Plant sesquiterpenes induce hyphal branching in arbuscular mycorrhizal fungi
2
2005
... 植物的根系分泌物中含有一些具有生物活性的大分子和小分子次生代谢产物, 这些物质调节着根系周围的土壤微生物种群与数量, 调节着植物与微生物之间的根际对话(rhizosphere talk).Peters等(1986)发现, 在缺氮条件下, 豆科植物根系通过分泌黄酮类和异黄酮类物质来诱导启动根瘤菌结瘤基因(nodD)的表达, 最终导致根瘤菌侵染根系并形成根瘤.Akiyama等(2005)研究发现, Lotus japonicus的根系分泌物中含有一种分枝因子(branching factor: 5-deoxystrigol), 在很低浓度时即可刺激菌根真菌萌发孢子的菌丝大量分枝. ...
... 真菌作为土壤微生物群落的主要组分之一, 在根际与植物形成密切关系, 尤其是丛枝菌根(arbuscular mycorrhizae, AM)能与大多数高等植物形成共生关系.丛枝菌根几乎在各种陆地生态系统中都有分布.许多研究表明菌根真菌和菌根植物之间存在互相感知的系统, 菌根植物的根系分泌物中存在能被菌根真菌识别的物质, 如来自Lotus japonicus的独脚金萌发素内酯(5-deoxystrigol)(Buee et al., 2000; Akiyama et al., 2005).在菌根真菌与植物的对话中, 植物产生的根系分泌物为菌根真菌的生长繁殖提供了丰富的碳源和能量, 而菌根真菌也能够帮助植物改善营养, 如通过溶解释放无机磷或水解有机磷来提高植物对磷元素的吸收利用, 抵御各种生物和非生物胁迫.然而也有研究发现, 植物与菌根真菌的关系也不是一成不变的, 在特定的环境下, 二者可能从互利共生变成寄生关系, 比如当为植物提供足够的化肥和有机肥时, 菌根真菌为植物提供磷元素的效益就可能无法补偿植物提供给菌根真菌碳源的成本, 从而引发二者关系的变化(Joh- nson et al., 1997).内生真菌(fungal endophyte)也与植物关系密切, 微生物学家Rusty J. Rodriguez甚至使用了一个新词“symbiogenics” (共生基因学)或称为“symbiosis-altered genetic expression” (共生改变基因表达)来形象地描述二者的关系.他的研究发现将分离自耐旱沙丘草本植物中的内生真菌与水稻共培养, 不但可以显著降低水稻对水分的需求量, 而且还可以促进水稻生长, 提高产量.同样, 将来自喜热稗的内生真菌与小麦共培养后, 小麦可在70 ℃高温下生长, 并且对水分的需求量减半(East, 2013). ...
Potential impact of soil microbiomes on the leaf metabolome and on herbivore feeding behavior
1
2013
... 植物根系分泌物的产生是植物根部长期进化的结果, 是适应环境的一种体现, 不是一个完全被动的过程, 因为植物的任何代谢功能都直接或间接地消耗碳源和能量.由于根部一直处于土壤中, 无时无刻不与周围的生物、非生物因子做斗争, 所以植物能够根据周围环境的变化不断地调整自身根系分泌物的组成和数量来积极响应(Walker et al., 2003).在根际中, 根系分泌物对某些微生物具有吸引作用, 这类具有趋化性的细菌或真菌就能够在根际中大量聚集和繁殖, 比如豆科植物对根瘤菌的诱导; 而对有些病原菌却具有抑制作用, 比如小麦根系分泌物中的酚酸具有抗细菌的活性, 并且其抗菌活性具有协同作用(Wu et al., 2001).Fierer等(2012)运用宏基因组学技术对取自寒漠、热沙漠、森林、草原、苔原的16份土样的微生物群落结构进行分析, 发现来自沙漠区的土壤微生物群落明显区别于非沙漠区的, 进一步的宏基因组功能分析发现来自沙漠区的微生物类群中与营养循环、分解代谢植物源有机化合物相关的基因含量很低, 尤其是来自寒漠地区的微生物群落, 其种类多样性和功能多样性都最低, 可见地上部物种多样性水平显著影响地下部微生物群落结构与功能多样性.相反, 地下部微生物群落也会对地上部植物的生理代谢、生长发育等产生重要影响.Badri等(2013)运用气相色谱-质谱联用仪(GC-MS)分析发现, 土壤微生物群落对植物叶片的代谢物组分有显著影响, 进而影响昆虫对叶片的取食行为.Mendes等(2011)运用PhyloChip芯片技术对抑病型土壤(disease suppressive soil)和利病型土壤(disease conducive soil)中的微生物群落结构进行分析, 发现种植于抑病型土壤的甜菜(Beta vulgaris)其根际优势群落普遍为拮抗病原菌相关的微生物, 如放线菌门、β变形菌门、γ变形菌门等, 尤其是其中的假单胞菌在抑病型土壤中的数量极显著高于利病型土壤中, 这是由根系分泌物介导下植物与根际特异微生物共同作用的结果.Zolla等(2013)研究发现, 在适度干旱胁迫下将拟南芥幼苗种植于同域土壤(sympatric soil, 即长期种植拟南芥的土壤)中, 可以显著地促进植物生长、提高生物量, 而种植于非同域土壤(non-sympatric soil, 如长期种植松树和玉米的土壤)中对植物生物量没有影响.进一步研究还发现, 这是由于同域土壤中存在很多与降低植物干旱胁迫响应相关的促生根际菌(plant growth promoting rhizobacteria, PGPR), 如伯克氏菌属、芽孢杆菌属、不动杆菌属等.Lakshmanan等(2012)研究还发现, 番茄(Lycopersicon esculentum)叶片受到病原菌侵染后, 可通过调节根系分泌物组分与含量, 如增加根系苹果酸分泌释放量, 使更多的苹果酸进入根际, 从而招募更多的有益菌向根际聚集, 这些有益菌可进一步引发植物的诱导性系统抗性(induced systemic resistance)以对病原菌产生防御反应, 可见根系分泌物在植物-微生物互作关系中所扮演的重要角色. ...
Root specific elicitation and antimicrobial activity of rosmarinic acid in hairy root cultures of Ocimum basilicum
1
2002a
... 由于土壤中不乏病原真菌, 植物也进化形成了各种抵御病原真菌的有效机制.Bais等(2002a)研究发现, 当受到终极腐霉菌Pythium ultimum侵害时, Ocimum basilicum根系会大量分泌迷迭香酸, 从而减轻该病菌的危害, 并杀灭多种土壤细菌和病原体, 显示出强大的抑菌活性.也有学者研究发现, Phytolacca americana的发根会向根际分泌许多抗真菌活性蛋白, 如核糖体失活蛋白(RIP)、几丁质酶等(Park et al., 2002a).RIP通过具N-糖苷酶活性来抑制蛋白合成, 或选择性地删除28S rRNA保守区的腺嘌呤残基(Nielsen & Boston, 2001; Vepachedu et al., 2003).根系分泌的RIP还具有抗真菌特性, 如能够抑制根腐菌Rhizoctonia solani和Trichoderma reesei (Park et al., 2002b). ...
Exudation of fluorescent β-carbolines from Oxalis tuberosa L. roots
1
2002b
... 当然, 土壤中也存在一些危害植物生长的有害土壤动物区系, 如植物病原线虫等, 但是某些植物能够通过释放特定的根系分泌物来抵御有害动物的威胁.Wu等(1999)通过体外发根-蚜虫共培试验发现, 在蚜虫的刺激下, 植物能够大量分泌一种植物抗毒素——聚乙烯.Bais等(2002b)发现, Oxalis tuberosa的根系分泌物中含有一种会发荧光的物质——β咔巴啉生物碱衍生物, 经鉴定, 为骆驼蓬碱(harmine)和骆驼蓬碱二氢(harmaline), 有抗菌杀虫作用.Larson等(1988)也发现, β咔巴啉生物碱衍生物对某些细菌、土壤动物具有光毒性. ...
Allelopathy and exotic plant invasion: from molecules and genes to species interactions
1
2003
... 植物根系分泌物中含有一类对邻近其他植物或自身产生抑制或促进作用的生物活性物质, 即为化感物质(allelochemicals).Tang和Young (1982)研究了Hemarthria altissima根系分泌物的化感作用, 发现供体植物根系分泌物中对受体植物生长呈抑制作用的主要是酚类化合物.在小麦(Triticum aestivum)、水稻(Oryza sativa)、玉米等作物的根系分泌物中也都检测到一些化感物质, 对农业杂草都具有抑制作用, 而且化感作用的强弱与品种有关(何海斌等, 2005).同时, 研究发现黄瓜(Cucumis sativus)、大豆(Glycine max)、烟草(Nicotiana tabacum)、地黄(Rehmannia glutinosa)、西洋参(Panax quinquefolius)等忌连作作物的连作障碍现象与根系分泌物中的化感物质密切相关.Yu和Matsui (1994)发现黄瓜根系分泌物中含有苯甲酸、对羟基苯甲酸、2, 5-二羟基苯甲酸、苯丙烯酸等11种酚酸物质, 其中有10种具有生物毒性.Pramanik等(2000)也从黄瓜根系分泌物中鉴定出苯甲酸及其衍生物、肉桂酸及其衍生物等, 并证明这些物质会阻碍黄瓜对养分的吸收.近年来的研究也表明, 根系分泌物对某些入侵植物的成功入侵起到决定性的化学生态学作用.Bais等(2003)研究发现, 矢车菊(Centaurea maculosa)根系分泌释放的儿茶素(+)-Catechin和(-)-Catechin对其成功入侵起重要作用. ...
The diversity of life in soil. In: Bardgett RD ed
1
2005a
... 高等植物光合作用固定的碳20%-60%被转移到植物地下部分, 其中释放到土壤中的碳多年生植物最高可达其转移量的70%, 一年生植物最高可达其转移量的40% (Kuzyakov & Domanski, 2000).在 “根际”这一特殊的生态环境中, 植物根系不断地向土壤中分泌大量的有机物质, 形成根际沉积, 这为根际微生物提供了丰富的营养和能源.根系分泌物的种类和数量决定了根际微生物的种类和数量, 并对微生物的生长繁殖及代谢过程产生影响(Singh & Mukerji, 2006); 而根际微生物群落的动态变化反过来也影响着根际生态系统的物质循环和能量流动, 从而影响植物的生长发育及植被多样性变化(Bar- dgett, 2005a).由此可见, 植物和土壤、微生物之间的物质交换、养分循环是一个复杂的开放系统. ...
Linkages between plant and soil biological communities
1
2005b
... 除了细菌、放线菌、真菌3大主要微生物类群外, 土壤中还存在个体大小差异明显的各式各样的动物区系.根据土壤动物个体大小可以将其划分为: (1)微动物区系(<0.1 mm, 如原生动物、线虫等); (2)中型动物区系(0.1-2.0 mm, 如甲螨、原尾虫等); (3)广动物区系(>2 mm, 如蚯蚓、白蚁、马陆等) (Bardgett, 2005b).土壤动物作为生态系统物质循环中的重要消费者, 在生态系统中起着重要的作用. ...
The rhizosphere microbiome and plant health
1
2012
... 在土壤中, 根系分泌物和根际微生物之间的相互作用是一个十分重要的过程, 植物根系通过分泌各种次生代谢物质对根际微生物的种类、数量和分布产生影响, 对根际微生物群落结构有选择塑造作用.研究证据表明, 根系分泌物介导下的植物-微生物互作关系变化对于土壤肥力、健康状况, 以及植物生长发育有着极其重要的作用(Eisenhauer et al., 2012).有学者甚至认为, 根际是植物体的一个组成部分, 根际微生物群落的宏基因组是植物体的第二基因组(Berendsen et al., 2012).与此类似, Daffonchio认为不应仅仅将植物体看作是一个独立的生物体(single organism), 而应该把它看成宏生物体(meta-organism), 它是植物自身与周围微生物组(microbiome)的结合体(East, 2013).本研究团队多年来一直致力于植物根际生物学特性的研究, 发现植物的他感作用、连作障碍、连作促进或是间套作增产等, 都是由其根系分泌物介导下的植物与特异微生物共同作用的结果. ...
The pre-symbiotic growth of arbuscular mycorrhizal fungi is induced by a branching factor partially purified from plant root exudates
1
2000
... 真菌作为土壤微生物群落的主要组分之一, 在根际与植物形成密切关系, 尤其是丛枝菌根(arbuscular mycorrhizae, AM)能与大多数高等植物形成共生关系.丛枝菌根几乎在各种陆地生态系统中都有分布.许多研究表明菌根真菌和菌根植物之间存在互相感知的系统, 菌根植物的根系分泌物中存在能被菌根真菌识别的物质, 如来自Lotus japonicus的独脚金萌发素内酯(5-deoxystrigol)(Buee et al., 2000; Akiyama et al., 2005).在菌根真菌与植物的对话中, 植物产生的根系分泌物为菌根真菌的生长繁殖提供了丰富的碳源和能量, 而菌根真菌也能够帮助植物改善营养, 如通过溶解释放无机磷或水解有机磷来提高植物对磷元素的吸收利用, 抵御各种生物和非生物胁迫.然而也有研究发现, 植物与菌根真菌的关系也不是一成不变的, 在特定的环境下, 二者可能从互利共生变成寄生关系, 比如当为植物提供足够的化肥和有机肥时, 菌根真菌为植物提供磷元素的效益就可能无法补偿植物提供给菌根真菌碳源的成本, 从而引发二者关系的变化(Joh- nson et al., 1997).内生真菌(fungal endophyte)也与植物关系密切, 微生物学家Rusty J. Rodriguez甚至使用了一个新词“symbiogenics” (共生基因学)或称为“symbiosis-altered genetic expression” (共生改变基因表达)来形象地描述二者的关系.他的研究发现将分离自耐旱沙丘草本植物中的内生真菌与水稻共培养, 不但可以显著降低水稻对水分的需求量, 而且还可以促进水稻生长, 提高产量.同样, 将来自喜热稗的内生真菌与小麦共培养后, 小麦可在70 ℃高温下生长, 并且对水分的需求量减半(East, 2013). ...
Humic acids isolated from earthworm compost enhance root elongation, lateral root emergence, and plasma membrane H+-ATPase activity in maize roots
1
2002
... 土壤动物群的生长、分布与植物根系分泌物关系密切.有研究表明, 根际土中的原生动物数量明显高于非根际土中的, 这可能是由于根系分泌物促进了土壤细菌、真菌的生长繁殖, 从而有利于以细菌、真菌为食的原生动物的生长(Zwart et al., 1994).土壤动物区系除了通过食用植物残体碎屑, 捕食细菌、真菌, 改变土壤微生物区系, 改善土壤理化性质来加速土壤物质循环外, 很多土壤动物(如原生动物、蚯蚓等)还能分泌一些激素类有机物来促进植物生长(Canellas et al., 2002). ...
根分泌作用与植物对金属毒害的抗性
1
2000
... 植物可以通过产生和释放根系分泌物来抵御各种非生物因素的干扰与生物胁迫.在金属污染物胁迫下, 某些植物的根系分泌物通过螯合、络合、沉淀等作用将金属污染物滞留在根外, 降低土壤中金属的生物有效性, 减少植物对有害金属的吸收(常学秀等, 2000).例如, 在Al胁迫下, 一些高等植物可分泌大量的柠檬酸、苹果酸、酚类化合物以及黏液、蛋白质复合物等来螯合游离的Aln+阳离子, 从而降低Al对植物根系的毒害作用(Neumann, 2007).也有研究表明, 某些植物通过分泌苹果酸、柠檬酸等根系分泌物来缓解重碳酸盐毒害, 同时这些分泌物也有利于植物吸收利用P、Fe、Zn、Mn等矿质元素(Hajiboland et al., 2003).近年来还陆续发现, 某些植物可以通过分泌几丁质酶、β-1, 3-葡聚糖酶、植物抗毒素(phytoalexin)等生物活性物质来抑制病原菌的生长(Neumann et al., 2000; Werner, 2000; Wasaki et al., 2005). ...
根分泌作用与植物对金属毒害的抗性
1
2000
... 植物可以通过产生和释放根系分泌物来抵御各种非生物因素的干扰与生物胁迫.在金属污染物胁迫下, 某些植物的根系分泌物通过螯合、络合、沉淀等作用将金属污染物滞留在根外, 降低土壤中金属的生物有效性, 减少植物对有害金属的吸收(常学秀等, 2000).例如, 在Al胁迫下, 一些高等植物可分泌大量的柠檬酸、苹果酸、酚类化合物以及黏液、蛋白质复合物等来螯合游离的Aln+阳离子, 从而降低Al对植物根系的毒害作用(Neumann, 2007).也有研究表明, 某些植物通过分泌苹果酸、柠檬酸等根系分泌物来缓解重碳酸盐毒害, 同时这些分泌物也有利于植物吸收利用P、Fe、Zn、Mn等矿质元素(Hajiboland et al., 2003).近年来还陆续发现, 某些植物可以通过分泌几丁质酶、β-1, 3-葡聚糖酶、植物抗毒素(phytoalexin)等生物活性物质来抑制病原菌的生长(Neumann et al., 2000; Werner, 2000; Wasaki et al., 2005). ...
Root exudation of phytochem- icals in Arabidopsis follows specific patterns that are developmentally programmed and correlate with soil microbial functions.
1
2013
... 在根际土壤中, 根系分泌物在介导植物与周围微生物的相互作用中起重要作用.Gschwendtner等(2011)研究发现, 不同的马铃薯(Solanum tuberosum)品种之间根际微生物群落差异很大, 而且在不同生长发育时期, 马铃薯根际微生物群落结构也发生变化, 这可能都与根系分泌物的组成和数量不同有关.同样, Chaparro等(2013)发现拟南芥(Arabidopsis thaliana)根系分泌物的组分与含量随生长发育时期而显著变化, 根部相关基因的表达水平也相应变化, 同时对根际微生物群落进行宏转录组测序分析还发现, 根际微生物功能基因的表达模式与拟南芥不同生长发育时期的根系分泌物分泌模式显著相关. ...
连作对白肋烟根际土壤细菌群落多样性的影响
1
2010
... 近年来, 对于植物化感作用的研究也倾向于根际微生物群落结构, 因为目前对于化感物质的成分与有效浓度仍存在较大争议, 有的学者认为酚酸类化感物质抑制杂草生长作用的有效浓度较高, 在田间自然条件下很难达到有效浓度, 故提出酚酸类不是化感物质, 而萜类和黄酮类才是化感物质(Kato-Noguchi, 2009).但是现在更多的学者认为, 化感物质进入根际土壤之后势必会受到微生物的代谢分解、加工转化等, 所以植物化感作用并非是在供体植物与受体植物之间直接发挥作用(林文雄等, 2007; Kaur et al., 2009).显然, 作物化感作用是植物化感物质与其根际土壤微生物综合作用的结果.本实验室运用现代生理生化技术、土壤宏基因组学并结合实时荧光定量PCR (qRT-PCR)技术等研究发现, 化感水稻与非化感水稻根际土壤微生物群落结构存在明显差异, 化感水稻根际存在30种特异微生物, 其中7种被鉴定为黏细菌(myxobacteria)(林瑞余等, 2007; 林辉锋, 2011), 它们对杂草(如稗(Echino- chloa crusgali))的发芽生长具有显著抑制作用.进一步研究还发现, 这些与水稻化感作用相关的黏细菌具有群体效应(quorum sensing), 而且水稻根系分泌物中的酚酸类物质对这类细菌具有明显的趋化促进作用(李颖哲, 2013).连作障碍, 也称再植病或土壤病, 是一种特殊的植物化感现象, 是指同一地块连续多年种植相同作物造成作物生长状况变差, 产量和品质降低, 病虫害发生加剧的现象.前人研究表明, 造成作物连作障碍的原因可能是多方面的, 比如土壤营养失衡、土壤理化性质恶化、自毒物质积累等(张重义和林文雄, 2009).但是, 近年来越来越多的学者认为, 根系分泌物生态效应的间接作用及土壤微生物区系紊乱是导致植物连作障碍形成的主要因素(Qi et al., 2009), 这可能是由于在根系分泌物特定组分的介导下, 某些类群的微生物(如土传病原菌)大量繁殖, 同时抑制其他有益微生物(如假单胞菌等拮抗菌)的生长, 进而改变了植物根系分泌物的组分和数量, 为趋化性病原微生物提供更多的碳源、能源, 形成恶性循环, 造成植物生长发育不良.本研究团队通过对地黄、太子参(Pseudostellariae hetero- phylla)、甘蔗(Saccharum officinarum)、烟草等不同作物连作下的根际微生态特性进行研究, 也均发现类似结果(陈冬梅等, 2010; 张重义等, 2010; 林茂兹等, 2012).Zhou和Wu (2012)通过外源添加黄瓜自毒物质——香豆酸至土壤中, 也发现香豆酸对土壤微生物群落结构产生显著影响, 导致厚壁菌门、β变形菌门等细菌大量增加, 而使拟杆菌门、δ变形菌门、浮霉菌门等细菌显著减少, 同时还造成土壤中病原菌(如尖孢镰刀菌等)大量繁殖增长.可见, 根系分泌物介导下的土壤微生物群落结构与功能多样性变化对其他植物或植物自身的生长发育都产生重要的影响. ...
连作对白肋烟根际土壤细菌群落多样性的影响
1
2010
... 近年来, 对于植物化感作用的研究也倾向于根际微生物群落结构, 因为目前对于化感物质的成分与有效浓度仍存在较大争议, 有的学者认为酚酸类化感物质抑制杂草生长作用的有效浓度较高, 在田间自然条件下很难达到有效浓度, 故提出酚酸类不是化感物质, 而萜类和黄酮类才是化感物质(Kato-Noguchi, 2009).但是现在更多的学者认为, 化感物质进入根际土壤之后势必会受到微生物的代谢分解、加工转化等, 所以植物化感作用并非是在供体植物与受体植物之间直接发挥作用(林文雄等, 2007; Kaur et al., 2009).显然, 作物化感作用是植物化感物质与其根际土壤微生物综合作用的结果.本实验室运用现代生理生化技术、土壤宏基因组学并结合实时荧光定量PCR (qRT-PCR)技术等研究发现, 化感水稻与非化感水稻根际土壤微生物群落结构存在明显差异, 化感水稻根际存在30种特异微生物, 其中7种被鉴定为黏细菌(myxobacteria)(林瑞余等, 2007; 林辉锋, 2011), 它们对杂草(如稗(Echino- chloa crusgali))的发芽生长具有显著抑制作用.进一步研究还发现, 这些与水稻化感作用相关的黏细菌具有群体效应(quorum sensing), 而且水稻根系分泌物中的酚酸类物质对这类细菌具有明显的趋化促进作用(李颖哲, 2013).连作障碍, 也称再植病或土壤病, 是一种特殊的植物化感现象, 是指同一地块连续多年种植相同作物造成作物生长状况变差, 产量和品质降低, 病虫害发生加剧的现象.前人研究表明, 造成作物连作障碍的原因可能是多方面的, 比如土壤营养失衡、土壤理化性质恶化、自毒物质积累等(张重义和林文雄, 2009).但是, 近年来越来越多的学者认为, 根系分泌物生态效应的间接作用及土壤微生物区系紊乱是导致植物连作障碍形成的主要因素(Qi et al., 2009), 这可能是由于在根系分泌物特定组分的介导下, 某些类群的微生物(如土传病原菌)大量繁殖, 同时抑制其他有益微生物(如假单胞菌等拮抗菌)的生长, 进而改变了植物根系分泌物的组分和数量, 为趋化性病原微生物提供更多的碳源、能源, 形成恶性循环, 造成植物生长发育不良.本研究团队通过对地黄、太子参(Pseudostellariae hetero- phylla)、甘蔗(Saccharum officinarum)、烟草等不同作物连作下的根际微生态特性进行研究, 也均发现类似结果(陈冬梅等, 2010; 张重义等, 2010; 林茂兹等, 2012).Zhou和Wu (2012)通过外源添加黄瓜自毒物质——香豆酸至土壤中, 也发现香豆酸对土壤微生物群落结构产生显著影响, 导致厚壁菌门、β变形菌门等细菌大量增加, 而使拟杆菌门、δ变形菌门、浮霉菌门等细菌显著减少, 同时还造成土壤中病原菌(如尖孢镰刀菌等)大量繁殖增长.可见, 根系分泌物介导下的土壤微生物群落结构与功能多样性变化对其他植物或植物自身的生长发育都产生重要的影响. ...
Quorum-sensing quenching by rhizobacterial volatiles
1
2011
... 随着根系分泌物与根际微生物互馈研究的不断深入, 群体效应这一概念也得到广泛关注.群体效应是指细菌利用自身分泌的自诱导剂(autoind- ucer, AI)作为信号分子进行交流, 来控制群体生理行为的一个过程.革兰氏阴性菌、阳性菌都具有群体效应, 前者存在自诱导剂合成蛋白(luxI)/自诱导剂受体蛋白(luxR)介导的群体感应系统, 以N-酰基高丝氨酸内酯(AHL)为自诱导剂; 后者存在寡肽介导的群体感应系统, 以寡肽为自诱导剂(Walker et al., 2003).细菌不断繁殖的过程中, 当自诱导剂的浓度达到一定阈值时, 自诱导剂就会与受体蛋白结合, 从而启动目的基因的表达, 引起一系列的生理反应(如图1所示).在自然界中, 很多与植物生长相关的细菌都具有群体效应现象, 它们或使植物致病(如Pseudomonas syringae和Erwinia carotovora细菌), 或促进植物生长(如Pseudomonas putida和Pseudomonas aureofaciens细菌), 或抑制植物发病(如Pseudomonas fluorescens细菌).这些细菌能够通过自诱导剂介导的群体感应系统来了解自身的群体密度, 同时该感应系统还介导细菌抗生素的合成、致病因子的产生、生物膜的产生、质粒接合转移以及细胞的聚集移动等.既然植物病原菌能够通过群体效应系统来成功地侵害植物, 有学者开始寻找各种干扰群体效应的途径, 即群体效应猝灭(quorum- quenching), 试图抑制植物病原菌的生长, 从而控制植物病害.Chernin等(2011)研究发现, 一些植物促生菌(如荧光假单胞菌(Pseudomonas fluorescens)和普城沙雷氏菌(Serratia plymuthica))能够分泌一些挥发性化合物来猝灭植物病原菌相互交流的信号分子, 干扰其群体效应系统, 进而抑制病原菌正常的生理代谢和生长繁殖.所以, 群体效应猝灭逐渐被认为是控制植物病害的一种新策略(Ma et al., 2013). ...
Citric acid excretion and precipitation of calcium citrate in the rhizosphere of white lupin (Lupinus albus L).
1
1989
... 已有的很多研究表明, 根系分泌物介导植物的营养吸收, 尤其是在营养胁迫的条件下.Dinkelaker等(1989)报道, 在缺磷的土壤中, 白羽扇豆(Lupinus albus)首先在根形态上产生变化, 即形成“簇状根”, 而后分泌大量的柠檬酸, 而且柠檬酸的增加量随植株年龄和缺磷程度而改变.双子叶植物根系在缺铁时会分泌出柠檬酸、草酸和咖啡酸等, 通过对土壤中难溶性铁的螯合作用来增加铁的有效性(史刚荣, 2004).Takagi等(1984)研究发现, 禾本科植物缺铁时, 根系通过分泌一种对Fe3+具有极强络合能力的铁载体(phytosiderophore, PS)来提高对Fe的吸收.而且PS对Fe3+的络合不是专一的, 还可与其他的微量金属元素Cu、Zn、Mn、Co、Ni进行络合, 提高了根际环境中这些元素的生物有效性(刘文菊等, 2000).Li等(2007)研究发现, 禾本科植物与豆科植物间套作时, 豆科植物如蚕豆(Vicia faba)根系会通过释放有机酸和质子来酸化土壤, 活化土壤难溶性磷, 从而促进禾本科植物如玉米(Zea mays)对磷的吸收利用. ...
Microbiome: soil science comes to life
2
2013
... 在土壤中, 根系分泌物和根际微生物之间的相互作用是一个十分重要的过程, 植物根系通过分泌各种次生代谢物质对根际微生物的种类、数量和分布产生影响, 对根际微生物群落结构有选择塑造作用.研究证据表明, 根系分泌物介导下的植物-微生物互作关系变化对于土壤肥力、健康状况, 以及植物生长发育有着极其重要的作用(Eisenhauer et al., 2012).有学者甚至认为, 根际是植物体的一个组成部分, 根际微生物群落的宏基因组是植物体的第二基因组(Berendsen et al., 2012).与此类似, Daffonchio认为不应仅仅将植物体看作是一个独立的生物体(single organism), 而应该把它看成宏生物体(meta-organism), 它是植物自身与周围微生物组(microbiome)的结合体(East, 2013).本研究团队多年来一直致力于植物根际生物学特性的研究, 发现植物的他感作用、连作障碍、连作促进或是间套作增产等, 都是由其根系分泌物介导下的植物与特异微生物共同作用的结果. ...
... 真菌作为土壤微生物群落的主要组分之一, 在根际与植物形成密切关系, 尤其是丛枝菌根(arbuscular mycorrhizae, AM)能与大多数高等植物形成共生关系.丛枝菌根几乎在各种陆地生态系统中都有分布.许多研究表明菌根真菌和菌根植物之间存在互相感知的系统, 菌根植物的根系分泌物中存在能被菌根真菌识别的物质, 如来自Lotus japonicus的独脚金萌发素内酯(5-deoxystrigol)(Buee et al., 2000; Akiyama et al., 2005).在菌根真菌与植物的对话中, 植物产生的根系分泌物为菌根真菌的生长繁殖提供了丰富的碳源和能量, 而菌根真菌也能够帮助植物改善营养, 如通过溶解释放无机磷或水解有机磷来提高植物对磷元素的吸收利用, 抵御各种生物和非生物胁迫.然而也有研究发现, 植物与菌根真菌的关系也不是一成不变的, 在特定的环境下, 二者可能从互利共生变成寄生关系, 比如当为植物提供足够的化肥和有机肥时, 菌根真菌为植物提供磷元素的效益就可能无法补偿植物提供给菌根真菌碳源的成本, 从而引发二者关系的变化(Joh- nson et al., 1997).内生真菌(fungal endophyte)也与植物关系密切, 微生物学家Rusty J. Rodriguez甚至使用了一个新词“symbiogenics” (共生基因学)或称为“symbiosis-altered genetic expression” (共生改变基因表达)来形象地描述二者的关系.他的研究发现将分离自耐旱沙丘草本植物中的内生真菌与水稻共培养, 不但可以显著降低水稻对水分的需求量, 而且还可以促进水稻生长, 提高产量.同样, 将来自喜热稗的内生真菌与小麦共培养后, 小麦可在70 ℃高温下生长, 并且对水分的需求量减半(East, 2013). ...
Bacterial diversity stabilizes community productivity
2
2012
... 根际是指受植物根系活动影响, 物理、化学和生物学性质上不同于土体的那部分靠近植物根系的微域土区, 是植物-土壤生态系统物质交换十分频繁的一个界面, 也是土壤微生物非常活跃的一个区域.而根系分泌物是调节根际微生态系统活力与功能的关键因素.根系分泌物介导下的植物-土壤-微生物相互作用的研究正成为现代科学研究的一个热点问题.近年来, 随着现代生物化学与分子生物学研究手段的日益发展, 有关植物根际生物学特性的研究成果也日益丰富与深入.研究发现, 根系分泌物对根际微生物群落结构具有选择塑造作用, 不同植物体的根际微生物群落结构具有独特性与代表性(Paterson et al., 2007); 反之, 根际微生物群落结构变化也对植物根系分泌物释放、土壤物质循环、能量流动、信息传递有重要的影响, 进而影响植物的生长发育过程(Eisenhauer et al., 2012).深入探索根系分泌物介导下的植物-微生物复杂的互作关系, 对于合理有效地调控根际生态系统, 最大限度地发挥该系统的功能, 提高养分资源利用效率与作物生产力, 实现农业生产可持续发展, 具有重要的理论参考与实践指导意义. ...
... 在土壤中, 根系分泌物和根际微生物之间的相互作用是一个十分重要的过程, 植物根系通过分泌各种次生代谢物质对根际微生物的种类、数量和分布产生影响, 对根际微生物群落结构有选择塑造作用.研究证据表明, 根系分泌物介导下的植物-微生物互作关系变化对于土壤肥力、健康状况, 以及植物生长发育有着极其重要的作用(Eisenhauer et al., 2012).有学者甚至认为, 根际是植物体的一个组成部分, 根际微生物群落的宏基因组是植物体的第二基因组(Berendsen et al., 2012).与此类似, Daffonchio认为不应仅仅将植物体看作是一个独立的生物体(single organism), 而应该把它看成宏生物体(meta-organism), 它是植物自身与周围微生物组(microbiome)的结合体(East, 2013).本研究团队多年来一直致力于植物根际生物学特性的研究, 发现植物的他感作用、连作障碍、连作促进或是间套作增产等, 都是由其根系分泌物介导下的植物与特异微生物共同作用的结果. ...
Cross-biome metagenomic analyses of soil microbial communities and their functional attributes
1
2012
... 植物根系分泌物的产生是植物根部长期进化的结果, 是适应环境的一种体现, 不是一个完全被动的过程, 因为植物的任何代谢功能都直接或间接地消耗碳源和能量.由于根部一直处于土壤中, 无时无刻不与周围的生物、非生物因子做斗争, 所以植物能够根据周围环境的变化不断地调整自身根系分泌物的组成和数量来积极响应(Walker et al., 2003).在根际中, 根系分泌物对某些微生物具有吸引作用, 这类具有趋化性的细菌或真菌就能够在根际中大量聚集和繁殖, 比如豆科植物对根瘤菌的诱导; 而对有些病原菌却具有抑制作用, 比如小麦根系分泌物中的酚酸具有抗细菌的活性, 并且其抗菌活性具有协同作用(Wu et al., 2001).Fierer等(2012)运用宏基因组学技术对取自寒漠、热沙漠、森林、草原、苔原的16份土样的微生物群落结构进行分析, 发现来自沙漠区的土壤微生物群落明显区别于非沙漠区的, 进一步的宏基因组功能分析发现来自沙漠区的微生物类群中与营养循环、分解代谢植物源有机化合物相关的基因含量很低, 尤其是来自寒漠地区的微生物群落, 其种类多样性和功能多样性都最低, 可见地上部物种多样性水平显著影响地下部微生物群落结构与功能多样性.相反, 地下部微生物群落也会对地上部植物的生理代谢、生长发育等产生重要影响.Badri等(2013)运用气相色谱-质谱联用仪(GC-MS)分析发现, 土壤微生物群落对植物叶片的代谢物组分有显著影响, 进而影响昆虫对叶片的取食行为.Mendes等(2011)运用PhyloChip芯片技术对抑病型土壤(disease suppressive soil)和利病型土壤(disease conducive soil)中的微生物群落结构进行分析, 发现种植于抑病型土壤的甜菜(Beta vulgaris)其根际优势群落普遍为拮抗病原菌相关的微生物, 如放线菌门、β变形菌门、γ变形菌门等, 尤其是其中的假单胞菌在抑病型土壤中的数量极显著高于利病型土壤中, 这是由根系分泌物介导下植物与根际特异微生物共同作用的结果.Zolla等(2013)研究发现, 在适度干旱胁迫下将拟南芥幼苗种植于同域土壤(sympatric soil, 即长期种植拟南芥的土壤)中, 可以显著地促进植物生长、提高生物量, 而种植于非同域土壤(non-sympatric soil, 如长期种植松树和玉米的土壤)中对植物生物量没有影响.进一步研究还发现, 这是由于同域土壤中存在很多与降低植物干旱胁迫响应相关的促生根际菌(plant growth promoting rhizobacteria, PGPR), 如伯克氏菌属、芽孢杆菌属、不动杆菌属等.Lakshmanan等(2012)研究还发现, 番茄(Lycopersicon esculentum)叶片受到病原菌侵染后, 可通过调节根系分泌物组分与含量, 如增加根系苹果酸分泌释放量, 使更多的苹果酸进入根际, 从而招募更多的有益菌向根际聚集, 这些有益菌可进一步引发植物的诱导性系统抗性(induced systemic resistance)以对病原菌产生防御反应, 可见根系分泌物在植物-微生物互作关系中所扮演的重要角色. ...
Altering plant-microbe interaction through artificially manipulating bacterial quorum sensing
1
2002
... 根系分泌物介导的植物-微生物互作关系是处在不断进化之中的.Walker等(2004)将Ocimum basilicum幼苗与病原微生物铜绿假单胞菌在试管内共培养, 发现在病原菌作用下Ocimum basilicum根系分泌物中酚酸类物质(如迷迭香酸(rosmarinic acid, RA)这一抗菌物质)含量显著增加, 进一步研究还发现铜绿假单胞菌具有群体效应, 能够在抗菌成分RA还未积累到有效作用浓度时便迅速产生生物被膜(biofilm), 以抵御植物分泌的抗菌物质对其造成伤害, 进而继续侵染植物根系.相反, 也有学者研究发现, 某些植物如豌豆(Pisum sativum)、大豆、水稻、苜蓿属(Medicago)植物等通过长期的进化, 对病原微生物形成一种防御机制, 即能够通过产生AHL类似物、AHL信号阻断物或降解酶等途径来干扰某些病原细菌的群体效应, 抑制其生长或干扰其正常生理功能, 从而保护植物自身(Fray, 2002). ...
The rhizosphere. In: Gregory PJ ed. Plant Roots: Growth, Activity and Interactions with the Soil.
1
2007
... 有研究表明, 根系分泌物对土壤微团聚体的大小、分布、稳定性、吸附性能及亲水性等物理性质有显著影响(Materechera et al., 1992).根系分泌物还显著地影响根际土壤pH值, 主要的影响途径有: (1)植物对阳、阴离子吸收利用的不平衡性; (2)有机阴离子的释放; (3)根部呼吸; (4)根际微生物产酸(Gregory, 2007).也有研究表明, 根际土壤尤其是根尖土壤的阳离子交换量(CEC)显著增加(Oades, 1978), 主要原因可能是根系分泌的黏胶物质(如聚糖醛酸)含有大量羧基, 而羧基是很好的阳离子交换基团. ...
Effects of genetically modified starch metabolism in potato plants on photosynthate fluxes into the rhizosphere and on microbial degraders of root exudates
1
2011
... 在根际土壤中, 根系分泌物在介导植物与周围微生物的相互作用中起重要作用.Gschwendtner等(2011)研究发现, 不同的马铃薯(Solanum tuberosum)品种之间根际微生物群落差异很大, 而且在不同生长发育时期, 马铃薯根际微生物群落结构也发生变化, 这可能都与根系分泌物的组成和数量不同有关.同样, Chaparro等(2013)发现拟南芥(Arabidopsis thaliana)根系分泌物的组分与含量随生长发育时期而显著变化, 根部相关基因的表达水平也相应变化, 同时对根际微生物群落进行宏转录组测序分析还发现, 根际微生物功能基因的表达模式与拟南芥不同生长发育时期的根系分泌物分泌模式显著相关. ...
Effects of bicarbonate and high pH on growth of Zn-efficient and Zn-inefficient genotypes of rice, wheat and rye
1
2003
... 植物可以通过产生和释放根系分泌物来抵御各种非生物因素的干扰与生物胁迫.在金属污染物胁迫下, 某些植物的根系分泌物通过螯合、络合、沉淀等作用将金属污染物滞留在根外, 降低土壤中金属的生物有效性, 减少植物对有害金属的吸收(常学秀等, 2000).例如, 在Al胁迫下, 一些高等植物可分泌大量的柠檬酸、苹果酸、酚类化合物以及黏液、蛋白质复合物等来螯合游离的Aln+阳离子, 从而降低Al对植物根系的毒害作用(Neumann, 2007).也有研究表明, 某些植物通过分泌苹果酸、柠檬酸等根系分泌物来缓解重碳酸盐毒害, 同时这些分泌物也有利于植物吸收利用P、Fe、Zn、Mn等矿质元素(Hajiboland et al., 2003).近年来还陆续发现, 某些植物可以通过分泌几丁质酶、β-1, 3-葡聚糖酶、植物抗毒素(phytoalexin)等生物活性物质来抑制病原菌的生长(Neumann et al., 2000; Werner, 2000; Wasaki et al., 2005). ...
化感水稻P1312777苗期根系分泌物中化学成分分析
1
2005
... 植物根系分泌物中含有一类对邻近其他植物或自身产生抑制或促进作用的生物活性物质, 即为化感物质(allelochemicals).Tang和Young (1982)研究了Hemarthria altissima根系分泌物的化感作用, 发现供体植物根系分泌物中对受体植物生长呈抑制作用的主要是酚类化合物.在小麦(Triticum aestivum)、水稻(Oryza sativa)、玉米等作物的根系分泌物中也都检测到一些化感物质, 对农业杂草都具有抑制作用, 而且化感作用的强弱与品种有关(何海斌等, 2005).同时, 研究发现黄瓜(Cucumis sativus)、大豆(Glycine max)、烟草(Nicotiana tabacum)、地黄(Rehmannia glutinosa)、西洋参(Panax quinquefolius)等忌连作作物的连作障碍现象与根系分泌物中的化感物质密切相关.Yu和Matsui (1994)发现黄瓜根系分泌物中含有苯甲酸、对羟基苯甲酸、2, 5-二羟基苯甲酸、苯丙烯酸等11种酚酸物质, 其中有10种具有生物毒性.Pramanik等(2000)也从黄瓜根系分泌物中鉴定出苯甲酸及其衍生物、肉桂酸及其衍生物等, 并证明这些物质会阻碍黄瓜对养分的吸收.近年来的研究也表明, 根系分泌物对某些入侵植物的成功入侵起到决定性的化学生态学作用.Bais等(2003)研究发现, 矢车菊(Centaurea maculosa)根系分泌释放的儿茶素(+)-Catechin和(-)-Catechin对其成功入侵起重要作用. ...
化感水稻P1312777苗期根系分泌物中化学成分分析
1
2005
... 植物根系分泌物中含有一类对邻近其他植物或自身产生抑制或促进作用的生物活性物质, 即为化感物质(allelochemicals).Tang和Young (1982)研究了Hemarthria altissima根系分泌物的化感作用, 发现供体植物根系分泌物中对受体植物生长呈抑制作用的主要是酚类化合物.在小麦(Triticum aestivum)、水稻(Oryza sativa)、玉米等作物的根系分泌物中也都检测到一些化感物质, 对农业杂草都具有抑制作用, 而且化感作用的强弱与品种有关(何海斌等, 2005).同时, 研究发现黄瓜(Cucumis sativus)、大豆(Glycine max)、烟草(Nicotiana tabacum)、地黄(Rehmannia glutinosa)、西洋参(Panax quinquefolius)等忌连作作物的连作障碍现象与根系分泌物中的化感物质密切相关.Yu和Matsui (1994)发现黄瓜根系分泌物中含有苯甲酸、对羟基苯甲酸、2, 5-二羟基苯甲酸、苯丙烯酸等11种酚酸物质, 其中有10种具有生物毒性.Pramanik等(2000)也从黄瓜根系分泌物中鉴定出苯甲酸及其衍生物、肉桂酸及其衍生物等, 并证明这些物质会阻碍黄瓜对养分的吸收.近年来的研究也表明, 根系分泌物对某些入侵植物的成功入侵起到决定性的化学生态学作用.Bais等(2003)研究发现, 矢车菊(Centaurea maculosa)根系分泌释放的儿茶素(+)-Catechin和(-)-Catechin对其成功入侵起重要作用. ...
根系分泌物及其根际效应
2
2006
... 根系分泌物(root exudate)是指在特定环境下, 活的植物通过根系的不同部位释放到根际环境中的有机物质的总称, 是一种复杂的非均一体系.广义的根系分泌物包括: (1)渗出物(diffusate), 即由根部细胞通过被动形式扩散出来的一些低分子量有机物质; (2)分泌物(secretion), 即由根部细胞主动释放的一些具有一定生理功能的有机物质, 对营养元素迁移、植物解毒、信号传递、抵御胁迫等起重要作用; (3)排泄物(excretion), 即根部细胞生物代谢过程中产生的分解产物(Neumann & Römheld, 2000; Werner, 2000).狭义的根系分泌物仅包括植物通过溢泌作用进入土壤的可溶性物质(贺永华等, 2006).根系分泌物可以通过改变根际物理、化学或生物学特性来提高植物根系对营养元素的吸收利用和适应外界环境的变化, 同时它也是调控根际微生态功能的关键因子. ...
... 根系分泌物收集按培养介质的不同可以分为水培收集法、土培收集法、基质培(蛭石培、砂培等)收集法.根据原位检测与否, 又可分为扰动收集和原位收集(如放射性检测法).根系分泌物的鉴定方法主要有仪器分析法和生物测试法.仪器分析法, 如紫外-可见光谱仪(UV-VIS)、红外光谱仪(IR)、毛细管电泳仪(CK)、气相色谱仪(GC)、液相色谱仪(LC)、离子色谱仪(HPIC)、质谱仪(MS)和核磁共振仪(NMR)分析以及色谱质谱联用技术(GC-MS、LC-MS)等; 生物测试法是根据某些细菌、真菌和植物幼苗对分泌物中特定组分的敏感性进行定性分析.因此, 可根据根际特异微生物类群的生物量、活性或群落分布对根系分泌物进行定性或定量研究(贺永华等, 2006). ...
根系分泌物及其根际效应
2
2006
... 根系分泌物(root exudate)是指在特定环境下, 活的植物通过根系的不同部位释放到根际环境中的有机物质的总称, 是一种复杂的非均一体系.广义的根系分泌物包括: (1)渗出物(diffusate), 即由根部细胞通过被动形式扩散出来的一些低分子量有机物质; (2)分泌物(secretion), 即由根部细胞主动释放的一些具有一定生理功能的有机物质, 对营养元素迁移、植物解毒、信号传递、抵御胁迫等起重要作用; (3)排泄物(excretion), 即根部细胞生物代谢过程中产生的分解产物(Neumann & Römheld, 2000; Werner, 2000).狭义的根系分泌物仅包括植物通过溢泌作用进入土壤的可溶性物质(贺永华等, 2006).根系分泌物可以通过改变根际物理、化学或生物学特性来提高植物根系对营养元素的吸收利用和适应外界环境的变化, 同时它也是调控根际微生态功能的关键因子. ...
... 根系分泌物收集按培养介质的不同可以分为水培收集法、土培收集法、基质培(蛭石培、砂培等)收集法.根据原位检测与否, 又可分为扰动收集和原位收集(如放射性检测法).根系分泌物的鉴定方法主要有仪器分析法和生物测试法.仪器分析法, 如紫外-可见光谱仪(UV-VIS)、红外光谱仪(IR)、毛细管电泳仪(CK)、气相色谱仪(GC)、液相色谱仪(LC)、离子色谱仪(HPIC)、质谱仪(MS)和核磁共振仪(NMR)分析以及色谱质谱联用技术(GC-MS、LC-MS)等; 生物测试法是根据某些细菌、真菌和植物幼苗对分泌物中特定组分的敏感性进行定性分析.因此, 可根据根际特异微生物类群的生物量、活性或群落分布对根系分泌物进行定性或定量研究(贺永华等, 2006). ...
Functioning of mycorrhizal associations along the mutualism-parasitism continuum
1
1997
... 真菌作为土壤微生物群落的主要组分之一, 在根际与植物形成密切关系, 尤其是丛枝菌根(arbuscular mycorrhizae, AM)能与大多数高等植物形成共生关系.丛枝菌根几乎在各种陆地生态系统中都有分布.许多研究表明菌根真菌和菌根植物之间存在互相感知的系统, 菌根植物的根系分泌物中存在能被菌根真菌识别的物质, 如来自Lotus japonicus的独脚金萌发素内酯(5-deoxystrigol)(Buee et al., 2000; Akiyama et al., 2005).在菌根真菌与植物的对话中, 植物产生的根系分泌物为菌根真菌的生长繁殖提供了丰富的碳源和能量, 而菌根真菌也能够帮助植物改善营养, 如通过溶解释放无机磷或水解有机磷来提高植物对磷元素的吸收利用, 抵御各种生物和非生物胁迫.然而也有研究发现, 植物与菌根真菌的关系也不是一成不变的, 在特定的环境下, 二者可能从互利共生变成寄生关系, 比如当为植物提供足够的化肥和有机肥时, 菌根真菌为植物提供磷元素的效益就可能无法补偿植物提供给菌根真菌碳源的成本, 从而引发二者关系的变化(Joh- nson et al., 1997).内生真菌(fungal endophyte)也与植物关系密切, 微生物学家Rusty J. Rodriguez甚至使用了一个新词“symbiogenics” (共生基因学)或称为“symbiosis-altered genetic expression” (共生改变基因表达)来形象地描述二者的关系.他的研究发现将分离自耐旱沙丘草本植物中的内生真菌与水稻共培养, 不但可以显著降低水稻对水分的需求量, 而且还可以促进水稻生长, 提高产量.同样, 将来自喜热稗的内生真菌与小麦共培养后, 小麦可在70 ℃高温下生长, 并且对水分的需求量减半(East, 2013). ...
Stress-induced allelopathic activity and momilactone B in rice
1
2009
... 近年来, 对于植物化感作用的研究也倾向于根际微生物群落结构, 因为目前对于化感物质的成分与有效浓度仍存在较大争议, 有的学者认为酚酸类化感物质抑制杂草生长作用的有效浓度较高, 在田间自然条件下很难达到有效浓度, 故提出酚酸类不是化感物质, 而萜类和黄酮类才是化感物质(Kato-Noguchi, 2009).但是现在更多的学者认为, 化感物质进入根际土壤之后势必会受到微生物的代谢分解、加工转化等, 所以植物化感作用并非是在供体植物与受体植物之间直接发挥作用(林文雄等, 2007; Kaur et al., 2009).显然, 作物化感作用是植物化感物质与其根际土壤微生物综合作用的结果.本实验室运用现代生理生化技术、土壤宏基因组学并结合实时荧光定量PCR (qRT-PCR)技术等研究发现, 化感水稻与非化感水稻根际土壤微生物群落结构存在明显差异, 化感水稻根际存在30种特异微生物, 其中7种被鉴定为黏细菌(myxobacteria)(林瑞余等, 2007; 林辉锋, 2011), 它们对杂草(如稗(Echino- chloa crusgali))的发芽生长具有显著抑制作用.进一步研究还发现, 这些与水稻化感作用相关的黏细菌具有群体效应(quorum sensing), 而且水稻根系分泌物中的酚酸类物质对这类细菌具有明显的趋化促进作用(李颖哲, 2013).连作障碍, 也称再植病或土壤病, 是一种特殊的植物化感现象, 是指同一地块连续多年种植相同作物造成作物生长状况变差, 产量和品质降低, 病虫害发生加剧的现象.前人研究表明, 造成作物连作障碍的原因可能是多方面的, 比如土壤营养失衡、土壤理化性质恶化、自毒物质积累等(张重义和林文雄, 2009).但是, 近年来越来越多的学者认为, 根系分泌物生态效应的间接作用及土壤微生物区系紊乱是导致植物连作障碍形成的主要因素(Qi et al., 2009), 这可能是由于在根系分泌物特定组分的介导下, 某些类群的微生物(如土传病原菌)大量繁殖, 同时抑制其他有益微生物(如假单胞菌等拮抗菌)的生长, 进而改变了植物根系分泌物的组分和数量, 为趋化性病原微生物提供更多的碳源、能源, 形成恶性循环, 造成植物生长发育不良.本研究团队通过对地黄、太子参(Pseudostellariae hetero- phylla)、甘蔗(Saccharum officinarum)、烟草等不同作物连作下的根际微生态特性进行研究, 也均发现类似结果(陈冬梅等, 2010; 张重义等, 2010; 林茂兹等, 2012).Zhou和Wu (2012)通过外源添加黄瓜自毒物质——香豆酸至土壤中, 也发现香豆酸对土壤微生物群落结构产生显著影响, 导致厚壁菌门、β变形菌门等细菌大量增加, 而使拟杆菌门、δ变形菌门、浮霉菌门等细菌显著减少, 同时还造成土壤中病原菌(如尖孢镰刀菌等)大量繁殖增长.可见, 根系分泌物介导下的土壤微生物群落结构与功能多样性变化对其他植物或植物自身的生长发育都产生重要的影响. ...
Taking ecological function seriously: soil microbial communities can obviate allelopathic effects of released metabolites
1
2009
... 近年来, 对于植物化感作用的研究也倾向于根际微生物群落结构, 因为目前对于化感物质的成分与有效浓度仍存在较大争议, 有的学者认为酚酸类化感物质抑制杂草生长作用的有效浓度较高, 在田间自然条件下很难达到有效浓度, 故提出酚酸类不是化感物质, 而萜类和黄酮类才是化感物质(Kato-Noguchi, 2009).但是现在更多的学者认为, 化感物质进入根际土壤之后势必会受到微生物的代谢分解、加工转化等, 所以植物化感作用并非是在供体植物与受体植物之间直接发挥作用(林文雄等, 2007; Kaur et al., 2009).显然, 作物化感作用是植物化感物质与其根际土壤微生物综合作用的结果.本实验室运用现代生理生化技术、土壤宏基因组学并结合实时荧光定量PCR (qRT-PCR)技术等研究发现, 化感水稻与非化感水稻根际土壤微生物群落结构存在明显差异, 化感水稻根际存在30种特异微生物, 其中7种被鉴定为黏细菌(myxobacteria)(林瑞余等, 2007; 林辉锋, 2011), 它们对杂草(如稗(Echino- chloa crusgali))的发芽生长具有显著抑制作用.进一步研究还发现, 这些与水稻化感作用相关的黏细菌具有群体效应(quorum sensing), 而且水稻根系分泌物中的酚酸类物质对这类细菌具有明显的趋化促进作用(李颖哲, 2013).连作障碍, 也称再植病或土壤病, 是一种特殊的植物化感现象, 是指同一地块连续多年种植相同作物造成作物生长状况变差, 产量和品质降低, 病虫害发生加剧的现象.前人研究表明, 造成作物连作障碍的原因可能是多方面的, 比如土壤营养失衡、土壤理化性质恶化、自毒物质积累等(张重义和林文雄, 2009).但是, 近年来越来越多的学者认为, 根系分泌物生态效应的间接作用及土壤微生物区系紊乱是导致植物连作障碍形成的主要因素(Qi et al., 2009), 这可能是由于在根系分泌物特定组分的介导下, 某些类群的微生物(如土传病原菌)大量繁殖, 同时抑制其他有益微生物(如假单胞菌等拮抗菌)的生长, 进而改变了植物根系分泌物的组分和数量, 为趋化性病原微生物提供更多的碳源、能源, 形成恶性循环, 造成植物生长发育不良.本研究团队通过对地黄、太子参(Pseudostellariae hetero- phylla)、甘蔗(Saccharum officinarum)、烟草等不同作物连作下的根际微生态特性进行研究, 也均发现类似结果(陈冬梅等, 2010; 张重义等, 2010; 林茂兹等, 2012).Zhou和Wu (2012)通过外源添加黄瓜自毒物质——香豆酸至土壤中, 也发现香豆酸对土壤微生物群落结构产生显著影响, 导致厚壁菌门、β变形菌门等细菌大量增加, 而使拟杆菌门、δ变形菌门、浮霉菌门等细菌显著减少, 同时还造成土壤中病原菌(如尖孢镰刀菌等)大量繁殖增长.可见, 根系分泌物介导下的土壤微生物群落结构与功能多样性变化对其他植物或植物自身的生长发育都产生重要的影响. ...
Carbon input by plants into the soil
1
2000
... 高等植物光合作用固定的碳20%-60%被转移到植物地下部分, 其中释放到土壤中的碳多年生植物最高可达其转移量的70%, 一年生植物最高可达其转移量的40% (Kuzyakov & Domanski, 2000).在 “根际”这一特殊的生态环境中, 植物根系不断地向土壤中分泌大量的有机物质, 形成根际沉积, 这为根际微生物提供了丰富的营养和能源.根系分泌物的种类和数量决定了根际微生物的种类和数量, 并对微生物的生长繁殖及代谢过程产生影响(Singh & Mukerji, 2006); 而根际微生物群落的动态变化反过来也影响着根际生态系统的物质循环和能量流动, 从而影响植物的生长发育及植被多样性变化(Bar- dgett, 2005a).由此可见, 植物和土壤、微生物之间的物质交换、养分循环是一个复杂的开放系统. ...
Microbe-associated molecular patterns-triggered root responses mediate beneficial rhizobacterial recruitment in Arabidopsis.
1
2012
... 植物根系分泌物的产生是植物根部长期进化的结果, 是适应环境的一种体现, 不是一个完全被动的过程, 因为植物的任何代谢功能都直接或间接地消耗碳源和能量.由于根部一直处于土壤中, 无时无刻不与周围的生物、非生物因子做斗争, 所以植物能够根据周围环境的变化不断地调整自身根系分泌物的组成和数量来积极响应(Walker et al., 2003).在根际中, 根系分泌物对某些微生物具有吸引作用, 这类具有趋化性的细菌或真菌就能够在根际中大量聚集和繁殖, 比如豆科植物对根瘤菌的诱导; 而对有些病原菌却具有抑制作用, 比如小麦根系分泌物中的酚酸具有抗细菌的活性, 并且其抗菌活性具有协同作用(Wu et al., 2001).Fierer等(2012)运用宏基因组学技术对取自寒漠、热沙漠、森林、草原、苔原的16份土样的微生物群落结构进行分析, 发现来自沙漠区的土壤微生物群落明显区别于非沙漠区的, 进一步的宏基因组功能分析发现来自沙漠区的微生物类群中与营养循环、分解代谢植物源有机化合物相关的基因含量很低, 尤其是来自寒漠地区的微生物群落, 其种类多样性和功能多样性都最低, 可见地上部物种多样性水平显著影响地下部微生物群落结构与功能多样性.相反, 地下部微生物群落也会对地上部植物的生理代谢、生长发育等产生重要影响.Badri等(2013)运用气相色谱-质谱联用仪(GC-MS)分析发现, 土壤微生物群落对植物叶片的代谢物组分有显著影响, 进而影响昆虫对叶片的取食行为.Mendes等(2011)运用PhyloChip芯片技术对抑病型土壤(disease suppressive soil)和利病型土壤(disease conducive soil)中的微生物群落结构进行分析, 发现种植于抑病型土壤的甜菜(Beta vulgaris)其根际优势群落普遍为拮抗病原菌相关的微生物, 如放线菌门、β变形菌门、γ变形菌门等, 尤其是其中的假单胞菌在抑病型土壤中的数量极显著高于利病型土壤中, 这是由根系分泌物介导下植物与根际特异微生物共同作用的结果.Zolla等(2013)研究发现, 在适度干旱胁迫下将拟南芥幼苗种植于同域土壤(sympatric soil, 即长期种植拟南芥的土壤)中, 可以显著地促进植物生长、提高生物量, 而种植于非同域土壤(non-sympatric soil, 如长期种植松树和玉米的土壤)中对植物生物量没有影响.进一步研究还发现, 这是由于同域土壤中存在很多与降低植物干旱胁迫响应相关的促生根际菌(plant growth promoting rhizobacteria, PGPR), 如伯克氏菌属、芽孢杆菌属、不动杆菌属等.Lakshmanan等(2012)研究还发现, 番茄(Lycopersicon esculentum)叶片受到病原菌侵染后, 可通过调节根系分泌物组分与含量, 如增加根系苹果酸分泌释放量, 使更多的苹果酸进入根际, 从而招募更多的有益菌向根际聚集, 这些有益菌可进一步引发植物的诱导性系统抗性(induced systemic resistance)以对病原菌产生防御反应, 可见根系分泌物在植物-微生物互作关系中所扮演的重要角色. ...
Genetic and genomic glimpses of the elusive arbuscular mycorrhizal fungi
1
2012
... 近年来, 根际生物学特性的研究不论是在研究方法与技术上, 还是在研究成果转化和生产实践指导上都取得了长足的进步.但是, 根际生物学特性的研究还需要从以下几方面进行突破: (1)根系分泌物原位收集、原位检测方法的改进与发展.目前对于根系分泌物的分析通常采用有机溶剂萃取并结合高效液相色谱法、气相质谱法等进行定性定量测定, 这些方法不但耗时, 而且无法实现田间实时动态原位检测.近年来, 生物传感器的问世在很大程度上促进了根系分泌物研究的发展, 如各种酚酸生物传感器的出现给农业科学与环境科学工作者提供了极大的便利(Soloducho & Cabaj, 2013).(2)植物响应根际生物胁迫的感知途径与机制.已知植物的免疫受体蛋白常分为模式识别受体(pattern recognition receptor)和效应子识别受体(effectors recognition receptor), 由这两类受体进行识别、激发的植物防卫反应分别称为病原物相关分子模式触发的免疫(PAMP-triggered immunity, PTI)和效应子触发的免疫(effector-triggered immunity, ETI).虽然PTI和ETI在植物天然免疫性中的作用已被认知, 但参与PTI及ETI信号转导的许多信号分子及蛋白组分仍未明确, 需要进一步确认.(3)植物与根际微生物“根际对话”的形式与信号分子.比如, 将来自耐旱或喜热植物中的内生真菌与其他植物进行共培, 可显著提高其他植物的耐旱性和耐热性, 但是内生真菌如何激活启动植物体内相关代谢途径以降低水分消耗和促进植物生长尚不清楚, 有待进一步研究.(4)微生物群体效应与植物生长发育的关系.在自然界中, 存在很多具有群体效应的有益或有害微生物, 如很多的植物病原菌都存在群体效应, 调控着病原菌在宿主根际的定殖过程及其对宿主的致病性, 通过深入研究这些植物病原菌的群体效应机制及其与宿主的互作过程, 可以帮助建立合理的农艺措施或通过转基因方法来控制这些具有群体效应的病原菌爆发成灾(von Bodman et al., 2003).当然, 通过对这些具有群体效应的有益菌的深入研究, 也可为农业生产服务, 本研究团队目前也在积极探索化感水稻根际中有关群体效应菌群(如黏细菌)在化感水稻抑制杂草过程中的作用, 但对于黏细菌与水稻根系分泌物的互作关系以及根系分泌物对黏细菌群体效应启动的研究还处于初步探索阶段.(5)微生物基因组学(microbial genomics)的运用.Lanfranco和Young (2012)通过对丛枝菌根转录组学的分析, 初步从基因水平上探索了丛枝菌根抑制宿主防御以及与宿主进行物质营养交换的过程.现在, 该项技术也被越来越多地运用于发现和鉴定介导植物-微生物二者互作关系的效应子(effector), 目前对于细菌释放的潜在效应子的研究呈指数增长, 而对于真菌和卵菌纲真菌效应子的研究还比较少(Ronald & Shirasu, 2012).(6)各种宏组学技术(如环境宏基因组学、宏转录组学、宏蛋白质组学、宏代谢组学)的进一步完善与突破, 以及它们在植物-微生物互作研究中的运用.如土壤蛋白质组学技术虽能够在蛋白质水平上为揭示土壤生态系统的生物化学过程提供依据, 但是它自身也存在一些不足之处, 特别是受到了技术的限制, 如土壤蛋白质提取纯化技术有待改善和环境微生物蛋白质库不完整.综上所述, 根际生物学特性研究还面临诸多的挑战, 需要多项技术的有机结合才能深入地揭示土壤这一“黑箱”所蕴含的奥秘. ...
β-Carboline alkaloids: mechanisms of phototoxicity to bacteria and insects
1
1988
... 当然, 土壤中也存在一些危害植物生长的有害土壤动物区系, 如植物病原线虫等, 但是某些植物能够通过释放特定的根系分泌物来抵御有害动物的威胁.Wu等(1999)通过体外发根-蚜虫共培试验发现, 在蚜虫的刺激下, 植物能够大量分泌一种植物抗毒素——聚乙烯.Bais等(2002b)发现, Oxalis tuberosa的根系分泌物中含有一种会发荧光的物质——β咔巴啉生物碱衍生物, 经鉴定, 为骆驼蓬碱(harmine)和骆驼蓬碱二氢(harmaline), 有抗菌杀虫作用.Larson等(1988)也发现, β咔巴啉生物碱衍生物对某些细菌、土壤动物具有光毒性. ...
Diversity enhances agricultural productivity via rhizosphere phosphorus facilitation on phosphorus-deficient soils
1
2007
... 已有的很多研究表明, 根系分泌物介导植物的营养吸收, 尤其是在营养胁迫的条件下.Dinkelaker等(1989)报道, 在缺磷的土壤中, 白羽扇豆(Lupinus albus)首先在根形态上产生变化, 即形成“簇状根”, 而后分泌大量的柠檬酸, 而且柠檬酸的增加量随植株年龄和缺磷程度而改变.双子叶植物根系在缺铁时会分泌出柠檬酸、草酸和咖啡酸等, 通过对土壤中难溶性铁的螯合作用来增加铁的有效性(史刚荣, 2004).Takagi等(1984)研究发现, 禾本科植物缺铁时, 根系通过分泌一种对Fe3+具有极强络合能力的铁载体(phytosiderophore, PS)来提高对Fe的吸收.而且PS对Fe3+的络合不是专一的, 还可与其他的微量金属元素Cu、Zn、Mn、Co、Ni进行络合, 提高了根际环境中这些元素的生物有效性(刘文菊等, 2000).Li等(2007)研究发现, 禾本科植物与豆科植物间套作时, 豆科植物如蚕豆(Vicia faba)根系会通过释放有机酸和质子来酸化土壤, 活化土壤难溶性磷, 从而促进禾本科植物如玉米(Zea mays)对磷的吸收利用. ...
水稻酚酸类化感物质的动态变化及其对根际特定微生物的影响
1
2013
... 近年来, 对于植物化感作用的研究也倾向于根际微生物群落结构, 因为目前对于化感物质的成分与有效浓度仍存在较大争议, 有的学者认为酚酸类化感物质抑制杂草生长作用的有效浓度较高, 在田间自然条件下很难达到有效浓度, 故提出酚酸类不是化感物质, 而萜类和黄酮类才是化感物质(Kato-Noguchi, 2009).但是现在更多的学者认为, 化感物质进入根际土壤之后势必会受到微生物的代谢分解、加工转化等, 所以植物化感作用并非是在供体植物与受体植物之间直接发挥作用(林文雄等, 2007; Kaur et al., 2009).显然, 作物化感作用是植物化感物质与其根际土壤微生物综合作用的结果.本实验室运用现代生理生化技术、土壤宏基因组学并结合实时荧光定量PCR (qRT-PCR)技术等研究发现, 化感水稻与非化感水稻根际土壤微生物群落结构存在明显差异, 化感水稻根际存在30种特异微生物, 其中7种被鉴定为黏细菌(myxobacteria)(林瑞余等, 2007; 林辉锋, 2011), 它们对杂草(如稗(Echino- chloa crusgali))的发芽生长具有显著抑制作用.进一步研究还发现, 这些与水稻化感作用相关的黏细菌具有群体效应(quorum sensing), 而且水稻根系分泌物中的酚酸类物质对这类细菌具有明显的趋化促进作用(李颖哲, 2013).连作障碍, 也称再植病或土壤病, 是一种特殊的植物化感现象, 是指同一地块连续多年种植相同作物造成作物生长状况变差, 产量和品质降低, 病虫害发生加剧的现象.前人研究表明, 造成作物连作障碍的原因可能是多方面的, 比如土壤营养失衡、土壤理化性质恶化、自毒物质积累等(张重义和林文雄, 2009).但是, 近年来越来越多的学者认为, 根系分泌物生态效应的间接作用及土壤微生物区系紊乱是导致植物连作障碍形成的主要因素(Qi et al., 2009), 这可能是由于在根系分泌物特定组分的介导下, 某些类群的微生物(如土传病原菌)大量繁殖, 同时抑制其他有益微生物(如假单胞菌等拮抗菌)的生长, 进而改变了植物根系分泌物的组分和数量, 为趋化性病原微生物提供更多的碳源、能源, 形成恶性循环, 造成植物生长发育不良.本研究团队通过对地黄、太子参(Pseudostellariae hetero- phylla)、甘蔗(Saccharum officinarum)、烟草等不同作物连作下的根际微生态特性进行研究, 也均发现类似结果(陈冬梅等, 2010; 张重义等, 2010; 林茂兹等, 2012).Zhou和Wu (2012)通过外源添加黄瓜自毒物质——香豆酸至土壤中, 也发现香豆酸对土壤微生物群落结构产生显著影响, 导致厚壁菌门、β变形菌门等细菌大量增加, 而使拟杆菌门、δ变形菌门、浮霉菌门等细菌显著减少, 同时还造成土壤中病原菌(如尖孢镰刀菌等)大量繁殖增长.可见, 根系分泌物介导下的土壤微生物群落结构与功能多样性变化对其他植物或植物自身的生长发育都产生重要的影响. ...
水稻酚酸类化感物质的动态变化及其对根际特定微生物的影响
1
2013
... 近年来, 对于植物化感作用的研究也倾向于根际微生物群落结构, 因为目前对于化感物质的成分与有效浓度仍存在较大争议, 有的学者认为酚酸类化感物质抑制杂草生长作用的有效浓度较高, 在田间自然条件下很难达到有效浓度, 故提出酚酸类不是化感物质, 而萜类和黄酮类才是化感物质(Kato-Noguchi, 2009).但是现在更多的学者认为, 化感物质进入根际土壤之后势必会受到微生物的代谢分解、加工转化等, 所以植物化感作用并非是在供体植物与受体植物之间直接发挥作用(林文雄等, 2007; Kaur et al., 2009).显然, 作物化感作用是植物化感物质与其根际土壤微生物综合作用的结果.本实验室运用现代生理生化技术、土壤宏基因组学并结合实时荧光定量PCR (qRT-PCR)技术等研究发现, 化感水稻与非化感水稻根际土壤微生物群落结构存在明显差异, 化感水稻根际存在30种特异微生物, 其中7种被鉴定为黏细菌(myxobacteria)(林瑞余等, 2007; 林辉锋, 2011), 它们对杂草(如稗(Echino- chloa crusgali))的发芽生长具有显著抑制作用.进一步研究还发现, 这些与水稻化感作用相关的黏细菌具有群体效应(quorum sensing), 而且水稻根系分泌物中的酚酸类物质对这类细菌具有明显的趋化促进作用(李颖哲, 2013).连作障碍, 也称再植病或土壤病, 是一种特殊的植物化感现象, 是指同一地块连续多年种植相同作物造成作物生长状况变差, 产量和品质降低, 病虫害发生加剧的现象.前人研究表明, 造成作物连作障碍的原因可能是多方面的, 比如土壤营养失衡、土壤理化性质恶化、自毒物质积累等(张重义和林文雄, 2009).但是, 近年来越来越多的学者认为, 根系分泌物生态效应的间接作用及土壤微生物区系紊乱是导致植物连作障碍形成的主要因素(Qi et al., 2009), 这可能是由于在根系分泌物特定组分的介导下, 某些类群的微生物(如土传病原菌)大量繁殖, 同时抑制其他有益微生物(如假单胞菌等拮抗菌)的生长, 进而改变了植物根系分泌物的组分和数量, 为趋化性病原微生物提供更多的碳源、能源, 形成恶性循环, 造成植物生长发育不良.本研究团队通过对地黄、太子参(Pseudostellariae hetero- phylla)、甘蔗(Saccharum officinarum)、烟草等不同作物连作下的根际微生态特性进行研究, 也均发现类似结果(陈冬梅等, 2010; 张重义等, 2010; 林茂兹等, 2012).Zhou和Wu (2012)通过外源添加黄瓜自毒物质——香豆酸至土壤中, 也发现香豆酸对土壤微生物群落结构产生显著影响, 导致厚壁菌门、β变形菌门等细菌大量增加, 而使拟杆菌门、δ变形菌门、浮霉菌门等细菌显著减少, 同时还造成土壤中病原菌(如尖孢镰刀菌等)大量繁殖增长.可见, 根系分泌物介导下的土壤微生物群落结构与功能多样性变化对其他植物或植物自身的生长发育都产生重要的影响. ...
不同水分条件下化感水稻根际功能微生物研究
1
2011
... 近年来, 对于植物化感作用的研究也倾向于根际微生物群落结构, 因为目前对于化感物质的成分与有效浓度仍存在较大争议, 有的学者认为酚酸类化感物质抑制杂草生长作用的有效浓度较高, 在田间自然条件下很难达到有效浓度, 故提出酚酸类不是化感物质, 而萜类和黄酮类才是化感物质(Kato-Noguchi, 2009).但是现在更多的学者认为, 化感物质进入根际土壤之后势必会受到微生物的代谢分解、加工转化等, 所以植物化感作用并非是在供体植物与受体植物之间直接发挥作用(林文雄等, 2007; Kaur et al., 2009).显然, 作物化感作用是植物化感物质与其根际土壤微生物综合作用的结果.本实验室运用现代生理生化技术、土壤宏基因组学并结合实时荧光定量PCR (qRT-PCR)技术等研究发现, 化感水稻与非化感水稻根际土壤微生物群落结构存在明显差异, 化感水稻根际存在30种特异微生物, 其中7种被鉴定为黏细菌(myxobacteria)(林瑞余等, 2007; 林辉锋, 2011), 它们对杂草(如稗(Echino- chloa crusgali))的发芽生长具有显著抑制作用.进一步研究还发现, 这些与水稻化感作用相关的黏细菌具有群体效应(quorum sensing), 而且水稻根系分泌物中的酚酸类物质对这类细菌具有明显的趋化促进作用(李颖哲, 2013).连作障碍, 也称再植病或土壤病, 是一种特殊的植物化感现象, 是指同一地块连续多年种植相同作物造成作物生长状况变差, 产量和品质降低, 病虫害发生加剧的现象.前人研究表明, 造成作物连作障碍的原因可能是多方面的, 比如土壤营养失衡、土壤理化性质恶化、自毒物质积累等(张重义和林文雄, 2009).但是, 近年来越来越多的学者认为, 根系分泌物生态效应的间接作用及土壤微生物区系紊乱是导致植物连作障碍形成的主要因素(Qi et al., 2009), 这可能是由于在根系分泌物特定组分的介导下, 某些类群的微生物(如土传病原菌)大量繁殖, 同时抑制其他有益微生物(如假单胞菌等拮抗菌)的生长, 进而改变了植物根系分泌物的组分和数量, 为趋化性病原微生物提供更多的碳源、能源, 形成恶性循环, 造成植物生长发育不良.本研究团队通过对地黄、太子参(Pseudostellariae hetero- phylla)、甘蔗(Saccharum officinarum)、烟草等不同作物连作下的根际微生态特性进行研究, 也均发现类似结果(陈冬梅等, 2010; 张重义等, 2010; 林茂兹等, 2012).Zhou和Wu (2012)通过外源添加黄瓜自毒物质——香豆酸至土壤中, 也发现香豆酸对土壤微生物群落结构产生显著影响, 导致厚壁菌门、β变形菌门等细菌大量增加, 而使拟杆菌门、δ变形菌门、浮霉菌门等细菌显著减少, 同时还造成土壤中病原菌(如尖孢镰刀菌等)大量繁殖增长.可见, 根系分泌物介导下的土壤微生物群落结构与功能多样性变化对其他植物或植物自身的生长发育都产生重要的影响. ...
不同水分条件下化感水稻根际功能微生物研究
1
2011
... 近年来, 对于植物化感作用的研究也倾向于根际微生物群落结构, 因为目前对于化感物质的成分与有效浓度仍存在较大争议, 有的学者认为酚酸类化感物质抑制杂草生长作用的有效浓度较高, 在田间自然条件下很难达到有效浓度, 故提出酚酸类不是化感物质, 而萜类和黄酮类才是化感物质(Kato-Noguchi, 2009).但是现在更多的学者认为, 化感物质进入根际土壤之后势必会受到微生物的代谢分解、加工转化等, 所以植物化感作用并非是在供体植物与受体植物之间直接发挥作用(林文雄等, 2007; Kaur et al., 2009).显然, 作物化感作用是植物化感物质与其根际土壤微生物综合作用的结果.本实验室运用现代生理生化技术、土壤宏基因组学并结合实时荧光定量PCR (qRT-PCR)技术等研究发现, 化感水稻与非化感水稻根际土壤微生物群落结构存在明显差异, 化感水稻根际存在30种特异微生物, 其中7种被鉴定为黏细菌(myxobacteria)(林瑞余等, 2007; 林辉锋, 2011), 它们对杂草(如稗(Echino- chloa crusgali))的发芽生长具有显著抑制作用.进一步研究还发现, 这些与水稻化感作用相关的黏细菌具有群体效应(quorum sensing), 而且水稻根系分泌物中的酚酸类物质对这类细菌具有明显的趋化促进作用(李颖哲, 2013).连作障碍, 也称再植病或土壤病, 是一种特殊的植物化感现象, 是指同一地块连续多年种植相同作物造成作物生长状况变差, 产量和品质降低, 病虫害发生加剧的现象.前人研究表明, 造成作物连作障碍的原因可能是多方面的, 比如土壤营养失衡、土壤理化性质恶化、自毒物质积累等(张重义和林文雄, 2009).但是, 近年来越来越多的学者认为, 根系分泌物生态效应的间接作用及土壤微生物区系紊乱是导致植物连作障碍形成的主要因素(Qi et al., 2009), 这可能是由于在根系分泌物特定组分的介导下, 某些类群的微生物(如土传病原菌)大量繁殖, 同时抑制其他有益微生物(如假单胞菌等拮抗菌)的生长, 进而改变了植物根系分泌物的组分和数量, 为趋化性病原微生物提供更多的碳源、能源, 形成恶性循环, 造成植物生长发育不良.本研究团队通过对地黄、太子参(Pseudostellariae hetero- phylla)、甘蔗(Saccharum officinarum)、烟草等不同作物连作下的根际微生态特性进行研究, 也均发现类似结果(陈冬梅等, 2010; 张重义等, 2010; 林茂兹等, 2012).Zhou和Wu (2012)通过外源添加黄瓜自毒物质——香豆酸至土壤中, 也发现香豆酸对土壤微生物群落结构产生显著影响, 导致厚壁菌门、β变形菌门等细菌大量增加, 而使拟杆菌门、δ变形菌门、浮霉菌门等细菌显著减少, 同时还造成土壤中病原菌(如尖孢镰刀菌等)大量繁殖增长.可见, 根系分泌物介导下的土壤微生物群落结构与功能多样性变化对其他植物或植物自身的生长发育都产生重要的影响. ...
太子参连作对根际土壤微生物的影响
1
2012
... 近年来, 对于植物化感作用的研究也倾向于根际微生物群落结构, 因为目前对于化感物质的成分与有效浓度仍存在较大争议, 有的学者认为酚酸类化感物质抑制杂草生长作用的有效浓度较高, 在田间自然条件下很难达到有效浓度, 故提出酚酸类不是化感物质, 而萜类和黄酮类才是化感物质(Kato-Noguchi, 2009).但是现在更多的学者认为, 化感物质进入根际土壤之后势必会受到微生物的代谢分解、加工转化等, 所以植物化感作用并非是在供体植物与受体植物之间直接发挥作用(林文雄等, 2007; Kaur et al., 2009).显然, 作物化感作用是植物化感物质与其根际土壤微生物综合作用的结果.本实验室运用现代生理生化技术、土壤宏基因组学并结合实时荧光定量PCR (qRT-PCR)技术等研究发现, 化感水稻与非化感水稻根际土壤微生物群落结构存在明显差异, 化感水稻根际存在30种特异微生物, 其中7种被鉴定为黏细菌(myxobacteria)(林瑞余等, 2007; 林辉锋, 2011), 它们对杂草(如稗(Echino- chloa crusgali))的发芽生长具有显著抑制作用.进一步研究还发现, 这些与水稻化感作用相关的黏细菌具有群体效应(quorum sensing), 而且水稻根系分泌物中的酚酸类物质对这类细菌具有明显的趋化促进作用(李颖哲, 2013).连作障碍, 也称再植病或土壤病, 是一种特殊的植物化感现象, 是指同一地块连续多年种植相同作物造成作物生长状况变差, 产量和品质降低, 病虫害发生加剧的现象.前人研究表明, 造成作物连作障碍的原因可能是多方面的, 比如土壤营养失衡、土壤理化性质恶化、自毒物质积累等(张重义和林文雄, 2009).但是, 近年来越来越多的学者认为, 根系分泌物生态效应的间接作用及土壤微生物区系紊乱是导致植物连作障碍形成的主要因素(Qi et al., 2009), 这可能是由于在根系分泌物特定组分的介导下, 某些类群的微生物(如土传病原菌)大量繁殖, 同时抑制其他有益微生物(如假单胞菌等拮抗菌)的生长, 进而改变了植物根系分泌物的组分和数量, 为趋化性病原微生物提供更多的碳源、能源, 形成恶性循环, 造成植物生长发育不良.本研究团队通过对地黄、太子参(Pseudostellariae hetero- phylla)、甘蔗(Saccharum officinarum)、烟草等不同作物连作下的根际微生态特性进行研究, 也均发现类似结果(陈冬梅等, 2010; 张重义等, 2010; 林茂兹等, 2012).Zhou和Wu (2012)通过外源添加黄瓜自毒物质——香豆酸至土壤中, 也发现香豆酸对土壤微生物群落结构产生显著影响, 导致厚壁菌门、β变形菌门等细菌大量增加, 而使拟杆菌门、δ变形菌门、浮霉菌门等细菌显著减少, 同时还造成土壤中病原菌(如尖孢镰刀菌等)大量繁殖增长.可见, 根系分泌物介导下的土壤微生物群落结构与功能多样性变化对其他植物或植物自身的生长发育都产生重要的影响. ...
太子参连作对根际土壤微生物的影响
1
2012
... 近年来, 对于植物化感作用的研究也倾向于根际微生物群落结构, 因为目前对于化感物质的成分与有效浓度仍存在较大争议, 有的学者认为酚酸类化感物质抑制杂草生长作用的有效浓度较高, 在田间自然条件下很难达到有效浓度, 故提出酚酸类不是化感物质, 而萜类和黄酮类才是化感物质(Kato-Noguchi, 2009).但是现在更多的学者认为, 化感物质进入根际土壤之后势必会受到微生物的代谢分解、加工转化等, 所以植物化感作用并非是在供体植物与受体植物之间直接发挥作用(林文雄等, 2007; Kaur et al., 2009).显然, 作物化感作用是植物化感物质与其根际土壤微生物综合作用的结果.本实验室运用现代生理生化技术、土壤宏基因组学并结合实时荧光定量PCR (qRT-PCR)技术等研究发现, 化感水稻与非化感水稻根际土壤微生物群落结构存在明显差异, 化感水稻根际存在30种特异微生物, 其中7种被鉴定为黏细菌(myxobacteria)(林瑞余等, 2007; 林辉锋, 2011), 它们对杂草(如稗(Echino- chloa crusgali))的发芽生长具有显著抑制作用.进一步研究还发现, 这些与水稻化感作用相关的黏细菌具有群体效应(quorum sensing), 而且水稻根系分泌物中的酚酸类物质对这类细菌具有明显的趋化促进作用(李颖哲, 2013).连作障碍, 也称再植病或土壤病, 是一种特殊的植物化感现象, 是指同一地块连续多年种植相同作物造成作物生长状况变差, 产量和品质降低, 病虫害发生加剧的现象.前人研究表明, 造成作物连作障碍的原因可能是多方面的, 比如土壤营养失衡、土壤理化性质恶化、自毒物质积累等(张重义和林文雄, 2009).但是, 近年来越来越多的学者认为, 根系分泌物生态效应的间接作用及土壤微生物区系紊乱是导致植物连作障碍形成的主要因素(Qi et al., 2009), 这可能是由于在根系分泌物特定组分的介导下, 某些类群的微生物(如土传病原菌)大量繁殖, 同时抑制其他有益微生物(如假单胞菌等拮抗菌)的生长, 进而改变了植物根系分泌物的组分和数量, 为趋化性病原微生物提供更多的碳源、能源, 形成恶性循环, 造成植物生长发育不良.本研究团队通过对地黄、太子参(Pseudostellariae hetero- phylla)、甘蔗(Saccharum officinarum)、烟草等不同作物连作下的根际微生态特性进行研究, 也均发现类似结果(陈冬梅等, 2010; 张重义等, 2010; 林茂兹等, 2012).Zhou和Wu (2012)通过外源添加黄瓜自毒物质——香豆酸至土壤中, 也发现香豆酸对土壤微生物群落结构产生显著影响, 导致厚壁菌门、β变形菌门等细菌大量增加, 而使拟杆菌门、δ变形菌门、浮霉菌门等细菌显著减少, 同时还造成土壤中病原菌(如尖孢镰刀菌等)大量繁殖增长.可见, 根系分泌物介导下的土壤微生物群落结构与功能多样性变化对其他植物或植物自身的生长发育都产生重要的影响. ...
苗期化感水稻对根际土壤微生物群落及其功能多样性的影响
1
2007
... 近年来, 对于植物化感作用的研究也倾向于根际微生物群落结构, 因为目前对于化感物质的成分与有效浓度仍存在较大争议, 有的学者认为酚酸类化感物质抑制杂草生长作用的有效浓度较高, 在田间自然条件下很难达到有效浓度, 故提出酚酸类不是化感物质, 而萜类和黄酮类才是化感物质(Kato-Noguchi, 2009).但是现在更多的学者认为, 化感物质进入根际土壤之后势必会受到微生物的代谢分解、加工转化等, 所以植物化感作用并非是在供体植物与受体植物之间直接发挥作用(林文雄等, 2007; Kaur et al., 2009).显然, 作物化感作用是植物化感物质与其根际土壤微生物综合作用的结果.本实验室运用现代生理生化技术、土壤宏基因组学并结合实时荧光定量PCR (qRT-PCR)技术等研究发现, 化感水稻与非化感水稻根际土壤微生物群落结构存在明显差异, 化感水稻根际存在30种特异微生物, 其中7种被鉴定为黏细菌(myxobacteria)(林瑞余等, 2007; 林辉锋, 2011), 它们对杂草(如稗(Echino- chloa crusgali))的发芽生长具有显著抑制作用.进一步研究还发现, 这些与水稻化感作用相关的黏细菌具有群体效应(quorum sensing), 而且水稻根系分泌物中的酚酸类物质对这类细菌具有明显的趋化促进作用(李颖哲, 2013).连作障碍, 也称再植病或土壤病, 是一种特殊的植物化感现象, 是指同一地块连续多年种植相同作物造成作物生长状况变差, 产量和品质降低, 病虫害发生加剧的现象.前人研究表明, 造成作物连作障碍的原因可能是多方面的, 比如土壤营养失衡、土壤理化性质恶化、自毒物质积累等(张重义和林文雄, 2009).但是, 近年来越来越多的学者认为, 根系分泌物生态效应的间接作用及土壤微生物区系紊乱是导致植物连作障碍形成的主要因素(Qi et al., 2009), 这可能是由于在根系分泌物特定组分的介导下, 某些类群的微生物(如土传病原菌)大量繁殖, 同时抑制其他有益微生物(如假单胞菌等拮抗菌)的生长, 进而改变了植物根系分泌物的组分和数量, 为趋化性病原微生物提供更多的碳源、能源, 形成恶性循环, 造成植物生长发育不良.本研究团队通过对地黄、太子参(Pseudostellariae hetero- phylla)、甘蔗(Saccharum officinarum)、烟草等不同作物连作下的根际微生态特性进行研究, 也均发现类似结果(陈冬梅等, 2010; 张重义等, 2010; 林茂兹等, 2012).Zhou和Wu (2012)通过外源添加黄瓜自毒物质——香豆酸至土壤中, 也发现香豆酸对土壤微生物群落结构产生显著影响, 导致厚壁菌门、β变形菌门等细菌大量增加, 而使拟杆菌门、δ变形菌门、浮霉菌门等细菌显著减少, 同时还造成土壤中病原菌(如尖孢镰刀菌等)大量繁殖增长.可见, 根系分泌物介导下的土壤微生物群落结构与功能多样性变化对其他植物或植物自身的生长发育都产生重要的影响. ...
苗期化感水稻对根际土壤微生物群落及其功能多样性的影响
1
2007
... 近年来, 对于植物化感作用的研究也倾向于根际微生物群落结构, 因为目前对于化感物质的成分与有效浓度仍存在较大争议, 有的学者认为酚酸类化感物质抑制杂草生长作用的有效浓度较高, 在田间自然条件下很难达到有效浓度, 故提出酚酸类不是化感物质, 而萜类和黄酮类才是化感物质(Kato-Noguchi, 2009).但是现在更多的学者认为, 化感物质进入根际土壤之后势必会受到微生物的代谢分解、加工转化等, 所以植物化感作用并非是在供体植物与受体植物之间直接发挥作用(林文雄等, 2007; Kaur et al., 2009).显然, 作物化感作用是植物化感物质与其根际土壤微生物综合作用的结果.本实验室运用现代生理生化技术、土壤宏基因组学并结合实时荧光定量PCR (qRT-PCR)技术等研究发现, 化感水稻与非化感水稻根际土壤微生物群落结构存在明显差异, 化感水稻根际存在30种特异微生物, 其中7种被鉴定为黏细菌(myxobacteria)(林瑞余等, 2007; 林辉锋, 2011), 它们对杂草(如稗(Echino- chloa crusgali))的发芽生长具有显著抑制作用.进一步研究还发现, 这些与水稻化感作用相关的黏细菌具有群体效应(quorum sensing), 而且水稻根系分泌物中的酚酸类物质对这类细菌具有明显的趋化促进作用(李颖哲, 2013).连作障碍, 也称再植病或土壤病, 是一种特殊的植物化感现象, 是指同一地块连续多年种植相同作物造成作物生长状况变差, 产量和品质降低, 病虫害发生加剧的现象.前人研究表明, 造成作物连作障碍的原因可能是多方面的, 比如土壤营养失衡、土壤理化性质恶化、自毒物质积累等(张重义和林文雄, 2009).但是, 近年来越来越多的学者认为, 根系分泌物生态效应的间接作用及土壤微生物区系紊乱是导致植物连作障碍形成的主要因素(Qi et al., 2009), 这可能是由于在根系分泌物特定组分的介导下, 某些类群的微生物(如土传病原菌)大量繁殖, 同时抑制其他有益微生物(如假单胞菌等拮抗菌)的生长, 进而改变了植物根系分泌物的组分和数量, 为趋化性病原微生物提供更多的碳源、能源, 形成恶性循环, 造成植物生长发育不良.本研究团队通过对地黄、太子参(Pseudostellariae hetero- phylla)、甘蔗(Saccharum officinarum)、烟草等不同作物连作下的根际微生态特性进行研究, 也均发现类似结果(陈冬梅等, 2010; 张重义等, 2010; 林茂兹等, 2012).Zhou和Wu (2012)通过外源添加黄瓜自毒物质——香豆酸至土壤中, 也发现香豆酸对土壤微生物群落结构产生显著影响, 导致厚壁菌门、β变形菌门等细菌大量增加, 而使拟杆菌门、δ变形菌门、浮霉菌门等细菌显著减少, 同时还造成土壤中病原菌(如尖孢镰刀菌等)大量繁殖增长.可见, 根系分泌物介导下的土壤微生物群落结构与功能多样性变化对其他植物或植物自身的生长发育都产生重要的影响. ...
Metaproteomic analysis of ratoon sugarcane rhizospheric soil
1
2013
... 近年来, 随着土壤蛋白质提取方法的不断优化, 土壤宏蛋白质的提取效率越来越高, 提取的纯度也越来越高, 足以进行土壤蛋白质双向电泳, 这就为土壤差异宏蛋白质组学研究奠定了基础.应用土壤宏蛋白质组学(soil metaproteomics)方法与技术研究植物根际生物学特性及其分子机理已成为当前科学研究的热点问题(Pierre-Alain et al., 2007; Wang et al., 2011).Wang等(2011)利用土壤蛋白质组学技术分析常见作物(水稻、太子参、地黄、甘蔗、烟草)根际生物学特性发现, 随机挑取的189个土壤蛋白质中, 有107个蛋白质来源于植物(56.61%), 有72个蛋白质来源于微生物(39.68%), 有10个蛋白质来源于动物(5.29%).该研究还通过T-RFLP技术对根际微生物群落结构进行分析, 结果发现, T-RFLP技术与土壤蛋白质组学技术既有相似性又有互补性, 二者都无法取代对方, 而是相互验证.Wu等(2011)进一步运用本实验优化建立的土壤宏蛋白质组学技术对连作下地黄根际土壤蛋白质表达谱变化进行研究, 发现地黄连作对来自植物、微生物的土壤蛋白质表达都产生显著影响, 发生差异表达的土壤蛋白质涉及能量、氨基酸、核酸代谢、次级代谢、胁迫防御、信号传递等功能, 它们在植物-微生物的“根际对话”中发挥着重要作用.同样, Lin等(2013)运用土壤宏蛋白质组学技术对新种植与宿根甘蔗根际土壤的宏蛋白质表达谱变化进行分析, 也发现了很多介导植物-微生物根际对话的关键土壤蛋白. ...
化感植物根际生物学特性研究现状与展望
1
2007
... 近年来, 对于植物化感作用的研究也倾向于根际微生物群落结构, 因为目前对于化感物质的成分与有效浓度仍存在较大争议, 有的学者认为酚酸类化感物质抑制杂草生长作用的有效浓度较高, 在田间自然条件下很难达到有效浓度, 故提出酚酸类不是化感物质, 而萜类和黄酮类才是化感物质(Kato-Noguchi, 2009).但是现在更多的学者认为, 化感物质进入根际土壤之后势必会受到微生物的代谢分解、加工转化等, 所以植物化感作用并非是在供体植物与受体植物之间直接发挥作用(林文雄等, 2007; Kaur et al., 2009).显然, 作物化感作用是植物化感物质与其根际土壤微生物综合作用的结果.本实验室运用现代生理生化技术、土壤宏基因组学并结合实时荧光定量PCR (qRT-PCR)技术等研究发现, 化感水稻与非化感水稻根际土壤微生物群落结构存在明显差异, 化感水稻根际存在30种特异微生物, 其中7种被鉴定为黏细菌(myxobacteria)(林瑞余等, 2007; 林辉锋, 2011), 它们对杂草(如稗(Echino- chloa crusgali))的发芽生长具有显著抑制作用.进一步研究还发现, 这些与水稻化感作用相关的黏细菌具有群体效应(quorum sensing), 而且水稻根系分泌物中的酚酸类物质对这类细菌具有明显的趋化促进作用(李颖哲, 2013).连作障碍, 也称再植病或土壤病, 是一种特殊的植物化感现象, 是指同一地块连续多年种植相同作物造成作物生长状况变差, 产量和品质降低, 病虫害发生加剧的现象.前人研究表明, 造成作物连作障碍的原因可能是多方面的, 比如土壤营养失衡、土壤理化性质恶化、自毒物质积累等(张重义和林文雄, 2009).但是, 近年来越来越多的学者认为, 根系分泌物生态效应的间接作用及土壤微生物区系紊乱是导致植物连作障碍形成的主要因素(Qi et al., 2009), 这可能是由于在根系分泌物特定组分的介导下, 某些类群的微生物(如土传病原菌)大量繁殖, 同时抑制其他有益微生物(如假单胞菌等拮抗菌)的生长, 进而改变了植物根系分泌物的组分和数量, 为趋化性病原微生物提供更多的碳源、能源, 形成恶性循环, 造成植物生长发育不良.本研究团队通过对地黄、太子参(Pseudostellariae hetero- phylla)、甘蔗(Saccharum officinarum)、烟草等不同作物连作下的根际微生态特性进行研究, 也均发现类似结果(陈冬梅等, 2010; 张重义等, 2010; 林茂兹等, 2012).Zhou和Wu (2012)通过外源添加黄瓜自毒物质——香豆酸至土壤中, 也发现香豆酸对土壤微生物群落结构产生显著影响, 导致厚壁菌门、β变形菌门等细菌大量增加, 而使拟杆菌门、δ变形菌门、浮霉菌门等细菌显著减少, 同时还造成土壤中病原菌(如尖孢镰刀菌等)大量繁殖增长.可见, 根系分泌物介导下的土壤微生物群落结构与功能多样性变化对其他植物或植物自身的生长发育都产生重要的影响. ...
化感植物根际生物学特性研究现状与展望
1
2007
... 近年来, 对于植物化感作用的研究也倾向于根际微生物群落结构, 因为目前对于化感物质的成分与有效浓度仍存在较大争议, 有的学者认为酚酸类化感物质抑制杂草生长作用的有效浓度较高, 在田间自然条件下很难达到有效浓度, 故提出酚酸类不是化感物质, 而萜类和黄酮类才是化感物质(Kato-Noguchi, 2009).但是现在更多的学者认为, 化感物质进入根际土壤之后势必会受到微生物的代谢分解、加工转化等, 所以植物化感作用并非是在供体植物与受体植物之间直接发挥作用(林文雄等, 2007; Kaur et al., 2009).显然, 作物化感作用是植物化感物质与其根际土壤微生物综合作用的结果.本实验室运用现代生理生化技术、土壤宏基因组学并结合实时荧光定量PCR (qRT-PCR)技术等研究发现, 化感水稻与非化感水稻根际土壤微生物群落结构存在明显差异, 化感水稻根际存在30种特异微生物, 其中7种被鉴定为黏细菌(myxobacteria)(林瑞余等, 2007; 林辉锋, 2011), 它们对杂草(如稗(Echino- chloa crusgali))的发芽生长具有显著抑制作用.进一步研究还发现, 这些与水稻化感作用相关的黏细菌具有群体效应(quorum sensing), 而且水稻根系分泌物中的酚酸类物质对这类细菌具有明显的趋化促进作用(李颖哲, 2013).连作障碍, 也称再植病或土壤病, 是一种特殊的植物化感现象, 是指同一地块连续多年种植相同作物造成作物生长状况变差, 产量和品质降低, 病虫害发生加剧的现象.前人研究表明, 造成作物连作障碍的原因可能是多方面的, 比如土壤营养失衡、土壤理化性质恶化、自毒物质积累等(张重义和林文雄, 2009).但是, 近年来越来越多的学者认为, 根系分泌物生态效应的间接作用及土壤微生物区系紊乱是导致植物连作障碍形成的主要因素(Qi et al., 2009), 这可能是由于在根系分泌物特定组分的介导下, 某些类群的微生物(如土传病原菌)大量繁殖, 同时抑制其他有益微生物(如假单胞菌等拮抗菌)的生长, 进而改变了植物根系分泌物的组分和数量, 为趋化性病原微生物提供更多的碳源、能源, 形成恶性循环, 造成植物生长发育不良.本研究团队通过对地黄、太子参(Pseudostellariae hetero- phylla)、甘蔗(Saccharum officinarum)、烟草等不同作物连作下的根际微生态特性进行研究, 也均发现类似结果(陈冬梅等, 2010; 张重义等, 2010; 林茂兹等, 2012).Zhou和Wu (2012)通过外源添加黄瓜自毒物质——香豆酸至土壤中, 也发现香豆酸对土壤微生物群落结构产生显著影响, 导致厚壁菌门、β变形菌门等细菌大量增加, 而使拟杆菌门、δ变形菌门、浮霉菌门等细菌显著减少, 同时还造成土壤中病原菌(如尖孢镰刀菌等)大量繁殖增长.可见, 根系分泌物介导下的土壤微生物群落结构与功能多样性变化对其他植物或植物自身的生长发育都产生重要的影响. ...
根分泌物对根际难溶性镉的活化作用及对水稻吸收、运输镉的影响
1
2000
... 已有的很多研究表明, 根系分泌物介导植物的营养吸收, 尤其是在营养胁迫的条件下.Dinkelaker等(1989)报道, 在缺磷的土壤中, 白羽扇豆(Lupinus albus)首先在根形态上产生变化, 即形成“簇状根”, 而后分泌大量的柠檬酸, 而且柠檬酸的增加量随植株年龄和缺磷程度而改变.双子叶植物根系在缺铁时会分泌出柠檬酸、草酸和咖啡酸等, 通过对土壤中难溶性铁的螯合作用来增加铁的有效性(史刚荣, 2004).Takagi等(1984)研究发现, 禾本科植物缺铁时, 根系通过分泌一种对Fe3+具有极强络合能力的铁载体(phytosiderophore, PS)来提高对Fe的吸收.而且PS对Fe3+的络合不是专一的, 还可与其他的微量金属元素Cu、Zn、Mn、Co、Ni进行络合, 提高了根际环境中这些元素的生物有效性(刘文菊等, 2000).Li等(2007)研究发现, 禾本科植物与豆科植物间套作时, 豆科植物如蚕豆(Vicia faba)根系会通过释放有机酸和质子来酸化土壤, 活化土壤难溶性磷, 从而促进禾本科植物如玉米(Zea mays)对磷的吸收利用. ...
根分泌物对根际难溶性镉的活化作用及对水稻吸收、运输镉的影响
1
2000
... 已有的很多研究表明, 根系分泌物介导植物的营养吸收, 尤其是在营养胁迫的条件下.Dinkelaker等(1989)报道, 在缺磷的土壤中, 白羽扇豆(Lupinus albus)首先在根形态上产生变化, 即形成“簇状根”, 而后分泌大量的柠檬酸, 而且柠檬酸的增加量随植株年龄和缺磷程度而改变.双子叶植物根系在缺铁时会分泌出柠檬酸、草酸和咖啡酸等, 通过对土壤中难溶性铁的螯合作用来增加铁的有效性(史刚荣, 2004).Takagi等(1984)研究发现, 禾本科植物缺铁时, 根系通过分泌一种对Fe3+具有极强络合能力的铁载体(phytosiderophore, PS)来提高对Fe的吸收.而且PS对Fe3+的络合不是专一的, 还可与其他的微量金属元素Cu、Zn、Mn、Co、Ni进行络合, 提高了根际环境中这些元素的生物有效性(刘文菊等, 2000).Li等(2007)研究发现, 禾本科植物与豆科植物间套作时, 豆科植物如蚕豆(Vicia faba)根系会通过释放有机酸和质子来酸化土壤, 活化土壤难溶性磷, 从而促进禾本科植物如玉米(Zea mays)对磷的吸收利用. ...
Quorum quenching in culturable phyllosphere bacteria from tobacco
1
2013
... 随着根系分泌物与根际微生物互馈研究的不断深入, 群体效应这一概念也得到广泛关注.群体效应是指细菌利用自身分泌的自诱导剂(autoind- ucer, AI)作为信号分子进行交流, 来控制群体生理行为的一个过程.革兰氏阴性菌、阳性菌都具有群体效应, 前者存在自诱导剂合成蛋白(luxI)/自诱导剂受体蛋白(luxR)介导的群体感应系统, 以N-酰基高丝氨酸内酯(AHL)为自诱导剂; 后者存在寡肽介导的群体感应系统, 以寡肽为自诱导剂(Walker et al., 2003).细菌不断繁殖的过程中, 当自诱导剂的浓度达到一定阈值时, 自诱导剂就会与受体蛋白结合, 从而启动目的基因的表达, 引起一系列的生理反应(如图1所示).在自然界中, 很多与植物生长相关的细菌都具有群体效应现象, 它们或使植物致病(如Pseudomonas syringae和Erwinia carotovora细菌), 或促进植物生长(如Pseudomonas putida和Pseudomonas aureofaciens细菌), 或抑制植物发病(如Pseudomonas fluorescens细菌).这些细菌能够通过自诱导剂介导的群体感应系统来了解自身的群体密度, 同时该感应系统还介导细菌抗生素的合成、致病因子的产生、生物膜的产生、质粒接合转移以及细胞的聚集移动等.既然植物病原菌能够通过群体效应系统来成功地侵害植物, 有学者开始寻找各种干扰群体效应的途径, 即群体效应猝灭(quorum- quenching), 试图抑制植物病原菌的生长, 从而控制植物病害.Chernin等(2011)研究发现, 一些植物促生菌(如荧光假单胞菌(Pseudomonas fluorescens)和普城沙雷氏菌(Serratia plymuthica))能够分泌一些挥发性化合物来猝灭植物病原菌相互交流的信号分子, 干扰其群体效应系统, 进而抑制病原菌正常的生理代谢和生长繁殖.所以, 群体效应猝灭逐渐被认为是控制植物病害的一种新策略(Ma et al., 2013). ...
1
1995
... 根系分泌物作为根际沉积的重要组成部分, 按分子量可分为高分子量与低分子量根系分泌物, 前者主要包括黏胶和胞外酶, 其中黏胶有聚多糖和多糖醛酸; 后者主要是低分子有机酸、糖、酚及各种氨基酸(包括非蛋白氨基酸, 如植物铁载体) (Marschner, 1995).根系分泌物按种类可分为糖类、氨基酸类、有机酸、酚酸类、脂肪酸、甾醇类、蛋白质、生长因子等(表1). ...
Formation of aggregates by plant roots in homogenised soils
1
1992
... 有研究表明, 根系分泌物对土壤微团聚体的大小、分布、稳定性、吸附性能及亲水性等物理性质有显著影响(Materechera et al., 1992).根系分泌物还显著地影响根际土壤pH值, 主要的影响途径有: (1)植物对阳、阴离子吸收利用的不平衡性; (2)有机阴离子的释放; (3)根部呼吸; (4)根际微生物产酸(Gregory, 2007).也有研究表明, 根际土壤尤其是根尖土壤的阳离子交换量(CEC)显著增加(Oades, 1978), 主要原因可能是根系分泌的黏胶物质(如聚糖醛酸)含有大量羧基, 而羧基是很好的阳离子交换基团. ...
Deciphering the rhizosphere microbiome for disease-suppressive bacteria
1
2011
... 植物根系分泌物的产生是植物根部长期进化的结果, 是适应环境的一种体现, 不是一个完全被动的过程, 因为植物的任何代谢功能都直接或间接地消耗碳源和能量.由于根部一直处于土壤中, 无时无刻不与周围的生物、非生物因子做斗争, 所以植物能够根据周围环境的变化不断地调整自身根系分泌物的组成和数量来积极响应(Walker et al., 2003).在根际中, 根系分泌物对某些微生物具有吸引作用, 这类具有趋化性的细菌或真菌就能够在根际中大量聚集和繁殖, 比如豆科植物对根瘤菌的诱导; 而对有些病原菌却具有抑制作用, 比如小麦根系分泌物中的酚酸具有抗细菌的活性, 并且其抗菌活性具有协同作用(Wu et al., 2001).Fierer等(2012)运用宏基因组学技术对取自寒漠、热沙漠、森林、草原、苔原的16份土样的微生物群落结构进行分析, 发现来自沙漠区的土壤微生物群落明显区别于非沙漠区的, 进一步的宏基因组功能分析发现来自沙漠区的微生物类群中与营养循环、分解代谢植物源有机化合物相关的基因含量很低, 尤其是来自寒漠地区的微生物群落, 其种类多样性和功能多样性都最低, 可见地上部物种多样性水平显著影响地下部微生物群落结构与功能多样性.相反, 地下部微生物群落也会对地上部植物的生理代谢、生长发育等产生重要影响.Badri等(2013)运用气相色谱-质谱联用仪(GC-MS)分析发现, 土壤微生物群落对植物叶片的代谢物组分有显著影响, 进而影响昆虫对叶片的取食行为.Mendes等(2011)运用PhyloChip芯片技术对抑病型土壤(disease suppressive soil)和利病型土壤(disease conducive soil)中的微生物群落结构进行分析, 发现种植于抑病型土壤的甜菜(Beta vulgaris)其根际优势群落普遍为拮抗病原菌相关的微生物, 如放线菌门、β变形菌门、γ变形菌门等, 尤其是其中的假单胞菌在抑病型土壤中的数量极显著高于利病型土壤中, 这是由根系分泌物介导下植物与根际特异微生物共同作用的结果.Zolla等(2013)研究发现, 在适度干旱胁迫下将拟南芥幼苗种植于同域土壤(sympatric soil, 即长期种植拟南芥的土壤)中, 可以显著地促进植物生长、提高生物量, 而种植于非同域土壤(non-sympatric soil, 如长期种植松树和玉米的土壤)中对植物生物量没有影响.进一步研究还发现, 这是由于同域土壤中存在很多与降低植物干旱胁迫响应相关的促生根际菌(plant growth promoting rhizobacteria, PGPR), 如伯克氏菌属、芽孢杆菌属、不动杆菌属等.Lakshmanan等(2012)研究还发现, 番茄(Lycopersicon esculentum)叶片受到病原菌侵染后, 可通过调节根系分泌物组分与含量, 如增加根系苹果酸分泌释放量, 使更多的苹果酸进入根际, 从而招募更多的有益菌向根际聚集, 这些有益菌可进一步引发植物的诱导性系统抗性(induced systemic resistance)以对病原菌产生防御反应, 可见根系分泌物在植物-微生物互作关系中所扮演的重要角色. ...
Root exudates and nutrient cycling
2
2011
... Reported functions of plant root exudates
Table 2 | 功能 Function (Neumann, 2007) | 组成 Compound |
| 营养功能 Nutritional function | |
为微生物生长提供碳、氮源
Available sources of carbon and nitrogen for microbes | 低分子量糖类、氨基酸、羧酸类
Low molecular weight sugar, amino acid, carboxylic acid |
| 植物营养吸收 Plant nutrient acquisition | |
通过金属螯合提高对P、Fe、Zn、Mn的吸收
Mobilization of P, Fe, Zn and Mn, by metal chelation | 柠檬酸、草酸、苹果酸、酒石酸、植物铁载体
Citrate, oxalate, malate, malonate, tartaric acid, phytosiderophore |
通过还原作用提高对Fe、Mn的吸收
Mobilization of Fe and Mn, by contact reduction | 酚类、苹果酸、柠檬酸
Phenolic, malate, citrate |
回收利用有机磷酯类中的P元素
Mobilization and retrieval of P from organic P esters | 根细胞分泌磷酸水解酶
Root-secretory phosphohydrolase |
吸引固氮菌
Chemoattraction of N2-fixing microbes | 类黄酮、苹果酸、植物黏液
Flavonoid, malate, mucilage |
诱导共生固氮菌表达Nod基因和抗性相关基因以抵御植物抗毒素
Nod-gene inducers and resistance-inducers against phytoalexins in symbiotic N2 fixation | 黄酮类、黄烷酮(衍生物)、异黄酮
Flavone, flavanone, isoflavone |
诱导形成菌根的信号物质
Signal functions in establishment of mycorrhizal associations | 类黄酮、糖、氨基酸
Flavonoid, sugar, amino acid |
为菌根真菌提高碳源
Available carbon source for mycorrhizal fungi | 转化酶
Invertase |
| 保护功能 Protective function | |
通过络合缓解铝毒
Detoxification of Al3+ by complexation | 柠檬酸、草酸、苹果酸、酚类、植物黏液、分泌蛋白
Citrate, oxalate, malate, phenolic, mucilage, secretory protein |
响应重碳酸盐毒胁迫
In response to bicarbonate toxicity | 苹果酸、柠檬酸
Malate, citrate |
保护根部分裂组织, 提高根部与土壤的接触, 提高土壤保水能力
Protection of the root meristem, improved root-soil contact, increased water-holding capacity | 植物黏液
Mucilage |
植物抗毒素, 抵御病原菌、寄生植物、竞争者
Phytoalexin, defense against of pathogens, parasites and competitors | 苯醌、氢化奎宁、皂角苷、几丁质酶、根边缘细胞
Quinone, hydroquinine, saponin, chitinase, root border cell |
2.1 根系分泌物与土壤物质循环 高等植物光合作用固定的碳20%-60%被转移到植物地下部分, 其中释放到土壤中的碳多年生植物最高可达其转移量的70%, 一年生植物最高可达其转移量的40% (Kuzyakov & Domanski, 2000).在 “根际”这一特殊的生态环境中, 植物根系不断地向土壤中分泌大量的有机物质, 形成根际沉积, 这为根际微生物提供了丰富的营养和能源.根系分泌物的种类和数量决定了根际微生物的种类和数量, 并对微生物的生长繁殖及代谢过程产生影响(Singh & Mukerji, 2006); 而根际微生物群落的动态变化反过来也影响着根际生态系统的物质循环和能量流动, 从而影响植物的生长发育及植被多样性变化(Bar- dgett, 2005a).由此可见, 植物和土壤、微生物之间的物质交换、养分循环是一个复杂的开放系统. ...
... 植物可以通过产生和释放根系分泌物来抵御各种非生物因素的干扰与生物胁迫.在金属污染物胁迫下, 某些植物的根系分泌物通过螯合、络合、沉淀等作用将金属污染物滞留在根外, 降低土壤中金属的生物有效性, 减少植物对有害金属的吸收(常学秀等, 2000).例如, 在Al胁迫下, 一些高等植物可分泌大量的柠檬酸、苹果酸、酚类化合物以及黏液、蛋白质复合物等来螯合游离的Aln+阳离子, 从而降低Al对植物根系的毒害作用(Neumann, 2007).也有研究表明, 某些植物通过分泌苹果酸、柠檬酸等根系分泌物来缓解重碳酸盐毒害, 同时这些分泌物也有利于植物吸收利用P、Fe、Zn、Mn等矿质元素(Hajiboland et al., 2003).近年来还陆续发现, 某些植物可以通过分泌几丁质酶、β-1, 3-葡聚糖酶、植物抗毒素(phytoalexin)等生物活性物质来抑制病原菌的生长(Neumann et al., 2000; Werner, 2000; Wasaki et al., 2005). ...
Physiological aspects of cluster root function and development in phosphorus-deficient white lupin (Lupinus albus L.).
1
2000
... 植物可以通过产生和释放根系分泌物来抵御各种非生物因素的干扰与生物胁迫.在金属污染物胁迫下, 某些植物的根系分泌物通过螯合、络合、沉淀等作用将金属污染物滞留在根外, 降低土壤中金属的生物有效性, 减少植物对有害金属的吸收(常学秀等, 2000).例如, 在Al胁迫下, 一些高等植物可分泌大量的柠檬酸、苹果酸、酚类化合物以及黏液、蛋白质复合物等来螯合游离的Aln+阳离子, 从而降低Al对植物根系的毒害作用(Neumann, 2007).也有研究表明, 某些植物通过分泌苹果酸、柠檬酸等根系分泌物来缓解重碳酸盐毒害, 同时这些分泌物也有利于植物吸收利用P、Fe、Zn、Mn等矿质元素(Hajiboland et al., 2003).近年来还陆续发现, 某些植物可以通过分泌几丁质酶、β-1, 3-葡聚糖酶、植物抗毒素(phytoalexin)等生物活性物质来抑制病原菌的生长(Neumann et al., 2000; Werner, 2000; Wasaki et al., 2005). ...
The release of root exudates as affected by the plant physiological status
1
2000
... 根系分泌物(root exudate)是指在特定环境下, 活的植物通过根系的不同部位释放到根际环境中的有机物质的总称, 是一种复杂的非均一体系.广义的根系分泌物包括: (1)渗出物(diffusate), 即由根部细胞通过被动形式扩散出来的一些低分子量有机物质; (2)分泌物(secretion), 即由根部细胞主动释放的一些具有一定生理功能的有机物质, 对营养元素迁移、植物解毒、信号传递、抵御胁迫等起重要作用; (3)排泄物(excretion), 即根部细胞生物代谢过程中产生的分解产物(Neumann & Römheld, 2000; Werner, 2000).狭义的根系分泌物仅包括植物通过溢泌作用进入土壤的可溶性物质(贺永华等, 2006).根系分泌物可以通过改变根际物理、化学或生物学特性来提高植物根系对营养元素的吸收利用和适应外界环境的变化, 同时它也是调控根际微生态功能的关键因子. ...
Ribosome-inactivating proteins: a plant perspective
1
2001
... 由于土壤中不乏病原真菌, 植物也进化形成了各种抵御病原真菌的有效机制.Bais等(2002a)研究发现, 当受到终极腐霉菌Pythium ultimum侵害时, Ocimum basilicum根系会大量分泌迷迭香酸, 从而减轻该病菌的危害, 并杀灭多种土壤细菌和病原体, 显示出强大的抑菌活性.也有学者研究发现, Phytolacca americana的发根会向根际分泌许多抗真菌活性蛋白, 如核糖体失活蛋白(RIP)、几丁质酶等(Park et al., 2002a).RIP通过具N-糖苷酶活性来抑制蛋白合成, 或选择性地删除28S rRNA保守区的腺嘌呤残基(Nielsen & Boston, 2001; Vepachedu et al., 2003).根系分泌的RIP还具有抗真菌特性, 如能够抑制根腐菌Rhizoctonia solani和Trichoderma reesei (Park et al., 2002b). ...
Mucilages at the root surface
1
1978
... 有研究表明, 根系分泌物对土壤微团聚体的大小、分布、稳定性、吸附性能及亲水性等物理性质有显著影响(Materechera et al., 1992).根系分泌物还显著地影响根际土壤pH值, 主要的影响途径有: (1)植物对阳、阴离子吸收利用的不平衡性; (2)有机阴离子的释放; (3)根部呼吸; (4)根际微生物产酸(Gregory, 2007).也有研究表明, 根际土壤尤其是根尖土壤的阳离子交换量(CEC)显著增加(Oades, 1978), 主要原因可能是根系分泌的黏胶物质(如聚糖醛酸)含有大量羧基, 而羧基是很好的阳离子交换基团. ...
Iso- lation and characterization of a novel ribosome-inactivat- ing protein from root cultures of pokeweed and its mechanism of secretion from roots
1
2002a
... 由于土壤中不乏病原真菌, 植物也进化形成了各种抵御病原真菌的有效机制.Bais等(2002a)研究发现, 当受到终极腐霉菌Pythium ultimum侵害时, Ocimum basilicum根系会大量分泌迷迭香酸, 从而减轻该病菌的危害, 并杀灭多种土壤细菌和病原体, 显示出强大的抑菌活性.也有学者研究发现, Phytolacca americana的发根会向根际分泌许多抗真菌活性蛋白, 如核糖体失活蛋白(RIP)、几丁质酶等(Park et al., 2002a).RIP通过具N-糖苷酶活性来抑制蛋白合成, 或选择性地删除28S rRNA保守区的腺嘌呤残基(Nielsen & Boston, 2001; Vepachedu et al., 2003).根系分泌的RIP还具有抗真菌特性, 如能够抑制根腐菌Rhizoctonia solani和Trichoderma reesei (Park et al., 2002b). ...
Enzymatic specificity of three ribosome-inactivating proteins against fungal ribosomes, and correlation with antifungal activity
1
200b
... 由于土壤中不乏病原真菌, 植物也进化形成了各种抵御病原真菌的有效机制.Bais等(2002a)研究发现, 当受到终极腐霉菌Pythium ultimum侵害时, Ocimum basilicum根系会大量分泌迷迭香酸, 从而减轻该病菌的危害, 并杀灭多种土壤细菌和病原体, 显示出强大的抑菌活性.也有学者研究发现, Phytolacca americana的发根会向根际分泌许多抗真菌活性蛋白, 如核糖体失活蛋白(RIP)、几丁质酶等(Park et al., 2002a).RIP通过具N-糖苷酶活性来抑制蛋白合成, 或选择性地删除28S rRNA保守区的腺嘌呤残基(Nielsen & Boston, 2001; Vepachedu et al., 2003).根系分泌的RIP还具有抗真菌特性, 如能够抑制根腐菌Rhizoctonia solani和Trichoderma reesei (Park et al., 2002b). ...
Rhizodeposition shapes rhizosphere microbial community structure in organic soil
1
2007
... 根际是指受植物根系活动影响, 物理、化学和生物学性质上不同于土体的那部分靠近植物根系的微域土区, 是植物-土壤生态系统物质交换十分频繁的一个界面, 也是土壤微生物非常活跃的一个区域.而根系分泌物是调节根际微生态系统活力与功能的关键因素.根系分泌物介导下的植物-土壤-微生物相互作用的研究正成为现代科学研究的一个热点问题.近年来, 随着现代生物化学与分子生物学研究手段的日益发展, 有关植物根际生物学特性的研究成果也日益丰富与深入.研究发现, 根系分泌物对根际微生物群落结构具有选择塑造作用, 不同植物体的根际微生物群落结构具有独特性与代表性(Paterson et al., 2007); 反之, 根际微生物群落结构变化也对植物根系分泌物释放、土壤物质循环、能量流动、信息传递有重要的影响, 进而影响植物的生长发育过程(Eisenhauer et al., 2012).深入探索根系分泌物介导下的植物-微生物复杂的互作关系, 对于合理有效地调控根际生态系统, 最大限度地发挥该系统的功能, 提高养分资源利用效率与作物生产力, 实现农业生产可持续发展, 具有重要的理论参考与实践指导意义. ...
A plant flavone, luteolin, induces expression of Rhizobium meliloti nodulation genes.
1
1986
... 植物的根系分泌物中含有一些具有生物活性的大分子和小分子次生代谢产物, 这些物质调节着根系周围的土壤微生物种群与数量, 调节着植物与微生物之间的根际对话(rhizosphere talk).Peters等(1986)发现, 在缺氮条件下, 豆科植物根系通过分泌黄酮类和异黄酮类物质来诱导启动根瘤菌结瘤基因(nodD)的表达, 最终导致根瘤菌侵染根系并形成根瘤.Akiyama等(2005)研究发现, Lotus japonicus的根系分泌物中含有一种分枝因子(branching factor: 5-deoxystrigol), 在很低浓度时即可刺激菌根真菌萌发孢子的菌丝大量分枝. ...
Protein extraction and fingerprinting optimization of bacterial communities in natural environment
1
2007
... 近年来, 随着土壤蛋白质提取方法的不断优化, 土壤宏蛋白质的提取效率越来越高, 提取的纯度也越来越高, 足以进行土壤蛋白质双向电泳, 这就为土壤差异宏蛋白质组学研究奠定了基础.应用土壤宏蛋白质组学(soil metaproteomics)方法与技术研究植物根际生物学特性及其分子机理已成为当前科学研究的热点问题(Pierre-Alain et al., 2007; Wang et al., 2011).Wang等(2011)利用土壤蛋白质组学技术分析常见作物(水稻、太子参、地黄、甘蔗、烟草)根际生物学特性发现, 随机挑取的189个土壤蛋白质中, 有107个蛋白质来源于植物(56.61%), 有72个蛋白质来源于微生物(39.68%), 有10个蛋白质来源于动物(5.29%).该研究还通过T-RFLP技术对根际微生物群落结构进行分析, 结果发现, T-RFLP技术与土壤蛋白质组学技术既有相似性又有互补性, 二者都无法取代对方, 而是相互验证.Wu等(2011)进一步运用本实验优化建立的土壤宏蛋白质组学技术对连作下地黄根际土壤蛋白质表达谱变化进行研究, 发现地黄连作对来自植物、微生物的土壤蛋白质表达都产生显著影响, 发生差异表达的土壤蛋白质涉及能量、氨基酸、核酸代谢、次级代谢、胁迫防御、信号传递等功能, 它们在植物-微生物的“根际对话”中发挥着重要作用.同样, Lin等(2013)运用土壤宏蛋白质组学技术对新种植与宿根甘蔗根际土壤的宏蛋白质表达谱变化进行分析, 也发现了很多介导植物-微生物根际对话的关键土壤蛋白. ...
Effects of temperature and photoperiod on the phytotoxic root exudates of cucumber (Cucumis sativus) in hydroponic culture.
1
2000
... 植物根系分泌物中含有一类对邻近其他植物或自身产生抑制或促进作用的生物活性物质, 即为化感物质(allelochemicals).Tang和Young (1982)研究了Hemarthria altissima根系分泌物的化感作用, 发现供体植物根系分泌物中对受体植物生长呈抑制作用的主要是酚类化合物.在小麦(Triticum aestivum)、水稻(Oryza sativa)、玉米等作物的根系分泌物中也都检测到一些化感物质, 对农业杂草都具有抑制作用, 而且化感作用的强弱与品种有关(何海斌等, 2005).同时, 研究发现黄瓜(Cucumis sativus)、大豆(Glycine max)、烟草(Nicotiana tabacum)、地黄(Rehmannia glutinosa)、西洋参(Panax quinquefolius)等忌连作作物的连作障碍现象与根系分泌物中的化感物质密切相关.Yu和Matsui (1994)发现黄瓜根系分泌物中含有苯甲酸、对羟基苯甲酸、2, 5-二羟基苯甲酸、苯丙烯酸等11种酚酸物质, 其中有10种具有生物毒性.Pramanik等(2000)也从黄瓜根系分泌物中鉴定出苯甲酸及其衍生物、肉桂酸及其衍生物等, 并证明这些物质会阻碍黄瓜对养分的吸收.近年来的研究也表明, 根系分泌物对某些入侵植物的成功入侵起到决定性的化学生态学作用.Bais等(2003)研究发现, 矢车菊(Centaurea maculosa)根系分泌释放的儿茶素(+)-Catechin和(-)-Catechin对其成功入侵起重要作用. ...
Soil microbial community composition and diversity in the rhizosphere of a Chinese medicinal plant
1
2009
... 近年来, 对于植物化感作用的研究也倾向于根际微生物群落结构, 因为目前对于化感物质的成分与有效浓度仍存在较大争议, 有的学者认为酚酸类化感物质抑制杂草生长作用的有效浓度较高, 在田间自然条件下很难达到有效浓度, 故提出酚酸类不是化感物质, 而萜类和黄酮类才是化感物质(Kato-Noguchi, 2009).但是现在更多的学者认为, 化感物质进入根际土壤之后势必会受到微生物的代谢分解、加工转化等, 所以植物化感作用并非是在供体植物与受体植物之间直接发挥作用(林文雄等, 2007; Kaur et al., 2009).显然, 作物化感作用是植物化感物质与其根际土壤微生物综合作用的结果.本实验室运用现代生理生化技术、土壤宏基因组学并结合实时荧光定量PCR (qRT-PCR)技术等研究发现, 化感水稻与非化感水稻根际土壤微生物群落结构存在明显差异, 化感水稻根际存在30种特异微生物, 其中7种被鉴定为黏细菌(myxobacteria)(林瑞余等, 2007; 林辉锋, 2011), 它们对杂草(如稗(Echino- chloa crusgali))的发芽生长具有显著抑制作用.进一步研究还发现, 这些与水稻化感作用相关的黏细菌具有群体效应(quorum sensing), 而且水稻根系分泌物中的酚酸类物质对这类细菌具有明显的趋化促进作用(李颖哲, 2013).连作障碍, 也称再植病或土壤病, 是一种特殊的植物化感现象, 是指同一地块连续多年种植相同作物造成作物生长状况变差, 产量和品质降低, 病虫害发生加剧的现象.前人研究表明, 造成作物连作障碍的原因可能是多方面的, 比如土壤营养失衡、土壤理化性质恶化、自毒物质积累等(张重义和林文雄, 2009).但是, 近年来越来越多的学者认为, 根系分泌物生态效应的间接作用及土壤微生物区系紊乱是导致植物连作障碍形成的主要因素(Qi et al., 2009), 这可能是由于在根系分泌物特定组分的介导下, 某些类群的微生物(如土传病原菌)大量繁殖, 同时抑制其他有益微生物(如假单胞菌等拮抗菌)的生长, 进而改变了植物根系分泌物的组分和数量, 为趋化性病原微生物提供更多的碳源、能源, 形成恶性循环, 造成植物生长发育不良.本研究团队通过对地黄、太子参(Pseudostellariae hetero- phylla)、甘蔗(Saccharum officinarum)、烟草等不同作物连作下的根际微生态特性进行研究, 也均发现类似结果(陈冬梅等, 2010; 张重义等, 2010; 林茂兹等, 2012).Zhou和Wu (2012)通过外源添加黄瓜自毒物质——香豆酸至土壤中, 也发现香豆酸对土壤微生物群落结构产生显著影响, 导致厚壁菌门、β变形菌门等细菌大量增加, 而使拟杆菌门、δ变形菌门、浮霉菌门等细菌显著减少, 同时还造成土壤中病原菌(如尖孢镰刀菌等)大量繁殖增长.可见, 根系分泌物介导下的土壤微生物群落结构与功能多样性变化对其他植物或植物自身的生长发育都产生重要的影响. ...
Front-runners in plant-microbe interactions
1
2012
... 近年来, 根际生物学特性的研究不论是在研究方法与技术上, 还是在研究成果转化和生产实践指导上都取得了长足的进步.但是, 根际生物学特性的研究还需要从以下几方面进行突破: (1)根系分泌物原位收集、原位检测方法的改进与发展.目前对于根系分泌物的分析通常采用有机溶剂萃取并结合高效液相色谱法、气相质谱法等进行定性定量测定, 这些方法不但耗时, 而且无法实现田间实时动态原位检测.近年来, 生物传感器的问世在很大程度上促进了根系分泌物研究的发展, 如各种酚酸生物传感器的出现给农业科学与环境科学工作者提供了极大的便利(Soloducho & Cabaj, 2013).(2)植物响应根际生物胁迫的感知途径与机制.已知植物的免疫受体蛋白常分为模式识别受体(pattern recognition receptor)和效应子识别受体(effectors recognition receptor), 由这两类受体进行识别、激发的植物防卫反应分别称为病原物相关分子模式触发的免疫(PAMP-triggered immunity, PTI)和效应子触发的免疫(effector-triggered immunity, ETI).虽然PTI和ETI在植物天然免疫性中的作用已被认知, 但参与PTI及ETI信号转导的许多信号分子及蛋白组分仍未明确, 需要进一步确认.(3)植物与根际微生物“根际对话”的形式与信号分子.比如, 将来自耐旱或喜热植物中的内生真菌与其他植物进行共培, 可显著提高其他植物的耐旱性和耐热性, 但是内生真菌如何激活启动植物体内相关代谢途径以降低水分消耗和促进植物生长尚不清楚, 有待进一步研究.(4)微生物群体效应与植物生长发育的关系.在自然界中, 存在很多具有群体效应的有益或有害微生物, 如很多的植物病原菌都存在群体效应, 调控着病原菌在宿主根际的定殖过程及其对宿主的致病性, 通过深入研究这些植物病原菌的群体效应机制及其与宿主的互作过程, 可以帮助建立合理的农艺措施或通过转基因方法来控制这些具有群体效应的病原菌爆发成灾(von Bodman et al., 2003).当然, 通过对这些具有群体效应的有益菌的深入研究, 也可为农业生产服务, 本研究团队目前也在积极探索化感水稻根际中有关群体效应菌群(如黏细菌)在化感水稻抑制杂草过程中的作用, 但对于黏细菌与水稻根系分泌物的互作关系以及根系分泌物对黏细菌群体效应启动的研究还处于初步探索阶段.(5)微生物基因组学(microbial genomics)的运用.Lanfranco和Young (2012)通过对丛枝菌根转录组学的分析, 初步从基因水平上探索了丛枝菌根抑制宿主防御以及与宿主进行物质营养交换的过程.现在, 该项技术也被越来越多地运用于发现和鉴定介导植物-微生物二者互作关系的效应子(effector), 目前对于细菌释放的潜在效应子的研究呈指数增长, 而对于真菌和卵菌纲真菌效应子的研究还比较少(Ronald & Shirasu, 2012).(6)各种宏组学技术(如环境宏基因组学、宏转录组学、宏蛋白质组学、宏代谢组学)的进一步完善与突破, 以及它们在植物-微生物互作研究中的运用.如土壤蛋白质组学技术虽能够在蛋白质水平上为揭示土壤生态系统的生物化学过程提供依据, 但是它自身也存在一些不足之处, 特别是受到了技术的限制, 如土壤蛋白质提取纯化技术有待改善和环境微生物蛋白质库不完整.综上所述, 根际生物学特性研究还面临诸多的挑战, 需要多项技术的有机结合才能深入地揭示土壤这一“黑箱”所蕴含的奥秘. ...
植物根系分泌物的生态效应
1
2004
... 已有的很多研究表明, 根系分泌物介导植物的营养吸收, 尤其是在营养胁迫的条件下.Dinkelaker等(1989)报道, 在缺磷的土壤中, 白羽扇豆(Lupinus albus)首先在根形态上产生变化, 即形成“簇状根”, 而后分泌大量的柠檬酸, 而且柠檬酸的增加量随植株年龄和缺磷程度而改变.双子叶植物根系在缺铁时会分泌出柠檬酸、草酸和咖啡酸等, 通过对土壤中难溶性铁的螯合作用来增加铁的有效性(史刚荣, 2004).Takagi等(1984)研究发现, 禾本科植物缺铁时, 根系通过分泌一种对Fe3+具有极强络合能力的铁载体(phytosiderophore, PS)来提高对Fe的吸收.而且PS对Fe3+的络合不是专一的, 还可与其他的微量金属元素Cu、Zn、Mn、Co、Ni进行络合, 提高了根际环境中这些元素的生物有效性(刘文菊等, 2000).Li等(2007)研究发现, 禾本科植物与豆科植物间套作时, 豆科植物如蚕豆(Vicia faba)根系会通过释放有机酸和质子来酸化土壤, 活化土壤难溶性磷, 从而促进禾本科植物如玉米(Zea mays)对磷的吸收利用. ...
植物根系分泌物的生态效应
1
2004
... 已有的很多研究表明, 根系分泌物介导植物的营养吸收, 尤其是在营养胁迫的条件下.Dinkelaker等(1989)报道, 在缺磷的土壤中, 白羽扇豆(Lupinus albus)首先在根形态上产生变化, 即形成“簇状根”, 而后分泌大量的柠檬酸, 而且柠檬酸的增加量随植株年龄和缺磷程度而改变.双子叶植物根系在缺铁时会分泌出柠檬酸、草酸和咖啡酸等, 通过对土壤中难溶性铁的螯合作用来增加铁的有效性(史刚荣, 2004).Takagi等(1984)研究发现, 禾本科植物缺铁时, 根系通过分泌一种对Fe3+具有极强络合能力的铁载体(phytosiderophore, PS)来提高对Fe的吸收.而且PS对Fe3+的络合不是专一的, 还可与其他的微量金属元素Cu、Zn、Mn、Co、Ni进行络合, 提高了根际环境中这些元素的生物有效性(刘文菊等, 2000).Li等(2007)研究发现, 禾本科植物与豆科植物间套作时, 豆科植物如蚕豆(Vicia faba)根系会通过释放有机酸和质子来酸化土壤, 活化土壤难溶性磷, 从而促进禾本科植物如玉米(Zea mays)对磷的吸收利用. ...
Root exudates as determinant of rhizospheric microbial biodiversity
1
2006
... 高等植物光合作用固定的碳20%-60%被转移到植物地下部分, 其中释放到土壤中的碳多年生植物最高可达其转移量的70%, 一年生植物最高可达其转移量的40% (Kuzyakov & Domanski, 2000).在 “根际”这一特殊的生态环境中, 植物根系不断地向土壤中分泌大量的有机物质, 形成根际沉积, 这为根际微生物提供了丰富的营养和能源.根系分泌物的种类和数量决定了根际微生物的种类和数量, 并对微生物的生长繁殖及代谢过程产生影响(Singh & Mukerji, 2006); 而根际微生物群落的动态变化反过来也影响着根际生态系统的物质循环和能量流动, 从而影响植物的生长发育及植被多样性变化(Bar- dgett, 2005a).由此可见, 植物和土壤、微生物之间的物质交换、养分循环是一个复杂的开放系统. ...
Phenolic compounds hybrid de-tectors
1
2013
... 近年来, 根际生物学特性的研究不论是在研究方法与技术上, 还是在研究成果转化和生产实践指导上都取得了长足的进步.但是, 根际生物学特性的研究还需要从以下几方面进行突破: (1)根系分泌物原位收集、原位检测方法的改进与发展.目前对于根系分泌物的分析通常采用有机溶剂萃取并结合高效液相色谱法、气相质谱法等进行定性定量测定, 这些方法不但耗时, 而且无法实现田间实时动态原位检测.近年来, 生物传感器的问世在很大程度上促进了根系分泌物研究的发展, 如各种酚酸生物传感器的出现给农业科学与环境科学工作者提供了极大的便利(Soloducho & Cabaj, 2013).(2)植物响应根际生物胁迫的感知途径与机制.已知植物的免疫受体蛋白常分为模式识别受体(pattern recognition receptor)和效应子识别受体(effectors recognition receptor), 由这两类受体进行识别、激发的植物防卫反应分别称为病原物相关分子模式触发的免疫(PAMP-triggered immunity, PTI)和效应子触发的免疫(effector-triggered immunity, ETI).虽然PTI和ETI在植物天然免疫性中的作用已被认知, 但参与PTI及ETI信号转导的许多信号分子及蛋白组分仍未明确, 需要进一步确认.(3)植物与根际微生物“根际对话”的形式与信号分子.比如, 将来自耐旱或喜热植物中的内生真菌与其他植物进行共培, 可显著提高其他植物的耐旱性和耐热性, 但是内生真菌如何激活启动植物体内相关代谢途径以降低水分消耗和促进植物生长尚不清楚, 有待进一步研究.(4)微生物群体效应与植物生长发育的关系.在自然界中, 存在很多具有群体效应的有益或有害微生物, 如很多的植物病原菌都存在群体效应, 调控着病原菌在宿主根际的定殖过程及其对宿主的致病性, 通过深入研究这些植物病原菌的群体效应机制及其与宿主的互作过程, 可以帮助建立合理的农艺措施或通过转基因方法来控制这些具有群体效应的病原菌爆发成灾(von Bodman et al., 2003).当然, 通过对这些具有群体效应的有益菌的深入研究, 也可为农业生产服务, 本研究团队目前也在积极探索化感水稻根际中有关群体效应菌群(如黏细菌)在化感水稻抑制杂草过程中的作用, 但对于黏细菌与水稻根系分泌物的互作关系以及根系分泌物对黏细菌群体效应启动的研究还处于初步探索阶段.(5)微生物基因组学(microbial genomics)的运用.Lanfranco和Young (2012)通过对丛枝菌根转录组学的分析, 初步从基因水平上探索了丛枝菌根抑制宿主防御以及与宿主进行物质营养交换的过程.现在, 该项技术也被越来越多地运用于发现和鉴定介导植物-微生物二者互作关系的效应子(effector), 目前对于细菌释放的潜在效应子的研究呈指数增长, 而对于真菌和卵菌纲真菌效应子的研究还比较少(Ronald & Shirasu, 2012).(6)各种宏组学技术(如环境宏基因组学、宏转录组学、宏蛋白质组学、宏代谢组学)的进一步完善与突破, 以及它们在植物-微生物互作研究中的运用.如土壤蛋白质组学技术虽能够在蛋白质水平上为揭示土壤生态系统的生物化学过程提供依据, 但是它自身也存在一些不足之处, 特别是受到了技术的限制, 如土壤蛋白质提取纯化技术有待改善和环境微生物蛋白质库不完整.综上所述, 根际生物学特性研究还面临诸多的挑战, 需要多项技术的有机结合才能深入地揭示土壤这一“黑箱”所蕴含的奥秘. ...
Physiological aspect of mugineic acid, a possible phytosiderophore of gramin- aceous plants
1
1984
... 已有的很多研究表明, 根系分泌物介导植物的营养吸收, 尤其是在营养胁迫的条件下.Dinkelaker等(1989)报道, 在缺磷的土壤中, 白羽扇豆(Lupinus albus)首先在根形态上产生变化, 即形成“簇状根”, 而后分泌大量的柠檬酸, 而且柠檬酸的增加量随植株年龄和缺磷程度而改变.双子叶植物根系在缺铁时会分泌出柠檬酸、草酸和咖啡酸等, 通过对土壤中难溶性铁的螯合作用来增加铁的有效性(史刚荣, 2004).Takagi等(1984)研究发现, 禾本科植物缺铁时, 根系通过分泌一种对Fe3+具有极强络合能力的铁载体(phytosiderophore, PS)来提高对Fe的吸收.而且PS对Fe3+的络合不是专一的, 还可与其他的微量金属元素Cu、Zn、Mn、Co、Ni进行络合, 提高了根际环境中这些元素的生物有效性(刘文菊等, 2000).Li等(2007)研究发现, 禾本科植物与豆科植物间套作时, 豆科植物如蚕豆(Vicia faba)根系会通过释放有机酸和质子来酸化土壤, 活化土壤难溶性磷, 从而促进禾本科植物如玉米(Zea mays)对磷的吸收利用. ...
Collection and identification of allelopathic compounds from the undisturbed root system of Bigalta limpograss (Hemarthria altissima).
1
1982
... 植物根系分泌物中含有一类对邻近其他植物或自身产生抑制或促进作用的生物活性物质, 即为化感物质(allelochemicals).Tang和Young (1982)研究了Hemarthria altissima根系分泌物的化感作用, 发现供体植物根系分泌物中对受体植物生长呈抑制作用的主要是酚类化合物.在小麦(Triticum aestivum)、水稻(Oryza sativa)、玉米等作物的根系分泌物中也都检测到一些化感物质, 对农业杂草都具有抑制作用, 而且化感作用的强弱与品种有关(何海斌等, 2005).同时, 研究发现黄瓜(Cucumis sativus)、大豆(Glycine max)、烟草(Nicotiana tabacum)、地黄(Rehmannia glutinosa)、西洋参(Panax quinquefolius)等忌连作作物的连作障碍现象与根系分泌物中的化感物质密切相关.Yu和Matsui (1994)发现黄瓜根系分泌物中含有苯甲酸、对羟基苯甲酸、2, 5-二羟基苯甲酸、苯丙烯酸等11种酚酸物质, 其中有10种具有生物毒性.Pramanik等(2000)也从黄瓜根系分泌物中鉴定出苯甲酸及其衍生物、肉桂酸及其衍生物等, 并证明这些物质会阻碍黄瓜对养分的吸收.近年来的研究也表明, 根系分泌物对某些入侵植物的成功入侵起到决定性的化学生态学作用.Bais等(2003)研究发现, 矢车菊(Centaurea maculosa)根系分泌释放的儿茶素(+)-Catechin和(-)-Catechin对其成功入侵起重要作用. ...
植物根系分泌物与根际营养关系评述
1
2000
... 除了为根际土壤微生物系统提供碳源、氮源之外, 根系分泌物还介导植物对矿质元素的吸收利用和对外界环境变化的适应等.前人研究表明, 有些植物在养分和环境胁迫时, 根系分泌物的成分和数量会产生急剧变化以适应变化的环境, 这些植物一般都具有较高的养分利用效率和利用能力, 具有较强的抗逆性(涂书新等, 2000).根系分泌物种类众多, 功能包括: (1)营养供给; (2)改变土壤质地; (3)提高植物营养吸收; (4)信号传导; (5)化感作用; (6)保护功能等.表2列出了某些根系分泌物的生态效应. ...
植物根系分泌物与根际营养关系评述
1
2000
... 除了为根际土壤微生物系统提供碳源、氮源之外, 根系分泌物还介导植物对矿质元素的吸收利用和对外界环境变化的适应等.前人研究表明, 有些植物在养分和环境胁迫时, 根系分泌物的成分和数量会产生急剧变化以适应变化的环境, 这些植物一般都具有较高的养分利用效率和利用能力, 具有较强的抗逆性(涂书新等, 2000).根系分泌物种类众多, 功能包括: (1)营养供给; (2)改变土壤质地; (3)提高植物营养吸收; (4)信号传导; (5)化感作用; (6)保护功能等.表2列出了某些根系分泌物的生态效应. ...
Molecular characterization and post-transcriptional regulation of ME1, a type-I ribosome-inactivating protein from Mirabilis expansa.
1
2003
... 由于土壤中不乏病原真菌, 植物也进化形成了各种抵御病原真菌的有效机制.Bais等(2002a)研究发现, 当受到终极腐霉菌Pythium ultimum侵害时, Ocimum basilicum根系会大量分泌迷迭香酸, 从而减轻该病菌的危害, 并杀灭多种土壤细菌和病原体, 显示出强大的抑菌活性.也有学者研究发现, Phytolacca americana的发根会向根际分泌许多抗真菌活性蛋白, 如核糖体失活蛋白(RIP)、几丁质酶等(Park et al., 2002a).RIP通过具N-糖苷酶活性来抑制蛋白合成, 或选择性地删除28S rRNA保守区的腺嘌呤残基(Nielsen & Boston, 2001; Vepachedu et al., 2003).根系分泌的RIP还具有抗真菌特性, 如能够抑制根腐菌Rhizoctonia solani和Trichoderma reesei (Park et al., 2002b). ...
Quorum sensing in plant-pathogenic bacteria
1
2003
... 近年来, 根际生物学特性的研究不论是在研究方法与技术上, 还是在研究成果转化和生产实践指导上都取得了长足的进步.但是, 根际生物学特性的研究还需要从以下几方面进行突破: (1)根系分泌物原位收集、原位检测方法的改进与发展.目前对于根系分泌物的分析通常采用有机溶剂萃取并结合高效液相色谱法、气相质谱法等进行定性定量测定, 这些方法不但耗时, 而且无法实现田间实时动态原位检测.近年来, 生物传感器的问世在很大程度上促进了根系分泌物研究的发展, 如各种酚酸生物传感器的出现给农业科学与环境科学工作者提供了极大的便利(Soloducho & Cabaj, 2013).(2)植物响应根际生物胁迫的感知途径与机制.已知植物的免疫受体蛋白常分为模式识别受体(pattern recognition receptor)和效应子识别受体(effectors recognition receptor), 由这两类受体进行识别、激发的植物防卫反应分别称为病原物相关分子模式触发的免疫(PAMP-triggered immunity, PTI)和效应子触发的免疫(effector-triggered immunity, ETI).虽然PTI和ETI在植物天然免疫性中的作用已被认知, 但参与PTI及ETI信号转导的许多信号分子及蛋白组分仍未明确, 需要进一步确认.(3)植物与根际微生物“根际对话”的形式与信号分子.比如, 将来自耐旱或喜热植物中的内生真菌与其他植物进行共培, 可显著提高其他植物的耐旱性和耐热性, 但是内生真菌如何激活启动植物体内相关代谢途径以降低水分消耗和促进植物生长尚不清楚, 有待进一步研究.(4)微生物群体效应与植物生长发育的关系.在自然界中, 存在很多具有群体效应的有益或有害微生物, 如很多的植物病原菌都存在群体效应, 调控着病原菌在宿主根际的定殖过程及其对宿主的致病性, 通过深入研究这些植物病原菌的群体效应机制及其与宿主的互作过程, 可以帮助建立合理的农艺措施或通过转基因方法来控制这些具有群体效应的病原菌爆发成灾(von Bodman et al., 2003).当然, 通过对这些具有群体效应的有益菌的深入研究, 也可为农业生产服务, 本研究团队目前也在积极探索化感水稻根际中有关群体效应菌群(如黏细菌)在化感水稻抑制杂草过程中的作用, 但对于黏细菌与水稻根系分泌物的互作关系以及根系分泌物对黏细菌群体效应启动的研究还处于初步探索阶段.(5)微生物基因组学(microbial genomics)的运用.Lanfranco和Young (2012)通过对丛枝菌根转录组学的分析, 初步从基因水平上探索了丛枝菌根抑制宿主防御以及与宿主进行物质营养交换的过程.现在, 该项技术也被越来越多地运用于发现和鉴定介导植物-微生物二者互作关系的效应子(effector), 目前对于细菌释放的潜在效应子的研究呈指数增长, 而对于真菌和卵菌纲真菌效应子的研究还比较少(Ronald & Shirasu, 2012).(6)各种宏组学技术(如环境宏基因组学、宏转录组学、宏蛋白质组学、宏代谢组学)的进一步完善与突破, 以及它们在植物-微生物互作研究中的运用.如土壤蛋白质组学技术虽能够在蛋白质水平上为揭示土壤生态系统的生物化学过程提供依据, 但是它自身也存在一些不足之处, 特别是受到了技术的限制, 如土壤蛋白质提取纯化技术有待改善和环境微生物蛋白质库不完整.综上所述, 根际生物学特性研究还面临诸多的挑战, 需要多项技术的有机结合才能深入地揭示土壤这一“黑箱”所蕴含的奥秘. ...
Pseudomonas aeruginosa-plant root interactions: pathogenicity, biofilm formation, and root exudation.
1
2004
... 根系分泌物介导的植物-微生物互作关系是处在不断进化之中的.Walker等(2004)将Ocimum basilicum幼苗与病原微生物铜绿假单胞菌在试管内共培养, 发现在病原菌作用下Ocimum basilicum根系分泌物中酚酸类物质(如迷迭香酸(rosmarinic acid, RA)这一抗菌物质)含量显著增加, 进一步研究还发现铜绿假单胞菌具有群体效应, 能够在抗菌成分RA还未积累到有效作用浓度时便迅速产生生物被膜(biofilm), 以抵御植物分泌的抗菌物质对其造成伤害, 进而继续侵染植物根系.相反, 也有学者研究发现, 某些植物如豌豆(Pisum sativum)、大豆、水稻、苜蓿属(Medicago)植物等通过长期的进化, 对病原微生物形成一种防御机制, 即能够通过产生AHL类似物、AHL信号阻断物或降解酶等途径来干扰某些病原细菌的群体效应, 抑制其生长或干扰其正常生理功能, 从而保护植物自身(Fray, 2002). ...
Root exudation and rhizosphere biology
2
2003
... 植物根系分泌物的产生是植物根部长期进化的结果, 是适应环境的一种体现, 不是一个完全被动的过程, 因为植物的任何代谢功能都直接或间接地消耗碳源和能量.由于根部一直处于土壤中, 无时无刻不与周围的生物、非生物因子做斗争, 所以植物能够根据周围环境的变化不断地调整自身根系分泌物的组成和数量来积极响应(Walker et al., 2003).在根际中, 根系分泌物对某些微生物具有吸引作用, 这类具有趋化性的细菌或真菌就能够在根际中大量聚集和繁殖, 比如豆科植物对根瘤菌的诱导; 而对有些病原菌却具有抑制作用, 比如小麦根系分泌物中的酚酸具有抗细菌的活性, 并且其抗菌活性具有协同作用(Wu et al., 2001).Fierer等(2012)运用宏基因组学技术对取自寒漠、热沙漠、森林、草原、苔原的16份土样的微生物群落结构进行分析, 发现来自沙漠区的土壤微生物群落明显区别于非沙漠区的, 进一步的宏基因组功能分析发现来自沙漠区的微生物类群中与营养循环、分解代谢植物源有机化合物相关的基因含量很低, 尤其是来自寒漠地区的微生物群落, 其种类多样性和功能多样性都最低, 可见地上部物种多样性水平显著影响地下部微生物群落结构与功能多样性.相反, 地下部微生物群落也会对地上部植物的生理代谢、生长发育等产生重要影响.Badri等(2013)运用气相色谱-质谱联用仪(GC-MS)分析发现, 土壤微生物群落对植物叶片的代谢物组分有显著影响, 进而影响昆虫对叶片的取食行为.Mendes等(2011)运用PhyloChip芯片技术对抑病型土壤(disease suppressive soil)和利病型土壤(disease conducive soil)中的微生物群落结构进行分析, 发现种植于抑病型土壤的甜菜(Beta vulgaris)其根际优势群落普遍为拮抗病原菌相关的微生物, 如放线菌门、β变形菌门、γ变形菌门等, 尤其是其中的假单胞菌在抑病型土壤中的数量极显著高于利病型土壤中, 这是由根系分泌物介导下植物与根际特异微生物共同作用的结果.Zolla等(2013)研究发现, 在适度干旱胁迫下将拟南芥幼苗种植于同域土壤(sympatric soil, 即长期种植拟南芥的土壤)中, 可以显著地促进植物生长、提高生物量, 而种植于非同域土壤(non-sympatric soil, 如长期种植松树和玉米的土壤)中对植物生物量没有影响.进一步研究还发现, 这是由于同域土壤中存在很多与降低植物干旱胁迫响应相关的促生根际菌(plant growth promoting rhizobacteria, PGPR), 如伯克氏菌属、芽孢杆菌属、不动杆菌属等.Lakshmanan等(2012)研究还发现, 番茄(Lycopersicon esculentum)叶片受到病原菌侵染后, 可通过调节根系分泌物组分与含量, 如增加根系苹果酸分泌释放量, 使更多的苹果酸进入根际, 从而招募更多的有益菌向根际聚集, 这些有益菌可进一步引发植物的诱导性系统抗性(induced systemic resistance)以对病原菌产生防御反应, 可见根系分泌物在植物-微生物互作关系中所扮演的重要角色. ...
... 随着根系分泌物与根际微生物互馈研究的不断深入, 群体效应这一概念也得到广泛关注.群体效应是指细菌利用自身分泌的自诱导剂(autoind- ucer, AI)作为信号分子进行交流, 来控制群体生理行为的一个过程.革兰氏阴性菌、阳性菌都具有群体效应, 前者存在自诱导剂合成蛋白(luxI)/自诱导剂受体蛋白(luxR)介导的群体感应系统, 以N-酰基高丝氨酸内酯(AHL)为自诱导剂; 后者存在寡肽介导的群体感应系统, 以寡肽为自诱导剂(Walker et al., 2003).细菌不断繁殖的过程中, 当自诱导剂的浓度达到一定阈值时, 自诱导剂就会与受体蛋白结合, 从而启动目的基因的表达, 引起一系列的生理反应(如图1所示).在自然界中, 很多与植物生长相关的细菌都具有群体效应现象, 它们或使植物致病(如Pseudomonas syringae和Erwinia carotovora细菌), 或促进植物生长(如Pseudomonas putida和Pseudomonas aureofaciens细菌), 或抑制植物发病(如Pseudomonas fluorescens细菌).这些细菌能够通过自诱导剂介导的群体感应系统来了解自身的群体密度, 同时该感应系统还介导细菌抗生素的合成、致病因子的产生、生物膜的产生、质粒接合转移以及细胞的聚集移动等.既然植物病原菌能够通过群体效应系统来成功地侵害植物, 有学者开始寻找各种干扰群体效应的途径, 即群体效应猝灭(quorum- quenching), 试图抑制植物病原菌的生长, 从而控制植物病害.Chernin等(2011)研究发现, 一些植物促生菌(如荧光假单胞菌(Pseudomonas fluorescens)和普城沙雷氏菌(Serratia plymuthica))能够分泌一些挥发性化合物来猝灭植物病原菌相互交流的信号分子, 干扰其群体效应系统, 进而抑制病原菌正常的生理代谢和生长繁殖.所以, 群体效应猝灭逐渐被认为是控制植物病害的一种新策略(Ma et al., 2013). ...
Characterization of metaproteomics in crop rhizospheric soil
2
2011
... 近年来, 随着土壤蛋白质提取方法的不断优化, 土壤宏蛋白质的提取效率越来越高, 提取的纯度也越来越高, 足以进行土壤蛋白质双向电泳, 这就为土壤差异宏蛋白质组学研究奠定了基础.应用土壤宏蛋白质组学(soil metaproteomics)方法与技术研究植物根际生物学特性及其分子机理已成为当前科学研究的热点问题(Pierre-Alain et al., 2007; Wang et al., 2011).Wang等(2011)利用土壤蛋白质组学技术分析常见作物(水稻、太子参、地黄、甘蔗、烟草)根际生物学特性发现, 随机挑取的189个土壤蛋白质中, 有107个蛋白质来源于植物(56.61%), 有72个蛋白质来源于微生物(39.68%), 有10个蛋白质来源于动物(5.29%).该研究还通过T-RFLP技术对根际微生物群落结构进行分析, 结果发现, T-RFLP技术与土壤蛋白质组学技术既有相似性又有互补性, 二者都无法取代对方, 而是相互验证.Wu等(2011)进一步运用本实验优化建立的土壤宏蛋白质组学技术对连作下地黄根际土壤蛋白质表达谱变化进行研究, 发现地黄连作对来自植物、微生物的土壤蛋白质表达都产生显著影响, 发生差异表达的土壤蛋白质涉及能量、氨基酸、核酸代谢、次级代谢、胁迫防御、信号传递等功能, 它们在植物-微生物的“根际对话”中发挥着重要作用.同样, Lin等(2013)运用土壤宏蛋白质组学技术对新种植与宿根甘蔗根际土壤的宏蛋白质表达谱变化进行分析, 也发现了很多介导植物-微生物根际对话的关键土壤蛋白. ...
... ).Wang等(2011)利用土壤蛋白质组学技术分析常见作物(水稻、太子参、地黄、甘蔗、烟草)根际生物学特性发现, 随机挑取的189个土壤蛋白质中, 有107个蛋白质来源于植物(56.61%), 有72个蛋白质来源于微生物(39.68%), 有10个蛋白质来源于动物(5.29%).该研究还通过T-RFLP技术对根际微生物群落结构进行分析, 结果发现, T-RFLP技术与土壤蛋白质组学技术既有相似性又有互补性, 二者都无法取代对方, 而是相互验证.Wu等(2011)进一步运用本实验优化建立的土壤宏蛋白质组学技术对连作下地黄根际土壤蛋白质表达谱变化进行研究, 发现地黄连作对来自植物、微生物的土壤蛋白质表达都产生显著影响, 发生差异表达的土壤蛋白质涉及能量、氨基酸、核酸代谢、次级代谢、胁迫防御、信号传递等功能, 它们在植物-微生物的“根际对话”中发挥着重要作用.同样, Lin等(2013)运用土壤宏蛋白质组学技术对新种植与宿根甘蔗根际土壤的宏蛋白质表达谱变化进行分析, 也发现了很多介导植物-微生物根际对话的关键土壤蛋白. ...
Root exudation, phosphorus acquisition, and microbial diversity in the rhizosphere of white lupine as affected by phosphorus supply and atmospheric carbon dioxide concentration
1
2005
... 植物可以通过产生和释放根系分泌物来抵御各种非生物因素的干扰与生物胁迫.在金属污染物胁迫下, 某些植物的根系分泌物通过螯合、络合、沉淀等作用将金属污染物滞留在根外, 降低土壤中金属的生物有效性, 减少植物对有害金属的吸收(常学秀等, 2000).例如, 在Al胁迫下, 一些高等植物可分泌大量的柠檬酸、苹果酸、酚类化合物以及黏液、蛋白质复合物等来螯合游离的Aln+阳离子, 从而降低Al对植物根系的毒害作用(Neumann, 2007).也有研究表明, 某些植物通过分泌苹果酸、柠檬酸等根系分泌物来缓解重碳酸盐毒害, 同时这些分泌物也有利于植物吸收利用P、Fe、Zn、Mn等矿质元素(Hajiboland et al., 2003).近年来还陆续发现, 某些植物可以通过分泌几丁质酶、β-1, 3-葡聚糖酶、植物抗毒素(phytoalexin)等生物活性物质来抑制病原菌的生长(Neumann et al., 2000; Werner, 2000; Wasaki et al., 2005). ...
Organic signals between plants and microor- ganisms.
2
2000
... 根系分泌物(root exudate)是指在特定环境下, 活的植物通过根系的不同部位释放到根际环境中的有机物质的总称, 是一种复杂的非均一体系.广义的根系分泌物包括: (1)渗出物(diffusate), 即由根部细胞通过被动形式扩散出来的一些低分子量有机物质; (2)分泌物(secretion), 即由根部细胞主动释放的一些具有一定生理功能的有机物质, 对营养元素迁移、植物解毒、信号传递、抵御胁迫等起重要作用; (3)排泄物(excretion), 即根部细胞生物代谢过程中产生的分解产物(Neumann & Römheld, 2000; Werner, 2000).狭义的根系分泌物仅包括植物通过溢泌作用进入土壤的可溶性物质(贺永华等, 2006).根系分泌物可以通过改变根际物理、化学或生物学特性来提高植物根系对营养元素的吸收利用和适应外界环境的变化, 同时它也是调控根际微生态功能的关键因子. ...
... 植物可以通过产生和释放根系分泌物来抵御各种非生物因素的干扰与生物胁迫.在金属污染物胁迫下, 某些植物的根系分泌物通过螯合、络合、沉淀等作用将金属污染物滞留在根外, 降低土壤中金属的生物有效性, 减少植物对有害金属的吸收(常学秀等, 2000).例如, 在Al胁迫下, 一些高等植物可分泌大量的柠檬酸、苹果酸、酚类化合物以及黏液、蛋白质复合物等来螯合游离的Aln+阳离子, 从而降低Al对植物根系的毒害作用(Neumann, 2007).也有研究表明, 某些植物通过分泌苹果酸、柠檬酸等根系分泌物来缓解重碳酸盐毒害, 同时这些分泌物也有利于植物吸收利用P、Fe、Zn、Mn等矿质元素(Hajiboland et al., 2003).近年来还陆续发现, 某些植物可以通过分泌几丁质酶、β-1, 3-葡聚糖酶、植物抗毒素(phytoalexin)等生物活性物质来抑制病原菌的生长(Neumann et al., 2000; Werner, 2000; Wasaki et al., 2005). ...
Allelochemicals in Wheat (Triticum aestivum L.): cultivar difference in the exudation of phenolic acids.
1
2001
... 植物根系分泌物的产生是植物根部长期进化的结果, 是适应环境的一种体现, 不是一个完全被动的过程, 因为植物的任何代谢功能都直接或间接地消耗碳源和能量.由于根部一直处于土壤中, 无时无刻不与周围的生物、非生物因子做斗争, 所以植物能够根据周围环境的变化不断地调整自身根系分泌物的组成和数量来积极响应(Walker et al., 2003).在根际中, 根系分泌物对某些微生物具有吸引作用, 这类具有趋化性的细菌或真菌就能够在根际中大量聚集和繁殖, 比如豆科植物对根瘤菌的诱导; 而对有些病原菌却具有抑制作用, 比如小麦根系分泌物中的酚酸具有抗细菌的活性, 并且其抗菌活性具有协同作用(Wu et al., 2001).Fierer等(2012)运用宏基因组学技术对取自寒漠、热沙漠、森林、草原、苔原的16份土样的微生物群落结构进行分析, 发现来自沙漠区的土壤微生物群落明显区别于非沙漠区的, 进一步的宏基因组功能分析发现来自沙漠区的微生物类群中与营养循环、分解代谢植物源有机化合物相关的基因含量很低, 尤其是来自寒漠地区的微生物群落, 其种类多样性和功能多样性都最低, 可见地上部物种多样性水平显著影响地下部微生物群落结构与功能多样性.相反, 地下部微生物群落也会对地上部植物的生理代谢、生长发育等产生重要影响.Badri等(2013)运用气相色谱-质谱联用仪(GC-MS)分析发现, 土壤微生物群落对植物叶片的代谢物组分有显著影响, 进而影响昆虫对叶片的取食行为.Mendes等(2011)运用PhyloChip芯片技术对抑病型土壤(disease suppressive soil)和利病型土壤(disease conducive soil)中的微生物群落结构进行分析, 发现种植于抑病型土壤的甜菜(Beta vulgaris)其根际优势群落普遍为拮抗病原菌相关的微生物, 如放线菌门、β变形菌门、γ变形菌门等, 尤其是其中的假单胞菌在抑病型土壤中的数量极显著高于利病型土壤中, 这是由根系分泌物介导下植物与根际特异微生物共同作用的结果.Zolla等(2013)研究发现, 在适度干旱胁迫下将拟南芥幼苗种植于同域土壤(sympatric soil, 即长期种植拟南芥的土壤)中, 可以显著地促进植物生长、提高生物量, 而种植于非同域土壤(non-sympatric soil, 如长期种植松树和玉米的土壤)中对植物生物量没有影响.进一步研究还发现, 这是由于同域土壤中存在很多与降低植物干旱胁迫响应相关的促生根际菌(plant growth promoting rhizobacteria, PGPR), 如伯克氏菌属、芽孢杆菌属、不动杆菌属等.Lakshmanan等(2012)研究还发现, 番茄(Lycopersicon esculentum)叶片受到病原菌侵染后, 可通过调节根系分泌物组分与含量, 如增加根系苹果酸分泌释放量, 使更多的苹果酸进入根际, 从而招募更多的有益菌向根际聚集, 这些有益菌可进一步引发植物的诱导性系统抗性(induced systemic resistance)以对病原菌产生防御反应, 可见根系分泌物在植物-微生物互作关系中所扮演的重要角色. ...
Comparative metaproteomic analysis on consecu- tively Rehmannia glutinosa-monocultured rhizosphere soil.
1
2011
... 近年来, 随着土壤蛋白质提取方法的不断优化, 土壤宏蛋白质的提取效率越来越高, 提取的纯度也越来越高, 足以进行土壤蛋白质双向电泳, 这就为土壤差异宏蛋白质组学研究奠定了基础.应用土壤宏蛋白质组学(soil metaproteomics)方法与技术研究植物根际生物学特性及其分子机理已成为当前科学研究的热点问题(Pierre-Alain et al., 2007; Wang et al., 2011).Wang等(2011)利用土壤蛋白质组学技术分析常见作物(水稻、太子参、地黄、甘蔗、烟草)根际生物学特性发现, 随机挑取的189个土壤蛋白质中, 有107个蛋白质来源于植物(56.61%), 有72个蛋白质来源于微生物(39.68%), 有10个蛋白质来源于动物(5.29%).该研究还通过T-RFLP技术对根际微生物群落结构进行分析, 结果发现, T-RFLP技术与土壤蛋白质组学技术既有相似性又有互补性, 二者都无法取代对方, 而是相互验证.Wu等(2011)进一步运用本实验优化建立的土壤宏蛋白质组学技术对连作下地黄根际土壤蛋白质表达谱变化进行研究, 发现地黄连作对来自植物、微生物的土壤蛋白质表达都产生显著影响, 发生差异表达的土壤蛋白质涉及能量、氨基酸、核酸代谢、次级代谢、胁迫防御、信号传递等功能, 它们在植物-微生物的“根际对话”中发挥着重要作用.同样, Lin等(2013)运用土壤宏蛋白质组学技术对新种植与宿根甘蔗根际土壤的宏蛋白质表达谱变化进行分析, 也发现了很多介导植物-微生物根际对话的关键土壤蛋白. ...
Root herbivory in vitro: interactions between roots and aphids grown in aseptic coculture.
1
1999
... 当然, 土壤中也存在一些危害植物生长的有害土壤动物区系, 如植物病原线虫等, 但是某些植物能够通过释放特定的根系分泌物来抵御有害动物的威胁.Wu等(1999)通过体外发根-蚜虫共培试验发现, 在蚜虫的刺激下, 植物能够大量分泌一种植物抗毒素——聚乙烯.Bais等(2002b)发现, Oxalis tuberosa的根系分泌物中含有一种会发荧光的物质——β咔巴啉生物碱衍生物, 经鉴定, 为骆驼蓬碱(harmine)和骆驼蓬碱二氢(harmaline), 有抗菌杀虫作用.Larson等(1988)也发现, β咔巴啉生物碱衍生物对某些细菌、土壤动物具有光毒性. ...
Phytotoxic substances in root exudates of cucumber (Cucumis sativus L.).
1
1994
... 植物根系分泌物中含有一类对邻近其他植物或自身产生抑制或促进作用的生物活性物质, 即为化感物质(allelochemicals).Tang和Young (1982)研究了Hemarthria altissima根系分泌物的化感作用, 发现供体植物根系分泌物中对受体植物生长呈抑制作用的主要是酚类化合物.在小麦(Triticum aestivum)、水稻(Oryza sativa)、玉米等作物的根系分泌物中也都检测到一些化感物质, 对农业杂草都具有抑制作用, 而且化感作用的强弱与品种有关(何海斌等, 2005).同时, 研究发现黄瓜(Cucumis sativus)、大豆(Glycine max)、烟草(Nicotiana tabacum)、地黄(Rehmannia glutinosa)、西洋参(Panax quinquefolius)等忌连作作物的连作障碍现象与根系分泌物中的化感物质密切相关.Yu和Matsui (1994)发现黄瓜根系分泌物中含有苯甲酸、对羟基苯甲酸、2, 5-二羟基苯甲酸、苯丙烯酸等11种酚酸物质, 其中有10种具有生物毒性.Pramanik等(2000)也从黄瓜根系分泌物中鉴定出苯甲酸及其衍生物、肉桂酸及其衍生物等, 并证明这些物质会阻碍黄瓜对养分的吸收.近年来的研究也表明, 根系分泌物对某些入侵植物的成功入侵起到决定性的化学生态学作用.Bais等(2003)研究发现, 矢车菊(Centaurea maculosa)根系分泌释放的儿茶素(+)-Catechin和(-)-Catechin对其成功入侵起重要作用. ...
连作对地黄根际土壤细菌群落多样性的影响
1
2010
... 近年来, 对于植物化感作用的研究也倾向于根际微生物群落结构, 因为目前对于化感物质的成分与有效浓度仍存在较大争议, 有的学者认为酚酸类化感物质抑制杂草生长作用的有效浓度较高, 在田间自然条件下很难达到有效浓度, 故提出酚酸类不是化感物质, 而萜类和黄酮类才是化感物质(Kato-Noguchi, 2009).但是现在更多的学者认为, 化感物质进入根际土壤之后势必会受到微生物的代谢分解、加工转化等, 所以植物化感作用并非是在供体植物与受体植物之间直接发挥作用(林文雄等, 2007; Kaur et al., 2009).显然, 作物化感作用是植物化感物质与其根际土壤微生物综合作用的结果.本实验室运用现代生理生化技术、土壤宏基因组学并结合实时荧光定量PCR (qRT-PCR)技术等研究发现, 化感水稻与非化感水稻根际土壤微生物群落结构存在明显差异, 化感水稻根际存在30种特异微生物, 其中7种被鉴定为黏细菌(myxobacteria)(林瑞余等, 2007; 林辉锋, 2011), 它们对杂草(如稗(Echino- chloa crusgali))的发芽生长具有显著抑制作用.进一步研究还发现, 这些与水稻化感作用相关的黏细菌具有群体效应(quorum sensing), 而且水稻根系分泌物中的酚酸类物质对这类细菌具有明显的趋化促进作用(李颖哲, 2013).连作障碍, 也称再植病或土壤病, 是一种特殊的植物化感现象, 是指同一地块连续多年种植相同作物造成作物生长状况变差, 产量和品质降低, 病虫害发生加剧的现象.前人研究表明, 造成作物连作障碍的原因可能是多方面的, 比如土壤营养失衡、土壤理化性质恶化、自毒物质积累等(张重义和林文雄, 2009).但是, 近年来越来越多的学者认为, 根系分泌物生态效应的间接作用及土壤微生物区系紊乱是导致植物连作障碍形成的主要因素(Qi et al., 2009), 这可能是由于在根系分泌物特定组分的介导下, 某些类群的微生物(如土传病原菌)大量繁殖, 同时抑制其他有益微生物(如假单胞菌等拮抗菌)的生长, 进而改变了植物根系分泌物的组分和数量, 为趋化性病原微生物提供更多的碳源、能源, 形成恶性循环, 造成植物生长发育不良.本研究团队通过对地黄、太子参(Pseudostellariae hetero- phylla)、甘蔗(Saccharum officinarum)、烟草等不同作物连作下的根际微生态特性进行研究, 也均发现类似结果(陈冬梅等, 2010; 张重义等, 2010; 林茂兹等, 2012).Zhou和Wu (2012)通过外源添加黄瓜自毒物质——香豆酸至土壤中, 也发现香豆酸对土壤微生物群落结构产生显著影响, 导致厚壁菌门、β变形菌门等细菌大量增加, 而使拟杆菌门、δ变形菌门、浮霉菌门等细菌显著减少, 同时还造成土壤中病原菌(如尖孢镰刀菌等)大量繁殖增长.可见, 根系分泌物介导下的土壤微生物群落结构与功能多样性变化对其他植物或植物自身的生长发育都产生重要的影响. ...
连作对地黄根际土壤细菌群落多样性的影响
1
2010
... 近年来, 对于植物化感作用的研究也倾向于根际微生物群落结构, 因为目前对于化感物质的成分与有效浓度仍存在较大争议, 有的学者认为酚酸类化感物质抑制杂草生长作用的有效浓度较高, 在田间自然条件下很难达到有效浓度, 故提出酚酸类不是化感物质, 而萜类和黄酮类才是化感物质(Kato-Noguchi, 2009).但是现在更多的学者认为, 化感物质进入根际土壤之后势必会受到微生物的代谢分解、加工转化等, 所以植物化感作用并非是在供体植物与受体植物之间直接发挥作用(林文雄等, 2007; Kaur et al., 2009).显然, 作物化感作用是植物化感物质与其根际土壤微生物综合作用的结果.本实验室运用现代生理生化技术、土壤宏基因组学并结合实时荧光定量PCR (qRT-PCR)技术等研究发现, 化感水稻与非化感水稻根际土壤微生物群落结构存在明显差异, 化感水稻根际存在30种特异微生物, 其中7种被鉴定为黏细菌(myxobacteria)(林瑞余等, 2007; 林辉锋, 2011), 它们对杂草(如稗(Echino- chloa crusgali))的发芽生长具有显著抑制作用.进一步研究还发现, 这些与水稻化感作用相关的黏细菌具有群体效应(quorum sensing), 而且水稻根系分泌物中的酚酸类物质对这类细菌具有明显的趋化促进作用(李颖哲, 2013).连作障碍, 也称再植病或土壤病, 是一种特殊的植物化感现象, 是指同一地块连续多年种植相同作物造成作物生长状况变差, 产量和品质降低, 病虫害发生加剧的现象.前人研究表明, 造成作物连作障碍的原因可能是多方面的, 比如土壤营养失衡、土壤理化性质恶化、自毒物质积累等(张重义和林文雄, 2009).但是, 近年来越来越多的学者认为, 根系分泌物生态效应的间接作用及土壤微生物区系紊乱是导致植物连作障碍形成的主要因素(Qi et al., 2009), 这可能是由于在根系分泌物特定组分的介导下, 某些类群的微生物(如土传病原菌)大量繁殖, 同时抑制其他有益微生物(如假单胞菌等拮抗菌)的生长, 进而改变了植物根系分泌物的组分和数量, 为趋化性病原微生物提供更多的碳源、能源, 形成恶性循环, 造成植物生长发育不良.本研究团队通过对地黄、太子参(Pseudostellariae hetero- phylla)、甘蔗(Saccharum officinarum)、烟草等不同作物连作下的根际微生态特性进行研究, 也均发现类似结果(陈冬梅等, 2010; 张重义等, 2010; 林茂兹等, 2012).Zhou和Wu (2012)通过外源添加黄瓜自毒物质——香豆酸至土壤中, 也发现香豆酸对土壤微生物群落结构产生显著影响, 导致厚壁菌门、β变形菌门等细菌大量增加, 而使拟杆菌门、δ变形菌门、浮霉菌门等细菌显著减少, 同时还造成土壤中病原菌(如尖孢镰刀菌等)大量繁殖增长.可见, 根系分泌物介导下的土壤微生物群落结构与功能多样性变化对其他植物或植物自身的生长发育都产生重要的影响. ...
药用植物的化感自毒作用与连作障碍
1
2009
... 近年来, 对于植物化感作用的研究也倾向于根际微生物群落结构, 因为目前对于化感物质的成分与有效浓度仍存在较大争议, 有的学者认为酚酸类化感物质抑制杂草生长作用的有效浓度较高, 在田间自然条件下很难达到有效浓度, 故提出酚酸类不是化感物质, 而萜类和黄酮类才是化感物质(Kato-Noguchi, 2009).但是现在更多的学者认为, 化感物质进入根际土壤之后势必会受到微生物的代谢分解、加工转化等, 所以植物化感作用并非是在供体植物与受体植物之间直接发挥作用(林文雄等, 2007; Kaur et al., 2009).显然, 作物化感作用是植物化感物质与其根际土壤微生物综合作用的结果.本实验室运用现代生理生化技术、土壤宏基因组学并结合实时荧光定量PCR (qRT-PCR)技术等研究发现, 化感水稻与非化感水稻根际土壤微生物群落结构存在明显差异, 化感水稻根际存在30种特异微生物, 其中7种被鉴定为黏细菌(myxobacteria)(林瑞余等, 2007; 林辉锋, 2011), 它们对杂草(如稗(Echino- chloa crusgali))的发芽生长具有显著抑制作用.进一步研究还发现, 这些与水稻化感作用相关的黏细菌具有群体效应(quorum sensing), 而且水稻根系分泌物中的酚酸类物质对这类细菌具有明显的趋化促进作用(李颖哲, 2013).连作障碍, 也称再植病或土壤病, 是一种特殊的植物化感现象, 是指同一地块连续多年种植相同作物造成作物生长状况变差, 产量和品质降低, 病虫害发生加剧的现象.前人研究表明, 造成作物连作障碍的原因可能是多方面的, 比如土壤营养失衡、土壤理化性质恶化、自毒物质积累等(张重义和林文雄, 2009).但是, 近年来越来越多的学者认为, 根系分泌物生态效应的间接作用及土壤微生物区系紊乱是导致植物连作障碍形成的主要因素(Qi et al., 2009), 这可能是由于在根系分泌物特定组分的介导下, 某些类群的微生物(如土传病原菌)大量繁殖, 同时抑制其他有益微生物(如假单胞菌等拮抗菌)的生长, 进而改变了植物根系分泌物的组分和数量, 为趋化性病原微生物提供更多的碳源、能源, 形成恶性循环, 造成植物生长发育不良.本研究团队通过对地黄、太子参(Pseudostellariae hetero- phylla)、甘蔗(Saccharum officinarum)、烟草等不同作物连作下的根际微生态特性进行研究, 也均发现类似结果(陈冬梅等, 2010; 张重义等, 2010; 林茂兹等, 2012).Zhou和Wu (2012)通过外源添加黄瓜自毒物质——香豆酸至土壤中, 也发现香豆酸对土壤微生物群落结构产生显著影响, 导致厚壁菌门、β变形菌门等细菌大量增加, 而使拟杆菌门、δ变形菌门、浮霉菌门等细菌显著减少, 同时还造成土壤中病原菌(如尖孢镰刀菌等)大量繁殖增长.可见, 根系分泌物介导下的土壤微生物群落结构与功能多样性变化对其他植物或植物自身的生长发育都产生重要的影响. ...
药用植物的化感自毒作用与连作障碍
1
2009
... 近年来, 对于植物化感作用的研究也倾向于根际微生物群落结构, 因为目前对于化感物质的成分与有效浓度仍存在较大争议, 有的学者认为酚酸类化感物质抑制杂草生长作用的有效浓度较高, 在田间自然条件下很难达到有效浓度, 故提出酚酸类不是化感物质, 而萜类和黄酮类才是化感物质(Kato-Noguchi, 2009).但是现在更多的学者认为, 化感物质进入根际土壤之后势必会受到微生物的代谢分解、加工转化等, 所以植物化感作用并非是在供体植物与受体植物之间直接发挥作用(林文雄等, 2007; Kaur et al., 2009).显然, 作物化感作用是植物化感物质与其根际土壤微生物综合作用的结果.本实验室运用现代生理生化技术、土壤宏基因组学并结合实时荧光定量PCR (qRT-PCR)技术等研究发现, 化感水稻与非化感水稻根际土壤微生物群落结构存在明显差异, 化感水稻根际存在30种特异微生物, 其中7种被鉴定为黏细菌(myxobacteria)(林瑞余等, 2007; 林辉锋, 2011), 它们对杂草(如稗(Echino- chloa crusgali))的发芽生长具有显著抑制作用.进一步研究还发现, 这些与水稻化感作用相关的黏细菌具有群体效应(quorum sensing), 而且水稻根系分泌物中的酚酸类物质对这类细菌具有明显的趋化促进作用(李颖哲, 2013).连作障碍, 也称再植病或土壤病, 是一种特殊的植物化感现象, 是指同一地块连续多年种植相同作物造成作物生长状况变差, 产量和品质降低, 病虫害发生加剧的现象.前人研究表明, 造成作物连作障碍的原因可能是多方面的, 比如土壤营养失衡、土壤理化性质恶化、自毒物质积累等(张重义和林文雄, 2009).但是, 近年来越来越多的学者认为, 根系分泌物生态效应的间接作用及土壤微生物区系紊乱是导致植物连作障碍形成的主要因素(Qi et al., 2009), 这可能是由于在根系分泌物特定组分的介导下, 某些类群的微生物(如土传病原菌)大量繁殖, 同时抑制其他有益微生物(如假单胞菌等拮抗菌)的生长, 进而改变了植物根系分泌物的组分和数量, 为趋化性病原微生物提供更多的碳源、能源, 形成恶性循环, 造成植物生长发育不良.本研究团队通过对地黄、太子参(Pseudostellariae hetero- phylla)、甘蔗(Saccharum officinarum)、烟草等不同作物连作下的根际微生态特性进行研究, 也均发现类似结果(陈冬梅等, 2010; 张重义等, 2010; 林茂兹等, 2012).Zhou和Wu (2012)通过外源添加黄瓜自毒物质——香豆酸至土壤中, 也发现香豆酸对土壤微生物群落结构产生显著影响, 导致厚壁菌门、β变形菌门等细菌大量增加, 而使拟杆菌门、δ变形菌门、浮霉菌门等细菌显著减少, 同时还造成土壤中病原菌(如尖孢镰刀菌等)大量繁殖增长.可见, 根系分泌物介导下的土壤微生物群落结构与功能多样性变化对其他植物或植物自身的生长发育都产生重要的影响. ...
P-Coumaric acid influenced cucumber rhizosphere soil microbial communities and the growth of Fusarium oxysporum f. sp. cucumerinum Owen.
1
2012
... 近年来, 对于植物化感作用的研究也倾向于根际微生物群落结构, 因为目前对于化感物质的成分与有效浓度仍存在较大争议, 有的学者认为酚酸类化感物质抑制杂草生长作用的有效浓度较高, 在田间自然条件下很难达到有效浓度, 故提出酚酸类不是化感物质, 而萜类和黄酮类才是化感物质(Kato-Noguchi, 2009).但是现在更多的学者认为, 化感物质进入根际土壤之后势必会受到微生物的代谢分解、加工转化等, 所以植物化感作用并非是在供体植物与受体植物之间直接发挥作用(林文雄等, 2007; Kaur et al., 2009).显然, 作物化感作用是植物化感物质与其根际土壤微生物综合作用的结果.本实验室运用现代生理生化技术、土壤宏基因组学并结合实时荧光定量PCR (qRT-PCR)技术等研究发现, 化感水稻与非化感水稻根际土壤微生物群落结构存在明显差异, 化感水稻根际存在30种特异微生物, 其中7种被鉴定为黏细菌(myxobacteria)(林瑞余等, 2007; 林辉锋, 2011), 它们对杂草(如稗(Echino- chloa crusgali))的发芽生长具有显著抑制作用.进一步研究还发现, 这些与水稻化感作用相关的黏细菌具有群体效应(quorum sensing), 而且水稻根系分泌物中的酚酸类物质对这类细菌具有明显的趋化促进作用(李颖哲, 2013).连作障碍, 也称再植病或土壤病, 是一种特殊的植物化感现象, 是指同一地块连续多年种植相同作物造成作物生长状况变差, 产量和品质降低, 病虫害发生加剧的现象.前人研究表明, 造成作物连作障碍的原因可能是多方面的, 比如土壤营养失衡、土壤理化性质恶化、自毒物质积累等(张重义和林文雄, 2009).但是, 近年来越来越多的学者认为, 根系分泌物生态效应的间接作用及土壤微生物区系紊乱是导致植物连作障碍形成的主要因素(Qi et al., 2009), 这可能是由于在根系分泌物特定组分的介导下, 某些类群的微生物(如土传病原菌)大量繁殖, 同时抑制其他有益微生物(如假单胞菌等拮抗菌)的生长, 进而改变了植物根系分泌物的组分和数量, 为趋化性病原微生物提供更多的碳源、能源, 形成恶性循环, 造成植物生长发育不良.本研究团队通过对地黄、太子参(Pseudostellariae hetero- phylla)、甘蔗(Saccharum officinarum)、烟草等不同作物连作下的根际微生态特性进行研究, 也均发现类似结果(陈冬梅等, 2010; 张重义等, 2010; 林茂兹等, 2012).Zhou和Wu (2012)通过外源添加黄瓜自毒物质——香豆酸至土壤中, 也发现香豆酸对土壤微生物群落结构产生显著影响, 导致厚壁菌门、β变形菌门等细菌大量增加, 而使拟杆菌门、δ变形菌门、浮霉菌门等细菌显著减少, 同时还造成土壤中病原菌(如尖孢镰刀菌等)大量繁殖增长.可见, 根系分泌物介导下的土壤微生物群落结构与功能多样性变化对其他植物或植物自身的生长发育都产生重要的影响. ...
Soil microbiomes vary in their ability to confer drought tolerance to Arabidopsis.
1
2013
... 植物根系分泌物的产生是植物根部长期进化的结果, 是适应环境的一种体现, 不是一个完全被动的过程, 因为植物的任何代谢功能都直接或间接地消耗碳源和能量.由于根部一直处于土壤中, 无时无刻不与周围的生物、非生物因子做斗争, 所以植物能够根据周围环境的变化不断地调整自身根系分泌物的组成和数量来积极响应(Walker et al., 2003).在根际中, 根系分泌物对某些微生物具有吸引作用, 这类具有趋化性的细菌或真菌就能够在根际中大量聚集和繁殖, 比如豆科植物对根瘤菌的诱导; 而对有些病原菌却具有抑制作用, 比如小麦根系分泌物中的酚酸具有抗细菌的活性, 并且其抗菌活性具有协同作用(Wu et al., 2001).Fierer等(2012)运用宏基因组学技术对取自寒漠、热沙漠、森林、草原、苔原的16份土样的微生物群落结构进行分析, 发现来自沙漠区的土壤微生物群落明显区别于非沙漠区的, 进一步的宏基因组功能分析发现来自沙漠区的微生物类群中与营养循环、分解代谢植物源有机化合物相关的基因含量很低, 尤其是来自寒漠地区的微生物群落, 其种类多样性和功能多样性都最低, 可见地上部物种多样性水平显著影响地下部微生物群落结构与功能多样性.相反, 地下部微生物群落也会对地上部植物的生理代谢、生长发育等产生重要影响.Badri等(2013)运用气相色谱-质谱联用仪(GC-MS)分析发现, 土壤微生物群落对植物叶片的代谢物组分有显著影响, 进而影响昆虫对叶片的取食行为.Mendes等(2011)运用PhyloChip芯片技术对抑病型土壤(disease suppressive soil)和利病型土壤(disease conducive soil)中的微生物群落结构进行分析, 发现种植于抑病型土壤的甜菜(Beta vulgaris)其根际优势群落普遍为拮抗病原菌相关的微生物, 如放线菌门、β变形菌门、γ变形菌门等, 尤其是其中的假单胞菌在抑病型土壤中的数量极显著高于利病型土壤中, 这是由根系分泌物介导下植物与根际特异微生物共同作用的结果.Zolla等(2013)研究发现, 在适度干旱胁迫下将拟南芥幼苗种植于同域土壤(sympatric soil, 即长期种植拟南芥的土壤)中, 可以显著地促进植物生长、提高生物量, 而种植于非同域土壤(non-sympatric soil, 如长期种植松树和玉米的土壤)中对植物生物量没有影响.进一步研究还发现, 这是由于同域土壤中存在很多与降低植物干旱胁迫响应相关的促生根际菌(plant growth promoting rhizobacteria, PGPR), 如伯克氏菌属、芽孢杆菌属、不动杆菌属等.Lakshmanan等(2012)研究还发现, 番茄(Lycopersicon esculentum)叶片受到病原菌侵染后, 可通过调节根系分泌物组分与含量, 如增加根系苹果酸分泌释放量, 使更多的苹果酸进入根际, 从而招募更多的有益菌向根际聚集, 这些有益菌可进一步引发植物的诱导性系统抗性(induced systemic resistance)以对病原菌产生防御反应, 可见根系分泌物在植物-微生物互作关系中所扮演的重要角色. ...
Rhizosphere protozoa: their significance in nutrient dynamics.
1
1994
... 土壤动物群的生长、分布与植物根系分泌物关系密切.有研究表明, 根际土中的原生动物数量明显高于非根际土中的, 这可能是由于根系分泌物促进了土壤细菌、真菌的生长繁殖, 从而有利于以细菌、真菌为食的原生动物的生长(Zwart et al., 1994).土壤动物区系除了通过食用植物残体碎屑, 捕食细菌、真菌, 改变土壤微生物区系, 改善土壤理化性质来加速土壤物质循环外, 很多土壤动物(如原生动物、蚯蚓等)还能分泌一些激素类有机物来促进植物生长(Canellas et al., 2002). ...



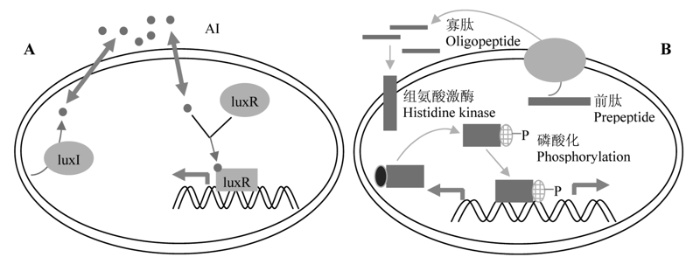


{kind=link}
{kind=link}