Aims Soil microbes play a key role in the biogeochemical cycling in terrestrial ecosystems and are important for the nutrient recovery of degraded soils due to disturbances. However, dynamics in soil microbial biomass during the development of the secondary forest after logging are little known. Our objectives were to examine the temporal dynamics and influencing factors of soil microbial biomass carbon content (Cmic) and nitrogen content (Nmic) along a temperate forest logging chronosequence.Methods The logging chronosequence included four sites with 0-year, 10-year, 25-year, and 56-year sites since clear cutting of a deciduous broadleaved forest and was established in 2014 in the Mao’ershan Forest Ecosystem Research Station, Northeast China. The Cmic and Nmic at all the sites were measured monthly during the growing season (from April to October) with the chloroform fumigation extraction method; the soil dissolved organic carbon content (Cdis), total nitrogen content (Ndis), soil water content and temperature were simultaneously measured. Important findings (1) There were significant differences in soil microbial biomass among the four sites: the means of Cmic at the 56-year and 0-year sites were significantly higher than those at the 25-year and 10-year sites; the means of Nmic at the 0-year and 56-year sites were significantly higher than those at the 10-year site, while the 25-year site had intermediate Nmic; The Cmic/Nmic ratios at the 56-year and 10-year sites were significantly higher than those at the 25-year and 0-year sites. (2) The Cmic and Nmic at the 0-year site tended to decrease at the end of the growing season compared to earlier times, while those at the rest sites showed an increasing trend or no significant change. Soil microbial biomass among the 10-year, 25-year, and 56-year sites differed at the early growing season, and its amplitude of variations decreased as the stand age increased. The Cmic/Nmic ratios at all sites showed a “W-shaped” seasonal pattern. (3) The main influencing factors of the seasonality of soil microbial biomass varied with the stand ages: they switched from soil water content at the 0-year and 10-year sites to the soil dissolved nutrients contents at the 10-year, 25-year, and 56-year sites. The seasonality of Cmic/Nmic ratios at the 0-year site was mainly influenced by soil temperature and Cdis, while those at the other three sites were driven by the Cdis/Ndisratio. It was concluded that with the forest development after clear cutting, the characteristics of vegetation and soil have been changing, inducing increased soil microbial biomass and thereby improved soil nutrient regime, which reflected strong links between aboveground changes in vegetation and belowground dynamics in soil microbes.
WANGXin-Qi, HANYi, WANGChuan-Kuan. Soil microbial biomass and its seasonality in deciduous broadleaved forests with different stand ages in the Mao’ershan region, Northeast China. Chinese Journal of Plant Ecology, 2017, 41(6): 597-609 https://doi.org/10.17521/cjpe.2017.0011
自20世纪初期以来, 中国东北地区原始森林遭到大规模的工业采伐, 现存森林类型主要为采伐干扰后形成的落叶阔叶次生林(Wang et al., 2006), 这为探索次生林发展过程中土壤微生物变化情况提供了研究基础。本研究通过空间代替时间的年龄系列法(Walker et al., 2010), 选取相近立地条件和干扰历史相似的3个林龄(10年, 25年, 56年)的落叶阔叶林以及56年林分采伐后的采伐迹地, 构成本地区落叶阔叶林的林龄序列样地, 旨在探讨在次生林发展过程中落叶阔叶林土壤微生物生物量的林龄变化和季节动态格局及其影响因子。
1 研究方法
1.1 研究区自然概况和试验设计
研究地设在黑龙江帽儿山森林生态系统国家野外科学观测研究站(45.40° N, 127.67° E)。气候为明显的大陆性温带季风气候, 年平均气温2.8 ℃, 年降水量629 mm。平均海拔400 m, 平均坡度10°-15°。地带性土壤为暗棕色森林土壤。植被是经干扰后形成的天然次生林及人工林。
在相同气候, 相似土壤基质、海拔、地形条件下, 选取阔叶红松原始林经采伐后天然更新形成的林龄分别为10年、25年和56年的落叶阔叶林(表1), 每个林龄的林分各设置3块面积为20 m × 30 m 的重复样地。另外, 在56年林分随机设置3块面积为10 m × 30 m的皆伐迹地样地, 作为0年的采伐迹地。为了尽可能减少采伐作业对地被物和土壤的影响, 采伐作业在冰雪覆盖、土壤冻结的2014年冬季(3月)进行。
Table 1
表1
表1 样地基本特征(平均值±标准偏差, n = 3)
Table 1 Characteristics of the sampled plots (mean ± SD, n = 3)
林龄 Site age (a)
坡度 Slope (°)
林分密度 Site density (trees•hm-2)
胸高断面积 Basal area (m2•hm-2)
平均胸径 Mean DBH (cm)
林分组成 Site composition
0-20 cm土壤pH值 Soil pH value at 0-20 cm depth
0
18
0
0
0
未评估 Not assessed
4.70 ± 0.16
10
15
6 200 ± 2 300
19.51 ± 2.40
5.0 ± 0.5
10BP+PU+PA+AM
4.83 ± 0.21
25
15
6 028 ± 804
25.62 ± 2.30
12.2 ± 0.4
5BP3PD1UJ1FM+AM-PA-QM
4.85 ± 0.10
56
18
1 833 ± 617
28.77 ± 4.12
26.8 ± 1.3
3BP2AM1UJ1JM1FM 1PD1TM+TA+PA-QM
4.45 ± 0.30
All sites are located at the middle slope. The site composition: AM, Acer mono; BP, Betula platyphylla; FM, Fraxinus mandschurica; JM, Juglans mandshurica; PA, Phellodendron amurense; PD, Populus davidiana; PU, Populus ussuriensis; QM, Quercus mogolica; TA, Tilia amurensis; TM, Tilia mandshurica; UJ, Ulmus davidiana var. japonica. Numbers in front of the tree species represent the tenths of the basal area of this tree species out of the whole area of the plot; “+” and “-” in front of the tree species mean that the percentage is <5% and <2%, respectively. DBH, diameter at breast height.所有样地均设在中坡。林分组成中, 树种AM、BP、FM、JM、PA、PD、PU、QM、TA、TM和UJ分别表示色木槭、白桦、水曲柳、胡桃楸、黄檗、山杨、大青杨、蒙古栎、紫椴、糠椴和春榆, 树种前面的数字表示该树种胸高断面积所占的成数, 树种前面的“+”和“-”分别表示组成比例<5%和<2%。
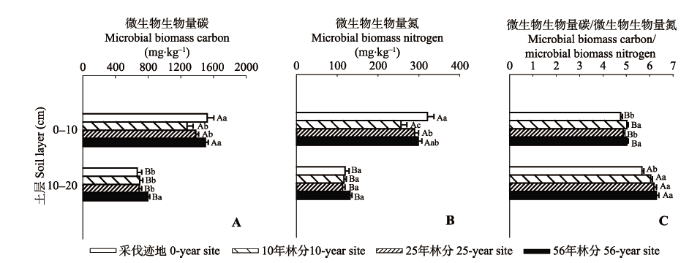
Fig. 1 Comparisons of the vertical changes in the means of soil microbial biomass carbon content (A), nitrogen content (B) and carbon and nitrogen ratio (C) at different sites during the growing season (mean ± SE). Different capital letters of the same site indicate significant differences between soil layers, and different lowercase letters of the same soil layer indicate significant differences among sites, while the same letter indicates no significant difference.
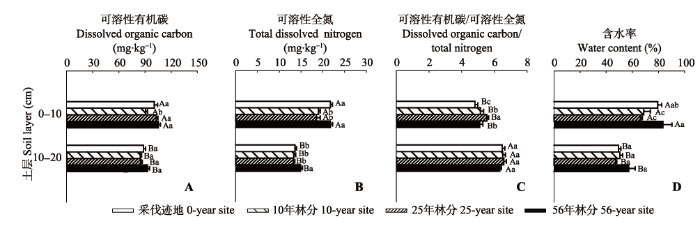
Fig. 2 Comparisons of the vertical changes in the means of soil dissolved organic carbon content (A), total nitrogen content (B), dissolved organic carbon and nitrogen ratio (C), and water content (D) among the four sites during the growing season (mean ± SE). Different capital letters of the same site indicate significant differences between soil layers, and different lowercase letters of the same soil layer indicate significant differences among sites, while the same letter indicates no significant difference.
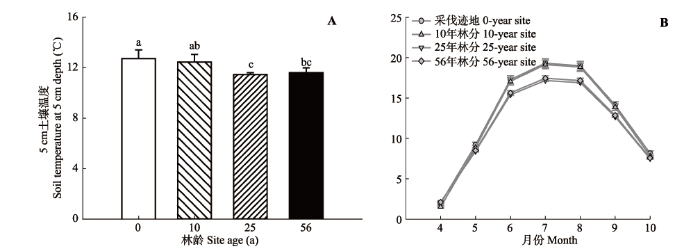
Fig. 3 Comparisons of the means of soil temperature at 5 cm depth during the growing season among sites and their seasonal dynamics (mean ± SE). Different lowercase letters indicate significant differences among sites.
Fig. 4 Seasonal dynamics in soil microbial biomass carbon content, nitrogen content and microbial biomass carbon and nitrogen ratio at different sites (mean ± SE).
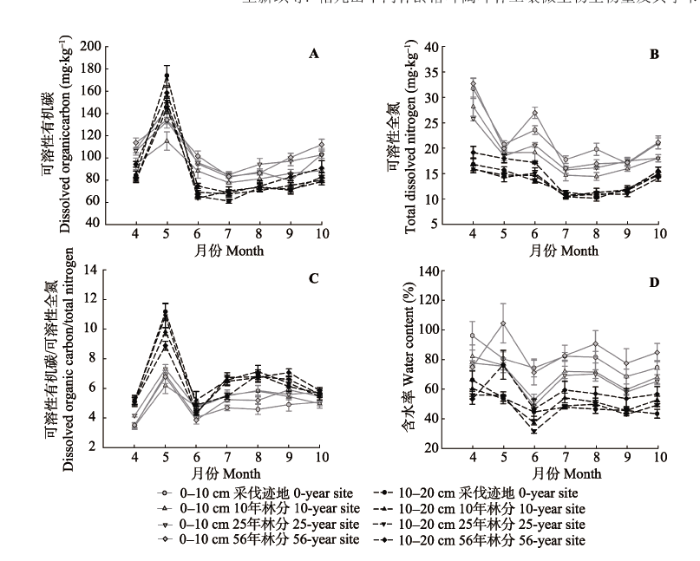
Fig. 5 Seasonal dynamics in soil dissolved organic carbon content (A), total nitrogen content (B), dissolved organic carbon and nitrogen ratio (C), and water content (D) at different sites (mean ± SE).
4个林龄林分Cmic/Nmic在生长季均呈现“W”形的变化格局(图4C), 表明土壤微生物群落结构能够随季节变化而转变(Björk et al., 2008; Smith et al., 2015)。4个林龄林分Cmic/Nmic在生长季前期显著下降, 这可能是由于5月温度适宜, 加之土壤完全解冻后液态水中的养分为微生物提供了丰富的分解底物, 更有利于细菌的生长和繁殖(Kaiser et al., 2014); 生长季中期, Cmic/Nmic维持在较低水平, 到后期显著上升, 这可能是由于真菌更适宜在温度较低的环境下生存(丁爽和王传宽, 2009; Robert et al., 2010; Kaiser et al., 2011)。除采伐迹地外, 其他3个林龄林分Cmic/Nmic均与Cdis/Ndis变化显著负相关(表3), 这与Song等(2015)的研究结果一致。说明随着季节更替, 地上植被通过改变土壤养分状况间接调节微生物群落结构的转变(Kaiser et al., 2014)。而采伐迹地Cmic/Nmic受T5和Cdis的共同影响(表3), 这表明植被的减少导致土壤微生物群落转变对温度的敏感性提高, 采伐干扰改变了土壤微生物群落转变的驱动因子。而Saynes等(2005)在热带次生林的研究表明, Cmic/Nmic的季节动态主要受降水的影响, 可能是由于该地区大部分时间内土壤较干旱, 因此降水后土壤环境剧变对微生物群落产生了明显的影响; 而在本研究区的生长季, 土壤一直比较湿润, 因而水分对微生物群落的限制作用不显著。
Seasonal dynamics of soil microorganisms and soil nutrients in fast-growing
Populus plantation forests of different ages in Yili, Xinjiang, China. Chinese Journal of Plant Ecology, 35, 389-401. (in Chinese with English abstract)[安然, 龚吉蕊, 尤鑫, 葛之葳, 段庆伟, 晏欣 (2011).
Development of soil microbial properties in topsoil layer during spontaneous succession in heaps after brown coal mining in relation to humus microstructure development.
Litter decomposition processes in the pure birch (
Betula platyphlla) forest and the birch and poplar(Populus davidiana) mixed forest. Acta Ecologica Sinica, 27, 1782-1790. (in Chinese with English abstract)[李雪峰, 张岩, 牛丽君, 韩士杰 (2007).
Soil-surface carbon dioxide efflux and microbial biomass in relation to tree density 13 years after a stand replacing fire in a lodgepole pine ecosystem.
Changes in labile soil organic matter fractions following land use change from monocropping to poplar-based agroforestry systems in a semiarid region of Northeast China.
Environmental Monitoring and Assessment, 184, 6845-6853.
Successional and seasonal variations in soil and litter microbial community structure and function during tropical post-agricultural forest regeneration: A multi-year study.
SternerRW,Elser JJ(2002).Ecological Stoichiometry: The Biology of Elements from Molecules to the Biosphere. Princeton University Press, Princeton, USA.
Forest succession on abandoned arable soils in European Russia—Impacts on microbial biomass, fungal-bacterial ratio, and basal CO2 respiration activity.
... 以往关于林龄对土壤微生物生物量影响的研究多集中于人工林(Chodak et al., 2009; 安然等, 2011; Mao et al., 2012; Yuan & Yue, 2012; Wen et al., 2014), 关于天然次生林的报道多集中在热带和亚热带地区(Saynes et al., 2005; Smith et al., 2015).虽然温带次生林在生态环境建设中起着举足轻重的作用, 但是温带次生林发展过程中土壤微生物生物量的变化情况及其影响因子尚不明确(Jia et al., 2005).以往研究表明: 随着林龄增加, 土壤微生物生物量可能逐渐增加(Chodak et al., 2009; Susyan et al., 2011; Mao et al., 2012)、减小(杨凯等, 2009; 安然等, 2011), 或达到最高值之后降低并保持稳定(Jia et al., 2005).这种结果的不一致可能是由研究对象以及林龄跨度不同造成的.随着次生林发展进程, 林分组成、生物量累积和分配等产生变化, 从而可能通过改变林内土壤温度、水分、凋落物质量等条件对土壤微生物产生间接影响(Litton et al., 2003; Jia et al., 2005), 也可能通过土壤酸化、有机质累积等过程直接影响微生物的生长繁殖及群落结构(Frouz & Nováková, 2005; Trap et al., 2011; Smith et al., 2015).由于不同林龄次生林土壤微生物的研究报道有限, 次生林发展过程中土壤微生物生物量对植被及环境因子的改变如何响应尚不明确. ...
... ; 安然等, 2011), 或达到最高值之后降低并保持稳定(Jia et al., 2005).这种结果的不一致可能是由研究对象以及林龄跨度不同造成的.随着次生林发展进程, 林分组成、生物量累积和分配等产生变化, 从而可能通过改变林内土壤温度、水分、凋落物质量等条件对土壤微生物产生间接影响(Litton et al., 2003; Jia et al., 2005), 也可能通过土壤酸化、有机质累积等过程直接影响微生物的生长繁殖及群落结构(Frouz & Nováková, 2005; Trap et al., 2011; Smith et al., 2015).由于不同林龄次生林土壤微生物的研究报道有限, 次生林发展过程中土壤微生物生物量对植被及环境因子的改变如何响应尚不明确. ...
... 由于真菌C/N高于细菌(Sterner & Elser, 2002), 因此通常将Cmic/Nmic作为衡量微生物群落变化的指标, 即Cmic/Nmic越高, 真菌生物量越高(Zhou & Wang, 2015).虽然采伐迹地Cmic、Nmic生长季均值与56年林分无显著差异, 但采伐迹地Cmic/Nmic显著小于56年林分(图1C).说明采伐干扰后土壤微生物群落发生转变, 即细菌所占比例增高.这是由于采伐干扰后死亡根系分解及植物吸收减少等导致可利用底物含量提高(Guo et al., 2004), 使得快速增长的细菌成为优势群落(Kaiser et al., 2014).虽然本研究中Cmic/Nmic与土壤pH值没有显著的负相关关系(表2), 但56年林分土壤pH值最低, 而两个土层的Cmic/Nmic最高.这可能是由于56年林分凋落物层较厚造成土壤酸化(表1), 而真菌更耐受酸性土壤环境, 导致微生物群落真菌比例升高(Trap et al., 2011).随着土层加深, Cmic/Nmic增高, 这与以往研究结果(Fierer et al., 2003; 安然等, 2011)一致, 主要是因为土壤表层养分限制较低, 更适宜细菌生存.以上结果说明: 在次生林发展过程中, 土壤微生物能够通过改变群落结构来适应土壤养分、酸度等性质的改变. ...
Ecology and Management of Forest Soils. John Wiley and Sons
Temporal variation in soil microbial communities in Alpine tundra.
1
2008
... 4个林龄林分Cmic/Nmic在生长季均呈现“W”形的变化格局(图4C), 表明土壤微生物群落结构能够随季节变化而转变(Björk et al., 2008; Smith et al., 2015).4个林龄林分Cmic/Nmic在生长季前期显著下降, 这可能是由于5月温度适宜, 加之土壤完全解冻后液态水中的养分为微生物提供了丰富的分解底物, 更有利于细菌的生长和繁殖(Kaiser et al., 2014); 生长季中期, Cmic/Nmic维持在较低水平, 到后期显著上升, 这可能是由于真菌更适宜在温度较低的环境下生存(丁爽和王传宽, 2009; Robert et al., 2010; Kaiser et al., 2011).除采伐迹地外, 其他3个林龄林分Cmic/Nmic均与Cdis/Ndis变化显著负相关(表3), 这与Song等(2015)的研究结果一致.说明随着季节更替, 地上植被通过改变土壤养分状况间接调节微生物群落结构的转变(Kaiser et al., 2014).而采伐迹地Cmic/Nmic受T5和Cdis的共同影响(表3), 这表明植被的减少导致土壤微生物群落转变对温度的敏感性提高, 采伐干扰改变了土壤微生物群落转变的驱动因子.而Saynes等(2005)在热带次生林的研究表明, Cmic/Nmic的季节动态主要受降水的影响, 可能是由于该地区大部分时间内土壤较干旱, 因此降水后土壤环境剧变对微生物群落产生了明显的影响; 而在本研究区的生长季, 土壤一直比较湿润, 因而水分对微生物群落的限制作用不显著. ...
Principles of Terrestrial Ecosystem Ecology. 2nd edn. Springer
Development of microbial properties in a chronosequence of sandy mine soils.
2
2009
... 以往关于林龄对土壤微生物生物量影响的研究多集中于人工林(Chodak et al., 2009; 安然等, 2011; Mao et al., 2012; Yuan & Yue, 2012; Wen et al., 2014), 关于天然次生林的报道多集中在热带和亚热带地区(Saynes et al., 2005; Smith et al., 2015).虽然温带次生林在生态环境建设中起着举足轻重的作用, 但是温带次生林发展过程中土壤微生物生物量的变化情况及其影响因子尚不明确(Jia et al., 2005).以往研究表明: 随着林龄增加, 土壤微生物生物量可能逐渐增加(Chodak et al., 2009; Susyan et al., 2011; Mao et al., 2012)、减小(杨凯等, 2009; 安然等, 2011), 或达到最高值之后降低并保持稳定(Jia et al., 2005).这种结果的不一致可能是由研究对象以及林龄跨度不同造成的.随着次生林发展进程, 林分组成、生物量累积和分配等产生变化, 从而可能通过改变林内土壤温度、水分、凋落物质量等条件对土壤微生物产生间接影响(Litton et al., 2003; Jia et al., 2005), 也可能通过土壤酸化、有机质累积等过程直接影响微生物的生长繁殖及群落结构(Frouz & Nováková, 2005; Trap et al., 2011; Smith et al., 2015).由于不同林龄次生林土壤微生物的研究报道有限, 次生林发展过程中土壤微生物生物量对植被及环境因子的改变如何响应尚不明确. ...
... ).以往研究表明: 随着林龄增加, 土壤微生物生物量可能逐渐增加(Chodak et al., 2009; Susyan et al., 2011; Mao et al., 2012)、减小(杨凯等, 2009; 安然等, 2011), 或达到最高值之后降低并保持稳定(Jia et al., 2005).这种结果的不一致可能是由研究对象以及林龄跨度不同造成的.随着次生林发展进程, 林分组成、生物量累积和分配等产生变化, 从而可能通过改变林内土壤温度、水分、凋落物质量等条件对土壤微生物产生间接影响(Litton et al., 2003; Jia et al., 2005), 也可能通过土壤酸化、有机质累积等过程直接影响微生物的生长繁殖及群落结构(Frouz & Nováková, 2005; Trap et al., 2011; Smith et al., 2015).由于不同林龄次生林土壤微生物的研究报道有限, 次生林发展过程中土壤微生物生物量对植被及环境因子的改变如何响应尚不明确. ...
... 4个林龄林分Cmic/Nmic在生长季均呈现“W”形的变化格局(图4C), 表明土壤微生物群落结构能够随季节变化而转变(Björk et al., 2008; Smith et al., 2015).4个林龄林分Cmic/Nmic在生长季前期显著下降, 这可能是由于5月温度适宜, 加之土壤完全解冻后液态水中的养分为微生物提供了丰富的分解底物, 更有利于细菌的生长和繁殖(Kaiser et al., 2014); 生长季中期, Cmic/Nmic维持在较低水平, 到后期显著上升, 这可能是由于真菌更适宜在温度较低的环境下生存(丁爽和王传宽, 2009; Robert et al., 2010; Kaiser et al., 2011).除采伐迹地外, 其他3个林龄林分Cmic/Nmic均与Cdis/Ndis变化显著负相关(表3), 这与Song等(2015)的研究结果一致.说明随着季节更替, 地上植被通过改变土壤养分状况间接调节微生物群落结构的转变(Kaiser et al., 2014).而采伐迹地Cmic/Nmic受T5和Cdis的共同影响(表3), 这表明植被的减少导致土壤微生物群落转变对温度的敏感性提高, 采伐干扰改变了土壤微生物群落转变的驱动因子.而Saynes等(2005)在热带次生林的研究表明, Cmic/Nmic的季节动态主要受降水的影响, 可能是由于该地区大部分时间内土壤较干旱, 因此降水后土壤环境剧变对微生物群落产生了明显的影响; 而在本研究区的生长季, 土壤一直比较湿润, 因而水分对微生物群落的限制作用不显著. ...
Inter-annual and seasonal dynamics of soil microbial biomass and nutrients in wet and dry low-Arctic sedge meadows.
3
2013
... 土壤微生物生物量的变化受土壤养分、水分、温度的共同影响, 而这些相关因子通常随季节转变而变化, 因此土壤微生物生物量通常呈现一定的季节动态(刘爽和王传宽, 2010; Kaiser et al., 2011; Wen et al., 2014).以往关于土壤微生物生物量季节变化的研究多针对不同森林类型(刘爽和王传宽, 2010; 刘纯等, 2014)、不同土地利用方式(Pandey et al., 2010)或不同林龄人工林(杨凯等, 2009), 而对不同林龄天然次生林土壤微生物生物量的季节变化尚不明确(Smith et al., 2015).不同林龄次生林植被活动(吸收养分、水分)及土壤养分、温湿度可能对季节变化产生不同的响应, 从而对微生物生物量的季节动态产生不同程度的影响.土壤可溶性养分较易被微生物吸收利用, 并且对季节变化的响应十分敏感(Edwards et al., 2006; Edwards & Jefferies, 2013; Freppaz et al., 2014); 土壤温度、湿度等环境因子的改变也调节着微生物的生长和繁殖(刘爽和王传宽, 2010; 刘纯等, 2014).因此, 探索不同林龄次生林土壤可溶性养分、土壤温湿度的季节变化格局及其与土壤微生物生物量的关系, 有利于充分理解次生林不同发展阶段土壤微生物生物量的驱动机制. ...
Variations in microbial community composition through two soil depth profiles.
1
2003
... 由于真菌C/N高于细菌(Sterner & Elser, 2002), 因此通常将Cmic/Nmic作为衡量微生物群落变化的指标, 即Cmic/Nmic越高, 真菌生物量越高(Zhou & Wang, 2015).虽然采伐迹地Cmic、Nmic生长季均值与56年林分无显著差异, 但采伐迹地Cmic/Nmic显著小于56年林分(图1C).说明采伐干扰后土壤微生物群落发生转变, 即细菌所占比例增高.这是由于采伐干扰后死亡根系分解及植物吸收减少等导致可利用底物含量提高(Guo et al., 2004), 使得快速增长的细菌成为优势群落(Kaiser et al., 2014).虽然本研究中Cmic/Nmic与土壤pH值没有显著的负相关关系(表2), 但56年林分土壤pH值最低, 而两个土层的Cmic/Nmic最高.这可能是由于56年林分凋落物层较厚造成土壤酸化(表1), 而真菌更耐受酸性土壤环境, 导致微生物群落真菌比例升高(Trap et al., 2011).随着土层加深, Cmic/Nmic增高, 这与以往研究结果(Fierer et al., 2003; 安然等, 2011)一致, 主要是因为土壤表层养分限制较低, 更适宜细菌生存.以上结果说明: 在次生林发展过程中, 土壤微生物能够通过改变群落结构来适应土壤养分、酸度等性质的改变. ...
Soil C and N storage and microbial biomass in US southern pine forests: Influence of forest management.
1
2015
... 本研究表明不同林龄落叶阔叶林土壤微生物生物量差异显著, 表现为随林龄增加先降低之后逐渐升高(图1).采伐迹地表层Cmic、Nmic与56年林分在整个生长季的均值差异不显著, 可能是由于采伐后残留在土壤里的根系死亡分解, 短期内为表层微生物提供了充足的可利用养分(Guo et al., 2004; Mäkiranta et al., 2012); 此外, 采伐后地上植被对养分的需求降低, 缓解了微生物与植物对养分的竞争(Kaiser et al., 2011), 因此短期维持了微生物生长和繁殖.但采伐迹地深层Cmic显著小于56年林分, 这主要可能是由于深层Cmic的主要限制因子为Ndis (表2), 而由于采伐干扰导致采伐迹地深层Ndis淋溶损失较多、Ndis含量较低, 因此抑制了微生物生长繁殖.采伐干扰10年后, 土壤微生物生物量显著降低, 之后随着林龄增长显著提高, 与Susyan等(2011)的研究结果一致.说明采伐的长期作用(土壤有机质降低、土壤紧实等)抑制了微生物的生长和繁殖(Foote et al., 2015), 但随着次生林的发展, 抑制作用能够逐渐解除.而黄土高原次生林发展过程中, 土壤微生物生物量在17年后达到最高值, 之后降低并保持稳定(Jia et al., 2005).造成这种差异的原因可能是黄土高原植被演替时间跨度较小, 但东北温带地区落叶阔叶林发展到阔叶红松林顶级群落需要200年左右.本研究中林龄较小, 只是森林演替进程中的一部分, 微生物生物量可能尚未达到最高值.因此, 为充分恢复东北温带地区退化土壤的性质及养分, 需要对天然次生林进行长期保护. ...
Winter-spring transition induces changes in nutrients and microbial biomass in mid-alpine forest soils.
1
2014
... 土壤微生物生物量的变化受土壤养分、水分、温度的共同影响, 而这些相关因子通常随季节转变而变化, 因此土壤微生物生物量通常呈现一定的季节动态(刘爽和王传宽, 2010; Kaiser et al., 2011; Wen et al., 2014).以往关于土壤微生物生物量季节变化的研究多针对不同森林类型(刘爽和王传宽, 2010; 刘纯等, 2014)、不同土地利用方式(Pandey et al., 2010)或不同林龄人工林(杨凯等, 2009), 而对不同林龄天然次生林土壤微生物生物量的季节变化尚不明确(Smith et al., 2015).不同林龄次生林植被活动(吸收养分、水分)及土壤养分、温湿度可能对季节变化产生不同的响应, 从而对微生物生物量的季节动态产生不同程度的影响.土壤可溶性养分较易被微生物吸收利用, 并且对季节变化的响应十分敏感(Edwards et al., 2006; Edwards & Jefferies, 2013; Freppaz et al., 2014); 土壤温度、湿度等环境因子的改变也调节着微生物的生长和繁殖(刘爽和王传宽, 2010; 刘纯等, 2014).因此, 探索不同林龄次生林土壤可溶性养分、土壤温湿度的季节变化格局及其与土壤微生物生物量的关系, 有利于充分理解次生林不同发展阶段土壤微生物生物量的驱动机制. ...
Development of soil microbial properties in topsoil layer during spontaneous succession in heaps after brown coal mining in relation to humus microstructure development.
1
2005
... 以往关于林龄对土壤微生物生物量影响的研究多集中于人工林(Chodak et al., 2009; 安然等, 2011; Mao et al., 2012; Yuan & Yue, 2012; Wen et al., 2014), 关于天然次生林的报道多集中在热带和亚热带地区(Saynes et al., 2005; Smith et al., 2015).虽然温带次生林在生态环境建设中起着举足轻重的作用, 但是温带次生林发展过程中土壤微生物生物量的变化情况及其影响因子尚不明确(Jia et al., 2005).以往研究表明: 随着林龄增加, 土壤微生物生物量可能逐渐增加(Chodak et al., 2009; Susyan et al., 2011; Mao et al., 2012)、减小(杨凯等, 2009; 安然等, 2011), 或达到最高值之后降低并保持稳定(Jia et al., 2005).这种结果的不一致可能是由研究对象以及林龄跨度不同造成的.随着次生林发展进程, 林分组成、生物量累积和分配等产生变化, 从而可能通过改变林内土壤温度、水分、凋落物质量等条件对土壤微生物产生间接影响(Litton et al., 2003; Jia et al., 2005), 也可能通过土壤酸化、有机质累积等过程直接影响微生物的生长繁殖及群落结构(Frouz & Nováková, 2005; Trap et al., 2011; Smith et al., 2015).由于不同林龄次生林土壤微生物的研究报道有限, 次生林发展过程中土壤微生物生物量对植被及环境因子的改变如何响应尚不明确. ...
Neotropical secondary forest succession: Changes in structural and functional characteristics.
Spatio-temporal patterns of soil available nutrients following experimental disturbance in a pine forest.
2
2004
... 本研究表明不同林龄落叶阔叶林土壤微生物生物量差异显著, 表现为随林龄增加先降低之后逐渐升高(图1).采伐迹地表层Cmic、Nmic与56年林分在整个生长季的均值差异不显著, 可能是由于采伐后残留在土壤里的根系死亡分解, 短期内为表层微生物提供了充足的可利用养分(Guo et al., 2004; Mäkiranta et al., 2012); 此外, 采伐后地上植被对养分的需求降低, 缓解了微生物与植物对养分的竞争(Kaiser et al., 2011), 因此短期维持了微生物生长和繁殖.但采伐迹地深层Cmic显著小于56年林分, 这主要可能是由于深层Cmic的主要限制因子为Ndis (表2), 而由于采伐干扰导致采伐迹地深层Ndis淋溶损失较多、Ndis含量较低, 因此抑制了微生物生长繁殖.采伐干扰10年后, 土壤微生物生物量显著降低, 之后随着林龄增长显著提高, 与Susyan等(2011)的研究结果一致.说明采伐的长期作用(土壤有机质降低、土壤紧实等)抑制了微生物的生长和繁殖(Foote et al., 2015), 但随着次生林的发展, 抑制作用能够逐渐解除.而黄土高原次生林发展过程中, 土壤微生物生物量在17年后达到最高值, 之后降低并保持稳定(Jia et al., 2005).造成这种差异的原因可能是黄土高原植被演替时间跨度较小, 但东北温带地区落叶阔叶林发展到阔叶红松林顶级群落需要200年左右.本研究中林龄较小, 只是森林演替进程中的一部分, 微生物生物量可能尚未达到最高值.因此, 为充分恢复东北温带地区退化土壤的性质及养分, 需要对天然次生林进行长期保护. ...
... 由于真菌C/N高于细菌(Sterner & Elser, 2002), 因此通常将Cmic/Nmic作为衡量微生物群落变化的指标, 即Cmic/Nmic越高, 真菌生物量越高(Zhou & Wang, 2015).虽然采伐迹地Cmic、Nmic生长季均值与56年林分无显著差异, 但采伐迹地Cmic/Nmic显著小于56年林分(图1C).说明采伐干扰后土壤微生物群落发生转变, 即细菌所占比例增高.这是由于采伐干扰后死亡根系分解及植物吸收减少等导致可利用底物含量提高(Guo et al., 2004), 使得快速增长的细菌成为优势群落(Kaiser et al., 2014).虽然本研究中Cmic/Nmic与土壤pH值没有显著的负相关关系(表2), 但56年林分土壤pH值最低, 而两个土层的Cmic/Nmic最高.这可能是由于56年林分凋落物层较厚造成土壤酸化(表1), 而真菌更耐受酸性土壤环境, 导致微生物群落真菌比例升高(Trap et al., 2011).随着土层加深, Cmic/Nmic增高, 这与以往研究结果(Fierer et al., 2003; 安然等, 2011)一致, 主要是因为土壤表层养分限制较低, 更适宜细菌生存.以上结果说明: 在次生林发展过程中, 土壤微生物能够通过改变群落结构来适应土壤养分、酸度等性质的改变. ...
Microbial biomass and nutrients in soil at the different stages of secondary forest succession in Ziwulin, northwest China.
7
2005
... 土壤微生物生物量是土壤养分中最活跃的部分, 不仅通过转化有机质为植物提供可利用养分, 调节陆地生态系统碳氮循环(Xu et al., 2013; Zhou & Wang, 2015), 而且在土壤养分的累积过程中扮演着重要的角色, 对于受到干扰后退化土壤的肥力恢复具有重要意义(Jia et al., 2005; Saynes et al., 2005; Spohn et al., 2016).经历农耕、采伐等人为干扰后形成的天然次生林是陆地生态系统的重要组成部分(Yang et al., 2011).充分理解天然次生林发展过程中土壤微生物生物量的变异及其影响因子, 对于退化土壤肥力的恢复、合理经营和保护次生林具有重要意义(Jia et al., 2005; Smith et al., 2015). ...
... ).充分理解天然次生林发展过程中土壤微生物生物量的变异及其影响因子, 对于退化土壤肥力的恢复、合理经营和保护次生林具有重要意义(Jia et al., 2005; Smith et al., 2015). ...
... 以往关于林龄对土壤微生物生物量影响的研究多集中于人工林(Chodak et al., 2009; 安然等, 2011; Mao et al., 2012; Yuan & Yue, 2012; Wen et al., 2014), 关于天然次生林的报道多集中在热带和亚热带地区(Saynes et al., 2005; Smith et al., 2015).虽然温带次生林在生态环境建设中起着举足轻重的作用, 但是温带次生林发展过程中土壤微生物生物量的变化情况及其影响因子尚不明确(Jia et al., 2005).以往研究表明: 随着林龄增加, 土壤微生物生物量可能逐渐增加(Chodak et al., 2009; Susyan et al., 2011; Mao et al., 2012)、减小(杨凯等, 2009; 安然等, 2011), 或达到最高值之后降低并保持稳定(Jia et al., 2005).这种结果的不一致可能是由研究对象以及林龄跨度不同造成的.随着次生林发展进程, 林分组成、生物量累积和分配等产生变化, 从而可能通过改变林内土壤温度、水分、凋落物质量等条件对土壤微生物产生间接影响(Litton et al., 2003; Jia et al., 2005), 也可能通过土壤酸化、有机质累积等过程直接影响微生物的生长繁殖及群落结构(Frouz & Nováková, 2005; Trap et al., 2011; Smith et al., 2015).由于不同林龄次生林土壤微生物的研究报道有限, 次生林发展过程中土壤微生物生物量对植被及环境因子的改变如何响应尚不明确. ...
... ), 或达到最高值之后降低并保持稳定(Jia et al., 2005).这种结果的不一致可能是由研究对象以及林龄跨度不同造成的.随着次生林发展进程, 林分组成、生物量累积和分配等产生变化, 从而可能通过改变林内土壤温度、水分、凋落物质量等条件对土壤微生物产生间接影响(Litton et al., 2003; Jia et al., 2005), 也可能通过土壤酸化、有机质累积等过程直接影响微生物的生长繁殖及群落结构(Frouz & Nováková, 2005; Trap et al., 2011; Smith et al., 2015).由于不同林龄次生林土壤微生物的研究报道有限, 次生林发展过程中土壤微生物生物量对植被及环境因子的改变如何响应尚不明确. ...
... ; Jia et al., 2005), 也可能通过土壤酸化、有机质累积等过程直接影响微生物的生长繁殖及群落结构(Frouz & Nováková, 2005; Trap et al., 2011; Smith et al., 2015).由于不同林龄次生林土壤微生物的研究报道有限, 次生林发展过程中土壤微生物生物量对植被及环境因子的改变如何响应尚不明确. ...
... 本研究表明不同林龄落叶阔叶林土壤微生物生物量差异显著, 表现为随林龄增加先降低之后逐渐升高(图1).采伐迹地表层Cmic、Nmic与56年林分在整个生长季的均值差异不显著, 可能是由于采伐后残留在土壤里的根系死亡分解, 短期内为表层微生物提供了充足的可利用养分(Guo et al., 2004; Mäkiranta et al., 2012); 此外, 采伐后地上植被对养分的需求降低, 缓解了微生物与植物对养分的竞争(Kaiser et al., 2011), 因此短期维持了微生物生长和繁殖.但采伐迹地深层Cmic显著小于56年林分, 这主要可能是由于深层Cmic的主要限制因子为Ndis (表2), 而由于采伐干扰导致采伐迹地深层Ndis淋溶损失较多、Ndis含量较低, 因此抑制了微生物生长繁殖.采伐干扰10年后, 土壤微生物生物量显著降低, 之后随着林龄增长显著提高, 与Susyan等(2011)的研究结果一致.说明采伐的长期作用(土壤有机质降低、土壤紧实等)抑制了微生物的生长和繁殖(Foote et al., 2015), 但随着次生林的发展, 抑制作用能够逐渐解除.而黄土高原次生林发展过程中, 土壤微生物生物量在17年后达到最高值, 之后降低并保持稳定(Jia et al., 2005).造成这种差异的原因可能是黄土高原植被演替时间跨度较小, 但东北温带地区落叶阔叶林发展到阔叶红松林顶级群落需要200年左右.本研究中林龄较小, 只是森林演替进程中的一部分, 微生物生物量可能尚未达到最高值.因此, 为充分恢复东北温带地区退化土壤的性质及养分, 需要对天然次生林进行长期保护. ...
Microbial community dynamics alleviate stoichiometric constraints during litter decay.
3
2014
... 由于真菌C/N高于细菌(Sterner & Elser, 2002), 因此通常将Cmic/Nmic作为衡量微生物群落变化的指标, 即Cmic/Nmic越高, 真菌生物量越高(Zhou & Wang, 2015).虽然采伐迹地Cmic、Nmic生长季均值与56年林分无显著差异, 但采伐迹地Cmic/Nmic显著小于56年林分(图1C).说明采伐干扰后土壤微生物群落发生转变, 即细菌所占比例增高.这是由于采伐干扰后死亡根系分解及植物吸收减少等导致可利用底物含量提高(Guo et al., 2004), 使得快速增长的细菌成为优势群落(Kaiser et al., 2014).虽然本研究中Cmic/Nmic与土壤pH值没有显著的负相关关系(表2), 但56年林分土壤pH值最低, 而两个土层的Cmic/Nmic最高.这可能是由于56年林分凋落物层较厚造成土壤酸化(表1), 而真菌更耐受酸性土壤环境, 导致微生物群落真菌比例升高(Trap et al., 2011).随着土层加深, Cmic/Nmic增高, 这与以往研究结果(Fierer et al., 2003; 安然等, 2011)一致, 主要是因为土壤表层养分限制较低, 更适宜细菌生存.以上结果说明: 在次生林发展过程中, 土壤微生物能够通过改变群落结构来适应土壤养分、酸度等性质的改变. ...
... 4个林龄林分Cmic/Nmic在生长季均呈现“W”形的变化格局(图4C), 表明土壤微生物群落结构能够随季节变化而转变(Björk et al., 2008; Smith et al., 2015).4个林龄林分Cmic/Nmic在生长季前期显著下降, 这可能是由于5月温度适宜, 加之土壤完全解冻后液态水中的养分为微生物提供了丰富的分解底物, 更有利于细菌的生长和繁殖(Kaiser et al., 2014); 生长季中期, Cmic/Nmic维持在较低水平, 到后期显著上升, 这可能是由于真菌更适宜在温度较低的环境下生存(丁爽和王传宽, 2009; Robert et al., 2010; Kaiser et al., 2011).除采伐迹地外, 其他3个林龄林分Cmic/Nmic均与Cdis/Ndis变化显著负相关(表3), 这与Song等(2015)的研究结果一致.说明随着季节更替, 地上植被通过改变土壤养分状况间接调节微生物群落结构的转变(Kaiser et al., 2014).而采伐迹地Cmic/Nmic受T5和Cdis的共同影响(表3), 这表明植被的减少导致土壤微生物群落转变对温度的敏感性提高, 采伐干扰改变了土壤微生物群落转变的驱动因子.而Saynes等(2005)在热带次生林的研究表明, Cmic/Nmic的季节动态主要受降水的影响, 可能是由于该地区大部分时间内土壤较干旱, 因此降水后土壤环境剧变对微生物群落产生了明显的影响; 而在本研究区的生长季, 土壤一直比较湿润, 因而水分对微生物群落的限制作用不显著. ...
... 4个林龄林分Cmic/Nmic在生长季均呈现“W”形的变化格局(图4C), 表明土壤微生物群落结构能够随季节变化而转变(Björk et al., 2008; Smith et al., 2015).4个林龄林分Cmic/Nmic在生长季前期显著下降, 这可能是由于5月温度适宜, 加之土壤完全解冻后液态水中的养分为微生物提供了丰富的分解底物, 更有利于细菌的生长和繁殖(Kaiser et al., 2014); 生长季中期, Cmic/Nmic维持在较低水平, 到后期显著上升, 这可能是由于真菌更适宜在温度较低的环境下生存(丁爽和王传宽, 2009; Robert et al., 2010; Kaiser et al., 2011).除采伐迹地外, 其他3个林龄林分Cmic/Nmic均与Cdis/Ndis变化显著负相关(表3), 这与Song等(2015)的研究结果一致.说明随着季节更替, 地上植被通过改变土壤养分状况间接调节微生物群落结构的转变(Kaiser et al., 2014).而采伐迹地Cmic/Nmic受T5和Cdis的共同影响(表3), 这表明植被的减少导致土壤微生物群落转变对温度的敏感性提高, 采伐干扰改变了土壤微生物群落转变的驱动因子.而Saynes等(2005)在热带次生林的研究表明, Cmic/Nmic的季节动态主要受降水的影响, 可能是由于该地区大部分时间内土壤较干旱, 因此降水后土壤环境剧变对微生物群落产生了明显的影响; 而在本研究区的生长季, 土壤一直比较湿润, 因而水分对微生物群落的限制作用不显著. ...
Soil-surface carbon dioxide efflux and microbial biomass in relation to tree density 13 years after a stand replacing fire in a lodgepole pine ecosystem.
2
2003
... 以往关于林龄对土壤微生物生物量影响的研究多集中于人工林(Chodak et al., 2009; 安然等, 2011; Mao et al., 2012; Yuan & Yue, 2012; Wen et al., 2014), 关于天然次生林的报道多集中在热带和亚热带地区(Saynes et al., 2005; Smith et al., 2015).虽然温带次生林在生态环境建设中起着举足轻重的作用, 但是温带次生林发展过程中土壤微生物生物量的变化情况及其影响因子尚不明确(Jia et al., 2005).以往研究表明: 随着林龄增加, 土壤微生物生物量可能逐渐增加(Chodak et al., 2009; Susyan et al., 2011; Mao et al., 2012)、减小(杨凯等, 2009; 安然等, 2011), 或达到最高值之后降低并保持稳定(Jia et al., 2005).这种结果的不一致可能是由研究对象以及林龄跨度不同造成的.随着次生林发展进程, 林分组成、生物量累积和分配等产生变化, 从而可能通过改变林内土壤温度、水分、凋落物质量等条件对土壤微生物产生间接影响(Litton et al., 2003; Jia et al., 2005), 也可能通过土壤酸化、有机质累积等过程直接影响微生物的生长繁殖及群落结构(Frouz & Nováková, 2005; Trap et al., 2011; Smith et al., 2015).由于不同林龄次生林土壤微生物的研究报道有限, 次生林发展过程中土壤微生物生物量对植被及环境因子的改变如何响应尚不明确. ...
The impact of logging residue on soil GHG fluxes in a drained peatland forest.
1
2012
... 本研究表明不同林龄落叶阔叶林土壤微生物生物量差异显著, 表现为随林龄增加先降低之后逐渐升高(图1).采伐迹地表层Cmic、Nmic与56年林分在整个生长季的均值差异不显著, 可能是由于采伐后残留在土壤里的根系死亡分解, 短期内为表层微生物提供了充足的可利用养分(Guo et al., 2004; Mäkiranta et al., 2012); 此外, 采伐后地上植被对养分的需求降低, 缓解了微生物与植物对养分的竞争(Kaiser et al., 2011), 因此短期维持了微生物生长和繁殖.但采伐迹地深层Cmic显著小于56年林分, 这主要可能是由于深层Cmic的主要限制因子为Ndis (表2), 而由于采伐干扰导致采伐迹地深层Ndis淋溶损失较多、Ndis含量较低, 因此抑制了微生物生长繁殖.采伐干扰10年后, 土壤微生物生物量显著降低, 之后随着林龄增长显著提高, 与Susyan等(2011)的研究结果一致.说明采伐的长期作用(土壤有机质降低、土壤紧实等)抑制了微生物的生长和繁殖(Foote et al., 2015), 但随着次生林的发展, 抑制作用能够逐渐解除.而黄土高原次生林发展过程中, 土壤微生物生物量在17年后达到最高值, 之后降低并保持稳定(Jia et al., 2005).造成这种差异的原因可能是黄土高原植被演替时间跨度较小, 但东北温带地区落叶阔叶林发展到阔叶红松林顶级群落需要200年左右.本研究中林龄较小, 只是森林演替进程中的一部分, 微生物生物量可能尚未达到最高值.因此, 为充分恢复东北温带地区退化土壤的性质及养分, 需要对天然次生林进行长期保护. ...
Changes in labile soil organic matter fractions following land use change from monocropping to poplar-based agroforestry systems in a semiarid region of Northeast China.
1
2012
... 以往关于林龄对土壤微生物生物量影响的研究多集中于人工林(Chodak et al., 2009; 安然等, 2011; Mao et al., 2012; Yuan & Yue, 2012; Wen et al., 2014), 关于天然次生林的报道多集中在热带和亚热带地区(Saynes et al., 2005; Smith et al., 2015).虽然温带次生林在生态环境建设中起着举足轻重的作用, 但是温带次生林发展过程中土壤微生物生物量的变化情况及其影响因子尚不明确(Jia et al., 2005).以往研究表明: 随着林龄增加, 土壤微生物生物量可能逐渐增加(Chodak et al., 2009; Susyan et al., 2011; Mao et al., 2012)、减小(杨凯等, 2009; 安然等, 2011), 或达到最高值之后降低并保持稳定(Jia et al., 2005).这种结果的不一致可能是由研究对象以及林龄跨度不同造成的.随着次生林发展进程, 林分组成、生物量累积和分配等产生变化, 从而可能通过改变林内土壤温度、水分、凋落物质量等条件对土壤微生物产生间接影响(Litton et al., 2003; Jia et al., 2005), 也可能通过土壤酸化、有机质累积等过程直接影响微生物的生长繁殖及群落结构(Frouz & Nováková, 2005; Trap et al., 2011; Smith et al., 2015).由于不同林龄次生林土壤微生物的研究报道有限, 次生林发展过程中土壤微生物生物量对植被及环境因子的改变如何响应尚不明确. ...
... 4个林龄林分Cmic/Nmic在生长季均呈现“W”形的变化格局(图4C), 表明土壤微生物群落结构能够随季节变化而转变(Björk et al., 2008; Smith et al., 2015).4个林龄林分Cmic/Nmic在生长季前期显著下降, 这可能是由于5月温度适宜, 加之土壤完全解冻后液态水中的养分为微生物提供了丰富的分解底物, 更有利于细菌的生长和繁殖(Kaiser et al., 2014); 生长季中期, Cmic/Nmic维持在较低水平, 到后期显著上升, 这可能是由于真菌更适宜在温度较低的环境下生存(丁爽和王传宽, 2009; Robert et al., 2010; Kaiser et al., 2011).除采伐迹地外, 其他3个林龄林分Cmic/Nmic均与Cdis/Ndis变化显著负相关(表3), 这与Song等(2015)的研究结果一致.说明随着季节更替, 地上植被通过改变土壤养分状况间接调节微生物群落结构的转变(Kaiser et al., 2014).而采伐迹地Cmic/Nmic受T5和Cdis的共同影响(表3), 这表明植被的减少导致土壤微生物群落转变对温度的敏感性提高, 采伐干扰改变了土壤微生物群落转变的驱动因子.而Saynes等(2005)在热带次生林的研究表明, Cmic/Nmic的季节动态主要受降水的影响, 可能是由于该地区大部分时间内土壤较干旱, 因此降水后土壤环境剧变对微生物群落产生了明显的影响; 而在本研究区的生长季, 土壤一直比较湿润, 因而水分对微生物群落的限制作用不显著. ...
Soil C and N dynamics in primary and secondary seasonally dry tropical forests in Mexico.
2
2005
... 土壤微生物生物量是土壤养分中最活跃的部分, 不仅通过转化有机质为植物提供可利用养分, 调节陆地生态系统碳氮循环(Xu et al., 2013; Zhou & Wang, 2015), 而且在土壤养分的累积过程中扮演着重要的角色, 对于受到干扰后退化土壤的肥力恢复具有重要意义(Jia et al., 2005; Saynes et al., 2005; Spohn et al., 2016).经历农耕、采伐等人为干扰后形成的天然次生林是陆地生态系统的重要组成部分(Yang et al., 2011).充分理解天然次生林发展过程中土壤微生物生物量的变异及其影响因子, 对于退化土壤肥力的恢复、合理经营和保护次生林具有重要意义(Jia et al., 2005; Smith et al., 2015). ...
... 以往关于林龄对土壤微生物生物量影响的研究多集中于人工林(Chodak et al., 2009; 安然等, 2011; Mao et al., 2012; Yuan & Yue, 2012; Wen et al., 2014), 关于天然次生林的报道多集中在热带和亚热带地区(Saynes et al., 2005; Smith et al., 2015).虽然温带次生林在生态环境建设中起着举足轻重的作用, 但是温带次生林发展过程中土壤微生物生物量的变化情况及其影响因子尚不明确(Jia et al., 2005).以往研究表明: 随着林龄增加, 土壤微生物生物量可能逐渐增加(Chodak et al., 2009; Susyan et al., 2011; Mao et al., 2012)、减小(杨凯等, 2009; 安然等, 2011), 或达到最高值之后降低并保持稳定(Jia et al., 2005).这种结果的不一致可能是由研究对象以及林龄跨度不同造成的.随着次生林发展进程, 林分组成、生物量累积和分配等产生变化, 从而可能通过改变林内土壤温度、水分、凋落物质量等条件对土壤微生物产生间接影响(Litton et al., 2003; Jia et al., 2005), 也可能通过土壤酸化、有机质累积等过程直接影响微生物的生长繁殖及群落结构(Frouz & Nováková, 2005; Trap et al., 2011; Smith et al., 2015).由于不同林龄次生林土壤微生物的研究报道有限, 次生林发展过程中土壤微生物生物量对植被及环境因子的改变如何响应尚不明确. ...
Successional and seasonal variations in soil and litter microbial community structure and function during tropical post-agricultural forest regeneration: A multi-year study.
5
2015
... 土壤微生物生物量是土壤养分中最活跃的部分, 不仅通过转化有机质为植物提供可利用养分, 调节陆地生态系统碳氮循环(Xu et al., 2013; Zhou & Wang, 2015), 而且在土壤养分的累积过程中扮演着重要的角色, 对于受到干扰后退化土壤的肥力恢复具有重要意义(Jia et al., 2005; Saynes et al., 2005; Spohn et al., 2016).经历农耕、采伐等人为干扰后形成的天然次生林是陆地生态系统的重要组成部分(Yang et al., 2011).充分理解天然次生林发展过程中土壤微生物生物量的变异及其影响因子, 对于退化土壤肥力的恢复、合理经营和保护次生林具有重要意义(Jia et al., 2005; Smith et al., 2015). ...
... 以往关于林龄对土壤微生物生物量影响的研究多集中于人工林(Chodak et al., 2009; 安然等, 2011; Mao et al., 2012; Yuan & Yue, 2012; Wen et al., 2014), 关于天然次生林的报道多集中在热带和亚热带地区(Saynes et al., 2005; Smith et al., 2015).虽然温带次生林在生态环境建设中起着举足轻重的作用, 但是温带次生林发展过程中土壤微生物生物量的变化情况及其影响因子尚不明确(Jia et al., 2005).以往研究表明: 随着林龄增加, 土壤微生物生物量可能逐渐增加(Chodak et al., 2009; Susyan et al., 2011; Mao et al., 2012)、减小(杨凯等, 2009; 安然等, 2011), 或达到最高值之后降低并保持稳定(Jia et al., 2005).这种结果的不一致可能是由研究对象以及林龄跨度不同造成的.随着次生林发展进程, 林分组成、生物量累积和分配等产生变化, 从而可能通过改变林内土壤温度、水分、凋落物质量等条件对土壤微生物产生间接影响(Litton et al., 2003; Jia et al., 2005), 也可能通过土壤酸化、有机质累积等过程直接影响微生物的生长繁殖及群落结构(Frouz & Nováková, 2005; Trap et al., 2011; Smith et al., 2015).由于不同林龄次生林土壤微生物的研究报道有限, 次生林发展过程中土壤微生物生物量对植被及环境因子的改变如何响应尚不明确. ...
... ; Smith et al., 2015).由于不同林龄次生林土壤微生物的研究报道有限, 次生林发展过程中土壤微生物生物量对植被及环境因子的改变如何响应尚不明确. ...
... 土壤微生物生物量的变化受土壤养分、水分、温度的共同影响, 而这些相关因子通常随季节转变而变化, 因此土壤微生物生物量通常呈现一定的季节动态(刘爽和王传宽, 2010; Kaiser et al., 2011; Wen et al., 2014).以往关于土壤微生物生物量季节变化的研究多针对不同森林类型(刘爽和王传宽, 2010; 刘纯等, 2014)、不同土地利用方式(Pandey et al., 2010)或不同林龄人工林(杨凯等, 2009), 而对不同林龄天然次生林土壤微生物生物量的季节变化尚不明确(Smith et al., 2015).不同林龄次生林植被活动(吸收养分、水分)及土壤养分、温湿度可能对季节变化产生不同的响应, 从而对微生物生物量的季节动态产生不同程度的影响.土壤可溶性养分较易被微生物吸收利用, 并且对季节变化的响应十分敏感(Edwards et al., 2006; Edwards & Jefferies, 2013; Freppaz et al., 2014); 土壤温度、湿度等环境因子的改变也调节着微生物的生长和繁殖(刘爽和王传宽, 2010; 刘纯等, 2014).因此, 探索不同林龄次生林土壤可溶性养分、土壤温湿度的季节变化格局及其与土壤微生物生物量的关系, 有利于充分理解次生林不同发展阶段土壤微生物生物量的驱动机制. ...
... 4个林龄林分Cmic/Nmic在生长季均呈现“W”形的变化格局(图4C), 表明土壤微生物群落结构能够随季节变化而转变(Björk et al., 2008; Smith et al., 2015).4个林龄林分Cmic/Nmic在生长季前期显著下降, 这可能是由于5月温度适宜, 加之土壤完全解冻后液态水中的养分为微生物提供了丰富的分解底物, 更有利于细菌的生长和繁殖(Kaiser et al., 2014); 生长季中期, Cmic/Nmic维持在较低水平, 到后期显著上升, 这可能是由于真菌更适宜在温度较低的环境下生存(丁爽和王传宽, 2009; Robert et al., 2010; Kaiser et al., 2011).除采伐迹地外, 其他3个林龄林分Cmic/Nmic均与Cdis/Ndis变化显著负相关(表3), 这与Song等(2015)的研究结果一致.说明随着季节更替, 地上植被通过改变土壤养分状况间接调节微生物群落结构的转变(Kaiser et al., 2014).而采伐迹地Cmic/Nmic受T5和Cdis的共同影响(表3), 这表明植被的减少导致土壤微生物群落转变对温度的敏感性提高, 采伐干扰改变了土壤微生物群落转变的驱动因子.而Saynes等(2005)在热带次生林的研究表明, Cmic/Nmic的季节动态主要受降水的影响, 可能是由于该地区大部分时间内土壤较干旱, 因此降水后土壤环境剧变对微生物群落产生了明显的影响; 而在本研究区的生长季, 土壤一直比较湿润, 因而水分对微生物群落的限制作用不显著. ...
Effects of historical logging on soil microbial communities in a subtropical forest in southern China.
2015
Dynamics of soil carbon, nitrogen, and phosphorus in calcareous soils after land-use abandonment—A chronosequence study.
1
2016
... 土壤微生物生物量是土壤养分中最活跃的部分, 不仅通过转化有机质为植物提供可利用养分, 调节陆地生态系统碳氮循环(Xu et al., 2013; Zhou & Wang, 2015), 而且在土壤养分的累积过程中扮演着重要的角色, 对于受到干扰后退化土壤的肥力恢复具有重要意义(Jia et al., 2005; Saynes et al., 2005; Spohn et al., 2016).经历农耕、采伐等人为干扰后形成的天然次生林是陆地生态系统的重要组成部分(Yang et al., 2011).充分理解天然次生林发展过程中土壤微生物生物量的变异及其影响因子, 对于退化土壤肥力的恢复、合理经营和保护次生林具有重要意义(Jia et al., 2005; Smith et al., 2015). ...
1
... 由于真菌C/N高于细菌(Sterner & Elser, 2002), 因此通常将Cmic/Nmic作为衡量微生物群落变化的指标, 即Cmic/Nmic越高, 真菌生物量越高(Zhou & Wang, 2015).虽然采伐迹地Cmic、Nmic生长季均值与56年林分无显著差异, 但采伐迹地Cmic/Nmic显著小于56年林分(图1C).说明采伐干扰后土壤微生物群落发生转变, 即细菌所占比例增高.这是由于采伐干扰后死亡根系分解及植物吸收减少等导致可利用底物含量提高(Guo et al., 2004), 使得快速增长的细菌成为优势群落(Kaiser et al., 2014).虽然本研究中Cmic/Nmic与土壤pH值没有显著的负相关关系(表2), 但56年林分土壤pH值最低, 而两个土层的Cmic/Nmic最高.这可能是由于56年林分凋落物层较厚造成土壤酸化(表1), 而真菌更耐受酸性土壤环境, 导致微生物群落真菌比例升高(Trap et al., 2011).随着土层加深, Cmic/Nmic增高, 这与以往研究结果(Fierer et al., 2003; 安然等, 2011)一致, 主要是因为土壤表层养分限制较低, 更适宜细菌生存.以上结果说明: 在次生林发展过程中, 土壤微生物能够通过改变群落结构来适应土壤养分、酸度等性质的改变. ...
Forest succession on abandoned arable soils in European Russia—Impacts on microbial biomass, fungal-bacterial ratio, and basal CO2 respiration activity.
1
2011
... 以往关于林龄对土壤微生物生物量影响的研究多集中于人工林(Chodak et al., 2009; 安然等, 2011; Mao et al., 2012; Yuan & Yue, 2012; Wen et al., 2014), 关于天然次生林的报道多集中在热带和亚热带地区(Saynes et al., 2005; Smith et al., 2015).虽然温带次生林在生态环境建设中起着举足轻重的作用, 但是温带次生林发展过程中土壤微生物生物量的变化情况及其影响因子尚不明确(Jia et al., 2005).以往研究表明: 随着林龄增加, 土壤微生物生物量可能逐渐增加(Chodak et al., 2009; Susyan et al., 2011; Mao et al., 2012)、减小(杨凯等, 2009; 安然等, 2011), 或达到最高值之后降低并保持稳定(Jia et al., 2005).这种结果的不一致可能是由研究对象以及林龄跨度不同造成的.随着次生林发展进程, 林分组成、生物量累积和分配等产生变化, 从而可能通过改变林内土壤温度、水分、凋落物质量等条件对土壤微生物产生间接影响(Litton et al., 2003; Jia et al., 2005), 也可能通过土壤酸化、有机质累积等过程直接影响微生物的生长繁殖及群落结构(Frouz & Nováková, 2005; Trap et al., 2011; Smith et al., 2015).由于不同林龄次生林土壤微生物的研究报道有限, 次生林发展过程中土壤微生物生物量对植被及环境因子的改变如何响应尚不明确. ...
Humus macro-morphology and soil microbial community changes along a 130-year- old
2
2011
... 以往关于林龄对土壤微生物生物量影响的研究多集中于人工林(Chodak et al., 2009; 安然等, 2011; Mao et al., 2012; Yuan & Yue, 2012; Wen et al., 2014), 关于天然次生林的报道多集中在热带和亚热带地区(Saynes et al., 2005; Smith et al., 2015).虽然温带次生林在生态环境建设中起着举足轻重的作用, 但是温带次生林发展过程中土壤微生物生物量的变化情况及其影响因子尚不明确(Jia et al., 2005).以往研究表明: 随着林龄增加, 土壤微生物生物量可能逐渐增加(Chodak et al., 2009; Susyan et al., 2011; Mao et al., 2012)、减小(杨凯等, 2009; 安然等, 2011), 或达到最高值之后降低并保持稳定(Jia et al., 2005).这种结果的不一致可能是由研究对象以及林龄跨度不同造成的.随着次生林发展进程, 林分组成、生物量累积和分配等产生变化, 从而可能通过改变林内土壤温度、水分、凋落物质量等条件对土壤微生物产生间接影响(Litton et al., 2003; Jia et al., 2005), 也可能通过土壤酸化、有机质累积等过程直接影响微生物的生长繁殖及群落结构(Frouz & Nováková, 2005; Trap et al., 2011; Smith et al., 2015).由于不同林龄次生林土壤微生物的研究报道有限, 次生林发展过程中土壤微生物生物量对植被及环境因子的改变如何响应尚不明确. ...
... 由于真菌C/N高于细菌(Sterner & Elser, 2002), 因此通常将Cmic/Nmic作为衡量微生物群落变化的指标, 即Cmic/Nmic越高, 真菌生物量越高(Zhou & Wang, 2015).虽然采伐迹地Cmic、Nmic生长季均值与56年林分无显著差异, 但采伐迹地Cmic/Nmic显著小于56年林分(图1C).说明采伐干扰后土壤微生物群落发生转变, 即细菌所占比例增高.这是由于采伐干扰后死亡根系分解及植物吸收减少等导致可利用底物含量提高(Guo et al., 2004), 使得快速增长的细菌成为优势群落(Kaiser et al., 2014).虽然本研究中Cmic/Nmic与土壤pH值没有显著的负相关关系(表2), 但56年林分土壤pH值最低, 而两个土层的Cmic/Nmic最高.这可能是由于56年林分凋落物层较厚造成土壤酸化(表1), 而真菌更耐受酸性土壤环境, 导致微生物群落真菌比例升高(Trap et al., 2011).随着土层加深, Cmic/Nmic增高, 这与以往研究结果(Fierer et al., 2003; 安然等, 2011)一致, 主要是因为土壤表层养分限制较低, 更适宜细菌生存.以上结果说明: 在次生林发展过程中, 土壤微生物能够通过改变群落结构来适应土壤养分、酸度等性质的改变. ...
An extraction method for measuring soil microbial biomass C.
Soil microbial biomass carbon and nitrogen in pure and mixed sites of Pinus massoniana and Cinnamomum camphora differing in stand age.
2
2014
... 以往关于林龄对土壤微生物生物量影响的研究多集中于人工林(Chodak et al., 2009; 安然等, 2011; Mao et al., 2012; Yuan & Yue, 2012; Wen et al., 2014), 关于天然次生林的报道多集中在热带和亚热带地区(Saynes et al., 2005; Smith et al., 2015).虽然温带次生林在生态环境建设中起着举足轻重的作用, 但是温带次生林发展过程中土壤微生物生物量的变化情况及其影响因子尚不明确(Jia et al., 2005).以往研究表明: 随着林龄增加, 土壤微生物生物量可能逐渐增加(Chodak et al., 2009; Susyan et al., 2011; Mao et al., 2012)、减小(杨凯等, 2009; 安然等, 2011), 或达到最高值之后降低并保持稳定(Jia et al., 2005).这种结果的不一致可能是由研究对象以及林龄跨度不同造成的.随着次生林发展进程, 林分组成、生物量累积和分配等产生变化, 从而可能通过改变林内土壤温度、水分、凋落物质量等条件对土壤微生物产生间接影响(Litton et al., 2003; Jia et al., 2005), 也可能通过土壤酸化、有机质累积等过程直接影响微生物的生长繁殖及群落结构(Frouz & Nováková, 2005; Trap et al., 2011; Smith et al., 2015).由于不同林龄次生林土壤微生物的研究报道有限, 次生林发展过程中土壤微生物生物量对植被及环境因子的改变如何响应尚不明确. ...
... 土壤微生物生物量的变化受土壤养分、水分、温度的共同影响, 而这些相关因子通常随季节转变而变化, 因此土壤微生物生物量通常呈现一定的季节动态(刘爽和王传宽, 2010; Kaiser et al., 2011; Wen et al., 2014).以往关于土壤微生物生物量季节变化的研究多针对不同森林类型(刘爽和王传宽, 2010; 刘纯等, 2014)、不同土地利用方式(Pandey et al., 2010)或不同林龄人工林(杨凯等, 2009), 而对不同林龄天然次生林土壤微生物生物量的季节变化尚不明确(Smith et al., 2015).不同林龄次生林植被活动(吸收养分、水分)及土壤养分、温湿度可能对季节变化产生不同的响应, 从而对微生物生物量的季节动态产生不同程度的影响.土壤可溶性养分较易被微生物吸收利用, 并且对季节变化的响应十分敏感(Edwards et al., 2006; Edwards & Jefferies, 2013; Freppaz et al., 2014); 土壤温度、湿度等环境因子的改变也调节着微生物的生长和繁殖(刘爽和王传宽, 2010; 刘纯等, 2014).因此, 探索不同林龄次生林土壤可溶性养分、土壤温湿度的季节变化格局及其与土壤微生物生物量的关系, 有利于充分理解次生林不同发展阶段土壤微生物生物量的驱动机制. ...
Substrate and environmental controls on microbial assimilation of soil organic carbon: A framework for Earth system models.
A global analysis of soil microbial biomass carbon, nitrogen and phosphorus in terrestrial ecosystems.
2
2013
... 土壤微生物生物量是土壤养分中最活跃的部分, 不仅通过转化有机质为植物提供可利用养分, 调节陆地生态系统碳氮循环(Xu et al., 2013; Zhou & Wang, 2015), 而且在土壤养分的累积过程中扮演着重要的角色, 对于受到干扰后退化土壤的肥力恢复具有重要意义(Jia et al., 2005; Saynes et al., 2005; Spohn et al., 2016).经历农耕、采伐等人为干扰后形成的天然次生林是陆地生态系统的重要组成部分(Yang et al., 2011).充分理解天然次生林发展过程中土壤微生物生物量的变异及其影响因子, 对于退化土壤肥力的恢复、合理经营和保护次生林具有重要意义(Jia et al., 2005; Smith et al., 2015). ...
... 以往关于林龄对土壤微生物生物量影响的研究多集中于人工林(Chodak et al., 2009; 安然等, 2011; Mao et al., 2012; Yuan & Yue, 2012; Wen et al., 2014), 关于天然次生林的报道多集中在热带和亚热带地区(Saynes et al., 2005; Smith et al., 2015).虽然温带次生林在生态环境建设中起着举足轻重的作用, 但是温带次生林发展过程中土壤微生物生物量的变化情况及其影响因子尚不明确(Jia et al., 2005).以往研究表明: 随着林龄增加, 土壤微生物生物量可能逐渐增加(Chodak et al., 2009; Susyan et al., 2011; Mao et al., 2012)、减小(杨凯等, 2009; 安然等, 2011), 或达到最高值之后降低并保持稳定(Jia et al., 2005).这种结果的不一致可能是由研究对象以及林龄跨度不同造成的.随着次生林发展进程, 林分组成、生物量累积和分配等产生变化, 从而可能通过改变林内土壤温度、水分、凋落物质量等条件对土壤微生物产生间接影响(Litton et al., 2003; Jia et al., 2005), 也可能通过土壤酸化、有机质累积等过程直接影响微生物的生长繁殖及群落结构(Frouz & Nováková, 2005; Trap et al., 2011; Smith et al., 2015).由于不同林龄次生林土壤微生物的研究报道有限, 次生林发展过程中土壤微生物生物量对植被及环境因子的改变如何响应尚不明确. ...
... 土壤微生物生物量的变化受土壤养分、水分、温度的共同影响, 而这些相关因子通常随季节转变而变化, 因此土壤微生物生物量通常呈现一定的季节动态(刘爽和王传宽, 2010; Kaiser et al., 2011; Wen et al., 2014).以往关于土壤微生物生物量季节变化的研究多针对不同森林类型(刘爽和王传宽, 2010; 刘纯等, 2014)、不同土地利用方式(Pandey et al., 2010)或不同林龄人工林(杨凯等, 2009), 而对不同林龄天然次生林土壤微生物生物量的季节变化尚不明确(Smith et al., 2015).不同林龄次生林植被活动(吸收养分、水分)及土壤养分、温湿度可能对季节变化产生不同的响应, 从而对微生物生物量的季节动态产生不同程度的影响.土壤可溶性养分较易被微生物吸收利用, 并且对季节变化的响应十分敏感(Edwards et al., 2006; Edwards & Jefferies, 2013; Freppaz et al., 2014); 土壤温度、湿度等环境因子的改变也调节着微生物的生长和繁殖(刘爽和王传宽, 2010; 刘纯等, 2014).因此, 探索不同林龄次生林土壤可溶性养分、土壤温湿度的季节变化格局及其与土壤微生物生物量的关系, 有利于充分理解次生林不同发展阶段土壤微生物生物量的驱动机制. ...
Carbon and nitrogen dynamics during forest stand development: A global synthesis.
1
2011
... 土壤微生物生物量是土壤养分中最活跃的部分, 不仅通过转化有机质为植物提供可利用养分, 调节陆地生态系统碳氮循环(Xu et al., 2013; Zhou & Wang, 2015), 而且在土壤养分的累积过程中扮演着重要的角色, 对于受到干扰后退化土壤的肥力恢复具有重要意义(Jia et al., 2005; Saynes et al., 2005; Spohn et al., 2016).经历农耕、采伐等人为干扰后形成的天然次生林是陆地生态系统的重要组成部分(Yang et al., 2011).充分理解天然次生林发展过程中土壤微生物生物量的变异及其影响因子, 对于退化土壤肥力的恢复、合理经营和保护次生林具有重要意义(Jia et al., 2005; Smith et al., 2015). ...
Soil microbial and enzymatic activities across a chronosequence of Chinese pine plantation development on the Loess Plateau of China.
1
2012
... 以往关于林龄对土壤微生物生物量影响的研究多集中于人工林(Chodak et al., 2009; 安然等, 2011; Mao et al., 2012; Yuan & Yue, 2012; Wen et al., 2014), 关于天然次生林的报道多集中在热带和亚热带地区(Saynes et al., 2005; Smith et al., 2015).虽然温带次生林在生态环境建设中起着举足轻重的作用, 但是温带次生林发展过程中土壤微生物生物量的变化情况及其影响因子尚不明确(Jia et al., 2005).以往研究表明: 随着林龄增加, 土壤微生物生物量可能逐渐增加(Chodak et al., 2009; Susyan et al., 2011; Mao et al., 2012)、减小(杨凯等, 2009; 安然等, 2011), 或达到最高值之后降低并保持稳定(Jia et al., 2005).这种结果的不一致可能是由研究对象以及林龄跨度不同造成的.随着次生林发展进程, 林分组成、生物量累积和分配等产生变化, 从而可能通过改变林内土壤温度、水分、凋落物质量等条件对土壤微生物产生间接影响(Litton et al., 2003; Jia et al., 2005), 也可能通过土壤酸化、有机质累积等过程直接影响微生物的生长繁殖及群落结构(Frouz & Nováková, 2005; Trap et al., 2011; Smith et al., 2015).由于不同林龄次生林土壤微生物的研究报道有限, 次生林发展过程中土壤微生物生物量对植被及环境因子的改变如何响应尚不明确. ...
Reviews and syntheses: Soil resources and climate jointly drive variations in microbial biomass carbon and nitrogen in China’s forest ecosystems.
4
2015
... 土壤微生物生物量是土壤养分中最活跃的部分, 不仅通过转化有机质为植物提供可利用养分, 调节陆地生态系统碳氮循环(Xu et al., 2013; Zhou & Wang, 2015), 而且在土壤养分的累积过程中扮演着重要的角色, 对于受到干扰后退化土壤的肥力恢复具有重要意义(Jia et al., 2005; Saynes et al., 2005; Spohn et al., 2016).经历农耕、采伐等人为干扰后形成的天然次生林是陆地生态系统的重要组成部分(Yang et al., 2011).充分理解天然次生林发展过程中土壤微生物生物量的变异及其影响因子, 对于退化土壤肥力的恢复、合理经营和保护次生林具有重要意义(Jia et al., 2005; Smith et al., 2015). ...




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}