Variations of non-structural carbohydrate concentration of Picea meyeri at different elevations of Luya Mountain, China
WANG Biao1, 2, JIANG Yuan1, 2, 3, , WANG Ming-Chang1, 2, DONG Man-Yu1, 2, ZHANG Yi-Ping1, 2
1Beijing Municipal Key Laboratory of Traditional Chinese Medicine Protection and Utilization, Beijing Normal University, Beijing 100875, China2College of Resources Science and Technology, Beijing Normal University, Beijing 100875, China3State Key Laboratory of Earth Surface Process and Resource Ecology, Beijing Normal University, Beijing 100875, China
Aims The alpine timberline is highly sensitive to environmental changes, although the mechanism controlling timberline formation is still inconclusive. Our objectives in this study were to test whether the alpine timberline formation is determined by carbon limitation or growth limitation, and explore physiological and ecological mechanisms of timberline tree species adapting to alpine environments. We examined the concentrations of the overall nonstructural carbohydrates (NSC) and tissues NSC of Picea meyeri at the end of growing season and in three elevations (low, medium and timberline) along an altitudinal gradient on the north slope of Luya Mountain, Shanxi, China. Methods We collected samples of leaf, stem and fine root tissues of P. meyeri on September 15, 2013. The total soluble sugar concentration of plant tissue was measured by an anthrone-sulfuric acid colorimetric method, and starch was extracted by a perchloric acid method. Important findings The overall NSC and tissues NSC increased significantly with elevation, suggesting that there was no carbon limitation at the alpine timberline. The NSC source and sink are all increased significantly with elevation, and there is no significant difference in the source-sink ratio among three elevations, indicating an adaptation of source-sink balances to altitudes and no restriction of carbon source activity in timberline trees. The ratio of sugar to starch in tissues showed an increasing trend with elevation, which suggests that the colder the environment was, the stronger the protective strategy adopted in plant tissues through resource investments, implying more growth limitation in trees near timberlines, The research results appear to support the “growth limit” hypothesis to some degree.
Keywords:elevation
;
nonstructural carbohydrate
;
source-sink balance
;
ratio of soluble sugar to starch
;
Picea meyeri
;
timberline
WANGBiao, JIANGYuan, WANGMing-Chang, DONGMan-Yu, ZHANGYi-Ping. Variations of non-structural carbohydrate concentration of Picea meyeri at different elevations of Luya Mountain, China[J]. Chinese Journal of Plant Ecology, 2015, 39(7): 746-752 https://doi.org/10.17521/cjpe.2015.0071
Table 1 Non-structural carbohydrate (NSC) across tissues, NSC in source (needles) and sink (carbon storage organs-fine roots and stem sapwood), and source-sink ratio of NSC (SSR-NSC = source NSC/sink NSC) in Picea meyeri growing at different elevations
海拔 Elevation (m)
NSC总体含量 Overall NSC (%)
NSC源 NSC in source (%)
NSC汇 NSC in sink (%)
源-汇比 SSR-NSC
2 040
12.09b
19.49b
9.62b
2.0a
2 400
12.62b
19.48b
10.33a
1.9a
2 740
14.56a
23.12a
11.71a
2.0a
Small letters indicate significant difference among different elevations (Duncan test, p < 0.05).不同小写字母表示不同海拔间的差异显著(Duncan多重比较, p < 0.05)。
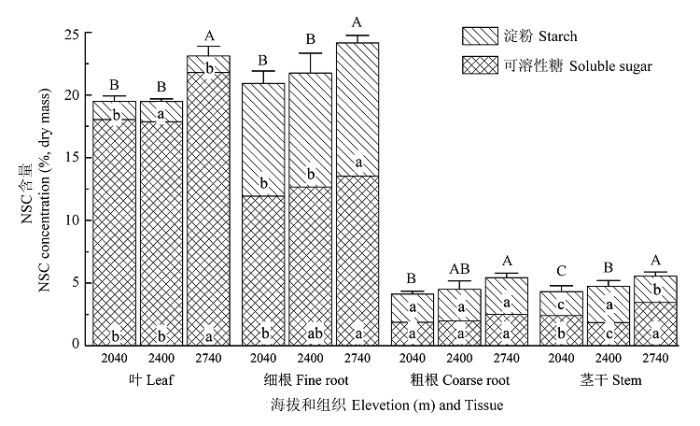
图1 不同海拔白杄各组织非结构性碳水化合物(NSC)含量(可溶性糖含量与淀粉含量之和)(平均值±标准偏差, n = 5)。不同大写字母(表示NSC)和小写字母(上部字母代表淀粉, 下部字母代表可溶性糖)表示同一组织不同海拔间NSC差异显著(p < 0.05, Duncan test)。
Fig. 1 Non-structural carbohydrate (NSC) concentrations in tissues of Picea meyeri growing at different elevations (mean ± SD, n = 5). Capital (for NSC ) and small letters (uppercase letters for sugars, and lowercase letters for starch) indicate significant differences (p < 0.05, Duncan test) among different altitudes.
Immediate responses and adaptative strategies of three olive cultivars under contrasting water availability regimes: Changes on structure and chemical composition of foliage and oxidative damage.
GoldsteinG, MeinzerFC, RadaF (1994). Environmental biology of a tropical treeline species, Polylepis sericea. In: Rundel PW, Smith AP, Meinzer FC eds. Tropical Alpine Environments: Plant Form and Function. Cambridge University Press, Cambridge. 129-149.
Studies on the relationship between the changes of fat, soluble sugar and flavoniods and the adaptation to environments of alpine subnival plants at the source of Ürümqi River.
Journal of Glaciology and Geocryology, 29, 947-952.
Altering flux through the sucrose biosynthesis pathway in transgenic Arabidopsis thaliana modifies photosynthetic acclimation at low temperatures and the development of freezing tolerance
Immediate responses and adaptative strategies of three olive cultivars under contrasting water availability regimes: Changes on structure and chemical composition of foliage and oxidative damage.
A re-assessment of high elevation treeline positions and their explanation.
2
1998
... 作为最明显的生态界限之一, 高山林线由于其特殊的结构、功能及对气候变化的高度敏感性, 已经成为全球气候变化研究的热点区域之一( Li et al., 2001, 2002).全球高山林线归根结底是一条低温界限, 其形成被认为是生长季低温影响了树木碳平衡, 从而限制树木海拔分布的结果(Körner, 1998, 2003a; Hoch & Körner, 2003; Körner & Paulsen, 2004; Shi et al., 2008).关于高山林线形成的原因已经争论了逾百年(Körner, 2003a), 并提出了多种假说(Körner, 2003b; Li et al., 2008a).Körner (2003a)和李迈和和Kräuchi (2005)将这些假说总结为5类, 即环境胁迫假说、干扰假说、更新繁殖障碍假说、碳平衡失调假说和生长限制假说, 并认为“碳平衡失调”或“生长限制”假说最有可能在全球尺度上解释高山林线现象.“碳平衡失调”假说(Stevens & Fox, 1991)认为: 低温、干旱、生长季节较短及其他环境胁迫因素, 使植物的碳吸收与碳消耗关系失调, 导致碳供应不足, 进而引起高山林线的形成.“生长限制”假说(Körner., 1998; Li et al., 2002)的前提条件是碳供应充足(即不存在“碳平衡失调”问题), 低温导致充足的碳不能被生长所利用, 从而使生长受限, 导致高山林线的形成.但是, 对于高山林线的形成是由于碳供应不足还是低温导致树木生长受限, 至今仍然没有明确的结论(Shi et al., 2006; Li et al., 2008a, 2008b). ...
... )认为: 低温、干旱、生长季节较短及其他环境胁迫因素, 使植物的碳吸收与碳消耗关系失调, 导致碳供应不足, 进而引起高山林线的形成.“生长限制”假说(Körner., 1998; Li et al., 2002)的前提条件是碳供应充足(即不存在“碳平衡失调”问题), 低温导致充足的碳不能被生长所利用, 从而使生长受限, 导致高山林线的形成.但是, 对于高山林线的形成是由于碳供应不足还是低温导致树木生长受限, 至今仍然没有明确的结论(Shi et al., 2006; Li et al., 2008a, 2008b). ...
Alpine Plant Life: Functional Plant Ecology of High Mountain Ecosystems
3
2003a
... 作为最明显的生态界限之一, 高山林线由于其特殊的结构、功能及对气候变化的高度敏感性, 已经成为全球气候变化研究的热点区域之一( Li et al., 2001, 2002).全球高山林线归根结底是一条低温界限, 其形成被认为是生长季低温影响了树木碳平衡, 从而限制树木海拔分布的结果(Körner, 1998, 2003a; Hoch & Körner, 2003; Körner & Paulsen, 2004; Shi et al., 2008).关于高山林线形成的原因已经争论了逾百年(Körner, 2003a), 并提出了多种假说(Körner, 2003b; Li et al., 2008a).Körner (2003a)和李迈和和Kräuchi (2005)将这些假说总结为5类, 即环境胁迫假说、干扰假说、更新繁殖障碍假说、碳平衡失调假说和生长限制假说, 并认为“碳平衡失调”或“生长限制”假说最有可能在全球尺度上解释高山林线现象.“碳平衡失调”假说(Stevens & Fox, 1991)认为: 低温、干旱、生长季节较短及其他环境胁迫因素, 使植物的碳吸收与碳消耗关系失调, 导致碳供应不足, 进而引起高山林线的形成.“生长限制”假说(Körner., 1998; Li et al., 2002)的前提条件是碳供应充足(即不存在“碳平衡失调”问题), 低温导致充足的碳不能被生长所利用, 从而使生长受限, 导致高山林线的形成.但是, 对于高山林线的形成是由于碳供应不足还是低温导致树木生长受限, 至今仍然没有明确的结论(Shi et al., 2006; Li et al., 2008a, 2008b). ...
... ).关于高山林线形成的原因已经争论了逾百年(Körner, 2003a), 并提出了多种假说(Körner, 2003b; Li et al., 2008a).Körner (2003a)和李迈和和Kräuchi (2005)将这些假说总结为5类, 即环境胁迫假说、干扰假说、更新繁殖障碍假说、碳平衡失调假说和生长限制假说, 并认为“碳平衡失调”或“生长限制”假说最有可能在全球尺度上解释高山林线现象.“碳平衡失调”假说(Stevens & Fox, 1991)认为: 低温、干旱、生长季节较短及其他环境胁迫因素, 使植物的碳吸收与碳消耗关系失调, 导致碳供应不足, 进而引起高山林线的形成.“生长限制”假说(Körner., 1998; Li et al., 2002)的前提条件是碳供应充足(即不存在“碳平衡失调”问题), 低温导致充足的碳不能被生长所利用, 从而使生长受限, 导致高山林线的形成.但是, 对于高山林线的形成是由于碳供应不足还是低温导致树木生长受限, 至今仍然没有明确的结论(Shi et al., 2006; Li et al., 2008a, 2008b). ...
... ).Körner (2003a)和李迈和和Kräuchi (2005)将这些假说总结为5类, 即环境胁迫假说、干扰假说、更新繁殖障碍假说、碳平衡失调假说和生长限制假说, 并认为“碳平衡失调”或“生长限制”假说最有可能在全球尺度上解释高山林线现象.“碳平衡失调”假说(Stevens & Fox, 1991)认为: 低温、干旱、生长季节较短及其他环境胁迫因素, 使植物的碳吸收与碳消耗关系失调, 导致碳供应不足, 进而引起高山林线的形成.“生长限制”假说(Körner., 1998; Li et al., 2002)的前提条件是碳供应充足(即不存在“碳平衡失调”问题), 低温导致充足的碳不能被生长所利用, 从而使生长受限, 导致高山林线的形成.但是, 对于高山林线的形成是由于碳供应不足还是低温导致树木生长受限, 至今仍然没有明确的结论(Shi et al., 2006; Li et al., 2008a, 2008b). ...
Carbon limitation in trees.
4
2003b
... 作为最明显的生态界限之一, 高山林线由于其特殊的结构、功能及对气候变化的高度敏感性, 已经成为全球气候变化研究的热点区域之一( Li et al., 2001, 2002).全球高山林线归根结底是一条低温界限, 其形成被认为是生长季低温影响了树木碳平衡, 从而限制树木海拔分布的结果(Körner, 1998, 2003a; Hoch & Körner, 2003; Körner & Paulsen, 2004; Shi et al., 2008).关于高山林线形成的原因已经争论了逾百年(Körner, 2003a), 并提出了多种假说(Körner, 2003b; Li et al., 2008a).Körner (2003a)和李迈和和Kräuchi (2005)将这些假说总结为5类, 即环境胁迫假说、干扰假说、更新繁殖障碍假说、碳平衡失调假说和生长限制假说, 并认为“碳平衡失调”或“生长限制”假说最有可能在全球尺度上解释高山林线现象.“碳平衡失调”假说(Stevens & Fox, 1991)认为: 低温、干旱、生长季节较短及其他环境胁迫因素, 使植物的碳吸收与碳消耗关系失调, 导致碳供应不足, 进而引起高山林线的形成.“生长限制”假说(Körner., 1998; Li et al., 2002)的前提条件是碳供应充足(即不存在“碳平衡失调”问题), 低温导致充足的碳不能被生长所利用, 从而使生长受限, 导致高山林线的形成.但是, 对于高山林线的形成是由于碳供应不足还是低温导致树木生长受限, 至今仍然没有明确的结论(Shi et al., 2006; Li et al., 2008a, 2008b). ...
... 生长于低温环境下的高山林线树木既需要足够的碳水化合物维持其生长, 也需要充足的可溶性碳水化合物来提高其在低温等不利环境下的生存能力(Körner, 2003b; 李迈和和Kräuchi, 2005).评价植物碳供应状态的一种可行方法是分析植物体内非结构性碳库的大小(Shi et al., 2006; Li et al., 2008a, 2008b).非结构性碳水化合物(non-structural carbohydrate, NSC)是树木休眠后恢复生长的主要碳供应者, 并能在植物光合作用不足时起到缓冲作用, 因此在很大程度上影响着植株的生长及对环境的响应(Li et al., 2001, 2002; Hoch & Körner, 2003; Körner, 2003b).植物组织中NSC含量(主要是可溶性糖和淀粉)的大小可看作是植物碳同化与碳消耗(即供应与需求)之间平衡关系的结果.树木组织中NSC含量随海拔升高而减少, 碳限制随海拔增加, 则意味着林线附近树木碳供应不足(碳平衡失调假说).相反, 树木组织形成受到限制(生长限制假说), 由于树木生长的碳需求减小, 则NSC含量随海拔升高而增加(Li et al., 2008a). ...
Spatial variability of mobile carbohydrates within Pinus cembra trees at the alpine treeline.
2
2001
... 作为最明显的生态界限之一, 高山林线由于其特殊的结构、功能及对气候变化的高度敏感性, 已经成为全球气候变化研究的热点区域之一( Li et al., 2001, 2002).全球高山林线归根结底是一条低温界限, 其形成被认为是生长季低温影响了树木碳平衡, 从而限制树木海拔分布的结果(Körner, 1998, 2003a; Hoch & Körner, 2003; Körner & Paulsen, 2004; Shi et al., 2008).关于高山林线形成的原因已经争论了逾百年(Körner, 2003a), 并提出了多种假说(Körner, 2003b; Li et al., 2008a).Körner (2003a)和李迈和和Kräuchi (2005)将这些假说总结为5类, 即环境胁迫假说、干扰假说、更新繁殖障碍假说、碳平衡失调假说和生长限制假说, 并认为“碳平衡失调”或“生长限制”假说最有可能在全球尺度上解释高山林线现象.“碳平衡失调”假说(Stevens & Fox, 1991)认为: 低温、干旱、生长季节较短及其他环境胁迫因素, 使植物的碳吸收与碳消耗关系失调, 导致碳供应不足, 进而引起高山林线的形成.“生长限制”假说(Körner., 1998; Li et al., 2002)的前提条件是碳供应充足(即不存在“碳平衡失调”问题), 低温导致充足的碳不能被生长所利用, 从而使生长受限, 导致高山林线的形成.但是, 对于高山林线的形成是由于碳供应不足还是低温导致树木生长受限, 至今仍然没有明确的结论(Shi et al., 2006; Li et al., 2008a, 2008b). ...
... 生长于低温环境下的高山林线树木既需要足够的碳水化合物维持其生长, 也需要充足的可溶性碳水化合物来提高其在低温等不利环境下的生存能力(Körner, 2003b; 李迈和和Kräuchi, 2005).评价植物碳供应状态的一种可行方法是分析植物体内非结构性碳库的大小(Shi et al., 2006; Li et al., 2008a, 2008b).非结构性碳水化合物(non-structural carbohydrate, NSC)是树木休眠后恢复生长的主要碳供应者, 并能在植物光合作用不足时起到缓冲作用, 因此在很大程度上影响着植株的生长及对环境的响应(Li et al., 2001, 2002; Hoch & Körner, 2003; Körner, 2003b).植物组织中NSC含量(主要是可溶性糖和淀粉)的大小可看作是植物碳同化与碳消耗(即供应与需求)之间平衡关系的结果.树木组织中NSC含量随海拔升高而减少, 碳限制随海拔增加, 则意味着林线附近树木碳供应不足(碳平衡失调假说).相反, 树木组织形成受到限制(生长限制假说), 由于树木生长的碳需求减小, 则NSC含量随海拔升高而增加(Li et al., 2008a). ...
Source/sink removal affects mobile carbohydrates in Pinus cembra at the Swiss treeline.
5
2002
... 作为最明显的生态界限之一, 高山林线由于其特殊的结构、功能及对气候变化的高度敏感性, 已经成为全球气候变化研究的热点区域之一( Li et al., 2001, 2002).全球高山林线归根结底是一条低温界限, 其形成被认为是生长季低温影响了树木碳平衡, 从而限制树木海拔分布的结果(Körner, 1998, 2003a; Hoch & Körner, 2003; Körner & Paulsen, 2004; Shi et al., 2008).关于高山林线形成的原因已经争论了逾百年(Körner, 2003a), 并提出了多种假说(Körner, 2003b; Li et al., 2008a).Körner (2003a)和李迈和和Kräuchi (2005)将这些假说总结为5类, 即环境胁迫假说、干扰假说、更新繁殖障碍假说、碳平衡失调假说和生长限制假说, 并认为“碳平衡失调”或“生长限制”假说最有可能在全球尺度上解释高山林线现象.“碳平衡失调”假说(Stevens & Fox, 1991)认为: 低温、干旱、生长季节较短及其他环境胁迫因素, 使植物的碳吸收与碳消耗关系失调, 导致碳供应不足, 进而引起高山林线的形成.“生长限制”假说(Körner., 1998; Li et al., 2002)的前提条件是碳供应充足(即不存在“碳平衡失调”问题), 低温导致充足的碳不能被生长所利用, 从而使生长受限, 导致高山林线的形成.但是, 对于高山林线的形成是由于碳供应不足还是低温导致树木生长受限, 至今仍然没有明确的结论(Shi et al., 2006; Li et al., 2008a, 2008b). ...
... ; Li et al., 2002)的前提条件是碳供应充足(即不存在“碳平衡失调”问题), 低温导致充足的碳不能被生长所利用, 从而使生长受限, 导致高山林线的形成.但是, 对于高山林线的形成是由于碳供应不足还是低温导致树木生长受限, 至今仍然没有明确的结论(Shi et al., 2006; Li et al., 2008a, 2008b). ...
... 生长于低温环境下的高山林线树木既需要足够的碳水化合物维持其生长, 也需要充足的可溶性碳水化合物来提高其在低温等不利环境下的生存能力(Körner, 2003b; 李迈和和Kräuchi, 2005).评价植物碳供应状态的一种可行方法是分析植物体内非结构性碳库的大小(Shi et al., 2006; Li et al., 2008a, 2008b).非结构性碳水化合物(non-structural carbohydrate, NSC)是树木休眠后恢复生长的主要碳供应者, 并能在植物光合作用不足时起到缓冲作用, 因此在很大程度上影响着植株的生长及对环境的响应(Li et al., 2001, 2002; Hoch & Körner, 2003; Körner, 2003b).植物组织中NSC含量(主要是可溶性糖和淀粉)的大小可看作是植物碳同化与碳消耗(即供应与需求)之间平衡关系的结果.树木组织中NSC含量随海拔升高而减少, 碳限制随海拔增加, 则意味着林线附近树木碳供应不足(碳平衡失调假说).相反, 树木组织形成受到限制(生长限制假说), 由于树木生长的碳需求减小, 则NSC含量随海拔升高而增加(Li et al., 2008a). ...
... 在每个样点选取直立、健康、无损伤、非孤立、大小(年龄、树高、胸径)基本一致的白杄5株, 分别采集叶、茎干木质部、粗根(直径>5 mm)及细根(直径<2 mm)样品.叶片样本取自上坡方向未遮阴的一年生和两年生成熟枝条, 进行混合; 木质部取样采用直径为5 mm的生长锥, 在胸高处平行于斜坡的相对两侧分别钻取2个树芯, 每树芯截取外侧3 cm木质部混合成每木木质部样; 细根(直径<2 mm, 带皮)样品采用挖掘法, 在5-30 cm深度的土层获取; 在每株样树的基部, 即分枝等级最高的根处用生长锥钻取3个根芯作为粗根样品.所取样品统一编号后, 在野外立即放入冷藏箱中保存, 并于当日带回驻地用微波炉杀青90 s (Li et al., 2002), 根部样品需将泥土洗净.然后, 将所有样品放在65-70 ℃恒温箱中烘至恒质量(Li et al., 2008a), 备用. ...
... 植物组织中NSC含量被用于评价光合作用的碳获得(源)与生长和呼吸过程的碳需求(汇)之间的平衡(Hoch et al., 2002; Li et al., 2002, 2008a; Körner, 2003b; Shi et al., 2008).高山林线树木源-汇活动受到低温等严酷环境因子的影响, 而一种较为平衡的碳源-汇关系有利于林线树木在严酷环境下生存(Li et al., 2008b).本研究发现生长季末林线白杄与中低海拔NSC源-汇比较为稳定, 约为2.0 (表1), 表明秋季源-汇关系对海拔的适应, 以及同化产物的分配较为平衡, 林线树木碳源活动没有受到限制.然而, Li等(2008b)对冷杉(Abies fabri)等3个物种的NSC源-汇关系的研究表明, 林线与低海拔NSC源-汇比在4月没有差异, 而7月林线树木显著低于低海拔.这种结果的差异性可能是因为夏季林线树木碳汇活动较强, 碳源活动受限, 而秋季和冬季, 树木处于或即将进入休眠状态, 源-汇活动均较弱.这些结果也许可以进一步表明林线树木碳源活动并没有受限, 而是碳汇活动受限(生长受限). ...
全球高山林线研究现状与发展方向
2
2005
... 作为最明显的生态界限之一, 高山林线由于其特殊的结构、功能及对气候变化的高度敏感性, 已经成为全球气候变化研究的热点区域之一( Li et al., 2001, 2002).全球高山林线归根结底是一条低温界限, 其形成被认为是生长季低温影响了树木碳平衡, 从而限制树木海拔分布的结果(Körner, 1998, 2003a; Hoch & Körner, 2003; Körner & Paulsen, 2004; Shi et al., 2008).关于高山林线形成的原因已经争论了逾百年(Körner, 2003a), 并提出了多种假说(Körner, 2003b; Li et al., 2008a).Körner (2003a)和李迈和和Kräuchi (2005)将这些假说总结为5类, 即环境胁迫假说、干扰假说、更新繁殖障碍假说、碳平衡失调假说和生长限制假说, 并认为“碳平衡失调”或“生长限制”假说最有可能在全球尺度上解释高山林线现象.“碳平衡失调”假说(Stevens & Fox, 1991)认为: 低温、干旱、生长季节较短及其他环境胁迫因素, 使植物的碳吸收与碳消耗关系失调, 导致碳供应不足, 进而引起高山林线的形成.“生长限制”假说(Körner., 1998; Li et al., 2002)的前提条件是碳供应充足(即不存在“碳平衡失调”问题), 低温导致充足的碳不能被生长所利用, 从而使生长受限, 导致高山林线的形成.但是, 对于高山林线的形成是由于碳供应不足还是低温导致树木生长受限, 至今仍然没有明确的结论(Shi et al., 2006; Li et al., 2008a, 2008b). ...
... 生长于低温环境下的高山林线树木既需要足够的碳水化合物维持其生长, 也需要充足的可溶性碳水化合物来提高其在低温等不利环境下的生存能力(Körner, 2003b; 李迈和和Kräuchi, 2005).评价植物碳供应状态的一种可行方法是分析植物体内非结构性碳库的大小(Shi et al., 2006; Li et al., 2008a, 2008b).非结构性碳水化合物(non-structural carbohydrate, NSC)是树木休眠后恢复生长的主要碳供应者, 并能在植物光合作用不足时起到缓冲作用, 因此在很大程度上影响着植株的生长及对环境的响应(Li et al., 2001, 2002; Hoch & Körner, 2003; Körner, 2003b).植物组织中NSC含量(主要是可溶性糖和淀粉)的大小可看作是植物碳同化与碳消耗(即供应与需求)之间平衡关系的结果.树木组织中NSC含量随海拔升高而减少, 碳限制随海拔增加, 则意味着林线附近树木碳供应不足(碳平衡失调假说).相反, 树木组织形成受到限制(生长限制假说), 由于树木生长的碳需求减小, 则NSC含量随海拔升高而增加(Li et al., 2008a). ...
全球高山林线研究现状与发展方向
2
2005
... 作为最明显的生态界限之一, 高山林线由于其特殊的结构、功能及对气候变化的高度敏感性, 已经成为全球气候变化研究的热点区域之一( Li et al., 2001, 2002).全球高山林线归根结底是一条低温界限, 其形成被认为是生长季低温影响了树木碳平衡, 从而限制树木海拔分布的结果(Körner, 1998, 2003a; Hoch & Körner, 2003; Körner & Paulsen, 2004; Shi et al., 2008).关于高山林线形成的原因已经争论了逾百年(Körner, 2003a), 并提出了多种假说(Körner, 2003b; Li et al., 2008a).Körner (2003a)和李迈和和Kräuchi (2005)将这些假说总结为5类, 即环境胁迫假说、干扰假说、更新繁殖障碍假说、碳平衡失调假说和生长限制假说, 并认为“碳平衡失调”或“生长限制”假说最有可能在全球尺度上解释高山林线现象.“碳平衡失调”假说(Stevens & Fox, 1991)认为: 低温、干旱、生长季节较短及其他环境胁迫因素, 使植物的碳吸收与碳消耗关系失调, 导致碳供应不足, 进而引起高山林线的形成.“生长限制”假说(Körner., 1998; Li et al., 2002)的前提条件是碳供应充足(即不存在“碳平衡失调”问题), 低温导致充足的碳不能被生长所利用, 从而使生长受限, 导致高山林线的形成.但是, 对于高山林线的形成是由于碳供应不足还是低温导致树木生长受限, 至今仍然没有明确的结论(Shi et al., 2006; Li et al., 2008a, 2008b). ...
... 生长于低温环境下的高山林线树木既需要足够的碳水化合物维持其生长, 也需要充足的可溶性碳水化合物来提高其在低温等不利环境下的生存能力(Körner, 2003b; 李迈和和Kräuchi, 2005).评价植物碳供应状态的一种可行方法是分析植物体内非结构性碳库的大小(Shi et al., 2006; Li et al., 2008a, 2008b).非结构性碳水化合物(non-structural carbohydrate, NSC)是树木休眠后恢复生长的主要碳供应者, 并能在植物光合作用不足时起到缓冲作用, 因此在很大程度上影响着植株的生长及对环境的响应(Li et al., 2001, 2002; Hoch & Körner, 2003; Körner, 2003b).植物组织中NSC含量(主要是可溶性糖和淀粉)的大小可看作是植物碳同化与碳消耗(即供应与需求)之间平衡关系的结果.树木组织中NSC含量随海拔升高而减少, 碳限制随海拔增加, 则意味着林线附近树木碳供应不足(碳平衡失调假说).相反, 树木组织形成受到限制(生长限制假说), 由于树木生长的碳需求减小, 则NSC含量随海拔升高而增加(Li et al., 2008a). ...
Mobile carbohydrates in Himalayan treeline trees I. Evidence for carbon gain limitation but not for growth limitation.
6
2008a
... 作为最明显的生态界限之一, 高山林线由于其特殊的结构、功能及对气候变化的高度敏感性, 已经成为全球气候变化研究的热点区域之一( Li et al., 2001, 2002).全球高山林线归根结底是一条低温界限, 其形成被认为是生长季低温影响了树木碳平衡, 从而限制树木海拔分布的结果(Körner, 1998, 2003a; Hoch & Körner, 2003; Körner & Paulsen, 2004; Shi et al., 2008).关于高山林线形成的原因已经争论了逾百年(Körner, 2003a), 并提出了多种假说(Körner, 2003b; Li et al., 2008a).Körner (2003a)和李迈和和Kräuchi (2005)将这些假说总结为5类, 即环境胁迫假说、干扰假说、更新繁殖障碍假说、碳平衡失调假说和生长限制假说, 并认为“碳平衡失调”或“生长限制”假说最有可能在全球尺度上解释高山林线现象.“碳平衡失调”假说(Stevens & Fox, 1991)认为: 低温、干旱、生长季节较短及其他环境胁迫因素, 使植物的碳吸收与碳消耗关系失调, 导致碳供应不足, 进而引起高山林线的形成.“生长限制”假说(Körner., 1998; Li et al., 2002)的前提条件是碳供应充足(即不存在“碳平衡失调”问题), 低温导致充足的碳不能被生长所利用, 从而使生长受限, 导致高山林线的形成.但是, 对于高山林线的形成是由于碳供应不足还是低温导致树木生长受限, 至今仍然没有明确的结论(Shi et al., 2006; Li et al., 2008a, 2008b). ...
... ; Li et al., 2008a, 2008b). ...
... 生长于低温环境下的高山林线树木既需要足够的碳水化合物维持其生长, 也需要充足的可溶性碳水化合物来提高其在低温等不利环境下的生存能力(Körner, 2003b; 李迈和和Kräuchi, 2005).评价植物碳供应状态的一种可行方法是分析植物体内非结构性碳库的大小(Shi et al., 2006; Li et al., 2008a, 2008b).非结构性碳水化合物(non-structural carbohydrate, NSC)是树木休眠后恢复生长的主要碳供应者, 并能在植物光合作用不足时起到缓冲作用, 因此在很大程度上影响着植株的生长及对环境的响应(Li et al., 2001, 2002; Hoch & Körner, 2003; Körner, 2003b).植物组织中NSC含量(主要是可溶性糖和淀粉)的大小可看作是植物碳同化与碳消耗(即供应与需求)之间平衡关系的结果.树木组织中NSC含量随海拔升高而减少, 碳限制随海拔增加, 则意味着林线附近树木碳供应不足(碳平衡失调假说).相反, 树木组织形成受到限制(生长限制假说), 由于树木生长的碳需求减小, 则NSC含量随海拔升高而增加(Li et al., 2008a). ...
... 作为最明显的生态界限之一, 高山林线由于其特殊的结构、功能及对气候变化的高度敏感性, 已经成为全球气候变化研究的热点区域之一( Li et al., 2001, 2002).全球高山林线归根结底是一条低温界限, 其形成被认为是生长季低温影响了树木碳平衡, 从而限制树木海拔分布的结果(Körner, 1998, 2003a; Hoch & Körner, 2003; Körner & Paulsen, 2004; Shi et al., 2008).关于高山林线形成的原因已经争论了逾百年(Körner, 2003a), 并提出了多种假说(Körner, 2003b; Li et al., 2008a).Körner (2003a)和李迈和和Kräuchi (2005)将这些假说总结为5类, 即环境胁迫假说、干扰假说、更新繁殖障碍假说、碳平衡失调假说和生长限制假说, 并认为“碳平衡失调”或“生长限制”假说最有可能在全球尺度上解释高山林线现象.“碳平衡失调”假说(Stevens & Fox, 1991)认为: 低温、干旱、生长季节较短及其他环境胁迫因素, 使植物的碳吸收与碳消耗关系失调, 导致碳供应不足, 进而引起高山林线的形成.“生长限制”假说(Körner., 1998; Li et al., 2002)的前提条件是碳供应充足(即不存在“碳平衡失调”问题), 低温导致充足的碳不能被生长所利用, 从而使生长受限, 导致高山林线的形成.但是, 对于高山林线的形成是由于碳供应不足还是低温导致树木生长受限, 至今仍然没有明确的结论(Shi et al., 2006; Li et al., 2008a, 2008b). ...
Altering flux through the sucrose biosynthesis pathway in transgenic Arabidopsis thaliana modifies photosynthetic acclimation at low temperatures and the development of freezing tolerance
1
2003
... 高山林线寒冷环境下, 植物的可持续性很大程度上取决于多年生器官中可溶性糖的积累(Morin et al., 2007).可溶性糖通过提高组织细胞液渗透压, 防止细胞内结冰, 保护植物顺利度过寒冷季节(Molina-Montenegro et al., 2012).本研究中, 林线树木可溶性糖含量高于低海拔树木(图1; 表2), 与周永斌等(2009)和Li等(2008a)的研究结果一致.其他一些研究显示, 林线树木叶片可溶性糖和低海拔没有差异(Hoch & Körner, 2003, 2005; Shi et al., 2006).这些研究结果表明, 在低温等严酷的环境下可溶性糖的有效性并没有限制林线树木的持续发展.Yu等(2014)认为这可能是因为生长季末期树木组织中可溶性糖含量水平与冬季树木存活密切相关, 而较高的可溶性糖含量是植物顺利度过寒冷而漫长的冬季的一种保护策略.同时, 植物组织的糖/淀粉比值也可能反映这种保护策略, 因为可溶性糖与淀粉的相互转化被公认为是一种有效的植物抗逆胁迫机制, 而它们的比例关系在高山植物抵御低温胁迫维持持续性中起着重要的作用(潘庆民等, 2002; 施征等, 2011).Li等(2008a)提出生长在高海拔的树木不仅依赖于总的NSC含量, 而且需要足够高的糖/淀粉比值顺利越冬.以往的研究发现生长在23 ℃的拟南芥(Arabidopsis thaliana)叶片的糖/淀粉比例低于生长在5 ℃下的叶片(Strand et al., 2003), 树木休眠期(10月至翌年4月)枝和茎干中的可溶性糖/淀粉比例(2.78-3.72)高于生长季(1.14-1.56)(Zhu et al., 2012), 本研究也发现林线树木组织可溶性糖/淀粉比值随海拔升高呈增大趋势(表3).这表明糖/淀粉比例可能随温度降低而升高, 以及植物中糖/淀粉比例对环境条件的一种积极的适应性响应, 即树木生长的环境越寒冷, 树木组织中表现出越明显的保护策略.这种结果也可能暗示林线区域的树木更多地受到生长限制. ...
Formation mechanisms of the alpine Erman’s birch (Betula ermanii) treeline on Changbai Mountain in Northeast China.
1
2014
... 以该地区高山林线树种白杄为研究对象, 根据白杄林在芦芽山北坡的分布特征, 在林线(2740 m)、中海拔(2400 m)和低海拔(2040 m)各设置一个样地, 进行采样.研究表明, NSC在生长季末期较稳定, 在NSC大量消耗的时期后, 能更好地反映碳源和碳汇之间的净平衡, 是最适合进行海拔对照研究的时期(Hoch et al., 2002; Yu et al., 2014), 因此采样均在生长季末(2013年9月15日)进行.为了减小光照和针叶表面温度的差异对碳水化合物的影响, 每次采样均在中午进行(Li et al., 2008b). ...
 , 王明昌
, 王明昌

{kind=link}
{kind=link}