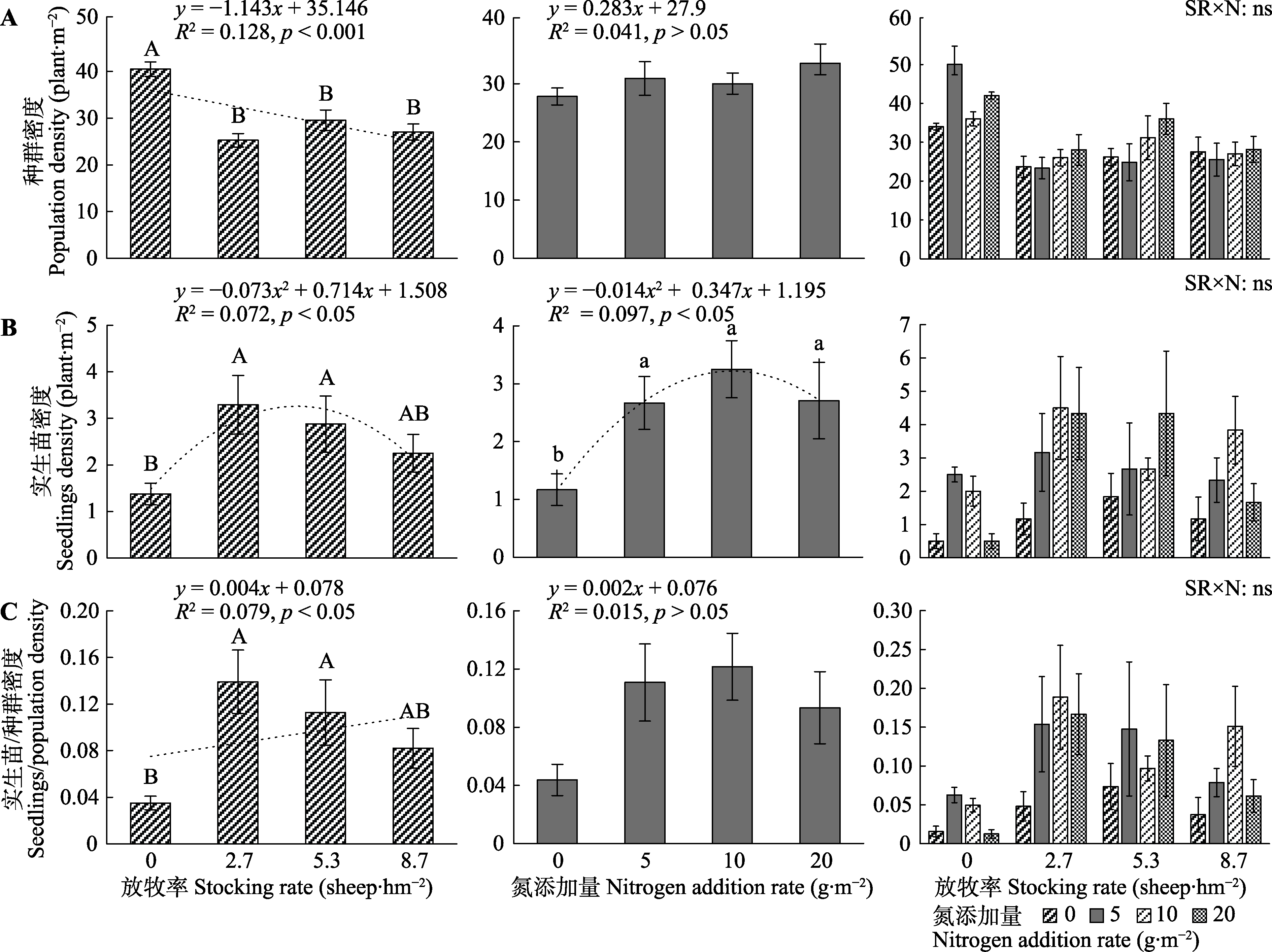 View image in article
图5
放牧和氮添加对长芒草繁殖性状的影响(平均值±标准误)。A, 种群密度。B, 实生苗密度。C, 实生苗密度/种群密度。不同大写字母表示不同放牧率间差异显著(p < 0.05), 不同小写字母表示不同氮添加量之间差异显著(p < 0.05)。N, 氮添加量; SR, 放牧。ns, p > 0.05。
正文中引用本图/表的段落
放牧对长芒草的种群密度有极显著影响, 放牧解释33%的种群密度的变化(图3A, 图5A)。种群密度随放牧率的增大而降低。氮添加、放牧和氮添加的交互作用对种群密度作用不显著(图5A)。
放牧和氮添加对长芒草的实生苗密度均有显著的影响, 两者分别解释8%和10%的实生苗密度的变化(图3A、3B, 图5B)。放牧或氮添加均促进了长芒草实生苗密度的增加, 这种促进作用随放牧率或氮添加量的增加呈“单峰”曲线变化, 放牧率为4.89 sheep·hm-2或氮添加量12.39 g·m-2时, 实生苗密度最高。放牧和氮添加的交互作用对实生苗密度的作用不显著(图5B)。
放牧对长芒草的实生苗密度/种群密度有显著影响, 实生苗密度/种群密度随放牧率的增加呈增加趋势(图5C), 说明放牧主要通过增加实生苗而促进种群更新。氮添加、放牧和氮添加的交互作用对实生苗密度/种群密度的作用不显著(图5C)。
分蘖密度和种群密度反映了植物无性繁殖的能力, 生殖枝密度和实生苗密度表征了植物有性繁殖的潜力和植被更新的活力(俞斌华等, 2005; 刘文亭等, 2022)。长芒草的繁殖性状对放牧和氮添加表现出不同的响应策略, 放牧增加了长芒草的分蘖密度(6%)和实生苗密度(8%), 降低了种群密度(33%) (图3-5)。这主要是受放牧行为的影响, 因为家畜的践踏和采食促进长芒草分株, 提高无性繁殖的能力(古琛等, 2017); 动物的选择性采食导致株丛较大、适口性较好的禾本科牧草优先被采食(周洁艳等, 2022), 因此长芒草的种群密度随放牧率的增加显著降低(图5A); 冬季放牧去除凋落物, 破坏地表结皮, 为来年种子的萌发和实生苗的生长提供了良好的环境(杨畅等, 2022)。放牧超过环境的承载能力后, 草地的抵抗力和恢复力下降(张爽等, 2017), 种间和种内资源竞争变大, 长芒草对生长资源的分配会从耐牧调整为避牧, 株丛活力减弱, 分蘖数量相对减少(Carneiro et al., 2021); 同时家畜在草地的过度活动导致土壤紧实度增加、含水量降低、化学计量比失衡(侯扶江等, 2004), 不利于种子的萌发, 导致放牧对分蘖密度(5.87 sheep·hm-2)和实生苗密度(4.89 sheep·hm-2)的作用有放牧率阈值的限制(图4, 图5)。长芒草的繁殖性状对放牧响应的结果增加了实生苗密度/种群密度(图5C), 说明放牧干扰下长芒草更倾向于通过增加实生苗密度来维持种群的更新。
本文的其它图/表
|


{kind=link}