


生物多样性与生态系统功能的关系是当前生态学领域内的一个重大科学问题(Díaz & Cabido, 2001; 张全国和张大勇, 2002, 2003; Hillebrand & Matthiessen, 2009; 赵新全, 2009)。已有很多实验研究表明群落初级生产力和稳定性随物种多样性增加, 该多样性效应得以实现的机制是超产效应(overyielding effect)、异步性效应(asynchrony effect)和投资组合效应(portfolio effect) (Isbell et al., 2009)。与此相反的观点认为, 由于环境筛或生态系统管理制度因素引起群落物种功能属性的趋同变化, 最终影响生态系统功能的主要因素可能是物种组成及其功能属性, 初级生产力和稳定性将由群落优势种控制, 而非物种多样性本身(Grime, 1998; Sasaki & Lauenroth, 2011)。因此, 有学者认为生态位互补效应在自然生态系统中是否还具有重要作用尚属未知(Grace et al., 2007), 较高物种多样性对群落的有效稳定作用可能不存在(Sasaki & Lauenroth, 2011)。我们以前的研究表明, 高寒灌丛草甸在经过12年连续放牧后, 不同放牧强度的群落结构差异明显, 由于物种在放牧梯度上的周转变化, 群落的物种组成相差18%-20%, 而物种多样性和均匀度并无显著差异(Zhu et al., 2008)。
群落中有些物种的相对多度对环境或扰动因素的变化会表现出正、负或单峰变化等多种响应模式, 这些物种称为响应物种(response species, RS), 而有些物种对此无响应, 即中立物种(neutral species, NS)。响应物种不一定就是最终影响生态系统功能的作用物种(effect species, ES) (Díaz & Cabido, 2001; Hooper et al., 2005), 两者可能相同, 也可能不完全相同, 但作用物种一定是响应物种。此外, 群落物种常常会在一定的环境梯度范围内发生周转, 但总有一些物种稳定存在于群落中, 且其重要值并无明显变化, 它们是共有物种(common species, CS)。尽管目前对群落物种多样性的生态系统功能效应有不同认识, 甚至对物种多样性本身是否有价值也提出了疑问, 但群落中这些不同“身份(identity)”物种的多样性与特定生态系统功能之间具有何种关系并不明确。比如, 它们是否执行不同的生态系统功能, 或者在同一功能中的作用有所不同?通过区分群落物种的“身份”, 揭示其物种多样性的生态系统功能效应与机制, 将进一步深化生物多样性的生态系统功能效应研究。
高寒草甸生态系统作为青藏高原最重要的自然生态系统类型之一, 长期受放牧家畜的强烈影响, 加之地处高寒, 虽然土壤有机质丰富, 但其矿化率较低, 可利用养分缺乏, 不能满足植物生长需要(赵新全, 2009)。因此, 高寒草甸群落及其主要物种对放牧的补偿响应模式强烈依赖于土壤有效养分资源的获得性, 解除养分限制能提高群落初级生产力和植物耐牧性(席博等, 2010; 朱志红等, 2010; 李晓刚等, 2011), 其初级生产力随物种多样性呈饱和递增模式(顾梦鹤等, 2011)。高寒灌丛草甸中对放牧扰动发生响应的物种数为17.4%, 而中立者高达82.6% (Zhu et al., 2012)。这说明放牧和土壤资源获得性对该群落的初级生产力有重要影响, 就生产力而言, 还可能存在物种冗余。但在放牧强度和资源获得性同时变化时, 生产力和稳定性功能究竟受控于哪类“身份”物种的多样性变化, 超产、异步性和投资组合效应是否共同起作用等问题均不明确。
本研究通过在高寒草甸为期5年(2007-2011年)的人工刈割、施肥和浇水控制实验, 比较了不同处理群落中全部物种、响应物种、作用物种和共有物种等4类物种的物种多样性与群落地上净初级生产力及稳定性的关系, 并就决定稳定性的3个参数(总多度、总协方差和总方差)与4类物种多样性的回归关系(它们分别表征超产、异步性和投资组合效应), 研究了多样性对稳定性的作用机制, 旨在揭示以下3个问题: (1) 4类物种的多样性与生产力及稳定性的关系模式及其相对影响; (2)多样性导致稳定性的作用机制; (3)刈割和土壤资源获得性对上述关系和功能的影响。
1 材料和方法
1.1 研究区概况
本研究在中国科学院海北高寒草甸生态系统定位研究站矮嵩草(Kobresia humilis)草甸内进行。定位站的地理坐标为37°29′-37°45′ N, 101°12′- 101°23′ E。研究区地形开阔, 多滩地和河流阶地, 海拔3200 m, 年平均气温-1.6 ℃, 年降水量560 mm, 6-9月降水量占全年降水量的68.6% (李英年等, 2004)。研究区地处高寒, 气温变幅大, 寒流和冷空气侵入频繁, 强度大, 霜冻严重, 全年无绝对无霜期, 相对无霜期约20天, 在最热月的7月仍可出现霜冻、结冰、降雪(雨夹雪)等冬季天气现象(李英年等, 2005)。矮嵩草草甸土壤为高山草甸土(王长庭等, 2005), 该群落在当地为冬春草场, 每年11月至次年5月底放牧, 放牧家畜为藏羊和牦牛。
1.2 研究方法
1.2.1 实验设计
研究样地建于2007年4月底。实验设刈割、施肥和浇水3种处理, 3区组, 每区组3个4 m × 4 m的大样方, 裂区实验设计。主区为刈割, 每年6月中旬进行, 模拟家畜春季放牧, 留茬高度分别为1 cm (H1)、3 cm (H3)和不刈割(NH), 刈割生物量分别为60%-70%、45%-50%和0。副区嵌套于主区中, 用4块长2 m、宽0.25 m的雪花铁皮十字形纵切嵌入草地0.25 m深, 将每个主区大样方分隔成4个2 m × 2 m的副区样方, 在其中分别设置4个处理, 即: (1)不施肥、不浇水(NFNW); (2)仅施肥(F); (3)仅浇水(W); (4)既施肥又浇水(FW)。在每个副区样方中央1.5 m ×1.5 m范围内进行刈割、施肥和浇水处理。每个副区样方内再设4个0.5 m × 0.5 m的观测样方, 其中1个为长期观察物种组成变化的永久样方, 剩余3个用于植物性状测定。每年施肥量以当地建植人工草地时的中等施用尿素量225 kg·km-2为依据, 分3次撒施, 分别于5-7月中旬进行。每次施用尿素2.5 g·m-2 (含N 20.4%), 磷酸二铵0.6 g·m-2 (含N 5.9%, P 28.0%)。施肥后用喷壶浇水4.5 kg·m-2, 浇水总量为年降水量的2.4%。对2007和2008年的数据分析显示, 浇水的作用不显著。故从2009年开始, 每次浇水量增至6.7 kg·m-2, 浇水总量为年降水量的3.6%。
1.2.2 取样步骤与方法
从2007年到2011年, 每年8月上旬群落初级生产力达到高峰时测定永久样方内的物种数、盖度(目测)、密度(双子叶植物为个体数, 丛生单子叶植物为分株数)、高度和总地上生物量(齐地面刈割, 60 ℃烘干72 h, kg·m-2), 共108个样方。将此生物量与6月份相同处理样方的刈割生物量(不刈割处理为0 kg·m-2)相加, 作为该样方地上净初级生产力(ANPP, kg·m-2·a-1)的估计。用于本文数据分析的数据采自2011年。
1.3 数据计算与统计分析
1.3.1 物种重要值
样方中物种重要值(IV)以下式(任继周, 1998)计算:
$IV=\left(相对密度+相对盖度+相对高度 \right)\text{/3}$
1.3.2 物种多样性
物种多样性用Simpson多样性指数D表征(马克平和刘玉明, 1994), 计算公式如下:
$D=1-\sum\limits_{i=1}^{s}{{{P}_{i}}^{2}}$
式中, Pi为样方中第i个物种的相对密度, s为物种数。本研究分别计算了物种丰富度指数和Shannon-Wiener多样性指数H', 但其在实验处理中的变化与D相同, 故文中结果仅以Simpson多样性指数D显示。
1.3.3 生态系统功能
以群落地上净初级生产力及密度稳定性表征生态系统功能。大部分进行群落时间稳定性的研究都采用生物量计算时间稳定性特征, 也有研究用密度(Sasaki & Lauenroth, 2011)或盖度(Yang et al., 2011)计算。我们对密度与地上生物量进行回归分析, 两者间具有显著的相关性(p < 0.05), 因此, 本文以密度数据计算稳定性。同时, 我们借用时间稳定性指数(temporal stability index, TSI) (Lehman & Tilman, 2000; Sasaki & Lauenroth, 2011), 目的在于比较不同处理群落的稳定性, 并通过其包含的3个重要参数, 分析稳定性机制, 公式如下:
$TSI=\frac{\mu }{\sigma }=\frac{\sum{Abu}}{\sqrt{\sum{Var+\sum{Cov}}}}$
式中, μ为平均密度, σ为密度的标准差, μ/σ 为变异系数的倒数, 用ICV表示(Tilman, 1999; Lehman & Tilman, 2000)。ΣAbu、ΣVar和ΣCov分别是各物种的总密度、密度总方差及所有成对物种密度变化的总协方差。这3个参数与物种多样性的关系可分别表征物种多样性作用的3种机制, 即: 超产效应、投资组合效应和异步性效应。ICV或TSI值越大, 稳定性越高。如果ICV或TSI随物种多样性增加, 那么物种多样性高的群落, 其ΣVar和ΣCov应越低(即存在由物种多样性增加引起的投资组合与异步性效应), 而ΣAbu应越高(即超产效应) (Tilman, 1999; Lehman & Tilman, 2000; Cottingham et al., 2001; Valone & Hoffman, 2003; Isbell et al., 2009; Sasaki & Lauenroth, 2011)。本文以各处理每区组中的3个样方计算1个TSI值, 因此, n = 9。
在自然群落中进行物种多样性效应研究时, 不同处理群落的物种数不相等, 不宜直接比较群落物种的异步性波动(Loreau & de Mazancourt, 2008)。Loreau和de Mazancourt (2008)和Isbell等(2009)采用群落尺度上物种的同步性变化间接表示物种的异步性, 同步性越高, 则异步性越低, 计算公式如下:
${{\phi }_{b}}=\frac{\mathop{\sigma }_{bT}^{\text{2}}}({\mathop{\sum\nolimits_{i=\text{1}}^{s}{{{\sigma }_{bi}}}})^{\text{2}}}$
式中, σ2bT是物种密度的总方差, σbi是物种i在具有S个物种的群落中密度的标准差。当φb = 1时, 表示群落物种密度发生同时增加或降低的绝对同步变化, 异步性最低, 群落的稳定性也最低; 当φb = 0时, 表示绝对异步, 群落稳定性最高(Loreau & de Mazancourt, 2008; Isbell et al., 2009)。
Taylor (1961)、Doak等(1998)认为, 在群落总密度给定的条件下, 如果群落物种密度的总方差随物种多样性下降, 则群落稳定性提高。但其条件是总密度的方差σ2与物种平均密度μ符合幂函数σ2 = cμz, c、z均是常数, 且z > 1, 即存在投资组合效应。我们计算了z值, 以验证是否存在该效应。
1.3.4 统计分析
1.3.4.1 实验处理效应 采用SPSS 17.0-GLM中Univariate, 进行裂区设计的方差分析。以区组作为随机因子, 主区为刈割强度, 副区为施肥、浇水和互作项, 对物种重要值、4类不同“身份”物种所占比例、物种多样性、净初级生产力和稳定性的变化进行分析。处理间平均值的多重比较采用Duncan氏检验法。统计分析前对数据进行正态性和方差齐性检验, 并分别对百分比数据和生物量数据进行反正弦和对数转换, 对其他数据进行平方根转换。统计检验的显著性水平为p < 0.05。
1.3.4.2 物种响应类型的确定 若方差分析中某物种的重要值在任一处理或互作影响下的F检验显著(p < 0.05), 则说明该物种对实验处理具有明显响应, 确定为响应物种。然后运用SPSS 17.0-GLM中Regression程序对响应物种重要值与群落净初级生产力间的关系进行线性分析、二次函数和逻辑斯蒂模型拟合(n = 36), 并以F检验显著(p < 0.05)且决定系数R2最大的回归模型为最优模型, 说明该响应物种对群落净初级生产力有显著影响, 依此确定作用物种及其响应模式。共有物种是指存在于所有样方中的物种。我们以前的研究(Zhu et al., 2008, 2012)显示高寒草甸物种在放牧强度梯度上多表现为线性或二次函数响应模式。这里考虑逻辑斯蒂模型是因为本实验有施肥×不刈割处理, 可能引起某些物种种群密度和地上生物量有较大增加, 此时该物种的种内相互作用应超过种间影响, 其种群重要值-生产力关系在不施肥×刈割到施肥×不刈割处理梯度上可能遵循逻辑斯蒂模型变化。
1.3.4.3 物种多样性与生态系统功能的关系 采用与上述相同的方法分别拟合4类不同“身份”物种的Simpson多样性指数与群落净初级生产力及稳定性间的关系方程。我们将12个处理组合分为10种处理组合梯度进行拟合。但由于浇水处理对4类物种的物种多样性(FTS = 0.638, FRS = 0.943, FES = 0.529, FCS = 0.624, p > 0.05)、初级生产力(F = 0.037, p > 0.05)和稳定性ICV (F = 0.001, p > 0.05)均无显著影响, 故对上述关系进行拟合分析时, 将浇水处理样方并入施肥处理样方, 相当于同一副区各有2个施肥与不施肥样方。文中仅显示了刈割与施肥处理下的物种多样性、初级生产力和稳定性变化, 及其组合梯度下生产力及稳定性之间的5种回归结果, 即: 不施肥-刈割强度梯度(n = 18)、施肥-刈割强度梯度(n = 18)、不刈割-施肥梯度(n = 12)、中度刈割-施肥梯度(n = 12)以及重度刈割-施肥梯度(n = 12)。
为了解释物种多样性与稳定性关系的机制, 对4类物种的多样性指数与稳定性的3个统计参数(ΣAbu、ΣVar和ΣCov)分别进行回归分析。
2 结果和分析
2.1 物种响应类型
群落中调查记录到的全部物种数为71个。其中, 响应物种、作用物种和共有物种数分别为26、13和46个, 分别占全部物种数的36.6%、18.3%和64.8%, 作用物种占响应物种数的50.0%。共有物种中, 响应物种和作用物种各占50.0%和23.9%。对刈割、施肥、浇水、刈割×施肥、刈割×浇水、施肥×浇水以及刈割×施肥×浇水处理发生响应的物种数分别占全部物种数的15.5%、28.4%、2.8%、7.0%、4.2%、0.0%和1.4% (表1)。
表1 响应物种重要值的方差分析及其与群落地上净初级生产力的回归分析
Table 1
| 植物种 Plant species | 变异来源(自由度) Sources of variance (df (m, n)) | R2 (n = 36) | 物种类型 Species type | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| B (2, 4) | C (2, 4) | F (1, 18) | W (1, 18) | C × F (2, 18) | C × W (2, 18) | F × W (1, 18) | C × F × W (2, 18) | |||
| 矮嵩草 Kobresia humilis | 4.43 | 0.32 | 5.29* | 0.96 | 1.25 | 0.23 | 0.52 | 0.24 | 0.006 | RS |
| 鹅绒委陵菜 Potentilla anserina | 12.71* | 6.96* | 14.87** | 0 | 5.52* | 4.86* | 0.01 | 0.89 | 0.014 | RS |
| 甘肃马先蒿 Pedicularis kansuensis | 0.27 | 2.56 | 5.51* | 4.55* | 0.54 | 1.01 | 3.91 | 1.19 | 0.013 | RS |
| 海乳草 Glaux maritima | 8.53* | 7.01* | 0.58 | 0.30 | 6.91** | 0.12 | 0.01 | 0.43 | 0 | RS |
| 尖叶龙胆 Gentiana lawrencei | 1.48 | 5.02 | 41.23** | 4.02 | 2.58 | 0.06 | 0.13 | 1.96 | 0.014 | RS |
| 美丽风毛菊 Saussurea superba | 2.61 | 7.16* | 1.33 | 0.02 | 0.80 | 0.91 | 1.28 | 0.05 | 0.024 | RS |
| 棉毛茛 Ranunculus membranaceus | 1.72 | 10.86* | 7.13* | 0.79 | 2.19 | 0.48 | 0.35 | 1.87 | 0.022 | RS |
| 蒲公英 Taraxacum mongolicum | 1.46 | 16.47* | 0.03 | 0.35 | 0.54 | 1.61 | 0.16 | 0.95 | 0.033 | RS |
| 溚草 Koeleria cristata | 1.00 | 1.46 | 15.83** | 0.83 | 1.77 | 0.12 | 0.46 | 0.53 | 0.009 | RS |
| 麻花艽 Gentiana macrophylla | 2.62 | 4.21 | 4.64* | 0.39 | 0.29 | 4.69* | 4.21 | 0.36 | 0.033 | RS |
| 小米草 Euphrasia regelii | 1.41 | 2.61 | 10.92** | 0.61 | 0.95 | 0.71 | 0.21 | 0.34 | 0.002 | RS |
| 亚洲蒲公英 Taraxacum leucanthum | 0.19 | 5.08** | 1.97 | 1.76 | 6.23** | 3.02 | 0.26 | 0.96 | 0.028 | RS |
| 紫菀 Aster alpinus | 1.81 | 8.21* | 0.68 | 0.86 | 3.43 | 0.40 | 0.28 | 5.06* | 0.016 | RS |
| 矮火绒草 Leontopodium nanum | 1.20 | 3.76 | 6.20* | 0.13 | 1.45 | 0.13 | 0.03 | 0.05 | 0.072** | RS, ES |
| 繁缕 Stellaria umbellata | 0.55 | 0.56 | 79.92** | 3.43 | 0.25 | 0.13 | 1.58 | 0.04 | 0.153** | RS, ES |
| 甘肃棘豆 Oxytropis kansuensis | 1.34 | 3.10 | 51.59** | 0.09 | 3.12 | 0.17 | 1.30 | 0.09 | 0.172** | RS, ES |
| 黄花棘豆 Oxytropis ochrocephala | 0.30 | 1.92 | 25.48** | 1.65 | 2.63 | 0.71 | 2.35 | 1.18 | 0.039* | RS, ES |
| 黄芪 Astragalus licentianus | 0.60 | 1.5 | 17.70** | 0.22 | 3.67* | 0.92 | 3.93 | 1.31 | 0.088** | RS, ES |
| 米口袋 Gueldenstaedtia verna | 2.21 | 3.91 | 20.17** | 0.27 | 0.98 | 0.14 | 1.17 | 0.79 | 0.101** | RS, ES |
| 垂穗披碱草 Elymus nutans | 0.63 | 1.66 | 26.18** | 0.26 | 0 | 0.13 | 0.07 | 2.05 | 0.145** | RS, ES |
| 青海棘豆 Oxytropis qinghaiensis | 0.63 | 1.22 | 35.19** | 1.88 | 1.73 | 0.56 | 0.04 | 1.56 | 0.130** | RS, ES |
| 三脉梅花草 Parnassia trinervis | 3.48 | 2.5 | 17.60** | 0 | 1.55 | 0.08 | 0 | 0.46 | 0.155** | RS, ES |
| 湿生扁蕾 Gentianopsis paludosa | 6.98 | 15.05* | 17.77** | 16.99** | 0.55 | 3.94* | 2.03 | 3.17 | 0.058* | RS, ES |
| 银莲花 Anemone obtusiloba | 1.05 | 11.88* | 8.32** | 1.85 | 2.95 | 2.69 | 1.3 | 2.79 | 0.125** | RS, ES |
| 圆萼刺参 Morina chinensis | 1.47 | 13.77* | 0 | 0.22 | 0 | 1.32 | 0.59 | 0.86 | 0.079* | RS, ES |
| 早熟禾 Poa sp. | 1.13 | 13.76* | 52.94** | 1.37 | 7.17** | 1.51 | 0.88 | 1.46 | 0.319** | RS, ES |
| 响应物种数(%) No. of response species (%) | 2 (2.8) | 11 (15.5) | 20 (28.4) | 2 (2.8) | 5 (7.0) | 3 (4.2) | 0 (0.0) | 1 (1.4) | 13 (18.3) | |
B, 区组; C, 刈割; ES, 作用物种; F, 施肥; RS, 响应物种; W, 浇水; ×, 交互作用; *, p < 0.05; **, p < 0.01。
B, block; C, clipping; ES, effect species; F, fertilizing; RS, response species; W, watering; ×, interaction; *, p < 0.05; **, p < 0.01.
表2 全部物种数及各类物种所占比例在不同处理间差异的方差分析
Table 2
| 物种类型 Species type | 不施肥 Unfertilized | 施肥 Fertilized | 施肥效应 Fertilized effect | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 不刈割 Unclipped | 中度刈割 Stubbled 3 cm | 重度刈割 Stubbled 1 cm | F | 平均值 Mean | 不刈割Unclipped | 中度刈割Stubbled 3 cm | 重度刈割Stubbled 1 cm | F | 平均值 Mean | F | ||
| 全部物种数 No. of total species | 28.7b | 33.4a | 28.9b | 13.79*** | 30.4 | 22.8c | 30.1a | 27.1b | 17.64*** | 26.6 | 20.33*** | |
| 响应物种(%) Response species (%) | 58.6a | 56.3a | 60.8a | 2.89 | 58.6 | 54.9b | 53.1b | 56.7a | 15.20*** | 54.9 | 40.32*** | |
| 作用物种(%) Effect species (%) | 28.4b | 28.4b | 31.5a | 3.76* | 29.5 | 23.9b | 24.2b | 27.5a | 10.23*** | 25.2 | 48.50*** | |
| 共有物种(%) Common species (%) | 94.0a | 91.5b | 93.8a | 12.64*** | 93.1 | 95.1a | 92.6b | 94.7a | 15.21*** | 94.1 | 20.55*** | |
同行数值后相同字母表示无显著差异(p > 0.05), 不同字母表示差异显著(p < 0.05)。
Values within the same row with same letters are no significant difference at 0.05 level, and different letters are significant difference at 0.05 level.
2.2 物种多样性变化
刈割与施肥的交互作用仅对全部物种(F(2,18) = 10.521; p < 0.01)、响应物种(F(2,18) = 14.585; p < 0.01)和共有物种(F(2,18) = 10.144; p < 0.01)的多样性有显著影响, 施肥后的不刈割群落有明显下降(图1), 下降程度分别为5.8%、10.8%和5.8%。其他处理对物种多样性均无显著影响。
图1
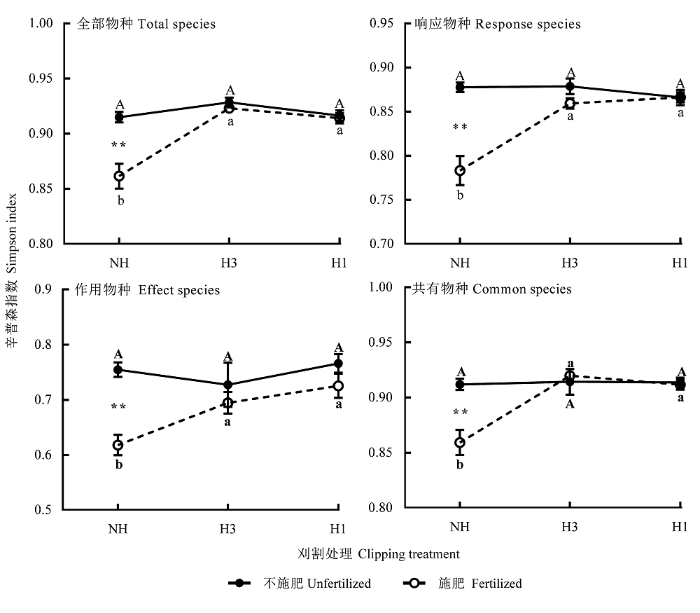
图1
刈割和施肥对物种多样性的影响(平均值±标准误差, n = 36)。
NH, 不刈割; H3, 中度刈割; H1, 重度刈割。相同的大写字母表示不施肥样方中刈割处理间无显著差异(p > 0.05); 相同的小写字母表示施肥样方中刈割处理间无显著差异(p > 0.05); **, p < 0.01。
Fig. 1
Effects of clipping and fertilizing on species diversity (mean ± SE, n = 36).
NH, unclipped; H3, stubbled 3 cm; H1, stubbled 1 cm. Same capital letters indicate no significant difference among clipping treatments in unfertilized quadrat (p < 0.05). Same small letters indicate no significant difference among clipping treatments in fertilized quadrat (p < 0.05); **, p < 0.01.
2.3 生态系统功能变化
刈割、施肥及刈割与施肥的交互作用对群落地上净初级生产力有显著影响(刈割: F(2,4) = 7.678, p <0.05; 施肥: F(1,18) = 80.311, p < 0.001; 刈割×施肥: F(2,18) = 4.081; p < 0.05)。不施肥时, 重度刈割群落的初级生产力比不刈割群落降低28.9%, 施肥后, 不刈割、中度刈割和重度刈割群落的初级生产力分别增加23.9%、42.8%和67.5%, 并引起初级生产力在不刈割和中度刈割群落以及中度和重度刈割群落间的差异消失(图2)。回归分析结果显示, 四类物种的密度对群落初级生产力的影响各异, 其中, 共有物种的密度对群落初级生产力影响最显著, Y = 6.083-0.325X (R2 = 0.153, F = 6.14, p < 0.05, Y =初级生产力, X =共有物种密度)
图2
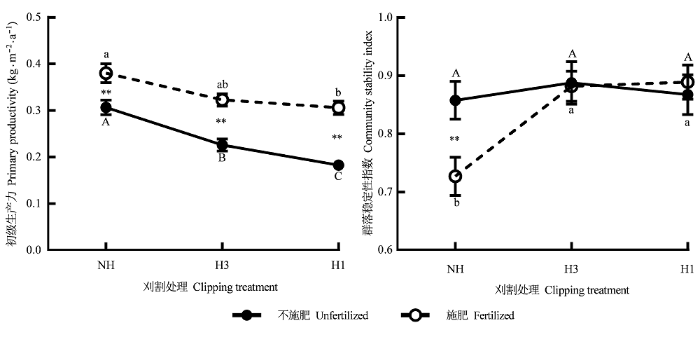
图2
不同处理对高寒草甸生态系统功能的影响(平均值±标准误差, n = 36)。
NH, 不刈割; H3, 中度刈割; H1, 重度刈割; **, p < 0.01; 相同的大写字母表示不施肥样方中刈割处理间无显著差异(p > 0.05); 相同的小写字母表示施肥样方中刈割处理间无显著差异(p > 0.05)。
Fig. 2
Effects of different treatments on ecosystem function in alpine meadow (mean ± SE, n = 36).
NH, unclipped; H3, stubbled 3 cm; H1, stubbled 1 cm; **, p < 0.01. Same capital letters indicate no significant difference among clipping treatments in unfertilized quadrat (p > 0.05). Small letters indicate no significant difference among clipping treatments in fertilized quadrat (p > 0.05).
群落稳定性ICV仅受刈割与施肥交互作用的影响(F(2,18) = 3.86, p < 0.05), 主要表现在施肥后引起不刈割群落的稳定性比不施肥群落下降15.3%, 而不施肥时, 稳定性在刈割处理间无显著差异(p > 0.05) (图2)。
2.4 物种多样性与生态系统功能
2.4.1 物种多样性与生产力的关系
表3 物种多样性与群落地上净初级生产力关系的回归分析
Table 3
| 物种类型 Species type | 施肥梯度 Fertility gradient | 刈割梯度 Clipping gradient | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 不施肥 Unfertilized | 施肥 Fertilized | 不刈割 Unclipped | 中度刈割 Stubbled 3 cm | 重度刈割 Stubbled 1 cm | |||||||||||||||
| K | F | R2 | K | F | R2 | K | F | R2 | K | F | R2 | K | F | R2 | |||||
| 全部物种 Total species | 2.90 | 2.7 | 0.05 | -0.84 | 1.4 | 0.03 | -0.71 | 0.7 | 0.02 | -1.00 | 0.1 | 0 | 2.41 | 0.8 | 0.02 | ||||
| 响应物种 Response species | 1.31 | 1.4 | 0.03 | -1.13 | 5.5* | 0.10 | -1.58 | 10.3** | 0.23 | -0.16 | 0 | 0 | 0.80 | 0.2 | 0.01 | ||||
| 作用物种 Effect species | 0.03 | 0 | 0 | -0.28 | 0.8 | 0.02 | -0.86 | 5.1* | 0.13 | 0.02 | 0 | 0 | -0.54 | 0.7 | 0.02 | ||||
| 共有物种 Common species | 2.84 | 2.6 | 0.05 | -0.81 | 1.3 | 0.02 | -0.68 | 0.6 | 0.02 | -0.55 | 0 | 0 | 2.62 | 0.9 | 0.03 | ||||
| 样方数 No. of quadrat | 54 | 54 | 36 | 36 | 36 | ||||||||||||||
K, 回归斜率; *, p < 0.05; **, p < 0.01。
K, regression slope; *, p < 0.05; **, p < 0.01.
图3
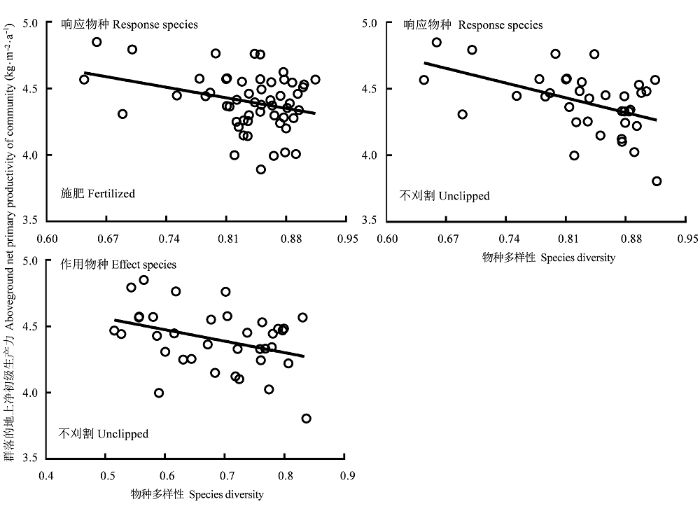
图3
物种多样性与群落地上净初级生产力的关系(对数尺度)。
Fig. 3
Relationships between species diversity and aboveground net primary productivity of community (log-scale).
2.4.2 物种多样性与稳定性的关系
表4 物种多样性与群落稳定性关系的回归分析
Table 4
| 物种类型 Species type | 施肥梯度 Fertility gradient | 刈割梯度 Clipping gradient | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 不施肥 Unfertilized | 施肥 Fertilized | 不刈割 Unclipped | 中度刈割 Stubbled 3 cm | 重度刈割 Stubbled 1 cm | |||||||||||||||
| K | F | R2 | K | F | R2 | K | F | R2 | K | F | R2 | K | F | R2 | |||||
| 全部物种 Total species | 6.18 | 161.8*** | 0.76 | 2.88 | 97.7*** | 0.65 | 2.85 | 85.8*** | 0.72 | 7.16 | 74.6*** | 0.69 | 5.52 | 62.1*** | 0.65 | ||||
| 响应物种 Response species | 2.39 | 22.3*** | 0.30 | 1.70 | 45.8*** | 0.47 | 1.31 | 19.5*** | 0.36 | 2.14 | 13.2*** | 0.28 | 2.95 | 28.1*** | 0.45 | ||||
| 作用物种 Effect species | 0.59 | 13.5*** | 0.21 | 0.83 | 22.0*** | 0.30 | 0.79 | 11.6** | 0.25 | 0.51 | 12.3** | 0.27 | 0.77 | 10.2** | 0.23 | ||||
| 共有物种 Common species | 6.18 | 160.4*** | 0.76 | 2.90 | 101.3*** | 0.66 | 2.83 | 87.3*** | 0.72 | 7.15 | 80.4*** | 0.70 | 5.79 | 66.2*** | 0.66 | ||||
| 样方数 No. of quadrats | 54 | 54 | 36 | 36 | 36 | ||||||||||||||
K, 回归斜率; **, p < 0.01; ***, p < 0.001。
K, regression slope; **, p < 0.01; ***, p < 0.001.
图4
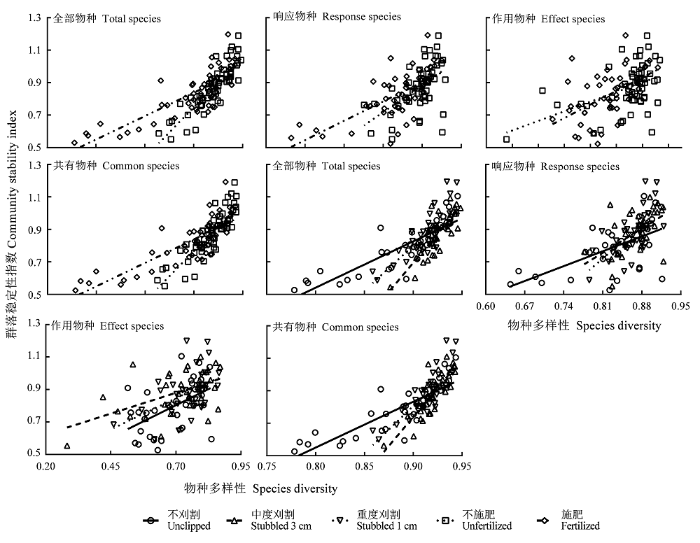
图4
物种多样性与群落稳定性的关系。
Fig. 4
Relationship between species diversity and community stability.
2.5 不同处理对稳定性参数的影响及其与物种多样性的关系
2.5.1 不同处理对稳定性参数的影响
方差分析显示, 各处理对物种的总密度均无显著影响。刈割和浇水对总方差也无显著影响(p > 0.05), 但施肥显著增加了全部物种的总方差(F(1,18)= 9.73, p < 0.01) (图5), 各处理对物种的同步性也无显著影响(p > 0.05)。
图5
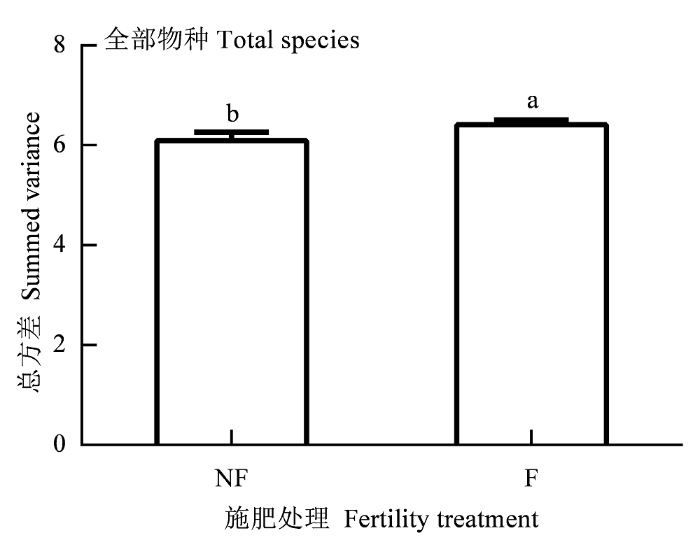
图5
施肥处理对全部物种总方差的影响(平均值±标准误差, n = 36)。
F, 施肥; NF, 不施肥。不同的小写字母表示差异显著(p < 0.05)。
Fig. 5
Effect of fertility treatment on summed variance of total species (mean ± SE, n = 36).
F, Fertilized; NF, unfertilized. Different small letters indicate significant difference (p < 0.05).
2.5.2 物种多样性与稳定性参数的关系
图6
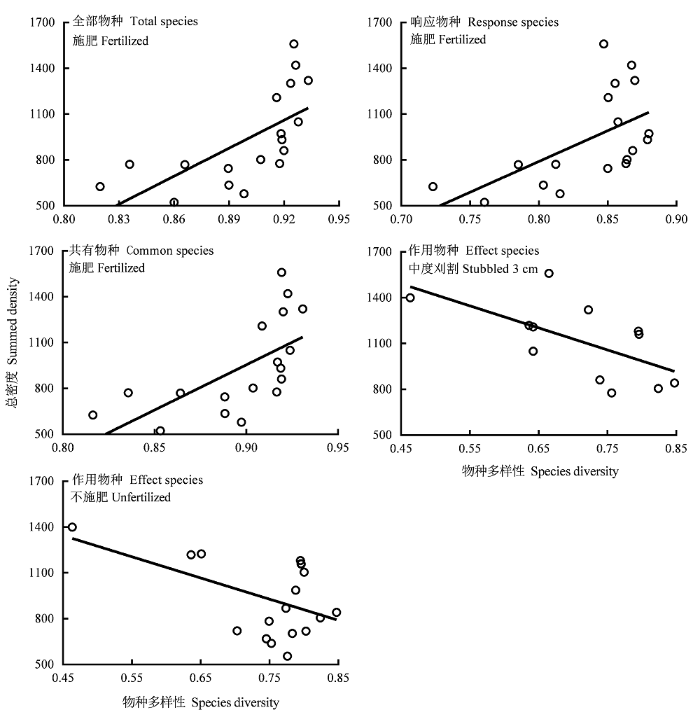
图6
物种多样性与群落物种总密度的关系。
Fig. 6
Relationships between species diversity and summed density of community.
4类物种的多样性与群落总方差在不施肥时显著负相关, 全部物种、响应物种、作用物种和共有物种的多样性变化分别解释了总方差变异的32%、25%、43%和50%, 但施肥后均变为无相关(表5; 图7), 回归斜率在不施肥与施肥处理间差异显著(pTS = 0.009, pRS = 0.031, pES = 0.020, pCS = 0.001)。全部物种和共有物种的多样性与群落总方差之间在不刈割及中度刈割群落均为显著负相关, 在重度刈割群落无相关。其多样性对总方差变异的解释能力由不刈割的54%和54%降至中度刈割的37%和53%(表5); 响应物种在两个刈割的群落为显著负相关, 其多样性对总方差变异的解释能力为27%-37%; 作用物种仅在中度刈割群落为显著负相关, 但能解释总方差变异的70% (表5; 图7)。
表5 物种多样性与群落稳定性参数关系的回归分析
Table 5
| 参数 Parameter | 物种类型 Species type | 施肥梯度 Fertility gradient | 刈割梯度 Clipping gradient | R2 的平均值 Mean of R2 | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 不施肥 Unfertilized | 施肥 Fertilized | 不刈割 Unclipped | 中度刈割 Stubbled 3 cm | 重度刈割 Stubbled 1 cm | |||||||||||||||||
| K | F | R2 | K | F | R2 | K | F | R2 | K | F | R2 | K | F | R2 | |||||||
| 总密度 Summed density | 全部物种Total species | -2 962.36 | 0.6 | 0.04 | 6 116.24 | 12.5** | 0.44 | 608.86 | 0.2 | 0.02 | -5 788.52 | 0.9 | 0.08 | 1 783.14 | 0.1 | 0.01 | 0.12 | ||||
| 响应物种Response species | -3 017.15 | 1.8 | 0.10 | 4 027.39 | 7.9** | 0.33 | 1 174.81 | 2.2 | 0.18 | -4 791.46 | 3.2 | 0.24 | -3 290.57 | 0.9 | 0.09 | 0.19 | |||||
| 作用物种Effect species | -1 387.72 | 5.2* | 0.25 | 1 854.35 | 3.2 | 0.17 | 919.64 | 2.6 | 0.21 | -1 443.65 | 6.1* | 0.38 | -333.98 | 0.1 | 0.01 | 0.20 | |||||
| 共有物种Common species | -3 983.58 | 2 | 0.11 | 5 920.40 | 11.1** | 0.41 | 622.84 | 0.2 | 0.02 | -3 339.45 | 1.0 | 0.09 | 2 521.8 | 0.2 | 0.02 | 0.13 | |||||
| 总方差 Summed variance | 全部物种Total species | -25.69 | 7.7** | 0.32 | -1.36 | 0.2 | 0.01 | -12.74 | 11.9** | 0.54 | -31.64 | 6.0* | 0.37 | -19.37 | 3.8 | 0.27 | 0.30 | ||||
| 响应物种Response species | -13.79 | 5.2* | 0.25 | -0.42 | 0 | 0 | -5.07 | 2.6 | 0.21 | -15.23 | 5.8* | 0.37 | -18.19 | 11.3** | 0.53 | 0.27 | |||||
| 作用物种Effects species | -5.28 | 11.8** | 0.43 | 0.19 | 0 | 0 | -2.93 | 1.5 | 0.13 | -5.06 | 23.1** | 0.70 | -3.80 | 2.8 | 0.22 | 0.30 | |||||
| 共有物种Common species | -24.51 | 16.1** | 0.50 | -1.63 | 0.3 | 0.02 | -12.61 | 11.7** | 0.54 | -20.84 | 11.2** | 0.53 | -19.44 | 3.0 | 0.23 | 0.36 | |||||
| 同步性 Synchrony | 全部物种Total species | -0.19 | 37.1** | 0.70 | -0.10 | 48.0** | 0.75 | -0.10 | 12.7** | 0.56 | -0.18 | 51.9** | 0.84 | -0.11 | 6.1* | 0.38 | 0.65 | ||||
| 响应物种Response species | -0.09 | 12.7** | 0.44 | -0.08 | 57.5** | 0.78 | -0.05 | 8.0* | 0.44 | -0.08 | 26.3** | 0.72 | -0.09 | 9.7** | 0.49 | 0.57 | |||||
| 作用物种Effects species | -0.02 | 8.4** | 0.34 | -0.03 | 7.0** | 0.30 | -0.04 | 5.5* | 0.35 | -0.02 | 20.2** | 0.67 | -0.02 | 1.8 | 0.15 | 0.36 | |||||
| 共有物种Common species | -0.09 | 6.5* | 0.29 | -0.10 | 47.6** | 0.75 | -0.09 | 12.2** | 0.55 | -0.05 | 2.8 | 0.22 | -0.12 | 5.8* | 0.37 | 0.44 | |||||
| 样方数 No. of quadrats | 18 | 18 | 12 | 12 | 12 | ||||||||||||||||
K, 回归斜率。**, p < 0.01; *, p < 0.05。
K, regression slope.**, p < 0.01; *, p < 0.05.
图7
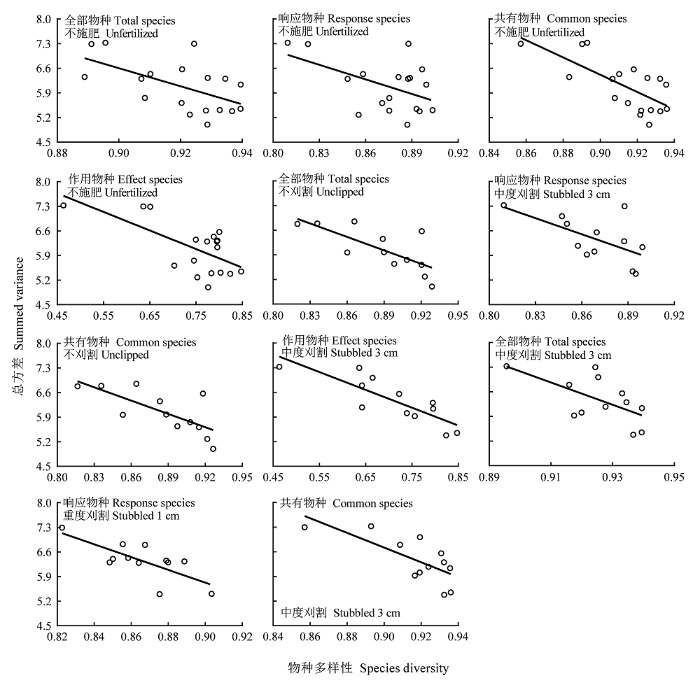
图7
物种多样性与群落总方差的关系。
Fig. 7
Relationships between species diversity and summed variance of community.
在对数尺度上进行的线性回归分析显示, 4类物种的密度总方差与其平均密度在所有处理中均存在显著的正相关关系(p < 0.001), 其z值均大于2 (图8), 这表明投资组合效应普遍存在于各类物种中, 但不同处理对该效应的影响不同。尽管施肥后4类物种的z值仍大于2, 但与不施肥相比显著减小(pTS = 0.015; pRS = 0.005; pES = 0.007; pCS = 0.014), 降低了该效应; 而刈割对z值的影响相对较小, 与不刈割相比, 仅中度和重度刈割分别引起作用物种和响应物种z值的增大(pES = 0.004)和减小(pRS = 0.003), 其他类物种的z值不受刈割的影响。但就变化趋势看, 除作用物种外, 其他类物种的z值均随刈割强度增加而减小。
不论施肥与否, 4类物种的物种多样性均与同步性显著负相关(表5; 图9)。施肥后全部物种、响应物种、作用物种和共有物种的多样性变化分别解释了同步性变异的75%、78%、30%和75%, 总体上高于不施肥时。施肥仅显著减小了全部物种的回归斜率(pTS = 0.013), 对其他3类物种则无影响(p > 0.05)。除作用物种和共有物种的物种多样性与同步性分别在重度和中度刈割群落无显著相关外, 其余4类物种在各刈割处理群落中均表现为显著负相关(表5)。刈割强度并未改变4类物种的回归斜率(p > 0.05), 全部物种、响应物种、作用物种和共有物种的多样性变化对同步性变异的解释能力分别为38%-84%、44%-72%、15%-67%和22%-55% (表5)。
图9
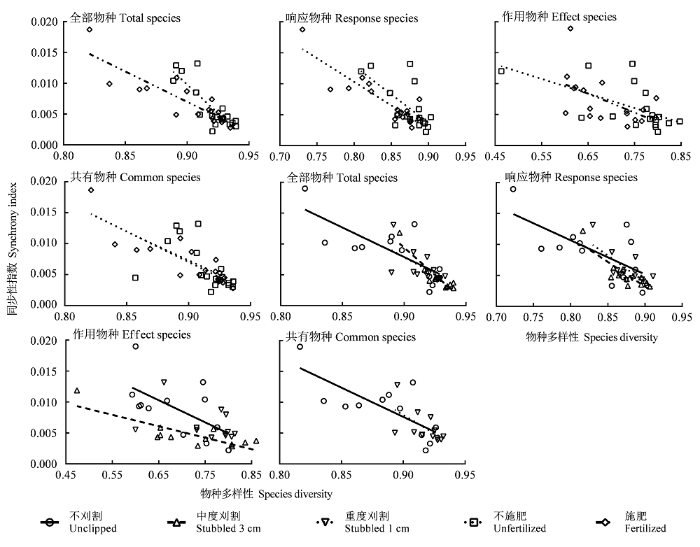
图9
物种多样性与群落同步性的关系。
Fig. 9
Relationships between species diversity and synchronism of community.
3 讨论和结论
3.1 物种多样性的生态系统功能效应
本研究根据群落物种重要值对刈割和土壤资源变化的响应与周转以及是否影响群落净初级生产力, 将群落物种划分为4种类型。研究结果表明, 物种多样性对生态系统净初级生产力和稳定性具有不同的效应, 且该效应源于不同类型的物种, 即少数物种的多样性变化影响着净初级生产力, 且为负效应(Hooper, 1998), 而多数物种的多样性变化决定着稳定性, 为正效应(Tilman, 1999; Cottingham et al., 2001; Valone & Hoffman, 2003; Tilman et al., 2006)。全部物种以及占64.8%的共有物种的多样性变化对净初级生产力无影响, 而占36.6%和18.3%的响应物种和作用物种的多样性与生产力负相关, 但其解释能力最高仅为23% (响应物种, 施肥梯度上不刈割群落) (表3; 图3), 然而共有物种的密度显著影响地上净初级生产力, 且为负效应, 4类物种的多样性对系统稳定性均具有正效应, 且共有物种的贡献最高, 响应物种和作用物种多样性的贡献则相对较低(表4; 图4)。这也说明群落物种多样性变化对净初级生产力的贡献可能低于其对稳定性的贡献, 更准确地说,是不同物种执行着不同的生态系统功能, 物种的“功能身份(functional identity)”可能比物种多样性本身更为关键, 从而表现出不同物种多样性效应的分异性。因此, 不加区别地笼统定义物种多样性与生态系统功能的关系可能欠妥。
3.2 物种多样性对群落稳定性的作用机制
尽管刈割梯度上全部物种、响应物种和共有物种的多样性与总多度在施肥后均极显著正相关(表5), 但并不表示高寒草甸生态系统在解除养分限制后是超产效应导致稳定性提高, 因为施肥后4类物种的多样性均有显著降低(图1), 而总多度并未发生显著变化(p > 0.05), 因此本研究中物种多样性与总多度间的正相关关系完全是由施肥后多样性的降低所致, 并不存在物种多样性本身增加后通过超产效应提高稳定性的证据。这与Fox (2003)的模型研究结果一致, 即泛化食草动物的取食(相当于本研究中刈割去除所有物种)导致物种共存时, 超产不可能发生(张全国和张大勇, 2003)。
尽管有研究表明物种的补偿机制(异步性)可以增加群落稳定性(Gonzalez & Loreau, 2009), 但也有研究对生态位互补效应是否具有重要作用提出质疑(Grace et al., 2007), 本研究并未发现异步性效应是提高群落稳定性的重要机制。因为尽管在本研究90%的实验处理-物种组合中, 4类物种的物种多样性与同步性之间均表现为显著负相关(表5; 图9), 可能意味着物种多样性增加会引起物种异步性的升高, 但由于各类物种的同步性在不同实验处理下并无显著变化(p > 0.05), 因此物种多样性与群落同步性间的负相关关系完全是由实验处理引起的多样性变化所致, 而异步性实际上并未发生相应改变, 因此也不存在异步性效应提高稳定性的证据。
但本研究普遍存在投资组合效应引起群落稳定性增加的证据。一是因为在所有实验处理中4类物种密度的总方差与其平均密度之间均存在显著的正相关(p < 0.001), 其幂函数的z值均大于2 (图8); 二是因为物种多样性与群落总方差在55%的实验处理-物种组合中均为显著负相关(表5); 三是因为刈割、施肥和浇水对物种的总密度无影响(p > 0.05), 但施肥却显著增加了每类物种的总方差(p < 0.01), 并降低了多样性(图1)。这符合发生投资组合效应的假定(Doak et al., 1998), 即在群落总密度不变的条件下, 物种多样性增加将减小群落物种密度的变异性, 从而提高群落稳定性。而且4类物种中, 共有物种的多样性变化对不同处理群落总方差的平均解释能力(即R2平均值, 表5)最高。因此, 可以认为高寒草甸群落稳定性的维持主要依赖于群落中共有物种的多样性增加, 其作用机制是投资组合效应, 而超产效应和异步性效应对稳定群落并无作用。
3.3 刈割和施肥对物种多样性和生态系统功能的影响
上述效应与4类物种的响应差异有关。例如, 刈割并未引起作用物种的多样性变化, 但施肥后其多样性降低程度最大(图1); 与之相反, 刈割后全部物种、响应物种和共有物种的多样性均有增加, 但施肥后其降低程度较低(图1)。这说明直接影响净初级生产力的少数作用物种的多样性受资源变化的影响更大, 而其他物种的多样性受刈割扰动的影响更大, 刈割后群落稳定性的提高主要与群落中大量物种的多样性增加以及群落优势度等级的调整引起稀有物种的补充有关。这与Sasaki和Lauenroth (2011)去除天然草地优势种的研究结果一致。
很多研究也都显示施肥具有提高净初级生产力(Tilman, 1982)、降低物种多样性和群落稳定性的作用(Lepš, 2004; Grman et al., 2010)。但本研究还同时表明, 尽管施肥后刈割群落的生产力增加更多(不刈割、中度和重度刈割群落分别增加23.9%、42.8%和67.5%), 但重度刈割群落的生产力仍低于不刈割群落(图2)。说明施肥后高寒草甸的补偿生长能力尚不足以抵消长期重度刈割的负效应, 这与我们过去关于该群落补偿生长的研究结果一致(周晓松等, 2011)。我们的其他研究显示, 在存有大量家畜粪尿、重度放牧的畜圈生境, 也会发生地上生物量的超补偿(席博等, 2010), 但其物种多样性很低, 仅由少量耐牧性强、并能快速生长的物种占优势(臧岳铭等, 2009)。结合本研究中群落物种多样性、净初级生产力和稳定性在模拟刈割和施肥梯度上的变化结果(图1, 图2), 我们认为, 将放牧强度控制在45%-50%的采食率是维持高寒草甸生态系统较高净初级生产力和物种多样性, 并保持长期稳定的首要前提。
致谢
感谢中国科学院海北高寒草甸生态系统定位研究站的支持。野外取样得到周晓松、李晓刚、许曼丽、李燕和关百盈等人的大力帮助, 特致谢忱。
参考文献
Biodiversity may regulate the temporal variability of ecological system
Vive la difference: plant functional diversity matters to ecosystem processes
The statistical inevitability of stability-diversity relationships in community ecology
The long-term relationship between plant diversity and total plant biomass depends on the mechanism maintaining diversity
The causes and consequences of compensatory dynamics in ecological communities
Does species diversity limit productivity in natural grassland communities?

Theoretical analyses and experimental studies of synthesized assemblages indicate that under particular circumstances species diversity can enhance community productivity through niche complementarity. It remains unclear whether this process has important effects in mature natural ecosystems where competitive feedbacks and complex environmental influences affect diversity-productivity relationships. In this study, we evaluated diversity-productivity relationships while statistically controlling for environmental influences in 12 natural grassland ecosystems. Because diversity-productivity relationships are conspicuously nonlinear, we developed a nonlinear structural equation modeling (SEM) methodology to separate the effects of diversity on productivity from the effects of productivity on diversity. Meta-analysis was used to summarize the SEM findings across studies. While competitive effects were readily detected, enhancement of production by diversity was not. These results suggest that the influence of small-scale diversity on productivity in mature natural systems is a weak force, both in absolute terms and relative to the effects of other controls on productivity.
Benefits of plant diversity to ecosystems: immediate, filter and founder effects
Mechanisms contributing to stability in ecosystem function depend on the environmental context
Primary productivity and species abundance of cultivated grassland under different clipping and seeding combinations
刈割留茬高度和不同播种组合对人工草地初级生产力和物种丰富度的影响
Biodiversity in a complex world: consolidation and progress in functional biodiversity research
The role of complementarity and competition in ecosystem responses to variation in plant diversity
Effects of biodiversity on ecosystem functioning: a consensus of current knowledge
Biodiversity, productivity and the temporal stability of productivity: patterns and processes
Theory predicts that the temporal stability of productivity, measured as the ratio of the mean to the standard deviation of community biomass, increases with species richness and evenness. We used experimental species mixtures of grassland plants to test this hypothesis and identified the mechanisms involved. Additionally, we tested whether biodiversity, productivity and temporal stability were similarly influenced by particular types of species interactions. We found that productivity was less variable among years in plots planted with more species. Temporal stability did not depend on whether the species were planted equally abundant (high evenness) or not (realistically low evenness). Greater richness increased temporal stability by increasing overyielding, asynchrony of species fluctuations and statistical averaging. Species interactions that favoured unproductive species increased both biodiversity and temporal stability. Species interactions that resulted in niche partitioning or facilitation increased both productivity and temporal stability. Thus, species interactions can promote biodiversity and ecosystem services.
Biodiversity, stability and productivity in competitive communities
Variability in population and community biomass in a grassland community affected by environmental productivity and diversity
Effects of clipping, fertilizing and watering on the relationship between species diversity, functional diversity and primary productivity in alpine meadow of China
刈割、施肥和浇水对高寒草甸物种多样性、功能多样性与初级生产力关系的影响
Analyses on climates and vegetation productivity background at Haibei alpine meadow ecosystem research station
海北高寒草甸生态系统定位站气候、植被生产力背景的分析
Seasonal frozen soil and its effect on vegetation production in Haibei alpine meadow
海北高寒草甸的季节冻土及在植被生产力形成过程中的作用
Species synchrony and its drivers: neutral and nonneutral community dynamics in fluctuating environments
Measurement of biotic community diversity I α diversity (Part 2)
生物群落多样性的测度方法I. α多样性的测度方法(下)
Dominant species, rather than diversity, regulates temporal stability of plant communities
Secondary succession and the pattern of plant dominance along experimental nitrogen gradients
The ecological consequences of changes in biodiversity: a search for general principles
Biodiversity and ecosystem stability in a decade-long grassland experiment
A mechanistic examination of diversity-stability relationship in annual plant communities
Relationship between species diversity and productivity in four types of alpine meadow plant communities
高寒草甸不同草地群落物种多样性与生产力关系研究
Effect of grazing disturbance and nutrient availability on the compensatory responses of community in alpine meadows
放牧强度和生境资源对高寒草甸群落补偿能力的影响
The effects of long-term fertilization on the temporal stability of alpine meadow communities
Effects of species diversity and functional diversity on primary productivity of alpine meadow
高寒矮嵩草草甸物种多样性与功能多样性对初级生产力的影响
Biodiversity and ecosystem functioning: recent advances and controversies
生物多样性与生态系统功能: 进展与争论
Biodiversity and ecosystem functioning: recent advances and trends
生物多样性与生态系统功能: 最新的进展与动向
Community compensatory mechanism under clipping, fertilizing and watering treatment in alpine meadow
刈割、施肥和浇水对高寒草甸物种多样性、功能多样性与初级生产力关系的影响
Response of Polygonum viviparum species and community level to long-term livestock grazing in alpine shrub meadow in Qinghai-Tibet Plateau
Grazing by domestic herbivores is generally recognized as a major ecological factor and an important evolutionary force in grasslands. Grazing has both extensive and profound effects on individual plants and communities. We investigated the response patterns of Polygonum viviparum species and the species diversity of an alpine shrub meadow in response to long-term livestock grazing by a field manipulative experiment controlling livestock numbers on the Qinghai-Tibet Plateau in China. Here, we hypothesize that within a range of grazing pressure, grazing can alter relative allocation to different plant parts without changing total biomass for some plant species if there is life history trade-offs between plant traits. The same type of communities exposed to different grazing pressures may only alter relative species' abundances or species composition and not vary species diversity because plant species differ in resistant capability to herbivory. The results show that plant height and biomass of different organs differed among grazing treatments but total biomass remained constant. Biomass allocation and absolute investments to both reproduction and growth decreased and to belowground storage increased with increased grazing pressure, indicating the increasing in storage function was attained at a cost of reducing reproduction of bulbils and represented an optimal allocation and an adaptive response of the species to long-term aboveground damage. Moreover, our results showed multiform response types for either species groups or single species along the gradient of grazing intensity. Heavy grazing caused a 13.2% increase in species richness. There was difference in species composition of about 18%-20% among grazing treatment. Shannon-Wiener (H') diversity index and species evenness (E) index did not differ among grazing treatments. These results support our hypothesis.
Compensatory growth of Carex scabrirostris in different habitats in alpine meadow
高寒草甸不同生境粗喙薹草补偿生长研究
Predicting plant traits and functional types response to grazing in an alpine shrub meadow on the Qinghai-Tibet Plateau

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}