



全球气候变化会对陆地生态系统的结构、功能和分布产生重要的影响, 尤其在中高纬度地区受气候变化的影响更大(Hu et al., 2002)。全球变暖导致极端气候事件发生频率、发生强度、发生范围的增加, 对森林生态系统产生了较大的负面影响(Webb, 2001), 因此气候变暖对森林生态系统所造成的影响已经成为目前研究的热点问题。
在森林生态系统中, 气候变化及多种生态因子直接或间接作用于树木的生长过程(Kramer et al., 2000), 导致树木表现出相应的径向生长变化(Goldblum & Rigg, 2005), 树木年轮的宽窄承载着多种气候因子和生态因子的信息。运用树木年轮学方法可去除生长趋势和其他非气候因素的影响, 从而最大限度保留树轮宽度年表的气候信息(Fritts, 1976), 进而探讨影响树木径向生长的关键气候因子。
树木径向生长对气候变化的响应不仅受到气候因子的影响, 同时还受到其他因素的作用, 以往研究发现树木径向生长对气候变化的响应因树种、环境因素和地域的差异而有明显不同(Leal et al., 2007), 例如由于地理位置(如纬度、经度、海拔)的不同会导致气候条件的变化, 进而使落叶松等树种呈现出不同的年轮-气候关系(Wang et al., 2009; Wu et al., 2017)。林龄也是影响年轮-气候关系的一个重要因素, 有研究表明幼龄树和老龄树的径向生长与气候因子间的关系存在显著差异, 例如祁连山脉不同林龄的祁连圆柏(Sabina przewalskii)在气候响应方面存在明显的不同(Yu et al., 2008)。同时不同树种径向生长对气候变化的响应也存在差异, 比如梁鹏鸿等(2016)研究发现杨桦林、蒙古栎林、阔叶红松林对气候变化的响应不同, 从杨桦林到阔叶红松林, 树木径向生长对气候波动的敏感性逐步降低。
目前对落叶松的年轮研究多集中在大兴安岭、长白山一带的落叶松天然林, 对辽东山区落叶松人工林径向生长的研究相对较少, 而且多数研究只探讨了气候条件对年轮-气候关系的影响, 或者在同一地点研究树种、林分因子对年轮-气候关系的影响, 然而这些因素如何共同作用来影响树木径向生长对气候变化的响应还不明确, 这些因素的相对作用大小还不太清楚。辽东山区广泛栽培有黄花落叶松(Larix olgensis)和日本落叶松(Larix kaempferi)人工林, 其对气候变化响应的影响机制还需进一步研究, 比如气候条件、树种和林分因子对年轮-气候关系影响作用的相对大小如何?研究这些问题对气候变化背景下落叶松人工林合理经营有重要意义。
本文以辽宁草河口和湾甸子林场不同林龄的黄花落叶松和日本落叶松人工林为研究对象, 通过研究树木径向生长与气候变化的关系, 来探讨以下三个问题: (1)不同林龄黄花落叶松和日本落叶松人工林对历史气候变化的响应; (2)气候条件、树种和林分因子(林龄、密度、蓄积量)是否影响落叶松对气候变化的响应(即年轮-气候关系); (3)气候条件和林分因子对年轮-气候关系影响作用的相对大小。
1 研究方法
1.1 研究区概况
研究地点位于辽宁省本溪满族自治县草河口林场(123.90° E, 40.87° N)和清原满族自治县湾甸子林场(125.80° E, 41.58° N), 都位于辽东山区, 属于温带大陆性季风气候。草河口林场属于长白山系千山支脉, 年平均气温6.5 ℃, 年降水量926.3 mm, 无霜期127天; 湾甸子林场属于长白山系龙岗山支脉北坡, 年平均气温3.4 ℃, 年降水量810 mm, 无霜期125天。研究区的海拔并不高, 介于300-600 m之间, 没有明显的植被垂直分布。地带性植被类型为针阔混交林, 天然落叶松林在辽宁多有分布, 同时落叶松人工林面积也较大, 目前全省拥有落叶松人工林557 201.5 hm2。作为辽宁省主要的造林树种(董厚德, 2011), 日本落叶松为20世纪30年代由日本引进, 黄花落叶松早在1912年在辽宁就被采用为人工造林树种(董厚德, 2011)。
1.2 样本采集与年表建立
在草河口和湾甸子林场, 选择不同林龄、不同立地条件的黄花落叶松和日本落叶松人工林设置样地。其中, 草河口林场设置黄花落叶松样地4块、日本落叶松样地6块, 湾甸子林场设置黄花落叶松样地3块、日本落叶松样地4块。样地面积50 m × 20 m, 由10个10 m × 10 m的样格组成。对样地中的乔木进行调查, 记录物种组成, 然后测定胸径大于3 cm树木的胸径和树高。同时, 记录样地的经纬度、海拔、坡度等环境因子, 并根据森林二类清查资料对样地的立地条件按“好” “中” “差”进行分级(方精云等, 2009)。根据落叶松的生长阶段将不同林龄落叶松人工林划分为中龄林(20-30 a)、成熟林(大于40 a)(曾凡鹏等, 2016)。群落调查和年轮采样在2016年完成, 各样地林分结构特征和基本信息见表1。
表1 草河口(CHK)、湾甸子(WDZ)黄花落叶松和日本落叶松的样地基本信息和林分结构特征
Table 1
| 样地编号 Plot ID | 年平均气温 Annual mean air temperature (℃) | 年降水量 Annual precipitation (mm) | 树种 Tree species | 林龄 Stand age | 立地条件 Site quality | 林分密度 Stem density (tree·hm-2) | 蓄积量 Wood volume (m³·hm-2) | 平均胸径 Mean DBH (cm) |
|---|---|---|---|---|---|---|---|---|
| CHK01 | 5.71 | 874.84 | 黄花落叶松 L. olgensis | 77 | 好 Well | 570 | 294.56 | 22.72 |
| CHK02 | 5.57 | 880.13 | 黄花落叶松 L. olgensis | 78 | 差 Poor | 990 | 324.91 | 19.72 |
| CHK03 | 5.90 | 867.84 | 日本落叶松 L. kaempferi | 24 | 中 Good | 680 | 105.02 | 15.56 |
| CHK04 | 5.97 | 865.17 | 日本落叶松 L. kaempferi | 25 | 差 Poor | 880 | 122.57 | 15.23 |
| CHK05 | 5.78 | 869.58 | 日本落叶松 L. kaempferi | 69 | 中 Good | 540 | 442.54 | 26.93 |
| CHK06 | 5.82 | 868.24 | 日本落叶松 L. kaempferi | 71 | 中 Good | 300 | 373.81 | 33.89 |
| CHK07 | 6.15 | 858.84 | 日本落叶松 L. kaempferi | 21 | 好 Well | 580 | 108.62 | 17.14 |
| CHK08 | 6.08 | 861.56 | 日本落叶松 L. kaempferi | 24 | 好 Well | 520 | 107.55 | 18.05 |
| CHK09 | 5.89 | 868.66 | 黄花落叶松 L. olgensis | 21 | 好 Well | 620 | 157.50 | 18.95 |
| CHK10 | 5.86 | 869.79 | 黄花落叶松 L. olgensis | 21 | 好 Well | 620 | 111.31 | 17.36 |
| WDZ01 | 4.19 | 832.08 | 黄花落叶松 L. olgensis | 49 | 中 Good | 520 | 353.52 | 25.97 |
| WDZ02 | 3.97 | 839.94 | 黄花落叶松 L. olgensis | 50 | 差 Poor | 540 | 322.13 | 26.26 |
| WDZ03 | 3.74 | 848.05 | 日本落叶松 L. kaempferi | 73 | 好 Well | 1490 | 761.10 | 23.66 |
| WDZ04 | 3.73 | 848.42 | 日本落叶松 L. kaempferi | 72 | 中 Good | 1090 | 608.39 | 24.09 |
| WDZ05 | 3.74 | 844.35 | 日本落叶松 L. kaempferi | 23 | 好 Well | 1030 | 272.70 | 17.58 |
| WDZ06 | 3.52 | 852.33 | 日本落叶松 L. kaempferi | 23 | 好 Well | 960 | 263.49 | 18.67 |
| WDZ07 | 3.91 | 838.52 | 黄花落叶松 L. olgensis | 20 | 差 Poor | 1290 | 169.18 | 15.04 |
DBH, diameter at breast height.
在每个样地内, 主要选择林冠层树木及少量中小径级个体钻取树芯, 每个样地钻取30-40株。用生长锥在胸径处(距地面1.3 m)沿东北、西南方向各钻取一根树芯。把从野外采集回来的树芯按照标准的处理方法进行干燥、固定、打磨(Stokes, 1996), 直至能在显微镜下清楚的辨别每根树芯的早材和晚材。然后通过精度为0.001 mm的LINTAB 6年轮宽度测量仪(Rintech, Heidelberg, Germany)和TSAP软件对年轮宽度进行测定。应用COFECHA程序对年轮序列进行交叉定年(Holmes, 1983), 以保证定年和测量的准确性, 并去除问题较大的树芯。由于树木的生长不仅受环境和生态因子的影响, 而且还受到自身生理因子的控制, 所以通过ARSTAN程序采用负指数函数模拟树木的生长趋势, 并消除非气候因素和树木生长趋势的影响, 进而得到标准年表, 年表中的年轮宽度指数(RWI)为实际测得年轮宽度与去趋势后的年轮宽度值的比值(Fritts, 1976)。
1.3 气候数据
气候数据由英国东安格利亚大学气候研究所建立的全球气候数据库(CRU TS 3.22 Global Climate Database, http://www.cru.uea.ac.uk/)下载(Mitchell & Jones, 2005)。我国诸多学者利用CRU网格数据对不同区域的降水、气温、蒸发等气候要素进行研究, 结果表明CRU网格数据在揭示气候要素变化特征方面有着较高的可靠性(黄秋霞等, 2013; 戚颖等, 2014), 该数据在国内外年轮年代学研究中得到了广泛的应用。因此本文气候资料采用CRU TS 3.22的月平均气温、月降水量和帕尔默干旱指数(PDSI)等气候数据。由于各样地落叶松人工林林龄不同, 因此气候数据选取成熟林年表共有的时间段(1986- 2015年), 而中林龄年表长度较短, 选取年表全部时间段进行分析, 以保持各样地结果的可比性。考虑到“滞后效应”, 即上一年气候可能会对当年的树木生长产生影响(李江风, 2000), 选取前一年夏季到当年秋季的平均气温、PDSI与RWI进行相关分析来研究落叶松人工林径向生长对气候变化的响应。
1.4 统计分析
为分析样地气候条件对落叶松年轮-气候关系的影响, 对相关气候数据进行提取和计算。根据调查样地的地理坐标, 利用ArcGIS 10.0从全球高分辨率气候数据库中提取各样地1-12月平均气温与降水量数据, 并计算下述指标: (1)年平均气温(MAT, ℃), 反映气温影响; (2)年降水量(MAP, mm), 反映水分的有效性; (3)潜在蒸散量(PET, mm), 是年轮-气候关系分析重要的气候指标, 反映能量有效性, 计算方法见文献(方精云, 1992)。这些气候指标的选取是因为热量积累、水分供应等因子对年轮-气候关系可能存在影响作用(Wang et al., 2009)。
首先用RWI和上年夏季到当年秋季各季节平均气温、PDSI进行相关分析。为了分析样地气候条件、立地条件、树种和林分因子对年轮-气候关系的影响大小, 对上述RWI与各季节气候指标的相关系数进行主成分分析(PCA), 利用一般线性模型(GLM)来分析探讨各种环境和生物因素对落叶松年轮-气候关系的影响。其中, 环境因素包括气候因子和立地条件。气候因子则包括潜在蒸发散、年降水量, 分别反映热量和水分的有效性, 因为以往研究表明热量和水分是影响东北地区森林生物量、生产力的关键气候要素(Wang et al., 2006; Xu et al., 2017)。第二类为生物因子, 包括树种(黄花落叶松和日本落叶松), 以及林分因子(密度、林龄和蓄积量), 有研究表明树种和林龄、蓄积量等林分因子可能会影响年轮-气候关系(Leal et al., 2007; Rais et al., 2014a)。
所有的数据分析均利用R 3.4.3软件进行, 检验水平设置值为0.1。
2 结果和分析
2.1 研究区气候变化趋势
CRU数据表明, 草河口林场由于纬度偏低, 年平均气温高于湾甸子林场, 但两个地点的水分条件没有明显的区别(图1)。半个世纪(1966-2015年)以来, 草河口地区的年平均气温以每年0.018 9 ℃ (R2 = 0.194, p < 0.01)的速度递增, 湾甸子地区的年平均气温以每年0.018 5 ℃ (R2 = 0.200, p < 0.01)的速度递增, 但PDSI变化都不显著。因此, 研究区近几十年来呈现出显著的气候变暖的趋势。
图1
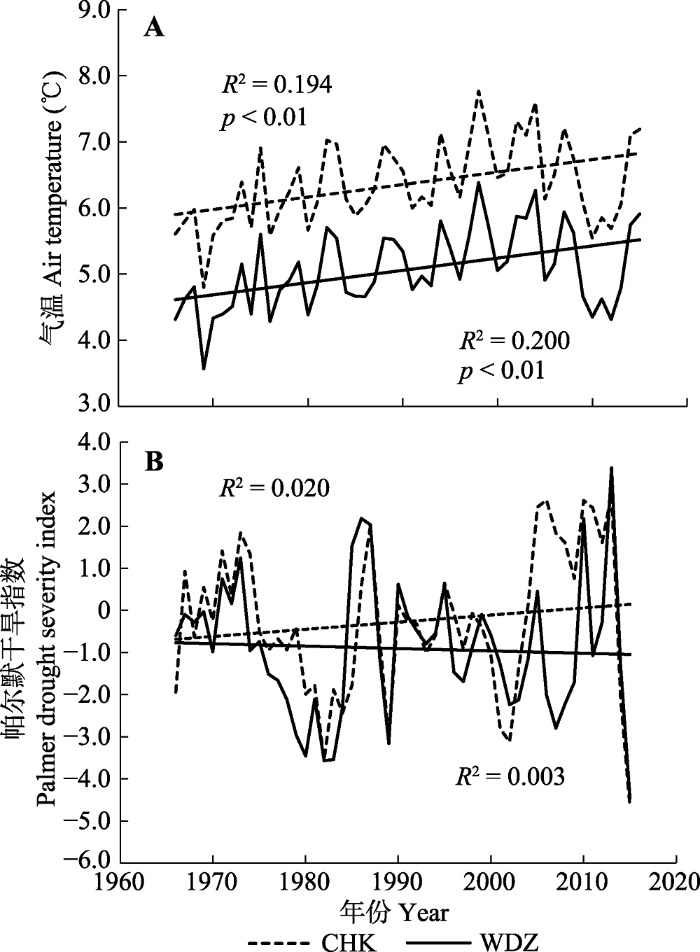
图1
草河口(CHK)和湾甸子(WDZ)年平均气温(A)与帕尔默干旱指数(B)的变化趋势。
Fig. 1
Trends in annual mean air temperature (A) and palmer drought severity index (B) in Caohekou (CHK) and Wandianzi (WDZ).
2.2 年表统计特征
通过负指数函数进行去趋势, 一共得到17个标准年表, 成熟林用于与气候进行分析的年表长度为1986-2016年, 而中林龄年表长度较短, 选取年表全部时间段与气候进行分析。各样地年表的统计参数如表2所示, CHK07的样芯长度最短, 只有15年, WDZ03的样芯长度最长, 达73年, 各样地的平均敏感度均大于0.140, 其中WDZ07最高, 为0.253。各样地的标准偏差(SD)介于0.142-0.278之间, 其中WDZ07的最大。17个样地树间相关系数为0.242- 0.603, CHK06最低, 草河口CHK10最高。CHK05的一阶自相关系数最高, 表明CHK05的落叶松径向生长受上一年气候因素的影响最大。各样地的样本总体代表性(EPS)均很高, 都明显超过0.85, 而且各样地信噪比都较高, 为11.489-51.739, 说明所建年表质量较好, 适合进一步进行轮宽与气候变化关系的分析(吴祥定, 1990)。表2 草河口(CHK)和湾甸子(WDZ)样地落叶松标准年表统计参数
Table 2 Summary of statistics for standard chronology in sampling plots of Caohekou (CHK) and Wandianzi (WDZ)
| 样地 Plot | 树芯/株数 Number of cores/trees | 样芯长度 Length of series (a) | 用于气候分析 的年表长度 Chronology length for climate analysis (a) | 平均敏感度 Mean sensitivity | 标准偏差 Standard deviation | R1样本间平均相关系数 Mean correlations among all radii | 一阶自相 关系数 Autocorrelation order 1 | 信噪比 Signal-to- noise ratio | 样本总体 代表性 Express population signal | 第一主成分所占方差量 PCA1 (%) |
|---|---|---|---|---|---|---|---|---|---|---|
| CHK01 | 39/23 | 1950-2016 | 1986-2016 | 0.170 | 0.217 | 0.340 | 0.610 | 18.064 | 0.948 | 0.411 |
| CHK02 | 63/34 | 1950-2016 | 1986-2016 | 0.199 | 0.250 | 0.431 | 0.586 | 38.694 | 0.975 | 0.463 |
| CHK03 | 43/23 | 1996-2016 | 1996-2016 | 0.160 | 0.142 | 0.324 | -0.054 | 14.382 | 0.935 | 0.396 |
| CHK04 | 46/23 | 1996-2016 | 1996-2016 | 0.165 | 0.151 | 0.395 | 0.244 | 20.857 | 0.954 | 0.432 |
| CHK05 | 38/20 | 1956-2016 | 1986-2016 | 0.147 | 0.212 | 0.307 | 0.680 | 13.743 | 0.932 | 0.368 |
| CHK06 | 49/27 | 1956-2016 | 1986-2016 | 0.154 | 0.167 | 0.242 | 0.449 | 13.431 | 0.931 | 0.283 |
| CHK07 | 39/20 | 2002-2016 | 2002-2016 | 0.213 | 0.217 | 0.475 | 0.409 | 33.504 | 0.971 | 0.524 |
| CHK08 | 38/20 | 2000-2016 | 2000-2016 | 0.151 | 0.145 | 0.285 | 0.142 | 15.149 | 0.938 | 0.354 |
| CHK09 | 46/26 | 1989-2016 | 1989-2016 | 0.161 | 0.186 | 0.493 | 0.392 | 36.937 | 0.974 | 0.535 |
| CHK10 | 40/22 | 1996-2016 | 1996-2016 | 0.181 | 0.239 | 0.603 | 0.491 | 51.739 | 0.981 | 0.639 |
| WDZ01 | 40/22 | 1968-2016 | 1986-2016 | 0.200 | 0.199 | 0.414 | 0.267 | 24.718 | 0.961 | 0.464 |
| WDZ02 | 41/21 | 1967-2016 | 1986-2016 | 0.225 | 0.220 | 0.501 | 0.246 | 36.094 | 0.973 | 0.533 |
| WDZ03 | 62/36 | 1944-2016 | 1986-2016 | 0.204 | 0.211 | 0.313 | 0.349 | 26.460 | 0.964 | 0.347 |
| WDZ04 | 44/24 | 1945-2016 | 1986-2016 | 0.166 | 0.178 | 0.284 | 0.247 | 11.489 | 0.920 | 0.329 |
| WDZ05 | 64/34 | 1994-2016 | 1994-2016 | 0.143 | 0.150 | 0.278 | 0.273 | 20.824 | 0.954 | 0.332 |
| WDZ06 | 42/28 | 1994-2016 | 1994-2016 | 0.154 | 0.155 | 0.430 | 0.100 | 26.446 | 0.964 | 0.470 |
| WDZ07 | 81/43 | 1971-2016 | 1986-2016 | 0.253 | 0.278 | 0.247 | 0.448 | 20.714 | 0.954 | 0.308 |
PCA1, variance in the first principal component.
2.3 径向生长与气候因子的关系
落叶松年轮-气候关系明显受到林龄等林分因素的影响, 比如图2中落叶松径向生长与上年夏季气温的相关关系随林龄增加呈显著下降的趋势(p < 0.001), 同样与上年秋季气温(p < 0.001)、当年夏季气温(p < 0.05)和当年秋季气温(p < 0.1)的相关关系随林龄增加呈显著下降的趋势。可以看出过去几十年的气候变暖对中龄落叶松人工林径向生长有促进作用, 而对成熟落叶松人工林有抑制作用。
图2
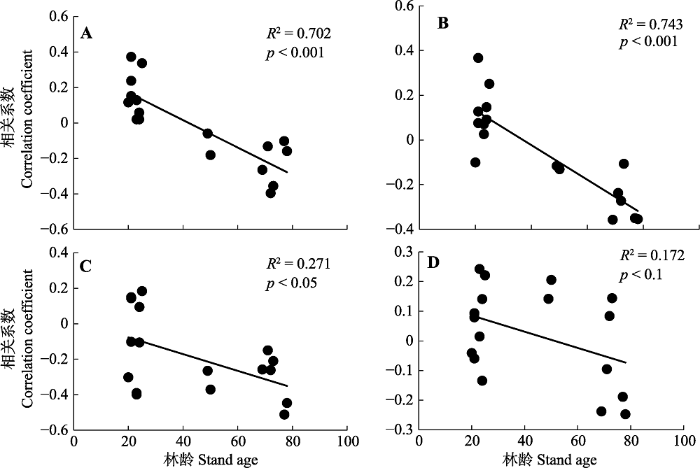
图2
草河口和湾甸子落叶松径向生长与不同季节气温相关关系随林龄的变化趋势。A, 上年夏季气温。B, 上年秋季气温。C, 当年夏季气温。D, 当年秋季气温。
Fig. 2
The trends of relationships between the radial growth of larch and air temperature in different seasons with increasing age of Caohekou and Wandianzi. A, Air temperature in summer of the previous year. B, Air temperature in autumn of the previous year. C, Air temperature in summer of the current year. D, Air temperature in autumn of the current year.
图3
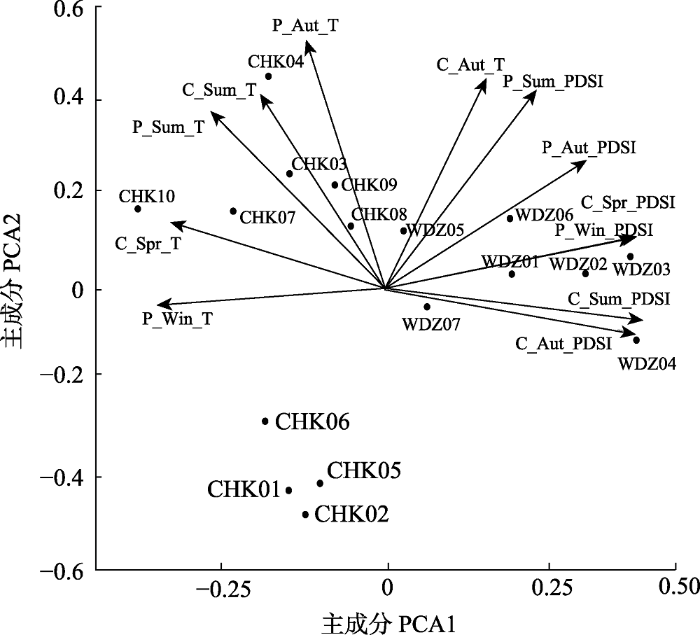
图3
草河口和湾甸子落叶松年轮宽度指数与气温(T)、帕尔默干旱指数(PDSI)的主成分分析。P表示上一年, C表示当年, Spr、Sum、Aut、Win分别表示春、夏、秋、冬季。
Fig. 3
Principal component analysis of ring width index of larch and air temperature (T), palmer drought severity index (PDSI) of Caohekou and Wandianzi. P designates the previous year, and C the current year. Spr, Sum, Aut and Win are abbreviations for spring, summer, autumn and winter, respectively.
RWI和季节气候因子的相关系数作主成分分析得前四轴载荷值, 由表3可知, PCA1轴的PDSI的载荷值都为正值, 因此主成分的正值越大, PDSI正相关关系越强, 主成分的负值越小, PDSI的负相关关系越强, 说明该轴主要反映了轮宽对水分条件历史变化的响应; PCA2轴的气温载荷值大多数为正值, 因此主成分的正值越大, 气温的正相关关系越强, 主成分的负值越小, 气温的负相关关系越强, 说明该轴主要反映了轮宽对历史气温变化的响应。
表3 季节气候因子在年轮宽度指数与气温、帕尔默干旱指数主成分分析各轴的载荷值
Table 3
| 变量 Variable | Comp. 1 | Comp. 2 | Comp. 3 | Comp. 4 |
|---|---|---|---|---|
| P_Sum_T | -0.253 | 0.361 | -0.318 | 0.233 |
| P_Aut_T | -0.115 | 0.507 | -0.048 | 0.084 |
| P_Win_T | -0.330 | -0.035 | 0.426 | 0.005 |
| C_Spr_T | -0.310 | 0.134 | -0.427 | 0.167 |
| C_Sum_T | -0.171 | 0.381 | 0.440 | -0.555 |
| C_Aut _T | 0.146 | 0.427 | -0.353 | -0.445 |
| P_Sum_PDSI | 0.219 | 0.405 | 0.212 | 0.475 |
| P_Aut_PDSI | 0.291 | 0.261 | 0.363 | 0.356 |
| P_Win_PDSI | 0.365 | 0.105 | -0.053 | -0.133 |
| C_Spr_PDSI | 0.359 | 0.102 | 0.002 | -0.183 |
| C_Sum_PDSI | 0.374 | -0.066 | -0.052 | -0.022 |
| C_Aut_PDSI | 0.364 | -0.092 | -0.178 | 0.019 |
P designates the previous year, and C is the current year, T is the mean air temperature, PDSI is the palmer drought severity index, Spr, Sum, Aut and Win are abbreviations for spring (March to May), summer (June to August), autumn (September to November), and winter (December to February), respectively.
P表示上一年, C表示当年, T为平均气温, PDSI为帕尔默干旱指数, Spr、Sum、Aut、Win分别表示春(3-5月)、夏(6-8月)、秋(9-11月)、冬季(12-2月)。
2.4 气候条件、林分因子对年轮-气候关系的影响
表4为环境因子、树种和林分因子对PCA1轴、2轴得分的单因素解释力(R2)。可以看出潜在蒸发散(PET)和蓄积量与PCA1轴的相关关系很显著(p < 0.001), 林分密度也达到了显著水平(p < 0.05), 这说明热量、水分条件、蓄积量和密度可能对年轮-气候关系有着重要的作用。林龄与PCA2轴存在显著相关关系(p < 0.001), 可以看出林分因子中的林龄对年轮-气候关系同样起到重要的作用。而立地条件、树种对PCA1、2轴均没有显著的解释力。
表4 环境因子、林分结构特征对年轮宽度指数与气温、帕尔默干旱指数的PCA1、2轴得分的解释力(R2)
Table 4
| 变量 Variable | 年降水量 Mean annual precipitation | 潜在蒸发量 Potential evapotranspiration | 林龄 Stand age | 林分密度 Stem density | 蓄积量 Wood volume | 立地条件 Site quality | 树种 Tree species |
|---|---|---|---|---|---|---|---|
| PCA1 | -0.522** | -0.722*** | 0.091 | 0.287* | 0.512** | 0.013 | 0.007 |
| PCA2 | -0.087 | 0.000 | -0.660*** | 0.011 | -0.197 | 0.090 | 0.056 |
“-” indicates a negative correlation, *, p < 0.05; **, p < 0.01; ***, p < 0.001
“-”表示负相关
附录I 年轮宽度指数与气温、帕尔默干旱指数的PCA1、2轴得分与环境因子、林分结构特征多元回归分析
Supplement I
| PCA1 | PCA2 | |||||
|---|---|---|---|---|---|---|
| df | % SS | p | df | % SS | p | |
| 树种 Tree species | 1 | 0.729 | 0.535 | 1 | 5.567 | 0.167 |
| 林分密度 Stem density | 1 | 28.043 | 0.004** | 1 | 0.625 | 0.625 |
| 林龄 Stand age | 1 | 8.225 | 0.061 | 1 | 64.605 | 0.000*** |
| 蓄积量 Wood volume | 1 | 34.052 | 0.002** | 1 | 4.956 | 0.190 |
| 潜在蒸发量Potential evapotranspiration | 1 | 11.502 | 0.032* | 1 | 1.905 | 0.400 |
| 年降水量 Annual precipitation | 1 | 2.555 | 0.260 | 1 | 0.662 | 0.615 |
| 立地条件 Site quality | 2 | 1.012 | 0.755 | 2 | 2.356 | 0.631 |
| 残差 Residuals | 8 | 13.882 | 8 | 19.323 | ||
*, p < 0.05; **, p < 0.01; ***, p < 0.001.
表5 年轮宽度指数与气温、帕尔默干旱指数的PCA1、2轴得分与环境因子、林分结构特征多元回归分析
Table 5
| PCA1 | PCA2 | |||||
|---|---|---|---|---|---|---|
| df | % SS | p | df | % SS | p | |
| 潜在蒸发量 Potential evapotranspiration | 1 | 72.170 | 0.000 2*** | 1 | 0.003 | 0.973 |
| 年降水量 Mean annual precipitation | 1 | 0.884 | 0.496 | 1 | 21.646 | 0.017* |
| 立地条件 Site quality | 2 | 1.122 | 0.733 | 2 | 13.291 | 0.123 |
| 树种 Tree species | 1 | 0.591 | 0.576 | 1 | 10.129 | 0.075 |
| 林分密度 Stem density | 1 | 0.212 | 0.736 | 1 | 1.219 | 0.498 |
| 林龄 Stand age | 1 | 7.990 | 0.064 | 1 | 27.443 | 0.009** |
| 蓄积量 Wood volume | 1 | 3.150 | 0.215 | 1 | 6.945 | 0.128 |
| 残差 Residuals | 8 | 13.881 | 8 | 19.323 | ||
*, p < 0.05; **, p < 0.01; ***, p < 0.001.
3 讨论
本研究结果表明, 研究地点的气候条件、林分因子对落叶松人工林径向生长对气候变化的响应(即年轮-气候关系)有显著影响。其中, 气候因子中热量和水分的有效性都有着重要作用; 林分因子中林龄、密度和蓄积量可能起主要作用, 而树种(黄花和日本落叶松)之间的差异影响似乎并不大。
3.1 气候条件与落叶松径向生长的关系
前人的研究表明由于地理位置的不同(如纬度、经度、海拔)会导致气候条件的变化, 进而使落叶松呈现出不同的年轮-气候关系(Wang et al., 2009)。不同的海拔树木径向生长对气候变化的敏感性不同(常锦峰等, 2009; 曾令兵等, 2012); 不同纬度红松(Pinus koraiensis)的径向生长对气候变化的响应也存在差异(刘敏等, 2016)。本研究与上述研究结果一致, 图3中PCA1轴主要反映了地点(气候条件)的差异, 因为草河口位于湾甸子的南部, 两地在气候上有一定的差异, 说明本研究中地理位置不同导致气候条件的差异是影响树木径向生长的重要因素, 表5中潜在蒸发散(PET)对PCA1轴的解释力(72.17%)最大, 而结合表5和附录I可以看出潜在蒸发量和林分密度、蓄积量存在共线性, 说明PET这么高的解释力(72.17%)包含了气候和林分密度、蓄积量的共同作用, 因此气候条件和林分密度、蓄积量共同影响年轮-气候关系。降水对PCA2轴的解释力也达到了显著水平, 为 21.6%。虽然草河口和湾甸子在地理位置上距离并不太远, 但PET在影响年轮-气候关系上仍然有这么高的解释力, 这充分说明气候条件是影响落叶松人工林径向生长对气候变化响应的重要因素。
3.2 林分因子与落叶松径向生长的关系
以前多数研究只探讨了气候条件对年轮-气候关系的影响, 或者在同一地点研究树种、林分因子对年轮-气候关系的影响, 然而这些因素如何共同作用来影响树木径向生长对气候变化的响应还不明确, 以及这些因素的相对作用大小也不太清楚。本文同时研究了气候条件、树种和林分因子对年轮-气候关系的影响, 进一步探讨了这些因素的相对作用大小。一些研究表明, 林龄是影响树木径向生长对气候变化响应的重要因素, 老龄林树木径向生长对水分因子的响应比幼龄林树木更加敏感(Wang et al., 2009; Merian & Lebourgeois, 2011; Schuster & Oberhuber, 2013), 本研究结果与之一致。由图2看出气温升高对林龄较小的落叶松人工林径向生长有促进作用, 而对林龄较大的林分有抑制作用。理论上来说, 在没有水分亏缺的情况下, 气候变暖是有利于树木生长的。径向生长和气温的负相关关系往往是由于气温升高造成的水分亏缺所导致, 这种情况常见于沿气候梯度开展的研究中(常锦峰等, 2009; 曾令兵等, 2012)。本研究发现的不同林龄林分生长与气温的相反关系, 可能反映了中龄林和成熟林之间水分生理的差异。因为中龄林在向成熟林生长发育的过程中, 树木径向生长所需要的营养和水分会增加, 当树龄增大到一定程度后, 随着生物量、树高等的增加, 树木内部的水分运输成为一个主要的限制因子(Ryan & Yoder, 1997)。此外, 林龄较大的树木的蒸腾作用也较幼龄林大得多, 水分需求更大。这两方面因素都导致在同样的降水条件下, 成熟林可能出现生理干旱, 而中龄林则不会。生理干旱会导致气孔过早关闭, 影响树木与外界进行气体交换, 进而限制树木的径向生长。因此本文结果说明, 气候变暖的确导致了水分有效性的下降, 已对林龄较大的林分产生了显著的不利影响。而林龄较小的林分由于水分需求和水分运输困难较小尚未导致生理干旱, 因此气候变暖反而有利于其生长。据此可以预测的是, 随着今后东北地区气候进一步变暖, 可能将逐步影响到林龄更小的林分的生长。对不同针叶树种年轮-气候关系的研究发现, 不同树种对干旱的响应存在差异(Lévesque et al., 2014); 同时有研究认为在较恶劣气候条件下不同树种对气候变化的响应是一致的, 而在相对温和的气候条件下不同树种对气候变化的响应差异较大(Ferrero et al., 2013), 本研究与上述研究结果不尽一致, 由表5可以看出树种对PCA1、2轴的解释力较低, 相对于其他环境、生物因素影响较小。这可能是因为黄花落叶松和日本落叶松同属于落叶松属植物, 分布的纬度差异不大, 种间差异相对不大, 因此对环境变化的适应方式比较相似, 所以在本研究中体现出树种对年轮-气候关系的影响相对于其他因素较小。林分密度会影响树木径向生长对气候变化的响应(Rais et al., 2014b), 通过疏伐实验发现低密度的林分对干旱的抵抗力和恢复力较强(D’Amato & Palik, 2013), 因为高密度的林分对水分的竞争更加激烈, 因此受水分胁迫更严重, 不利于树木的生长甚至导致死亡。同时有研究表明蓄积量对年轮-气候关系也有一定的影响作用, 因为蓄积量较大的林分对水分需求较大, 进而竞争较激烈, 因此对水分条件的响应更为敏感(Rais et al., 2014a)。在本研究中林分密度和蓄积量对年轮-气候关系也起到了重要的作用, 由表5和附录I可以看出林分密度、蓄积量和气候因子存在共线性, 林分密度和蓄积量对年轮- 气候关系都具有重要的影响(p < 0.01), 所以气候和林分密度、蓄积量共同影响着树木径向生长对气候变化的响应。在气候变化背景下, 成熟林一般蓄积量较大, 对水分需求较大, 容易产生生理干旱, 因此老林龄树木径向生长对水分因子更加敏感, 进而导致密度对林龄较大树木的限制作用更大(Primicia et al., 2015), 所以对人工林进行经营时, 要保证成熟林生长环境中水分充足, 可以通过疏伐、减少竞争来减缓气候变化对树木径向生长的影响。
4 结论
本研究通过对辽东山区落叶松人工林进行群落调查, 运用树木年轮气候学的方法来探讨径向生长与气候变化的关系, 结果表明:1)在气候变化的背景下, 气温的适当升高有利于中龄落叶松人工林的生长, 而成熟落叶松人工林受到生理干旱的限制作用, 在对气候变化的响应中更为脆弱。2)落叶松人工林径向生长对气候变化的响应主要受气候条件和林分因子(密度、林龄和蓄积量)的影响, 其中气候条件(热量和水分的有效性)的影响比林分因子要大。3)对黄花落叶松和日本落叶松人工林进行经营时要因地制宜, 针对不同林龄的落叶松人工林采取不同的经营措施, 尤其对林龄较大的落叶松人工林需要进一步研究, 如何通过疏伐减少对水分的竞争, 以合理应对气候变化的威胁。
致谢 感谢北京林业大学林学院博士生梁鹏鸿、孙晗博士在工作中给予的帮助。

参考文献
Alpine timberline dynamics in relation to climatic variability in the northern Daxing’an Mountains
DOI:10.3969/j.issn.1008-2786.2009.06.010
URL
[本文引用: 2]

高山林线植被对气候变化十分敏感,已成为全球变化研究的热点。研究了大兴安岭北部大白山高山 林线的树木生长和群落璺新动态及其与气候变化、火干扰等因素的关系。结果表明,林线树木的生长对气候变化十分敏感,但其敏感性随着海拔的降低而减弱;在高 海拔,林线树木的径向生长与上年生长季后期(8月)降水负相关,而与上年初秋(9月)温度正相关,这些限制作用随着海拔的降低而逐渐消失;与此相反,低海 拔树木生长与当年冬末春初(3月)的温度负相关,但随着海拔上升这种限制作用消失。分析结果还表明,本地区的群落更新主要受火干扰驱动,而与气候变化没有 显著关系。不同树种在火灾后更新的时间存在差异,这种差异又因海拔的不同而异,反映出不同树种的更新策略及对环境变化适应能力的差异。大兴安岭北部的高山 林线在树木生长对气候变化的敏感性、与气候因子的关系及群落更新动态等方面均与干旱区林线有明显的差异,这些差异与气候条件、树种、更新驱动力等方面的不 同有密切的关系。
大兴安岭北部大白山高山林线动态与气候变化的关系
DOI:10.3969/j.issn.1008-2786.2009.06.010
URL
[本文引用: 2]
高山林线植被对气候变化十分敏感,已成为全球变化研究的热点。研究了大兴安岭北部大白山高山 林线的树木生长和群落璺新动态及其与气候变化、火干扰等因素的关系。结果表明,林线树木的生长对气候变化十分敏感,但其敏感性随着海拔的降低而减弱;在高 海拔,林线树木的径向生长与上年生长季后期(8月)降水负相关,而与上年初秋(9月)温度正相关,这些限制作用随着海拔的降低而逐渐消失;与此相反,低海 拔树木生长与当年冬末春初(3月)的温度负相关,但随着海拔上升这种限制作用消失。分析结果还表明,本地区的群落更新主要受火干扰驱动,而与气候变化没有 显著关系。不同树种在火灾后更新的时间存在差异,这种差异又因海拔的不同而异,反映出不同树种的更新策略及对环境变化适应能力的差异。大兴安岭北部的高山 林线在树木生长对气候变化的敏感性、与气候因子的关系及群落更新动态等方面均与干旱区林线有明显的差异,这些差异与气候条件、树种、更新驱动力等方面的不 同有密切的关系。
Effects of thinning on drought vulnerability and climate response in north temperate forest ecosystems
DOI:10.1890/13-0677.1
URL
PMID:24555305
[本文引用: 1]
Reducing tree densities through silvicultural thinning has been widely advocated as a strategy for enhancing resistance and resilience to drought, yet few empirical evaluations of this approach exist. We examined detailed dendrochronological data from a long-term (>50 years) replicated thinning experiment to determine if density reductions conferred greater resistance and/or resilience to droughts, assessed by the magnitude of stand-level growth reductions. Our results suggest that thinning generally enhanced drought resistance and resilience; however, this relationship showed a pronounced reversal over time in stands maintained at lower tree densities. Specifically, lower-density stands exhibited greater resistance and resilience at younger ages (49 years), yet exhibited lower resistance and resilience at older ages (76 years), relative to higher-density stands. We attribute this reversal to significantly greater tree sizes attained within the lower-density stands through stand development, which in turn increased tree-level water demand during the later droughts. Results from response-function analyses indicate that thinning altered growth-climate relationships, such that higher-density stands were more sensitive to growing-season precipitation relative to lower-density stands. These results confirm the potential of density management to moderate drought impacts on growth, and they highlight the importance of accounting for stand structure when predicting climate-change impacts to forests.
Study on the geographic elements affecting temperature distribution in China
地理要素对我国气温分布影响的数量评价
Methods and protocols for plant community inventory
DOI:10.3724/SP.J.1003.2009.09253
URL
[本文引用: 1]
植物群落是不同植物在长期环境变化中相互作用、相互适应而形成的组合。它提供着人类赖以生存的主要物质资源, 维系着地球生态系统的健康和功能, 也为各种动物和其他生物提供食物来源和栖息地, 是人类生存和发展不可或缺的物质基础, 具有不可替代的作用。我国植物群落类型多样, 在世界上首屈一指, 但我国至今尚没有一次全面和系统的植物群落清查, 不仅影响了人们对我国植物资源的了解、利用和保护, 也不利于我国生态学、环境科学和地理学等相关学科的发展。采用统一的方法体系和技术规范开展我国植物群落的清查工作势在必行, 并具有紧迫性。本文基于作者长期的野外工作实践和国内外的群落调查方法, 首先简要定义了与植物群落清查有关的重要概念, 在此基础上, 论述了调查样地的设置原则和体系、群落清查的技术指标和方法、主要优势种生态属性的测定方法和规范, 并介绍了大样地调查的主要步骤。通过本文的介绍、归纳和总结, 试图为制定我国植物群落清查的技术规范提供基础材料和技术储备。
植物群落清查的主要内容、方法和技术规范
DOI:10.3724/SP.J.1003.2009.09253
URL
[本文引用: 1]
植物群落是不同植物在长期环境变化中相互作用、相互适应而形成的组合。它提供着人类赖以生存的主要物质资源, 维系着地球生态系统的健康和功能, 也为各种动物和其他生物提供食物来源和栖息地, 是人类生存和发展不可或缺的物质基础, 具有不可替代的作用。我国植物群落类型多样, 在世界上首屈一指, 但我国至今尚没有一次全面和系统的植物群落清查, 不仅影响了人们对我国植物资源的了解、利用和保护, 也不利于我国生态学、环境科学和地理学等相关学科的发展。采用统一的方法体系和技术规范开展我国植物群落的清查工作势在必行, 并具有紧迫性。本文基于作者长期的野外工作实践和国内外的群落调查方法, 首先简要定义了与植物群落清查有关的重要概念, 在此基础上, 论述了调查样地的设置原则和体系、群落清查的技术指标和方法、主要优势种生态属性的测定方法和规范, 并介绍了大样地调查的主要步骤。通过本文的介绍、归纳和总结, 试图为制定我国植物群落清查的技术规范提供基础材料和技术储备。
Tree-growth responses across environmental gradients in subtropical Argentinean forests
DOI:10.1007/s11258-013-0254-2
URL
[本文引用: 1]
Subtropical forests in montane ecosystems grow under a wide range of environmental conditions. However, little is known about the growth responses of subtropical trees to climate along ecological gradients. To assess how, and to what extent climate controls tree growth, we analyzed tree responses to climate for 15 chronologies from 4 different species (Schinopsis lorentzii, Juglans australis, Cedrela lilloi, Alnus acuminata) across a variety of environments in subtropical forests from northwestern Argentina (22-28A degrees S, 64-66A degrees W). Using correlation and principal component analysis, site and species differences in tree-growth responses to precipitation and temperature were determined along the elevation gradient from the dry-warm Chaco lowlands to the wet-cool montane Yungas. Our results show that species responses differ according to the severity in climate conditions along the elevation gradient. At sites with unfavorable conditions, mainly located at the extremes of the environmental gradient, responses of different species to climate variations are similar; in contrast, at sites with relatively mild conditions, tree growth displays a large variety of responses reflecting differences in both local environmental conditions and species physiology. Our research suggests that individualistic responses to environmental variability would determine differences in the type and timing of the responses of dominant trees to climate, which ultimately may shift species' assemblages in montane subtropical regions of South America under future climate changes.
Tree growth response to climate change at the deciduous boreal forest ecotone, Ontario, Canada
DOI:10.1139/x05-185 URL [本文引用: 1]
Computer-assisted quality control in tree-ring dating and measurement
Response of tundra ecosystem in southwestern Alaska to Younger-Dryas climatic oscillation
DOI:10.1046/j.1365-2486.2002.00550.x
URL
[本文引用: 1]
Climatic warming during the last glacial 搃nterglacial transition (LGIT) was punctuated by reversals to glacial-like conditions. Palaeorecords of ecosystem change can help document the geographical extent of these events and improve our understanding of biotic sensitivity to climatic forcing. To reconstruct ecosystem and climatic variations during the LGIT, we analyzed lake sediments from southwestern Alaska for fossil pollen assemblages, biogenic-silica content (BSiO 2 %), and organic-carbon content (OC%). Betula shrub tundra replaced herb tundra as the dominant vegetation of the region around 13 600 cal BP (cal BP: 14 C calibrated calendar years before present), as inferred from an increase of Betula pollen percentages from << 5% to >> 20% with associated decreases in Cyperaceae, Poaceae, and Artemisia . At c . 13 000 cal BP, a decrease of Betula pollen from 28 to << 5% suggests that shrub tundra reverted to herb tundra. Shrub tundra replaced herb tundra to resume as the dominant vegetation at 11 600 cal BP. Higher OC% and BSiO 2 % values suggest more stable soils and higher aquatic productivity during shrub-tundra periods than during herb-tundra periods, although pollen changes lagged behind changes in the biogeochemical indicators before c . 13 000 cal BP. Comparison of our palaeoecological data with the ice-core d 18 O record from Greenland reveals strikingly similar patterns from the onset through the termination of the Younger Dryas (YD). This similarity supports the hypothesis that, as in the North Atlantic region, pronounced YD climatic oscillations occurred in the North Pacific region. The rapidity and magnitude of ecological changes at the termination of the YD are consistent with greenhouse experiments and historic photographs demonstrating tundra sensitivity to climatic forcing.
Climatic characteristics in Central Asia based on CRU data
Based on the CRU data in Central Asia during the period of 1971 2000, in this paper the method of linear regression was used to analyze the climate change in Central Asia in recent 30 years. The results showed that the desert regions in Turkmenistan and Uzbekistan were the aridest areas with the highest temperature in Central Asia. Precipitation in Tajikistan and Kyrgyzstan in winter and spring was higher than that in summer and autumn, and temperature range was relatively low. In Kazakhstan, precipitation occurred mainly in summer, and it decreased from the west to the east; the range of temperature was high, and it was warmer in the west than that in the east. The annual range of temperature in Central Asia was low. There was a significant difference of climate between Xinjiang of China and the five countries in Central Asia, precipitation in Xinjiang occurred mainly in the Tianshan Mountains and in summer, where the warming trend was obvious, and the change trend of maximum and minimum temperature was opposite to that in the western part of Central Asia.
基于CRU资料的中亚地区气候特征
Based on the CRU data in Central Asia during the period of 1971 2000, in this paper the method of linear regression was used to analyze the climate change in Central Asia in recent 30 years. The results showed that the desert regions in Turkmenistan and Uzbekistan were the aridest areas with the highest temperature in Central Asia. Precipitation in Tajikistan and Kyrgyzstan in winter and spring was higher than that in summer and autumn, and temperature range was relatively low. In Kazakhstan, precipitation occurred mainly in summer, and it decreased from the west to the east; the range of temperature was high, and it was warmer in the west than that in the east. The annual range of temperature in Central Asia was low. There was a significant difference of climate between Xinjiang of China and the five countries in Central Asia, precipitation in Xinjiang occurred mainly in the Tianshan Mountains and in summer, where the warming trend was obvious, and the change trend of maximum and minimum temperature was opposite to that in the western part of Central Asia.
The importance of phenology for the evaluation of impact of climate change on growth of boreal, temperate and Mediterranean forests ecosystems: An overview
DOI:10.1007/s004840000066
URL
PMID:10993560
[本文引用: 1]
Abstract An overview is presented of the phenological models relevant for boreal coniferous, temperate-zone deciduous and Mediterranean coniferous forest ecosystems. The phenology of the boreal forests is mainly driven by temperature, affecting the timing of the start of the growing season and thereby its duration, and the level of frost hardiness and thereby the reduction of foliage area and photosynthetic capacity by severe frost events. The phenology of temperate-zone forests is also mainly driven by temperature. Since temperate-zone forests are mostly mixed-species deciduous forests, differences in phenological response may affect competition between tree species. The phenology of Mediterranean coniferous forests is mainly driven by water availability, affecting the development of leaf area, rather than the timing of phenological events. These phenological models were subsequently coupled to the process-based forest model FORGRO to evaluate the effect of different climate change scenarios on growth. The results indicate that the phenology of each of the forest types significantly affects the growth response to a given climate change scenario. The absolute responses presented in this study should, however, be used with caution as there are still uncertainties in the phenological models, the growth models, the parameter values obtained and the climate change scenarios used. Future research should attempt to reduce these uncertainties. It is recommended that phenological models that describe the mechanisms by which seasonality in climatic drivers affects the phenological aspects of trees should be developed and carefully tested. Only by using such models may we make an assessment of the impact of climate change on the functioning and productivity of different forest ecosystems.
Tree-ring growth variability in the Austrian Alps: The influence of site, altitude, tree species and climate
DOI:10.1080/03009480701267063
URL
[本文引用: 2]
We present an extensive new network of ring-width chronologies, comprising data from 100 sites within the Austrian Eastern Alps, made up of multiple tree species. Principal components analysis and cluster analysis were used to identify five separate high-frequency tree-growth signals from these data. Tree-growth variability at these sites is explained by site altitude and species differences that moderate the effects of the annually varying climatic forcing on tree growth. Recently developed, two-century long gridded climate data sets for the Greater Alpine Region were used to assess the relationship between climate and tree growth. Tree growth at low altitudes is controlled mainly by spring-summer moisture availability. At high altitudes precipitation is no longer a limiting factor and growth is mostly determined by summer temperature. In the intermediate altitudinal range, we did not find any direct relationships with specific climatic variables. High-altitude chronologies suitable for reconstructing past temperatures and low-altitude chronologies suitable for reconstructing past precipitation were identified.
Increased water-use efficiency does not lead to enhanced tree growth under xeric and mesic conditions
DOI:10.1111/nph.12772
URL
PMID:24635031
[本文引用: 1]
Higher atmospheric CO2 concentrations (ca) can under certain conditions increase tree growth by enhancing photosynthesis, resulting in an increase of intrinsic water-use efficiency (iWUE) in trees. However, the magnitude of these effects and their interactions with changing climatic conditions are still poorly understood under xeric and mesic conditions.We combined radial growth analysis with intra- and interannual 020713C and 020718O measurements to investigate growth and physiological responses of Larix decidua, Picea abies, Pinus sylvestris, Pinus nigra and Pseudotsuga menziesii in relation to rising ca and changing climate at a xeric site in the dry inner Alps and at a mesic site in the Swiss lowlands.iWUE increased significantly over the last 50 yr by 809000929% and varied depending on species, site water availability, and seasons. Regardless of species and increased iWUE, radial growth has significantly declined under xeric conditions, whereas growth has not increased as expected under mesic conditions. Overall, drought-induced stomatal closure has reduced transpiration at the cost of reduced carbon uptake and growth.Our results indicate that, even under mesic conditions, the temperature-induced drought stress has overridden the potential CO2 090004fertilization090005 on tree growth, hence challenging today's predictions of improved forest productivity of temperate forests.
Growth responses of broad-leaf and Korean pine mixed forests at different successional stages to climate change in the Shengshan Nature Reserve of Heilongjiang Province, China
DOI:10.17521/cjpe.2015.0357
URL
[本文引用: 1]
黑龙江黑河为我国红松(Pinus koraiensis)分布的北界,在研究红松林的生长、演替、分布,及其对气候变化的响应上有独特的意义。该文通过研究胜山保护区内阔叶红松林的演替系列(软阔叶林、硬阔叶林、阔叶红松近熟林和成熟林4个阶段),分析了树木径向生长与气候变化的关系在不同演替阶段的差异。结果表明:从演替早期的软阔叶林到晚期的红松成熟林,年表统计特征表明树木径向生长对气候波动的敏感性逐步降低。对年轮-气候关系的分析结果也表明气候对树木径向生长的影响随着演替的进展呈现规律性的变化。上年6月和12月的气温与红松成熟林的径向生长显著正相关,表现出明显的“滞后效应”。红松成熟林的轮宽指数与当年6月气温显著负相关,而与当年6月降水量显著正相关,反映出生长季水分对红松生长的限制。上述这些限制作用均随着演替的进展而增强,但在演替的早期影响不显著。相反,上年6月降水量与软阔叶林的生长显著负相关,但该限制作用在演替的中晚期消失。这些差异反映出随着演替的进展,优势树种对水分的需求逐步提高。滑动相关分析表明研究区近几十年明显的气候干暖化趋势对各林型的生长兼具有利和不利的影响。不同因素综合作用下,软、硬阔叶林阶段生长尚未产生清晰的长期变化趋势。但由于水分对红松林生长的限制作用增强,红松林生长明显下降。今后气候进一步干暖化可能对红松林的生长、恢复演替和分布有不利影响。
黑龙江胜山保护区阔叶红松林不同演替阶段径向生长与气候变化的关系
DOI:10.17521/cjpe.2015.0357
URL
[本文引用: 1]
黑龙江黑河为我国红松(Pinus koraiensis)分布的北界,在研究红松林的生长、演替、分布,及其对气候变化的响应上有独特的意义。该文通过研究胜山保护区内阔叶红松林的演替系列(软阔叶林、硬阔叶林、阔叶红松近熟林和成熟林4个阶段),分析了树木径向生长与气候变化的关系在不同演替阶段的差异。结果表明:从演替早期的软阔叶林到晚期的红松成熟林,年表统计特征表明树木径向生长对气候波动的敏感性逐步降低。对年轮-气候关系的分析结果也表明气候对树木径向生长的影响随着演替的进展呈现规律性的变化。上年6月和12月的气温与红松成熟林的径向生长显著正相关,表现出明显的“滞后效应”。红松成熟林的轮宽指数与当年6月气温显著负相关,而与当年6月降水量显著正相关,反映出生长季水分对红松生长的限制。上述这些限制作用均随着演替的进展而增强,但在演替的早期影响不显著。相反,上年6月降水量与软阔叶林的生长显著负相关,但该限制作用在演替的中晚期消失。这些差异反映出随着演替的进展,优势树种对水分的需求逐步提高。滑动相关分析表明研究区近几十年明显的气候干暖化趋势对各林型的生长兼具有利和不利的影响。不同因素综合作用下,软、硬阔叶林阶段生长尚未产生清晰的长期变化趋势。但由于水分对红松林生长的限制作用增强,红松林生长明显下降。今后气候进一步干暖化可能对红松林的生长、恢复演替和分布有不利影响。
Response of radial growth of Pinus koraiensis in broad-leaved Korean pine forests with different latitudes to climatical factors.
DOI:10.13287/j.1001-9332.201605.020
URL
[本文引用: 1]
运用树木年轮气候学方法,研究了不同纬度阔叶红松林红松径向生长趋势及其与气候因子的关系,以期揭示不同纬度红松径向生长对气候因子响应的差异以及气候变化影响下红松的动态特征、适应性及敏感性.结果表明:不同纬度红松径向生长对当地气候因子的响应存在差异,最南部的白石砬子自然保护区红松径向生长与生长季的平均相对湿度呈显著正相关,与平均最高气温呈显著负相关.中部的长白山自然保护区的低海拔区域红松径向生长与生长季的水分因子(降水、相对湿度和帕尔默干旱指数PDSI)呈显著正相关,而与平均最高气温呈显著负相关;凉水自然保护区红松径向生长与生长季的水分因子(相对湿度和PDSI)呈显著正相关,与气温因子(平均气温和平均最高气温)呈显著负相关.而最北部的胜山自然保护区红松径向生长则与大部分月份的气温因子呈显著正相关.当年6月气候因子是影响所有纬度红松径向生长的关键气候因子,4个样地都与当年6月平均最高气温呈显著负相关.在气温不断上升的近40年,最南部的红松径向生长呈显著下降趋势,最北部呈显著上升趋势,中部的变化不显著.如果未来气温升高而降水不变,红松分布区可能缩小.
不同纬度阔叶红松林红松径向生长对气候因子的响应
DOI:10.13287/j.1001-9332.201605.020
URL
[本文引用: 1]
运用树木年轮气候学方法,研究了不同纬度阔叶红松林红松径向生长趋势及其与气候因子的关系,以期揭示不同纬度红松径向生长对气候因子响应的差异以及气候变化影响下红松的动态特征、适应性及敏感性.结果表明:不同纬度红松径向生长对当地气候因子的响应存在差异,最南部的白石砬子自然保护区红松径向生长与生长季的平均相对湿度呈显著正相关,与平均最高气温呈显著负相关.中部的长白山自然保护区的低海拔区域红松径向生长与生长季的水分因子(降水、相对湿度和帕尔默干旱指数PDSI)呈显著正相关,而与平均最高气温呈显著负相关;凉水自然保护区红松径向生长与生长季的水分因子(相对湿度和PDSI)呈显著正相关,与气温因子(平均气温和平均最高气温)呈显著负相关.而最北部的胜山自然保护区红松径向生长则与大部分月份的气温因子呈显著正相关.当年6月气候因子是影响所有纬度红松径向生长的关键气候因子,4个样地都与当年6月平均最高气温呈显著负相关.在气温不断上升的近40年,最南部的红松径向生长呈显著下降趋势,最北部呈显著上升趋势,中部的变化不显著.如果未来气温升高而降水不变,红松分布区可能缩小.
Size-mediated climat-growth relationships in temperate forests: A multi-species analysis
DOI:10.1016/j.foreco.2011.01.019
URL
[本文引用: 1]
In most dendrochronological studies, climate rowth relationships are established on dominant trees to minimize non-climatic signals. However, response to environmental factors may be affected by tree-size, which begs the question of the representativeness of dominant trees on the stand level. To highlight the variations in climate rowth relationships among sizes and species, under a wide range of ecological conditions (climate and soil properties), 61 pure even-aged stands were sampled across France. At each stand, two tree-ring chronologies were established from 10 big- to 10 small-diameter trees. Our objectives were, (1) to assess variations in climate sensitivity between the two size-diameter classes, and (2) to investigate the role of species and ecological conditions on these variations. The climate-growth relationships were evaluated from 122 tree-ring chronologies (1 220 trees) through extreme growth years and correlation function analyses. Sensitivity to climate of shade-intolerant and moderately shade-tolerant species ( Picea abies (L.) Karst., Pinus sylvestris L. and Quercus petraea (Matt.) Liebl.) remained constant between the size-diameter classes for both temperature and hydric balance, while the shade-tolerant species Abies alba Mill. and Fagus sylvatica L. displayed significant differences, with larger trees being more sensitive to summer drought than smaller trees. This difference increased with increasing climatic xericity. Our results suggest that, for shade-tolerant species, (1) big trees could be more sensitive to climatic change especially under xeric climate, and (2) future tree ring studies should include trees stratified by size to produce unbiased estimation of sensitivity to climate.
An improved method of constructing a database of monthly climate observations and associated high-resolution grids
DOI:10.1002/(ISSN)1097-0088 URL [本文引用: 1]
Age, competition, disturbance and elevation effects on tree and stand growth response of primary Picea abies forest to climate.
Application of CRU dataset to calculation of ET0 of Heilongjiang Province
CRU数据集在黑龙江省ET0计算中的应用
Growth reaction patterns of tree height, diameter, and volume of Douglas-fir (Pseudotsuga menziesii( Mirb.) Franco) under acute drought stress in Southern Germany.
Influence of initial plant density on sawn timber properties for Douglas-fir (Pseudotsuga menziesii( Mirb.) Franco).
Hydraulic limits to tree height and tree growth
DOI:10.2307/1313077 URL [本文引用: 1]
Drought sensitivity of three co-occurring conifers within a dry inner alpine environment
DOI:10.1007/s00468-012-0768-6 URL [本文引用: 1]
Spatial and age-dependent tree-ring growth responses of Larix gmelinii to climate in northeastern China.
Climatic control of primary forest structure and DBH-height allometry in Northeast China
Past global changes and their significance for the future
DOI:10.1023/A:1011133708229 URL [本文引用: 1]
Application of tree ring analysis to the study on environment variation
树木年轮分析在环境变化研究中的应用
A test of BIOME-BGC with dendrochronology for forests along the altitudinal gradient of Mt. Changbai in Northeast China
Tree-ring widths are good proxies of annual variation in forest productivity in temperate forests
DOI:10.1038/s41598-017-02022-6 URL [本文引用: 1]
Age-dependent tree-ring growth responses to climate in Qilian juniper (Sabina przewalskii Kom.).
The stoichiometric characteristics of C, N and P in soil and root of larch (Larix spp.) plantation at different stand ages in mountainous region of eastern Liaoning Province, China.
辽东山区不同林龄落叶松人工林土壤-根系C:N:P生态化学计量特征
Alpine timberline ecotone tree growth in relation to climatic variability for Picea crassifolia forests in the middle Qilian Mountains, northwestern China.
祁连山中段青海云杉高山林线交错区树轮宽度与气候变化的关系

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}