


任明迅(2008)对雄蕊的合生现象(Stamen fusion)进行了研究, 区分出3种主要的雄蕊合生方式: 花丝合生、花药合生、花丝花药均合生。其中, 花内所有花药合生形成的聚药雄蕊(Synantherous stamen)以及花丝花药均合生形成的花药筒(柱)可把花药以及花粉粒都集中在一起, 可能影响到花药与柱头之间的空间距离(雌雄异位, Herkogamy)、花粉呈现规律及其与传粉者的相互作用。任明迅(2008)根据花粉呈现理论(Pollen presentation theory, Harder & Wilson, 1994)认为, 花药在生长上的联合, 增加了花内所有花粉接触小型传粉者的可能性, 并使得花粉散播在访花昆虫的同一部位, 可能降低花粉损耗和提高异交传粉的准确性, 因此可促进植物的雄性功能和繁殖效率。但是这方面的实验研究几乎没有, 人们对于花内雄蕊之间为何存在不同形式、不同程度的雄蕊合生现象还缺乏认识。
为揭示雄蕊合生对植物花粉输出以及自交与异交水平等繁殖过程的影响、雄蕊合生与哪些花部特征可能形成稳定的组合(花部综合征, floral syndrome)而适应传粉过程, 本文选取花丝花药均合生的典型植物——半边莲(Lobelia chinensis), 研究其花部综合征和繁育系统, 以验证以上假说、认识其传粉机制和开展更深入的人为操纵实验揭示其适应意义。
1 研究材料与地点
半边莲为桔梗科半边莲亚科的多年生湿生草本植物(有资料将半边莲亚科独立为一个科, 本文根据《中国植物志》及APG (2003)仍将之作为桔梗科的一个亚科), 主要分布在我国中南部地区田垄、草地和小路边, 较为常见。半边莲茎柔弱, 常匍匐丛生, 茎末端直立, 叶相对互生。花果期在5~9月, 盛花期在7~8月。
研究地点位于中国科学院武汉植物园内的一个半自然的水塘, 岸边为草本植物较多的斜坡。在水陆交界处自然分布有大量的半边莲。与半边莲共存的主要物种为圆叶节节菜(Rotala rotundifolia)、田字萍(Marsilea quadrifolia)、水花生(Alternanthera philoxeroides)和一些低矮的禾草类和莎草科植物。
2 研究方法
2.1 花部综合征与传粉生态学观察
2008年5月, 半边莲花开始大量盛开。选择有代表性的10朵花进行以下观察: 单花开花进程、单花结构、花直径、颜色、分泌物质类型等。
选取和标记5个未开的花蕾进行更准确的长期观测, 每天8:00~9:00、12:00~13:00和 17:00 ~ 18:00各观察1次, 直到花朵完全萎蔫。每次观察均记录花瓣颜色、花药筒与花粉颜色、柱头的伸长与膨大等。半边莲单花寿命可达3~5 d, 因此设置5个花期阶段: 花蕾期(花瓣下部敞开露出雌蕊与雄蕊)、开放期(花开第1天)、盛开期1(花开第2天)、盛开期2(柱头有明显探出和膨大, 通常为花开第3天)和花谢期(花后第4天或第5天)。每个时期取5朵花进行显微解剖, 在显微镜下分析花内各部位尤其是雌蕊与雄蕊的结构及其随发育时间的变化。
参照龚燕兵和黄双全(2007)的方法, 在晴朗天气的清晨、中午和傍晚, 分别进行访花昆虫的观察。对访花昆虫的类型与访花行为、访花频率、花间运动式样等进行观察与比较分析。初步判断半边莲的常见访花昆虫和有效的传粉者。并对常见访花者进行体表镜检, 判断其是否带有半边莲的花粉。
2.2 繁育系统
2.2.1 异交指数(Outcrossing index)
2.2.2 花粉胚珠比(Pollen-ovule ratio, P/O)
2.2.3 花粉活力和柱头活性
同上分5个花期梯度(花蕾期、开放期、盛开期1、盛开期2和花谢期), 取不同时间梯度时的花药放在盛有15% (g∙ml-1)蔗糖溶液的离心管中让花粉萌发。24 h后放入冰箱4 ℃停止萌发, 并加入2滴5%的亚甲基蓝溶液染色30 min。用光学显微镜统计萌发花粉的比率, 根据花粉萌发率来判断花粉活力和花粉寿命。以有活性花粉的比率来判断不同花期阶段的雄性功能水平。
柱头活性可用双氧水检测(Dafni, 1992): 将不同时期的柱头泡入3%的双氧水。大量产生气泡的柱头为可受柱头(有活性), 无气泡或气泡很少的柱头没有活性。由于半边莲特殊的雄蕊合生结构, 使得自花花粉会落置在柱头上; 因此测量柱头活性的时候, 先通过显微镜下细心地剔除黏附的所有花粉粒, 然后再进行上述的柱头活性检测。
2.2.4 人工授粉套袋实验
为揭示半边莲是否存在无融合生殖和自发自交现象, 以及自交亲和性程度, 设置了以下几个套袋处理实验: 1)直接套袋: 对花部不做任何处理, 直接在花未开放时套袋; 2)去雄套袋: 在花开初期将花药筒及合生花丝去除, 然后套袋; 3)人工自交授粉: 花蕾套袋, 待柱头膨大可受时, 轻剥花药筒将其中的花药涂抹于柱头上, 然后套袋; 4)人工异交授粉: 花蕾套袋, 在花开初期去雄。然后在柱头成熟时涂抹适量的异交花粉, 之后套袋。每个处理5个重复。在花萎蔫和将这些处理后得到的结实率与种子数等与自然状态下对应的指标进行比较分析。
2.3 数据分析
采用SPSS 13.0 for Windows 统计软件中One-way ANOVA分析套袋实验中不同处理间的结实率、每果平均种子数, 以及主要传粉昆虫在不同时间段访花频率的差异。数据间的两两比较采用Student-Newman-Keuls检验法(S-N-K法)。
3 结果
3.1 花部综合征与传粉生态学过程
半边莲花大, 具有长的花轴, 单生于直立茎上的叶腋处。花瓣颜色为粉红或浅红, 单花寿命可达3~5 d。花瓣5, 左右两侧的花瓣为细长的披针形, 中间3花瓣椭圆状。花瓣中下部90°弯折并联合成筒状, 花瓣上部平展成一个平面, 偏向一侧形成半圆形的花瓣面(图1A)。花丝与花柱基部有花蜜分泌。花冠筒喉部有毛, 有草绿色斑。5个雄蕊的花药完全合生在一起, 形成长约2 mm的灰蓝色的花药管, 花药管顶端有小口, 小口周围有髯毛。雄蕊的花丝中上部也合生在一起; 花丝下部分离插生于花筒内壁, 有细绒毛。柱头和花柱上端被包围在雄蕊的合生结构内, 形成类似合蕊柱的柱状结构。在开花的第2天到第3天, 柱头从花药筒顶端小口伸出, 并逐渐裂开、膨大(图1B、1C)。
图1

图1
半边莲的开花进程与主要访花昆虫
A: 花开放后的第1天, 柱头还包裹在花药筒内 The 1st day after flower opening, with the stigma and styly enclosed by the cylinder, which is formed by the fused anthers and filaments B: 开花2 d后, 柱头已经伸出了花药筒, 但还未膨大(箭头所示为柱头)Two days after flower opening, stigma extends out of the cylinder (stigma is indicated by arrow head) C: 开花3~4 d后, 柱头变膨大(箭头所示) Three to four days after flower opening, the exserted stigma (indicated by arrow head) opens and becomes receptive. D、E、F: 分别是蚂蚁、食蚜蝇和苍蝇访问半边莲的花 The most frequent visitors to L. chinensis flower are ant (D), hoverfly (E), and fly (F), respectively
Fig. 1
The flowering phenology and main pollinators of Lobelia chinensis
传粉生态学观察发现, 经常有蚂蚁(Campon- otus sp.)从花冠筒裂缝处的花柱基部与花丝的空隙处轻易取食花蜜(图1D), 也有食蚜蝇(Episyrphus sp.)采集花粉或探入花冠筒吸食花蜜(图1E)。蚂蚁访问频率最高, 在中午和傍晚的时候常见于植株与花上, 食蚜蝇的访问频率较低。这两类是半边莲的常见访花者, 其中蚂蚁是观察到的最频繁、访问时间最长的访花者。两类主要访花者都能在访花过程中接触到花内的性器官(图1D、1E)。在体式显微镜下对访问半边莲花后的蚂蚁和食蚜蝇体表观察发现, 约有10%的个体带有半边莲花粉(蚂蚁: n = 15; 食蚜蝇: n = 5)。因此, 蚂蚁和食蚜蝇可能是半边莲的传粉者。实验期间也发现了少量小型蝇类(Muscidae)采食花粉(图1F), 期间没有发现蜂类、蝶类和蛾类等大型访花者。半边莲花在中午(12: 00 ~ 14: 00)的时候, 具有最多的访问者, 而清晨的访花者最少。
3.2 繁育系统
3.2.1 异交指数
半边莲花直径大, 最小处的直径也约为1 cm; 雄蕊先熟。由于特殊的雄蕊合生结构将柱头包围在其中, 柱头和花药空间距离接近, 没有明显的雌雄异位。根据Dafni (1992)异交指数的计算规则和标准, 半边莲的异交指数为4, 繁育系统以异交为主, 部分自交亲和, 需要传粉者。
3.2.2 花粉胚珠比(P/O比)
半边莲单花的花粉数量有(5 200±130)粒, 胚珠数量有(55±6)个; 花粉胚珠比(P/O比)为94.54。依据Cruden (1977)的标准, 半边莲的繁育系统为兼性自交。
3.2.3 花粉与柱头的活性
图2
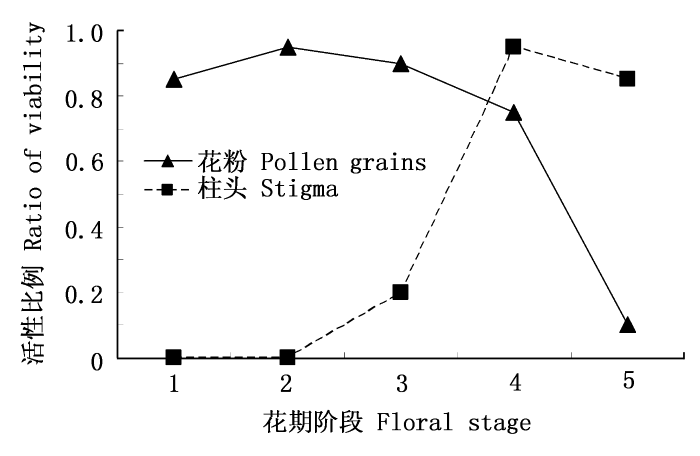
图2
半边莲花粉与柱头的活性变化
Fig. 2
The viability of pollen grains and stigma of Lobelia chinensis
3.2.4 套袋实验
套袋实验结果发现, 去雄套袋没有种子生成, 说明半边莲不存在无融合生殖; 自然情况下套袋(不去雄+套袋)也没有种子生成, 表明不存在自发自交。人工自交授粉得到了与人工异交授粉相似的结实率, 表明该植物自交亲合性很高。人工授粉比自然状态下的结实率高(表1), 说明半边莲野外群体存在一定的花粉限制。
表1 套袋实验的结实情况
Table 1
| 处理 Treatment | 实验花数/个体数 Flowers / individuals | 结实率 Seed-set ratio | 平均每花种子数 Seed number per flower |
| 自然对照 Natural (unbagged) | 33/15 | 33% b | 16 ± 7 b |
| 自然对照 + 套袋 Natural (bagged) | 20/10 | 0 a | 0 a |
| 去雄 + 套袋 Emasculated + bagged | 15/15 | 0 a | 0 a |
| 人工自交授粉 + 套袋 Hand-pollinated with self pollen + bagged | 30/15 | 90% c | 45 ± 10 c |
| 人工异交授粉 + 套袋 Hand-pollinated with outcrossing pollen + bagged | 35/15 | 85% c | 50 ± 5 c |
| 表中有不同字母的数值表示在0.01水平有显著差异 Values with different letters are significantly different at the 0.01 level | |||
4 讨论
4. 1 繁育系统与P/O比
半边莲雄蕊的花药紧密合生成花药管, 花粉向管内散粉, 管内底部由柱头及其收集毛承载花粉粒, 形成花粉的二次呈现(Secondary pollen presentation)(Howell et al., 1993; Castro et al., 2008)。这与菊科植物聚药雄蕊的结构与散粉模式很类似。这种花药合生成筒且花粉释放在筒内的聚药雄蕊结构, 与兰科和萝藦科植物雄蕊花粉形成花粉块一样, 能使花粉集中呈现在同一个空间位置, 多个花粉粒能作为整体一起释放或被传粉者接触, 有利于降低花粉损耗、促进花粉准确有效地输出, 使得植物不必通过生产大量花粉来保证异交花粉的成功散布。因此, 这可能是导致半边莲P/O比偏低的一个原因。在二强雄蕊、花药成对合生的厚萼凌霄(Campsis radicans)中也发现, 这个以异交为主的雄蕊合生植物P/O比也偏低, 与半边莲的P/O值近似(任明迅, 未发表数据)。
半边莲中的低花粉胚珠比的另外一个原因可能与花粉聚联(Pollen aggregation)相似。花粉聚联是指花药内的花粉不是以单个花粉粒的形式存在, 而是通过不同方式结合成2个一体, 4个一体或者多个一体的黏合; 如兰科和萝藦科中的花粉块(Pollinia)、柳叶菜科柳叶菜属和杜鹃花科杜鹃花属等植物中的四合花粉(Tetrads)和四合花粉之间的粘丝(Viscin threads), 以及川苔草科和芝菜科等的二合花粉(Dyads)(Harder & Johnson, 2008)。Cruden和Jensen (1979)以及Cruden (2000)发现, 花粉聚联植物的P/O比通常明显低于不存在花粉聚联的植物。Harder和Johnson (2008)认为, 这可能是因为花粉粒聚联在一起进行传粉和授粉, 不仅能提高传粉效率, 而且通过集体传粉和授粉过程提高了每个花粉粒成功授粉的可能性, 进而增加了子房内所有胚珠都被同一来源花粉受精的可能性。因此, 花粉聚联提高了植物的局域配偶竞争的强度(Harder & Johnson, 2008)。局域配偶竞争的选择压力提高了个体对雌性功能的投入(胚珠数量增加), 减少了雄性功能的投入(花粉数量降低), 因而导致了低P/O比(Hamilton, 1967; Harder & Johnson, 2008)。从传粉机制来看, 雄蕊合生(尤其是花药的合生)与花粉聚联的传粉过程与机制非常类似, 都通过把花粉聚集在一起提高了传粉效率, 降低了移出花药内所有花粉的所需昆虫访问数, 可能也因此提高了单个花粉粒的授粉成功率, 并增加了来自同一个体的花粉授精子房内所有胚珠的可能性。因此, 在半边莲这类雄蕊合生的植物中发现的低P/O比, 可能与花粉聚联植物的低P/O比具有近似的进化来源。
4.2 传粉生态学过程
实验期间, 蚂蚁是半边莲最常见的访花者, 访花时间较久, 也经常从花瓣开口处进入花内, 接触雌雄蕊。由于半边莲平卧地表, 花冠筒未封闭(在花冠筒后部雌蕊与雄蕊的生长之处未闭合, 暴露花蜜), 蚂蚁可方便地到达花冠基部, 吸取花蜜(图1D)。这些特征似乎都有利于蚂蚁的访问。蚂蚁对半边莲的访花方式, 通常是抱着性器官形成的柱状结构, 从花外往下探寻花冠筒裂缝处的花蜜(图1D)。这个过程中, 蚂蚁腹部可接触到柱头和花药筒开口部位。蚂蚁不仅可从花冠筒裂缝采蜜, 也会从花冠开口处贴着雄蕊与雌蕊形成的合生柱状结构深入花内, 并在这个过程中接触花药与柱头。对访问过半边莲的蚂蚁体表进行了镜检, 也发现有部分个体其后腿与胸腹部确实粘有半边莲的花粉, 说明频繁地长时间访问半边莲的蚂蚁可能是半边莲的主要传粉者之一。由于半边莲花部特征符合蚂蚁传粉综合征, 且花部结构较为特化、与蚂蚁的访问方式配合较好, 这种蚂蚁传粉机制可能是长期进化产生的普遍适应, 而非实验地半边莲群体局域形成的暂时性传粉机制。但这还尚待更严谨的实验证明。
实验期间, 也有较多的食蚜蝇和苍蝇访问半边莲, 它们经常停落在花药筒上, 或从花瓣开口处进入花内接触到性器官。对访问半边莲花之后的食蚜蝇体表镜检发现, 约有2/5的个体带有半边莲的少量花粉。因此, 食蚜蝇也可能是半边莲的有效传粉者。蝇类访花较少, 只捕捉到了2个访花个体, 且在其体表未能检测到半边莲花粉, 因此蝇类可能不是半边莲的主要传粉者。
4.3 雄蕊合生的花部综合征
花丝花药均合生(尤其是花药紧密合生成一体)的现象, 不仅在桔梗科半边莲亚科中广泛存在, 也分布在菊科、凤仙花科、堇菜科、香茜科、葫芦科、茄科以及苦苣苔科苦苣苔属(Conandron spp.)等很多植物中, 在系统进化上表现为多次起源(任明迅, 2008)。花药合生以及花丝花药均合生的植物大多有以下共同特征: 1)花冠形成花冠筒或起码花冠基部合生成管状; 2)雄性先熟; 如果花药合生成花药筒, 花粉会落置在筒内未成熟的柱头上形成花粉的二次呈现; 3)传粉者多为小型昆虫或用喙伸入花内吸蜜的蝶蛾类(任明迅, 2008)。这些特征能较为普遍地关联出现, 可能是由于小型传粉者以及用喙进行传粉的昆虫与花药或花粉接触面有限, 围拢的花冠筒以及紧密合生在一起的花药与花粉能提高花内所有花粉接触到传粉者的可能性, 促进花粉的输出; 同时, 合生在一起的花药也提高了传粉的准确性, 降低了花粉的损耗, 减少了移出花药内所有花粉的所需昆虫访问数(任明迅, 2008; Harder & Johnson, 2008)。
本研究结果发现, 半边莲确实具有以上3个特征, 从而初步证实了花药合生(包括花丝花药均合生)很可能与这些特征形成较稳定的组合, 即“花部综合征”(传粉综合征)。这一花部综合征可能通过管状的花冠筒降低了小型传粉者活动的任意性, 通过合生的花药促进了花内所有花粉接触到传粉者和提高传粉效率, 通过雌雄异熟(主要是雄性先熟)避免了自交可能性(合生结构往往导致了雌蕊与雄蕊的靠近)。当然, 这种花部综合征(花药合生+花被筒+雄性先熟+小型传粉者)的普遍性及其具体的传粉过程, 还有待于更多的实验研究加以证实。
李晓琼和张家亮同学协助传粉观察和显微分析, 特此致谢
参考文献
An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG II
Sexual interference of floral kind

Floral hermaphroditism results in conflicts and compromise in the parental roles of plants during pollination and mating. A potential cost of hermaphroditism is sexual interference between maternal and paternal functions resulting in gamete wastage and reduced fitness. Sexual interference may or may not be associated with self-pollination. In cases where self-pollination occurs, ovule or pollen discounting may reduce mating opportunities. Here I describe forms of sexual interference in flowering plants, distinguishing whether physical or biochemical interactions are involved and whether fitness costs associated with gamete wastage arise from intra-floral versus inter-floral processes. I review the limited experimental evidence for interference between sex functions and evaluate the hypothesis that some floral adaptations usually interpreted as anti-selfing mechanisms may serve an alternative function in reducing mating costs arising from this form of sexual conflict.
How does secondary pollen presentation affect the fitness of Polygala vayredae (Polygalaceae)?
Pollen-ovule ratios: a conservative indicator of breeding systems in flowering plants
Pollen grains: why so many?
Viscin threads, pollination efficiency and low pollen-ovule ratios
On methodology of foraging behavior of pollinating insects
Pollen removal from tristylous Pontederia cordata: effects of anther position and pollinator specialization
Function and evolution of aggregated pollen in angiosperms
Evolutionary options for maximizing pollen dispersal of animal-pollinated plants
Floral evolution and male reproductive success: optimal dispersing schedules for pollen dispersal by animal-pollinated plants
Secondary pollen presentation in angiosperms and its biological significance
Anther arrangement influences pollen deposition and removal in hermaphrodite flowers
Chenorchis: a new orchid genus and its eco-strategy of ant pollination
Evolutionary implications of pollen presentation schedules in animal-pollinated plants
Pseudocopulation of an orchid by male ants: a test of two hypotheses accounting for the rarity of ant pollination
Outcrossing in ant pollinated clonal orchid
Ant pollination of the perennial herb Blandfordia grandiflora (Liliaceae)
Stamen fusion in plants: diversity, adaptive significance, and taxonomic implications
Ants pollinate Neottia listeroides (Orchidaceae) in Sichuan, China

{kind=link}
{kind=link}
{kind=link}
{kind=link}