
|
|
||
|
广西红锥人工林径向生长的季节格局及其对气候因子的响应
植物生态学报
2024, 48 (8):
1021-1034.
DOI: 10.17521/cjpe.2023.0192
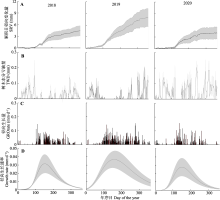
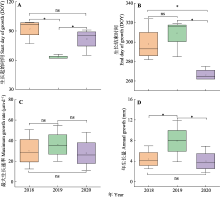
树干径向生长量(GROrate)和水分亏缺量(TWD)是树木响应环境因子的重要表征, 分别受不同环境因子的影响, 对环境因子的响应也不同。研究径向变化动态及其对环境因子的响应关系, 对了解树木生长和生理特性应对气候变化具有重要意义。该研究利用高分辨率径向变化记录仪, 连续记录2018-2020年红锥(Castanopsis hystrix)树干径向变化过程, 同步监测环境因子, 分析GROrate和TWD的动态变化及其与环境因子的关系。结果表明: 径向生长的开始时间为3月4日至4月1日, 结束时间为9月23日至11月5日, 最大生长速率的出现时间为5月31日至6月8日。生长季内红锥生长呈不连续性, 实际生长天数占整个生长季长度的47.8%-74.1%。生长季越长, 生长发生的天数越多。日尺度上, 在主要生长期(4-9月)环境因子中空气相对湿度(RH)、降水量(P)、光合有效辐射(PAR)和饱和水汽压差(VPD)与GROrate的相关性最强, 而TWD的环境相关性与GROrate类似, 但方向相反。21天滑动相关结果显示, 在3年生长季的绝大部分时间VPD、P和RH是影响红锥径向变化的关键因素。月尺度上, GROrate与月降雨事件高度同步, 而TWD与干旱期同步。因此红锥径向变化主要响应水分相关的环境因子, 这将有助于更好地预测气候变化下森林动态的生长响应。 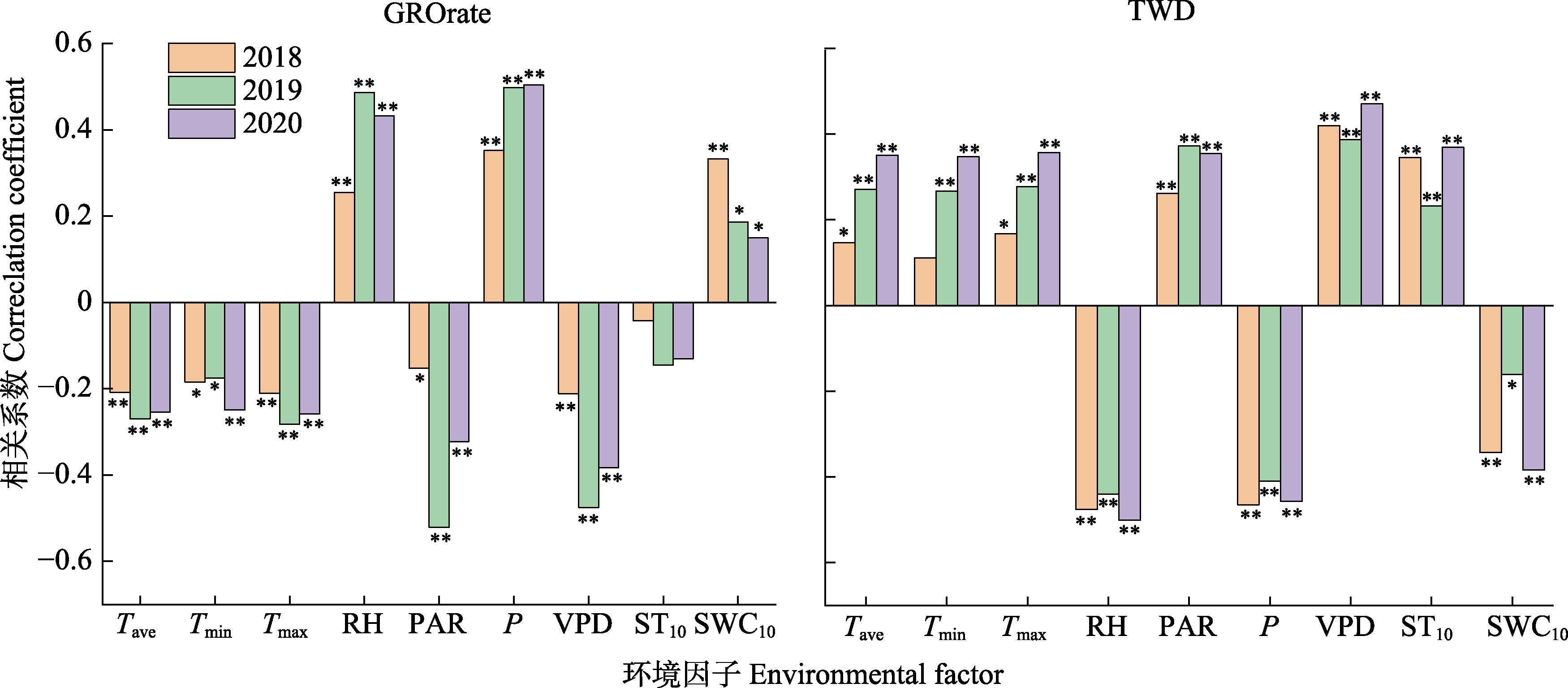 View image in article
图6
广西红锥林标准化日径向生长量(GROrate)和树木水分亏缺量(TWD)与环境因子的关系。*, p < 0.05; **, p < 0.01。P, 降水量; PAR, 光合有效辐射; RH, 相对湿度; SWC, 10 cm深度土壤含水率; Tave, 平均气温; Tmax, 最高气温; Tmin, 最低气温; TS10, 10 cm深度土壤温度; VPD, 饱和水汽压差。
正文中引用本图/表的段落
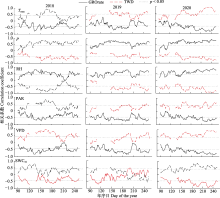
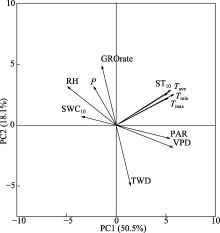
在日尺度上, 相关分析表明, 在所有观测年份的主要生长期(4-9月), GROrate与气温(负相关)、降水量(正相关)、空气相对湿度(正相关)、饱和水汽压差(负相关)、光合有效辐射(负相关)和10 cm 深度土壤含水率(正相关)显著相关(图6)。这些环境因子都是直接(降水量、空气相对湿度、10 cm深度土壤含水率)或间接(气温、饱和水汽压差、光合有效辐射)地影响树木的水分供应, 从而影响树木的径向生长, 表明在本研究区域红锥径向生长受到了水分的限制。TWD和环境因子之间的相关性与GROrate类似, 但方向相反(图6)。在月尺度上, 径向生长量与月降雨事件高度同步, 而水分亏缺量与干旱期同步(图7)。
本研究中, 红锥径向生长停止时间发生在9-11月, 年际差异超过1个月(DOY 266-309) (图5B; 表1), 表明空气或土壤温度可能不是影响生长停止的主要因素。Etzold等(2022)认为生长概率与水分条件密切相关, 尤其是生长后期, 生长停止受水分亏缺的限制大于温度。本研究中, 2018年7月和2020年6-7月降水量少, 气温高(图3), 形成明显的夏季干旱, 出现大量和持续的树木水分亏缺(图4B, 图6), 导致2018和2020年生长结束日期比2019年分别早了11和43天, 这与Oberhuber等(2014)认为夏季水分亏缺可以导致形成层活动和木质部形成提前停止的结果一致。Eilmann等(2011)对欧洲赤松(Pinus sylvestris)进行的灌溉/对照实验结果也证实了水的可用性对径向生长停止的关键作用。研究表明, 形成层的分裂和次生木质部细胞的分化需要由光合同化物转化的蔗糖的持续供应(Deslauriers et al., 2016), 干旱可能会导致气孔关闭和随之而来的蔗糖减少, 使径向生长提前停止。Guo等(2022)开展的模拟降雨减少处理下红锥生长的干旱实验研究表明, 减雨处理下叶和分枝非结构性碳水化合物(NSC)含量显著增加, 而叶和分枝的NSC库主要支持地上过程, 包括出芽、生叶、开花、结果、分枝生长和呼吸维持(Palacio et al., 2018; Huang et al., 2021)。由此可见, 在干旱胁迫下, 红锥倾向于分配有限的资源用于初级生长和繁殖, 而不是径向生长。
本研究中, 红锥径向生长主要发生在4-9月, 这表明亚热带森林树木生长的季节性与降雨的季节性十分吻合(Gheyret et al., 2021)。无论是降雨多的年份还是降雨较少的年份, 在主要生长期, 径向变化与空气温度、降水量、空气相对湿度、饱和水汽压差、光合有效辐射和10 cm深土壤含水率显著相关(图6), 说明与树木水分状况相关的气候因子对红锥径向变化的影响较大。
TWD可以作为树木水分状况的指示因子, 反映水分损失对树干径向变化的影响。在主要生长期, TWD与最高气温、光合有效辐射和饱和水汽压差正相关, 与降水量、空气相对湿度和土壤含水率负相关(图6)。最高气温、光合有效辐射和饱和水汽压差可降低大气水势, 促进蒸腾作用, 对细胞膨压产生负面影响, 抑制细胞扩大和生长。相反, 降水量、空气相对湿度和土壤含水率可以通过增加树干中的水分状态直接有利于细胞增大。Garnier和Berger (1986)研究表明, 干旱导致径向收缩增加。本研究结果显示, 在所有观测年份, TWD与季节干旱期同步, 尤其是2020年的夏季极端干旱导致径向生长收缩时间较长, 收缩强度大(图4B, 图7; 表3)。全球变暖导致的气温升高将加强干旱频率和强度, 从而增加干旱对径向生长的影响。因此, 在全球变化的背景下, 干旱胁迫对树木生长的影响应给予更多的关注。
本文的其它图/表
|

{kind=link}