A review study on past 40 years of research on effects of tropospheric O3 on belowground structure, functioning, and processes of trees: a linkage with potential ecological implications
1
2016
... (2)加强地表O3污染对地下生态过程的影响研究: 地下部分是生态系统水分供给、养分循环、碳分配的核心环节, 探究地下生态系统对地表O3浓度升高的响应对于准确理解地表O3浓度升高对整个生态系统的影响具有重要意义.事实上, 高浓度O3对地下部分的干扰比地上部分更明显且更早出现(Hofstra et al., 1981).然而, 目前有关O3对地下生态过程动态变化过程的研究还十分缺乏, 其响应机制尚不明确.已有的研究多集中在地表O3浓度升高对地下碳分配、根系生长和生物量的影响上, 少部分涉及凋落物、根系分泌物、土壤呼吸、微生物活性和结构等(Agathokleous et al., 2016), 而地表O3浓度升高对土壤微食物网的影响及其反馈几乎都没有涉及(Grulke & Heath, 2020), 更无法将土壤微生物过程(如微生物活性、生物量、群落组成、多样性和功能)与土壤关键生物地球化学循环过程, 以及与植物地上生态过程关联起来(Andersen, 2003; 冯兆忠等, 2018).未来的研究应加强O3对地下生态过程的长期定位研究, 特别是O3对地下碳分配动态、土壤微生物动态及根际氮循环的研究, 量化地下生态过程对O3响应的程度及其对生态系统长期稳定的反馈. ...
Understanding and improving global crop response to ozone pollution
2
2017
... 地表臭氧(O3)特指距离地球表面15 km范围内的近地层O3, 除少量来自平流层大气传输外, 其余大部分是氮氧化物(NOx)、非甲烷类挥发性有机化合物(NMVOCs)、一氧化碳(CO)和甲烷(CH4)等前体物在强烈光照下发生光化学反应而产生(冯兆忠等, 2018).自工业革命以来, 伴随城市化进程加快和化石燃料的过度燃烧, 地表O3浓度在世界范围内普遍升高.近年来, 模型和观测数据都表明地表O3浓度小时峰值在欧洲和北美等地已有所下降, 但亚洲地区仍呈不断上升的趋势(Cooper et al., 2014).中国国家环境监测网络2013-2017年的数据表明, 过去5年中国地表O3浓度以每年3 μg·kg-1的速率攀升(Wang et al., 2019), 目前全国地表O3浓度每日8 h滑动平均最大值(MDA8)的年平均值已达(41.2 ± 6.3) μg·kg-1, 尤其华北平原、长江三角洲和珠江三角洲等地(MDA8峰值年平均值已处于60-70 μg·kg-1), 地表O3污染形势十分严峻(Lu et al., 2018).地表O3是具有植物毒性的气体污染物, 具有强氧化性.高浓度地表O3或长期低浓度O3能诱导植物叶片出现可见损伤, 加速植物衰老, 降低光合作用, 改变碳分配, 抑制植物生长和生物量积累, 影响作物品质和产量, 最终降低生态系统碳汇能力, 改变生态系统结构和功能(Ashmore, 2005; Ainsworth, 2017; 高峰, 2018). ...
... 通常认为, 植物通过气孔防御和非气孔防御对O3胁迫做出响应.气孔作为叶片与外界气体交换的“门户”, 是O3进入植物体内的主要通道, 控制着植物蒸腾、呼吸和光合等重要生理过程.正常光热条件下, 气孔处于打开状态, 但高浓度O3可以直接影响调节气孔开张的保卫细胞上钾离子浓度, 改变细胞膨压, 诱导气孔关闭, 引发避逆性的防御响应(Wieser et al., 2002).因此, 气孔闭合被认为是植物抵御O3胁迫的第一道防线(Ainsworth, 2017).然而, 气孔对O3的响应(关闭)也并不是始终如一的.研究发现随着O3暴露时间的延长, O3诱导的气孔关闭会出现滞后甚至失灵的现象(Hoshika et al., 2012), 从而导致大量O3进入细胞间隙, 气孔防御机制 失效. ...
The influence of rising tropospheric carbon dioxide and ozone on plant productivity
1
2020
... (1)开展O3-FACE研究: 现有研究多为OTC系统下的实验, OTC气室内“微气候” (如温度、水分、光照和风力等生态因素)与外界环境状态存在明显差异, 这些生态因子的变化可能影响植物应对O3的响应, 缓解或加重O3污染对农作物产量或森林植被生产力的损伤(Long et al., 2005; Feng et al., 2018a).全球模型评估结果显示, OTC实验结果低估了O3对作物产量损失的影响(van Dingenen et al., 2009).最近, Feng等(2018b)也发现, 同样浓度的O3对OTC实验下小麦和水稻产量的影响要低于FACE实验; 而对大豆产量的影响则相反.因此, OTC实验结果并不能完全应用于实际O3区域风险的评估.O3-FACE平台是未来研究实际大田环境下O3浓度升高对农林生态系统影响的主要依托平台.为减少区域O3风险评估的不确定性, 未来应加大O3-FACE平台的实验研究.考虑到目前已开展的FACE实验多坐落在温带地区, 主要探究温带植被或作物对高浓度O3的响应, 较少涉及热带和寒带植物或其他区域典型生态系统(Ainsworth et al., 2020).最理想的情况是, 不同区域、不同生态系统类型共同开展O3-FACE实验研究(Ainsworth et al., 2012). ...
The effects of tropospheric ozone on net primary productivity and implications for climate change
3
2012
... O3不仅抑制植物生长, 还导致农作物大量减产(Fuhrer, 2009; Ainsworth et al., 2012; Feng et al., 2015a; Singh & Agrawal, 2017).有关O3对作物产量的影响已经在北美、欧洲和亚洲多地的几十种作物上有所研究报道, 包括对O3敏感的小麦、西瓜(Citrullus lanatus)、豆类、棉花(Gossypium hirsutum)、萝卜(Raphanus sativus)、番茄(Lycopersicon esclentum)、洋葱(Allium cepa)、大豆(Glycine max)和莴苣(Lactuca sativa), 对O3适度敏感的甜菜(Beta vulgaris)、马铃薯(Solanum tuberosum)、油菜(Brassica napus)、烟草(Nicotiana tabacum)、水稻、玉米(Zea mays)、葡萄和西兰花(Brassica oleracea), 和对O3耐受的作物大麦(Hordeum vulgare)、李(Prunus salicina)和草莓(Fragaria ananassa)等(Mills et al., 2007; Booker et al., 2009; Feng & Kobayashi, 2009; Avnery et al., 2011; Broberg et al., 2015).通过汇总406个控制实验的数据进行meta分析发现, 与活性炭过滤环境(≤26 μg·kg-1)相比, 当前环境O3浓度(平均值40 μg·kg-1)可导致主要粮食作物(小麦、水稻、大豆和马铃薯等)减产10%左右; 未来地表O3浓度升高情景下(51-71 μg·kg-1), 作物产量将进一步降低10%- 20% (Feng & Kobayashi, 2009).最近, 有关美国、法国、印度、中国和俄罗斯等小麦主产国的模型评估数据显示, 2010-2012年间高浓度O3浓度造成北半球和南半球小麦平均减产9.9%和6.2%, 抵消了因水肥管理或改良品种等措施对其增产所做的努力(Mills et al., 2018).我国这方面的研究起步较晚, 但发展迅速.从20世纪90年代末开始, 学者先后利用OTC和O3-FACE系统研究了地表O3浓度升高对我国主要粮食作物和蔬菜的影响, 涉及品种包括冬小麦、水稻、大豆、玉米、油菜、菠菜(Spinacia oleracea)和青菜(Brassica rapa)等(冯兆忠等, 2018).研究显示, 高浓度O3暴露下农作物均出现不同程度减产, 不同浓度O3对水稻、小麦、大豆等不同品种作物的减产幅度为10%-60% (贾一磊, 2016; 曹嘉晨等, 2017), 对油菜、菠菜和青菜等蔬菜生物产量的降幅为4.0%-89% (白月明等, 2002; 郭建平等, 2003). ...
... 地表O3浓度升高对植物光合作用、生长和生物量积累、物质分配等过程的连锁负效应导致陆地生态系统生产力和固碳能力降低(Reich, 1987; Wittig et al., 2009; Ainsworth et al., 2012), 削弱了自然生态系统的碳汇能力(Felzer et al., 2005; Sitch et al., 2007; Collins et al., 2010).基于大量OTCs控制实验下北半球中纬度北方温带树种的meta分析发现, O3浓度升高导致树木的净光合作用和总生物量分别降低11%和7% (Wittig et al., 2009).最近, 有关中国热带、亚热带和温带树种的meta分析则发现, O3浓度升高导致中国木本植物的净光合作用和总生物量分别降低28%和14% (Li et al., 2017). ...
... (1)开展O3-FACE研究: 现有研究多为OTC系统下的实验, OTC气室内“微气候” (如温度、水分、光照和风力等生态因素)与外界环境状态存在明显差异, 这些生态因子的变化可能影响植物应对O3的响应, 缓解或加重O3污染对农作物产量或森林植被生产力的损伤(Long et al., 2005; Feng et al., 2018a).全球模型评估结果显示, OTC实验结果低估了O3对作物产量损失的影响(van Dingenen et al., 2009).最近, Feng等(2018b)也发现, 同样浓度的O3对OTC实验下小麦和水稻产量的影响要低于FACE实验; 而对大豆产量的影响则相反.因此, OTC实验结果并不能完全应用于实际O3区域风险的评估.O3-FACE平台是未来研究实际大田环境下O3浓度升高对农林生态系统影响的主要依托平台.为减少区域O3风险评估的不确定性, 未来应加大O3-FACE平台的实验研究.考虑到目前已开展的FACE实验多坐落在温带地区, 主要探究温带植被或作物对高浓度O3的响应, 较少涉及热带和寒带植物或其他区域典型生态系统(Ainsworth et al., 2020).最理想的情况是, 不同区域、不同生态系统类型共同开展O3-FACE实验研究(Ainsworth et al., 2012). ...
Source-sink balance and carbon allocation below ground in plants exposed to ozone: tansley review
2
2003
... 此外, 虽然地下生态系统并不是直接暴露在O3环境下, 但大量证据表明O3对地上生态系统的影响将会转移至地下(Andersen, 2003; Lindroth, 2010).如高浓度O3环境下, 植物分配到根的生物量将会大大减少, 基于100多组实验数据的meta分析研究表明O3暴露导致生物量分配到地下的量平均减少5.6% (Grantz et al., 2006).另外, 凋落物分解是生态系统营养循环的关键过程.通过影响凋落物的数量、质量和分解速率, 高浓度地表O3会间接改变土壤微生物群落活性、组成和结构, 土壤化学属性及土壤营养循环(Larson et al., 2002; Fuhrer et al., 2016).然而, 比较遗憾的是, 目前有关O3对地下生态过程影响的研究仅集中在农田生态系统, 仅有少部分研究涉及森林土壤(主要来自Aspen FACE)或草地生态系统(ICP Vegetation, 2013).而且, 由于凋落物种类、化学成分、分解者种群、土壤和大气环境条件的不同, 高浓度O3对凋落物分解动态的影响还存在很大争议(Gessner et al., 2010; Fuhrer et al., 2016; Grulke & Heath, 2020).例如, 研究发现O3对欧洲白桦(Betula pendula)叶片凋落物质量和损失率有明显影响(Kasurinen et al., 2006, 2007), 但不影响欧洲山毛榉凋落物的分解速率(Schloter et al., 2005).有关地表O3浓度升高对地下生态系统结构和功能的影响还有待进一步探究. ...
... (2)加强地表O3污染对地下生态过程的影响研究: 地下部分是生态系统水分供给、养分循环、碳分配的核心环节, 探究地下生态系统对地表O3浓度升高的响应对于准确理解地表O3浓度升高对整个生态系统的影响具有重要意义.事实上, 高浓度O3对地下部分的干扰比地上部分更明显且更早出现(Hofstra et al., 1981).然而, 目前有关O3对地下生态过程动态变化过程的研究还十分缺乏, 其响应机制尚不明确.已有的研究多集中在地表O3浓度升高对地下碳分配、根系生长和生物量的影响上, 少部分涉及凋落物、根系分泌物、土壤呼吸、微生物活性和结构等(Agathokleous et al., 2016), 而地表O3浓度升高对土壤微食物网的影响及其反馈几乎都没有涉及(Grulke & Heath, 2020), 更无法将土壤微生物过程(如微生物活性、生物量、群落组成、多样性和功能)与土壤关键生物地球化学循环过程, 以及与植物地上生态过程关联起来(Andersen, 2003; 冯兆忠等, 2018).未来的研究应加强O3对地下生态过程的长期定位研究, 特别是O3对地下碳分配动态、土壤微生物动态及根际氮循环的研究, 量化地下生态过程对O3响应的程度及其对生态系统长期稳定的反馈. ...
Assessing the future global impacts of ozone on vegetation
2
2005
... 地表臭氧(O3)特指距离地球表面15 km范围内的近地层O3, 除少量来自平流层大气传输外, 其余大部分是氮氧化物(NOx)、非甲烷类挥发性有机化合物(NMVOCs)、一氧化碳(CO)和甲烷(CH4)等前体物在强烈光照下发生光化学反应而产生(冯兆忠等, 2018).自工业革命以来, 伴随城市化进程加快和化石燃料的过度燃烧, 地表O3浓度在世界范围内普遍升高.近年来, 模型和观测数据都表明地表O3浓度小时峰值在欧洲和北美等地已有所下降, 但亚洲地区仍呈不断上升的趋势(Cooper et al., 2014).中国国家环境监测网络2013-2017年的数据表明, 过去5年中国地表O3浓度以每年3 μg·kg-1的速率攀升(Wang et al., 2019), 目前全国地表O3浓度每日8 h滑动平均最大值(MDA8)的年平均值已达(41.2 ± 6.3) μg·kg-1, 尤其华北平原、长江三角洲和珠江三角洲等地(MDA8峰值年平均值已处于60-70 μg·kg-1), 地表O3污染形势十分严峻(Lu et al., 2018).地表O3是具有植物毒性的气体污染物, 具有强氧化性.高浓度地表O3或长期低浓度O3能诱导植物叶片出现可见损伤, 加速植物衰老, 降低光合作用, 改变碳分配, 抑制植物生长和生物量积累, 影响作物品质和产量, 最终降低生态系统碳汇能力, 改变生态系统结构和功能(Ashmore, 2005; Ainsworth, 2017; 高峰, 2018). ...
... 地表O3的发现最早可追溯到1839年(Leeds, 1880), 但直到1945年“洛杉矶烟雾”爆发后, 地表O3生态效应的研究才逐渐进入学者的视野(Rogers et al., 1956).经过近百年的发展, 涉及地表O3对植物、土壤及生态系统影响的研究, 已经遍及世界上大多数国家, 发表了2 500多篇期刊论文和十余本英文专著.到目前为止, O3表观伤害症状的描述及症状等级的鉴定(如叶片萎黄、褪绿和坏死等典型症状分类), O3伤害机理的探究(如O3降低光合速率的气孔与非气孔因素等), O3对植物叶片(光合作用、气孔导度及抗氧化系统等), 个体(生物量、产量或品质), 种群(种间关系、群落构成)及生态系统(生产力和固碳能力等)的影响, 及O3污染防治对策的探讨(前体物减排措施、化学防护剂的应用和作物栽培管理模式的筛选)等方面都进行了大量相关研究(Ashmore, 2005; Grulke & Heath, 2020).然而地表O3浓度升高对陆地生态系统水平碳、氮和水循环、营养交互、群落更替及地下生态系统动态变化过程(如根系功能变化、根际过程机制、土壤微生物活性及反馈等)方面的研究仍鲜有报道(Fuhrer et al., 2016).通过系统梳理和总结目前国内外研究成果, 本文对地表O3浓度升高对生态系统影响这一领域的研究历史、研究方法、评估指标、重大进展和存在的主要问题进行了回顾分析, 并对未来研究进行了展望. ...
Global crop yield reductions due to surface ozone exposure: 1. Year 2000 crop production losses and economic damage
2
2011
... O3不仅抑制植物生长, 还导致农作物大量减产(Fuhrer, 2009; Ainsworth et al., 2012; Feng et al., 2015a; Singh & Agrawal, 2017).有关O3对作物产量的影响已经在北美、欧洲和亚洲多地的几十种作物上有所研究报道, 包括对O3敏感的小麦、西瓜(Citrullus lanatus)、豆类、棉花(Gossypium hirsutum)、萝卜(Raphanus sativus)、番茄(Lycopersicon esclentum)、洋葱(Allium cepa)、大豆(Glycine max)和莴苣(Lactuca sativa), 对O3适度敏感的甜菜(Beta vulgaris)、马铃薯(Solanum tuberosum)、油菜(Brassica napus)、烟草(Nicotiana tabacum)、水稻、玉米(Zea mays)、葡萄和西兰花(Brassica oleracea), 和对O3耐受的作物大麦(Hordeum vulgare)、李(Prunus salicina)和草莓(Fragaria ananassa)等(Mills et al., 2007; Booker et al., 2009; Feng & Kobayashi, 2009; Avnery et al., 2011; Broberg et al., 2015).通过汇总406个控制实验的数据进行meta分析发现, 与活性炭过滤环境(≤26 μg·kg-1)相比, 当前环境O3浓度(平均值40 μg·kg-1)可导致主要粮食作物(小麦、水稻、大豆和马铃薯等)减产10%左右; 未来地表O3浓度升高情景下(51-71 μg·kg-1), 作物产量将进一步降低10%- 20% (Feng & Kobayashi, 2009).最近, 有关美国、法国、印度、中国和俄罗斯等小麦主产国的模型评估数据显示, 2010-2012年间高浓度O3浓度造成北半球和南半球小麦平均减产9.9%和6.2%, 抵消了因水肥管理或改良品种等措施对其增产所做的努力(Mills et al., 2018).我国这方面的研究起步较晚, 但发展迅速.从20世纪90年代末开始, 学者先后利用OTC和O3-FACE系统研究了地表O3浓度升高对我国主要粮食作物和蔬菜的影响, 涉及品种包括冬小麦、水稻、大豆、玉米、油菜、菠菜(Spinacia oleracea)和青菜(Brassica rapa)等(冯兆忠等, 2018).研究显示, 高浓度O3暴露下农作物均出现不同程度减产, 不同浓度O3对水稻、小麦、大豆等不同品种作物的减产幅度为10%-60% (贾一磊, 2016; 曹嘉晨等, 2017), 对油菜、菠菜和青菜等蔬菜生物产量的降幅为4.0%-89% (白月明等, 2002; 郭建平等, 2003). ...
... 基于OTCs和O3-FACE平台下单个站点的控制实验可探究O3对植物个体、群落或生态系统的影响, 但将站点或样地尺度的研究外推到区域或全球, 评估O3污染区域风险的研究主要通过响应关系和模型.目前, 其研究领域主要集中在地表O3污染对农作物产量和森林生产力等方面.早在1994年, Chameides等(1994)就利用经济模型初步预测O3浓度升高可能导致2025年全球30%-75%的作物面临产量降低的风险.基于O3前体物排放、化学反应传输模型和M12、AOT40指标的评估显示, 2000年全球O3污染引起全球小麦、玉米和大豆产量平均降低2.2%-15%, 每年因上述作物产量降低所导致的经济损失达110-180亿美元(Avnery et al., 2011).最近, 基于PODY评估指标的研究则表明, O3对小麦造成的产量损失北半球(9.9%)明显高于南半球(6.2%), 平均产量损失最高的地区主要位于北半球暖温带湿润地区(12.5%)、热带湿热(12.5%)和热带潮湿气候区(17.2%)(Mills et al., 2018).在区域尺度上, 基于1990年O3浓度数据, Wang和Mauzerall (2004)利用M7和M12指标评估了O3对中国、日本和韩国主要农作物产量的影响, 结果显示O3污染造成小麦、玉米、棉花减产1%-9%, 大豆减产23%-27%, 造成2.4-35亿美元的经济损失, 在维持现有的污染物排放、管理措施和农业生产技术不变的情况下, 预计到2020年小麦、玉米、棉花减产幅度为2%-15%, 大豆为28%-35%.最近, 利用AOT40指标的研究显示, 2014年当前O3浓度导致中国农作物产量损失7 840万t, 小麦、水稻和玉米相对减产量分别为8.5%-14%、3.9%-15%和2.2%-5.5% (Lin et al., 2018).而只聚焦于长江三角洲地区的研究则发现, 2015年区域O3污染分别造成小麦和水稻减产9.9%-36.1%和7.3%- 23.9% (Zhao et al., 2018).虽然国内也有少量研究建立了PODY与个别树种或作物相对生物量或相对产量之间的通量响应关系(Feng et al., 2012; Hu et al., 2015; Gao et al., 2017), 但并没有研究利用PODY指标评估区域O3污染对农作物产量损失的影响. ...
O3浓度增加对油菜影响的诊断试验研究
1
2002
... O3不仅抑制植物生长, 还导致农作物大量减产(Fuhrer, 2009; Ainsworth et al., 2012; Feng et al., 2015a; Singh & Agrawal, 2017).有关O3对作物产量的影响已经在北美、欧洲和亚洲多地的几十种作物上有所研究报道, 包括对O3敏感的小麦、西瓜(Citrullus lanatus)、豆类、棉花(Gossypium hirsutum)、萝卜(Raphanus sativus)、番茄(Lycopersicon esclentum)、洋葱(Allium cepa)、大豆(Glycine max)和莴苣(Lactuca sativa), 对O3适度敏感的甜菜(Beta vulgaris)、马铃薯(Solanum tuberosum)、油菜(Brassica napus)、烟草(Nicotiana tabacum)、水稻、玉米(Zea mays)、葡萄和西兰花(Brassica oleracea), 和对O3耐受的作物大麦(Hordeum vulgare)、李(Prunus salicina)和草莓(Fragaria ananassa)等(Mills et al., 2007; Booker et al., 2009; Feng & Kobayashi, 2009; Avnery et al., 2011; Broberg et al., 2015).通过汇总406个控制实验的数据进行meta分析发现, 与活性炭过滤环境(≤26 μg·kg-1)相比, 当前环境O3浓度(平均值40 μg·kg-1)可导致主要粮食作物(小麦、水稻、大豆和马铃薯等)减产10%左右; 未来地表O3浓度升高情景下(51-71 μg·kg-1), 作物产量将进一步降低10%- 20% (Feng & Kobayashi, 2009).最近, 有关美国、法国、印度、中国和俄罗斯等小麦主产国的模型评估数据显示, 2010-2012年间高浓度O3浓度造成北半球和南半球小麦平均减产9.9%和6.2%, 抵消了因水肥管理或改良品种等措施对其增产所做的努力(Mills et al., 2018).我国这方面的研究起步较晚, 但发展迅速.从20世纪90年代末开始, 学者先后利用OTC和O3-FACE系统研究了地表O3浓度升高对我国主要粮食作物和蔬菜的影响, 涉及品种包括冬小麦、水稻、大豆、玉米、油菜、菠菜(Spinacia oleracea)和青菜(Brassica rapa)等(冯兆忠等, 2018).研究显示, 高浓度O3暴露下农作物均出现不同程度减产, 不同浓度O3对水稻、小麦、大豆等不同品种作物的减产幅度为10%-60% (贾一磊, 2016; 曹嘉晨等, 2017), 对油菜、菠菜和青菜等蔬菜生物产量的降幅为4.0%-89% (白月明等, 2002; 郭建平等, 2003). ...
O3浓度增加对油菜影响的诊断试验研究
1
2002
... O3不仅抑制植物生长, 还导致农作物大量减产(Fuhrer, 2009; Ainsworth et al., 2012; Feng et al., 2015a; Singh & Agrawal, 2017).有关O3对作物产量的影响已经在北美、欧洲和亚洲多地的几十种作物上有所研究报道, 包括对O3敏感的小麦、西瓜(Citrullus lanatus)、豆类、棉花(Gossypium hirsutum)、萝卜(Raphanus sativus)、番茄(Lycopersicon esclentum)、洋葱(Allium cepa)、大豆(Glycine max)和莴苣(Lactuca sativa), 对O3适度敏感的甜菜(Beta vulgaris)、马铃薯(Solanum tuberosum)、油菜(Brassica napus)、烟草(Nicotiana tabacum)、水稻、玉米(Zea mays)、葡萄和西兰花(Brassica oleracea), 和对O3耐受的作物大麦(Hordeum vulgare)、李(Prunus salicina)和草莓(Fragaria ananassa)等(Mills et al., 2007; Booker et al., 2009; Feng & Kobayashi, 2009; Avnery et al., 2011; Broberg et al., 2015).通过汇总406个控制实验的数据进行meta分析发现, 与活性炭过滤环境(≤26 μg·kg-1)相比, 当前环境O3浓度(平均值40 μg·kg-1)可导致主要粮食作物(小麦、水稻、大豆和马铃薯等)减产10%左右; 未来地表O3浓度升高情景下(51-71 μg·kg-1), 作物产量将进一步降低10%- 20% (Feng & Kobayashi, 2009).最近, 有关美国、法国、印度、中国和俄罗斯等小麦主产国的模型评估数据显示, 2010-2012年间高浓度O3浓度造成北半球和南半球小麦平均减产9.9%和6.2%, 抵消了因水肥管理或改良品种等措施对其增产所做的努力(Mills et al., 2018).我国这方面的研究起步较晚, 但发展迅速.从20世纪90年代末开始, 学者先后利用OTC和O3-FACE系统研究了地表O3浓度升高对我国主要粮食作物和蔬菜的影响, 涉及品种包括冬小麦、水稻、大豆、玉米、油菜、菠菜(Spinacia oleracea)和青菜(Brassica rapa)等(冯兆忠等, 2018).研究显示, 高浓度O3暴露下农作物均出现不同程度减产, 不同浓度O3对水稻、小麦、大豆等不同品种作物的减产幅度为10%-60% (贾一磊, 2016; 曹嘉晨等, 2017), 对油菜、菠菜和青菜等蔬菜生物产量的降幅为4.0%-89% (白月明等, 2002; 郭建平等, 2003). ...
大豆对臭氧、二氧化碳及其复合效应的响应
1
2005
... 除单因子O3胁迫外, 由于全球气候变化和极端气候常态化, 植物经常受到O3和其他环境因子交互作用的影响(如O3与CO2、升温、干旱或氮沉降等)(Schmadel-Hagebölling et al., 1998; Ollinger et al., 2002; Eltayeb et al., 2006; Ghimire et al., 2018).然而, 植物应对O3与其他因子交互作用的响应并不是两者或单独作用的简单叠加, 往往取决于植物种类、生长时期、对特定环境因子的敏感性、胁迫浓度及时间, 以及具体的微气候条件(列淦文和薛立, 2014).一方面, 通过诱导气孔关闭、限制叶片O3吸收(CO2浓度升高/干旱胁迫), 或提高碳同化速率、促进植物光合作用(CO2浓度升高/氮增加), CO2浓度升高、干旱胁迫和氮沉降经常被报道可增强植物抵御O3的能力, 减缓O3伤害(白月明等, 2005; Handley & Grulke, 2008; Gao et al., 2017).另一方面, 与之相反, 也有研究发现长期高浓度CO2或干旱降低气孔导度的同时, 会引起植物蒸腾速率、光合产物合成和积累的降低(McDonald et al., 2002; Pollastrini et al., 2010); 而超过植物承载阈值的高氮负荷也会对植物的光合生理活性产生负效应(Yamaguchi et al., 2010), 进而加重植物在复合环境下的碳损失、水分蒸散损失和自由基积累, 对植物产生协同伤害(Karnosky et al., 2005; 列淦文和薛立, 2014).高峰(2018)认为O3与其他环境胁迫因子出现的先后次序也会影响植物对复合作用的响应. ...
大豆对臭氧、二氧化碳及其复合效应的响应
1
2005
... 除单因子O3胁迫外, 由于全球气候变化和极端气候常态化, 植物经常受到O3和其他环境因子交互作用的影响(如O3与CO2、升温、干旱或氮沉降等)(Schmadel-Hagebölling et al., 1998; Ollinger et al., 2002; Eltayeb et al., 2006; Ghimire et al., 2018).然而, 植物应对O3与其他因子交互作用的响应并不是两者或单独作用的简单叠加, 往往取决于植物种类、生长时期、对特定环境因子的敏感性、胁迫浓度及时间, 以及具体的微气候条件(列淦文和薛立, 2014).一方面, 通过诱导气孔关闭、限制叶片O3吸收(CO2浓度升高/干旱胁迫), 或提高碳同化速率、促进植物光合作用(CO2浓度升高/氮增加), CO2浓度升高、干旱胁迫和氮沉降经常被报道可增强植物抵御O3的能力, 减缓O3伤害(白月明等, 2005; Handley & Grulke, 2008; Gao et al., 2017).另一方面, 与之相反, 也有研究发现长期高浓度CO2或干旱降低气孔导度的同时, 会引起植物蒸腾速率、光合产物合成和积累的降低(McDonald et al., 2002; Pollastrini et al., 2010); 而超过植物承载阈值的高氮负荷也会对植物的光合生理活性产生负效应(Yamaguchi et al., 2010), 进而加重植物在复合环境下的碳损失、水分蒸散损失和自由基积累, 对植物产生协同伤害(Karnosky et al., 2005; 列淦文和薛立, 2014).高峰(2018)认为O3与其他环境胁迫因子出现的先后次序也会影响植物对复合作用的响应. ...
Diversity of an early successional plant community as in?uenced by ozone
1
1998
... 植物长期过量摄取O3还可以引起物种组成、冠层结构改变, 影响生态系统种群均匀度和丰富度, 威胁生态系统多样性(Fuhrer et al., 2016; Bergmann et al., 2017), 最终影响生态系统碳、氮和水循环(McLaughlin et al., 2007; Sun et al., 2012; Grulke & Heath, 2020).基于温室或OTCs控制实验下有关草本植物的多项研究都发现, 随着O3暴露时间的延长, 对O3相对敏感的物种/种群将会被对O3相对耐受的物种/种群替代(Fuhrer et al., 1994; Hayes et al., 2009), 非禾本科杂草与禾本科杂草的比例也在发生改变(Hayes et al., 2010).有关Aspen FACE森林生态系统的研究也表明, 尽管整个森林林分结构没有受到明显影响, 但敏感型欧洲山杨树种在O3熏蒸7年后几乎从森林冠层中消失(Kubiske et al., 2007).然而, 由于种内及种间复杂的竞争关系, O3对生态系统群落结构的影响有时候并不是统一的, 高浓度O3暴露环境下O3敏感型种群首先达到较高覆盖度的情况也时有发生(Evans & Ashmore, 1992; Barbo et al., 1998).此外, 由于生态系统各营养级之间的交互作用, 高浓度O3影响植物群落多样性的同时也潜在影响植物组织、化学组成、二次代谢产物(如酚苷类化合物、植物源挥发性有机化合物等)释放, 进而改变植物种群周边与之相关的动植物、真菌、细菌和昆虫行为或活动(ICP Vegetation, 2013; Fuhrer et al., 2016).例如, O3暴露使得西黄松(Pinus ponderosa)更易受到小蠹虫的袭击(Dahlsten et al., 1997), 但O3诱导红云杉(Picea rubens)萜烯物质释放增加的同时减少了其上蚜虫幼虫的数量(Cannon, 1990); 而有关欧洲山杨的研究则发现O3暴露降低柳皮苷和特里杨苷含量, 增加了缩合单宁含量, 引起林分中蚜虫和森林天幕毛虫幼虫数量剧增, 但蚜虫捕食者或天敌(如瓢虫、草蛉和蜘蛛)的数量显著减少(Percy et al., 2002; Kopper & Lindroth, 2003). ...
Impact of tropospheric ozone on terrestrial biodiversity: a literature analysis to identify ozone sensitive taxa
1
2017
... 植物长期过量摄取O3还可以引起物种组成、冠层结构改变, 影响生态系统种群均匀度和丰富度, 威胁生态系统多样性(Fuhrer et al., 2016; Bergmann et al., 2017), 最终影响生态系统碳、氮和水循环(McLaughlin et al., 2007; Sun et al., 2012; Grulke & Heath, 2020).基于温室或OTCs控制实验下有关草本植物的多项研究都发现, 随着O3暴露时间的延长, 对O3相对敏感的物种/种群将会被对O3相对耐受的物种/种群替代(Fuhrer et al., 1994; Hayes et al., 2009), 非禾本科杂草与禾本科杂草的比例也在发生改变(Hayes et al., 2010).有关Aspen FACE森林生态系统的研究也表明, 尽管整个森林林分结构没有受到明显影响, 但敏感型欧洲山杨树种在O3熏蒸7年后几乎从森林冠层中消失(Kubiske et al., 2007).然而, 由于种内及种间复杂的竞争关系, O3对生态系统群落结构的影响有时候并不是统一的, 高浓度O3暴露环境下O3敏感型种群首先达到较高覆盖度的情况也时有发生(Evans & Ashmore, 1992; Barbo et al., 1998).此外, 由于生态系统各营养级之间的交互作用, 高浓度O3影响植物群落多样性的同时也潜在影响植物组织、化学组成、二次代谢产物(如酚苷类化合物、植物源挥发性有机化合物等)释放, 进而改变植物种群周边与之相关的动植物、真菌、细菌和昆虫行为或活动(ICP Vegetation, 2013; Fuhrer et al., 2016).例如, O3暴露使得西黄松(Pinus ponderosa)更易受到小蠹虫的袭击(Dahlsten et al., 1997), 但O3诱导红云杉(Picea rubens)萜烯物质释放增加的同时减少了其上蚜虫幼虫的数量(Cannon, 1990); 而有关欧洲山杨的研究则发现O3暴露降低柳皮苷和特里杨苷含量, 增加了缩合单宁含量, 引起林分中蚜虫和森林天幕毛虫幼虫数量剧增, 但蚜虫捕食者或天敌(如瓢虫、草蛉和蜘蛛)的数量显著减少(Percy et al., 2002; Kopper & Lindroth, 2003). ...
The ozone component of global change: potential effects on agricultural and horticultural plant yield, product quality and interactions with invasive species
3
2009
... O3不仅抑制植物生长, 还导致农作物大量减产(Fuhrer, 2009; Ainsworth et al., 2012; Feng et al., 2015a; Singh & Agrawal, 2017).有关O3对作物产量的影响已经在北美、欧洲和亚洲多地的几十种作物上有所研究报道, 包括对O3敏感的小麦、西瓜(Citrullus lanatus)、豆类、棉花(Gossypium hirsutum)、萝卜(Raphanus sativus)、番茄(Lycopersicon esclentum)、洋葱(Allium cepa)、大豆(Glycine max)和莴苣(Lactuca sativa), 对O3适度敏感的甜菜(Beta vulgaris)、马铃薯(Solanum tuberosum)、油菜(Brassica napus)、烟草(Nicotiana tabacum)、水稻、玉米(Zea mays)、葡萄和西兰花(Brassica oleracea), 和对O3耐受的作物大麦(Hordeum vulgare)、李(Prunus salicina)和草莓(Fragaria ananassa)等(Mills et al., 2007; Booker et al., 2009; Feng & Kobayashi, 2009; Avnery et al., 2011; Broberg et al., 2015).通过汇总406个控制实验的数据进行meta分析发现, 与活性炭过滤环境(≤26 μg·kg-1)相比, 当前环境O3浓度(平均值40 μg·kg-1)可导致主要粮食作物(小麦、水稻、大豆和马铃薯等)减产10%左右; 未来地表O3浓度升高情景下(51-71 μg·kg-1), 作物产量将进一步降低10%- 20% (Feng & Kobayashi, 2009).最近, 有关美国、法国、印度、中国和俄罗斯等小麦主产国的模型评估数据显示, 2010-2012年间高浓度O3浓度造成北半球和南半球小麦平均减产9.9%和6.2%, 抵消了因水肥管理或改良品种等措施对其增产所做的努力(Mills et al., 2018).我国这方面的研究起步较晚, 但发展迅速.从20世纪90年代末开始, 学者先后利用OTC和O3-FACE系统研究了地表O3浓度升高对我国主要粮食作物和蔬菜的影响, 涉及品种包括冬小麦、水稻、大豆、玉米、油菜、菠菜(Spinacia oleracea)和青菜(Brassica rapa)等(冯兆忠等, 2018).研究显示, 高浓度O3暴露下农作物均出现不同程度减产, 不同浓度O3对水稻、小麦、大豆等不同品种作物的减产幅度为10%-60% (贾一磊, 2016; 曹嘉晨等, 2017), 对油菜、菠菜和青菜等蔬菜生物产量的降幅为4.0%-89% (白月明等, 2002; 郭建平等, 2003). ...
... (4)关注O3对作物品质及食品安全的危害: 世界上主要粮食作物要么对O3敏感(小麦、大豆等), 要么对O3适度敏感(玉米、水稻和马铃薯等).到2050年, 世界人口预计增加到90亿, 粮食供应需增加至少50%才能满足人口增长的需求, 如何保证粮食供应安全将成为21世纪最重要的一个挑战(Tai et al., 2014).目前为止, 除作物产量之外, 有关地表O3浓度升高对作物品质的影响研究多集中在蔬菜作物或粮食作物可食部位糖分、蛋白质和淀粉含量等食用口感方面(Booker et al., 2009), 很少涉及其背后对人体健康的直接影响.随着生活水平的提高, 人们更加关注食品的营养与安全(如微量元素含量或健康风险等), 而长期高浓度O3暴露可改变粮食作物籽粒微量元素及营养物质含量(Booker et al., 2009; 冯兆忠等, 2018).此外, 也有研究显示O3暴露导致马铃薯的糖苷生物碱含量增加, 糖苷生物碱不仅导致味苦而且浓度较高时具有毒性(Pell & Pearson, 1984).因此, 在全球环境变化大背景下, 长期食用暴露在O3污染下的粮食作物是否影响人体营养平衡或危及健康值得关注.未来的研究应关注高浓度O3对农作物外观品质(如籽粒形态、发芽率和成活率等)、加工特性、口感(如黏弹性、柔软性等)、烹饪特性和营养特性等外在和内在价值的综合影响. ...
... 暴露可改变粮食作物籽粒微量元素及营养物质含量(Booker et al., 2009; 冯兆忠等, 2018).此外, 也有研究显示O3暴露导致马铃薯的糖苷生物碱含量增加, 糖苷生物碱不仅导致味苦而且浓度较高时具有毒性(Pell & Pearson, 1984).因此, 在全球环境变化大背景下, 长期食用暴露在O3污染下的粮食作物是否影响人体营养平衡或危及健康值得关注.未来的研究应关注高浓度O3对农作物外观品质(如籽粒形态、发芽率和成活率等)、加工特性、口感(如黏弹性、柔软性等)、烹饪特性和营养特性等外在和内在价值的综合影响. ...
Ozone effects on wheat grain quality—A summary
2
2015
... O3不仅抑制植物生长, 还导致农作物大量减产(Fuhrer, 2009; Ainsworth et al., 2012; Feng et al., 2015a; Singh & Agrawal, 2017).有关O3对作物产量的影响已经在北美、欧洲和亚洲多地的几十种作物上有所研究报道, 包括对O3敏感的小麦、西瓜(Citrullus lanatus)、豆类、棉花(Gossypium hirsutum)、萝卜(Raphanus sativus)、番茄(Lycopersicon esclentum)、洋葱(Allium cepa)、大豆(Glycine max)和莴苣(Lactuca sativa), 对O3适度敏感的甜菜(Beta vulgaris)、马铃薯(Solanum tuberosum)、油菜(Brassica napus)、烟草(Nicotiana tabacum)、水稻、玉米(Zea mays)、葡萄和西兰花(Brassica oleracea), 和对O3耐受的作物大麦(Hordeum vulgare)、李(Prunus salicina)和草莓(Fragaria ananassa)等(Mills et al., 2007; Booker et al., 2009; Feng & Kobayashi, 2009; Avnery et al., 2011; Broberg et al., 2015).通过汇总406个控制实验的数据进行meta分析发现, 与活性炭过滤环境(≤26 μg·kg-1)相比, 当前环境O3浓度(平均值40 μg·kg-1)可导致主要粮食作物(小麦、水稻、大豆和马铃薯等)减产10%左右; 未来地表O3浓度升高情景下(51-71 μg·kg-1), 作物产量将进一步降低10%- 20% (Feng & Kobayashi, 2009).最近, 有关美国、法国、印度、中国和俄罗斯等小麦主产国的模型评估数据显示, 2010-2012年间高浓度O3浓度造成北半球和南半球小麦平均减产9.9%和6.2%, 抵消了因水肥管理或改良品种等措施对其增产所做的努力(Mills et al., 2018).我国这方面的研究起步较晚, 但发展迅速.从20世纪90年代末开始, 学者先后利用OTC和O3-FACE系统研究了地表O3浓度升高对我国主要粮食作物和蔬菜的影响, 涉及品种包括冬小麦、水稻、大豆、玉米、油菜、菠菜(Spinacia oleracea)和青菜(Brassica rapa)等(冯兆忠等, 2018).研究显示, 高浓度O3暴露下农作物均出现不同程度减产, 不同浓度O3对水稻、小麦、大豆等不同品种作物的减产幅度为10%-60% (贾一磊, 2016; 曹嘉晨等, 2017), 对油菜、菠菜和青菜等蔬菜生物产量的降幅为4.0%-89% (白月明等, 2002; 郭建平等, 2003). ...
... 相对于农作物产量, O3对粮食品质影响的研究集中在几种主要农作物(如小麦、水稻和大豆)(冯兆忠等, 2018).基于北美、欧洲和亚洲已有的O3对小麦影响的meta分析发现, O3显著降低小麦淀粉浓度, 但显著增加蛋白质和一些营养元素浓度(如钾、镁、钙、磷、锌、锰和铜)(Broberg et al., 2015).印度北部热带地区有关小麦的研究则显示, O3显著降低小麦籽粒糖含量, 但没有改变籽粒蛋白质含量(Tomer et al., 2015).有关水稻的研究还发现, O3增加粳稻直链淀粉含量, 但对籼稻直链淀粉含量无显著影响(Sawada et al., 2016).国内研究则表明, 尽管高浓度O3显著降低冬小麦面粉中支链淀粉和淀粉含量, 但是增加直链淀粉含量(Zhang et al., 2013).O3显著降低水稻稻米直链淀粉浓度、分解率, 增加黏度、起糊温度、蛋白质和微量矿物质浓度, 但蛋白质总量没有改变(Wang et al., 2012).此外, 研究还发现O3熏蒸显著降低了小麦籽粒出粉率和硬度, 但增加冬小麦面粉峰值黏度、低谷黏度、最终黏度和反弹值(贾一磊, 2016). ...
New flux based dose-response relationships for ozone for European forest tree species
1
2015
... 然而, O3对植物的影响不仅依赖于大气O3浓度同时也依赖于叶片气孔导度(O3的吸收)和植物自身抗氧化能力(Matyssek et al., 2007).近年来, 基于气孔O3吸收量的评估指标PODY (植物毒性O3剂量, 整个生长季单位面积上气孔O3吸收通量超过临界值Y nmol m-2·s-1的积累量)被开发并逐步替代浓度暴露剂量指标, 用于区域或全球O3对农作物和森林树种生长影响的评估(Büker et al., 2015; LRTAR Convention, 2015).目前, 哪个指标更适合区域O3风险评估的争论一直存在, 但欧洲和亚洲地区多项O3对农作物或植被影响的研究均表明: 考虑水分供应的情况下, O3暴露造成的生物量或作物产量的降低与PODY的拟合度比AOTX更好(Mills et al., 2011; Gao et al., 2017), 能更好表征O3对生态系统的影响.整体来看, 各个评估指标的计算方式和所需参数从简单到复杂, 从单一考虑O3浓度到综合考虑环境因子对植物O3吸收通量的影响, 逐渐可以精确评估地表O3污染对生态系统农林生产力的影响(表3). ...
Olfactory response of eastern spruce budworm larvae to red spruce needles exposed to acid-rain and elevated levels of ozone
1
1990
... 植物长期过量摄取O3还可以引起物种组成、冠层结构改变, 影响生态系统种群均匀度和丰富度, 威胁生态系统多样性(Fuhrer et al., 2016; Bergmann et al., 2017), 最终影响生态系统碳、氮和水循环(McLaughlin et al., 2007; Sun et al., 2012; Grulke & Heath, 2020).基于温室或OTCs控制实验下有关草本植物的多项研究都发现, 随着O3暴露时间的延长, 对O3相对敏感的物种/种群将会被对O3相对耐受的物种/种群替代(Fuhrer et al., 1994; Hayes et al., 2009), 非禾本科杂草与禾本科杂草的比例也在发生改变(Hayes et al., 2010).有关Aspen FACE森林生态系统的研究也表明, 尽管整个森林林分结构没有受到明显影响, 但敏感型欧洲山杨树种在O3熏蒸7年后几乎从森林冠层中消失(Kubiske et al., 2007).然而, 由于种内及种间复杂的竞争关系, O3对生态系统群落结构的影响有时候并不是统一的, 高浓度O3暴露环境下O3敏感型种群首先达到较高覆盖度的情况也时有发生(Evans & Ashmore, 1992; Barbo et al., 1998).此外, 由于生态系统各营养级之间的交互作用, 高浓度O3影响植物群落多样性的同时也潜在影响植物组织、化学组成、二次代谢产物(如酚苷类化合物、植物源挥发性有机化合物等)释放, 进而改变植物种群周边与之相关的动植物、真菌、细菌和昆虫行为或活动(ICP Vegetation, 2013; Fuhrer et al., 2016).例如, O3暴露使得西黄松(Pinus ponderosa)更易受到小蠹虫的袭击(Dahlsten et al., 1997), 但O3诱导红云杉(Picea rubens)萜烯物质释放增加的同时减少了其上蚜虫幼虫的数量(Cannon, 1990); 而有关欧洲山杨的研究则发现O3暴露降低柳皮苷和特里杨苷含量, 增加了缩合单宁含量, 引起林分中蚜虫和森林天幕毛虫幼虫数量剧增, 但蚜虫捕食者或天敌(如瓢虫、草蛉和蜘蛛)的数量显著减少(Percy et al., 2002; Kopper & Lindroth, 2003). ...
地表臭氧浓度升高对冬小麦和大豆生长和产量的影响
1
2017
... O3不仅抑制植物生长, 还导致农作物大量减产(Fuhrer, 2009; Ainsworth et al., 2012; Feng et al., 2015a; Singh & Agrawal, 2017).有关O3对作物产量的影响已经在北美、欧洲和亚洲多地的几十种作物上有所研究报道, 包括对O3敏感的小麦、西瓜(Citrullus lanatus)、豆类、棉花(Gossypium hirsutum)、萝卜(Raphanus sativus)、番茄(Lycopersicon esclentum)、洋葱(Allium cepa)、大豆(Glycine max)和莴苣(Lactuca sativa), 对O3适度敏感的甜菜(Beta vulgaris)、马铃薯(Solanum tuberosum)、油菜(Brassica napus)、烟草(Nicotiana tabacum)、水稻、玉米(Zea mays)、葡萄和西兰花(Brassica oleracea), 和对O3耐受的作物大麦(Hordeum vulgare)、李(Prunus salicina)和草莓(Fragaria ananassa)等(Mills et al., 2007; Booker et al., 2009; Feng & Kobayashi, 2009; Avnery et al., 2011; Broberg et al., 2015).通过汇总406个控制实验的数据进行meta分析发现, 与活性炭过滤环境(≤26 μg·kg-1)相比, 当前环境O3浓度(平均值40 μg·kg-1)可导致主要粮食作物(小麦、水稻、大豆和马铃薯等)减产10%左右; 未来地表O3浓度升高情景下(51-71 μg·kg-1), 作物产量将进一步降低10%- 20% (Feng & Kobayashi, 2009).最近, 有关美国、法国、印度、中国和俄罗斯等小麦主产国的模型评估数据显示, 2010-2012年间高浓度O3浓度造成北半球和南半球小麦平均减产9.9%和6.2%, 抵消了因水肥管理或改良品种等措施对其增产所做的努力(Mills et al., 2018).我国这方面的研究起步较晚, 但发展迅速.从20世纪90年代末开始, 学者先后利用OTC和O3-FACE系统研究了地表O3浓度升高对我国主要粮食作物和蔬菜的影响, 涉及品种包括冬小麦、水稻、大豆、玉米、油菜、菠菜(Spinacia oleracea)和青菜(Brassica rapa)等(冯兆忠等, 2018).研究显示, 高浓度O3暴露下农作物均出现不同程度减产, 不同浓度O3对水稻、小麦、大豆等不同品种作物的减产幅度为10%-60% (贾一磊, 2016; 曹嘉晨等, 2017), 对油菜、菠菜和青菜等蔬菜生物产量的降幅为4.0%-89% (白月明等, 2002; 郭建平等, 2003). ...
地表臭氧浓度升高对冬小麦和大豆生长和产量的影响
1
2017
... O3不仅抑制植物生长, 还导致农作物大量减产(Fuhrer, 2009; Ainsworth et al., 2012; Feng et al., 2015a; Singh & Agrawal, 2017).有关O3对作物产量的影响已经在北美、欧洲和亚洲多地的几十种作物上有所研究报道, 包括对O3敏感的小麦、西瓜(Citrullus lanatus)、豆类、棉花(Gossypium hirsutum)、萝卜(Raphanus sativus)、番茄(Lycopersicon esclentum)、洋葱(Allium cepa)、大豆(Glycine max)和莴苣(Lactuca sativa), 对O3适度敏感的甜菜(Beta vulgaris)、马铃薯(Solanum tuberosum)、油菜(Brassica napus)、烟草(Nicotiana tabacum)、水稻、玉米(Zea mays)、葡萄和西兰花(Brassica oleracea), 和对O3耐受的作物大麦(Hordeum vulgare)、李(Prunus salicina)和草莓(Fragaria ananassa)等(Mills et al., 2007; Booker et al., 2009; Feng & Kobayashi, 2009; Avnery et al., 2011; Broberg et al., 2015).通过汇总406个控制实验的数据进行meta分析发现, 与活性炭过滤环境(≤26 μg·kg-1)相比, 当前环境O3浓度(平均值40 μg·kg-1)可导致主要粮食作物(小麦、水稻、大豆和马铃薯等)减产10%左右; 未来地表O3浓度升高情景下(51-71 μg·kg-1), 作物产量将进一步降低10%- 20% (Feng & Kobayashi, 2009).最近, 有关美国、法国、印度、中国和俄罗斯等小麦主产国的模型评估数据显示, 2010-2012年间高浓度O3浓度造成北半球和南半球小麦平均减产9.9%和6.2%, 抵消了因水肥管理或改良品种等措施对其增产所做的努力(Mills et al., 2018).我国这方面的研究起步较晚, 但发展迅速.从20世纪90年代末开始, 学者先后利用OTC和O3-FACE系统研究了地表O3浓度升高对我国主要粮食作物和蔬菜的影响, 涉及品种包括冬小麦、水稻、大豆、玉米、油菜、菠菜(Spinacia oleracea)和青菜(Brassica rapa)等(冯兆忠等, 2018).研究显示, 高浓度O3暴露下农作物均出现不同程度减产, 不同浓度O3对水稻、小麦、大豆等不同品种作物的减产幅度为10%-60% (贾一磊, 2016; 曹嘉晨等, 2017), 对油菜、菠菜和青菜等蔬菜生物产量的降幅为4.0%-89% (白月明等, 2002; 郭建平等, 2003). ...
Growth of continental-scale metro-agro-plexes, regional ozone pollution and world food production
1
1994
... 基于OTCs和O3-FACE平台下单个站点的控制实验可探究O3对植物个体、群落或生态系统的影响, 但将站点或样地尺度的研究外推到区域或全球, 评估O3污染区域风险的研究主要通过响应关系和模型.目前, 其研究领域主要集中在地表O3污染对农作物产量和森林生产力等方面.早在1994年, Chameides等(1994)就利用经济模型初步预测O3浓度升高可能导致2025年全球30%-75%的作物面临产量降低的风险.基于O3前体物排放、化学反应传输模型和M12、AOT40指标的评估显示, 2000年全球O3污染引起全球小麦、玉米和大豆产量平均降低2.2%-15%, 每年因上述作物产量降低所导致的经济损失达110-180亿美元(Avnery et al., 2011).最近, 基于PODY评估指标的研究则表明, O3对小麦造成的产量损失北半球(9.9%)明显高于南半球(6.2%), 平均产量损失最高的地区主要位于北半球暖温带湿润地区(12.5%)、热带湿热(12.5%)和热带潮湿气候区(17.2%)(Mills et al., 2018).在区域尺度上, 基于1990年O3浓度数据, Wang和Mauzerall (2004)利用M7和M12指标评估了O3对中国、日本和韩国主要农作物产量的影响, 结果显示O3污染造成小麦、玉米、棉花减产1%-9%, 大豆减产23%-27%, 造成2.4-35亿美元的经济损失, 在维持现有的污染物排放、管理措施和农业生产技术不变的情况下, 预计到2020年小麦、玉米、棉花减产幅度为2%-15%, 大豆为28%-35%.最近, 利用AOT40指标的研究显示, 2014年当前O3浓度导致中国农作物产量损失7 840万t, 小麦、水稻和玉米相对减产量分别为8.5%-14%、3.9%-15%和2.2%-5.5% (Lin et al., 2018).而只聚焦于长江三角洲地区的研究则发现, 2015年区域O3污染分别造成小麦和水稻减产9.9%-36.1%和7.3%- 23.9% (Zhao et al., 2018).虽然国内也有少量研究建立了PODY与个别树种或作物相对生物量或相对产量之间的通量响应关系(Feng et al., 2012; Hu et al., 2015; Gao et al., 2017), 但并没有研究利用PODY指标评估区域O3污染对农作物产量损失的影响. ...
Ambient ozone effects on forest trees of the eastern United States: a review
1
1998
... 区域尺度上, 地表O3污染对陆地生态系统固碳能力的影响主要依靠模式算法的估算.早在20世纪90年代, 基于AOT40指标的研究就发现高浓度O3降低美国东北部阔叶林净初级生产力(NPP) 3%-16%, 对地上生物量的降低每年为0-10% (Chappelka & Samuelson, 1998).另有研究表明O3污染造成美国植被碳固定降低18-38 Tg·a-1, NPP降低至少9 g·m-2·a-1, 且高浓度O3导致植物-土壤系统碳积累和固定能力下降(Felzer et al., 2004).此外, 运用自主开发的基于过程的陆地生态系统动态模型(DLEM), 国内学者发现近40年来O3浓度升高使中国森林净固碳量减少7.7%, 且不同的森林类型降低幅度不同: 如北方落叶阔叶林降低21.8%, 北方落叶针叶林降低5.2%, 温带落叶阔叶林降低21.1%, 温带常绿阔叶林降低7.6%, 温带常绿针叶林降低0.4%, 温带落叶针叶林降低6.2%, 热带落叶阔叶林降低43.1%, 热带常绿阔叶林降低9.6% (Ren et al., 2011).最近, Yue等(2017)用地球系统模式(NASA ModelE2-YIBs)得出O3污染造成中国陆地植被NPP降低600 Tg·a-1, 且对于不同功能类型的植物, O3对总初级生产力(GPP)的降低都随着O3浓度的升高而加大.总体来看, 模型驱动的结果与大量控制实验的发现相吻合, 均表明O3污染在不同尺度都对自然生态系统碳固定有显著的负效应, 未来O3浓度升高有可能抵消因大气CO2浓度升高所增加的GPP (18%-34%)(Sitch et al., 2007). ...
How vegetation impacts affect climate metrics for ozone precursors
1
2010
... 地表O3浓度升高对植物光合作用、生长和生物量积累、物质分配等过程的连锁负效应导致陆地生态系统生产力和固碳能力降低(Reich, 1987; Wittig et al., 2009; Ainsworth et al., 2012), 削弱了自然生态系统的碳汇能力(Felzer et al., 2005; Sitch et al., 2007; Collins et al., 2010).基于大量OTCs控制实验下北半球中纬度北方温带树种的meta分析发现, O3浓度升高导致树木的净光合作用和总生物量分别降低11%和7% (Wittig et al., 2009).最近, 有关中国热带、亚热带和温带树种的meta分析则发现, O3浓度升高导致中国木本植物的净光合作用和总生物量分别降低28%和14% (Li et al., 2017). ...
Global distribution and trends of tropospheric ozone: an observation- based review
1
2014
... 地表臭氧(O3)特指距离地球表面15 km范围内的近地层O3, 除少量来自平流层大气传输外, 其余大部分是氮氧化物(NOx)、非甲烷类挥发性有机化合物(NMVOCs)、一氧化碳(CO)和甲烷(CH4)等前体物在强烈光照下发生光化学反应而产生(冯兆忠等, 2018).自工业革命以来, 伴随城市化进程加快和化石燃料的过度燃烧, 地表O3浓度在世界范围内普遍升高.近年来, 模型和观测数据都表明地表O3浓度小时峰值在欧洲和北美等地已有所下降, 但亚洲地区仍呈不断上升的趋势(Cooper et al., 2014).中国国家环境监测网络2013-2017年的数据表明, 过去5年中国地表O3浓度以每年3 μg·kg-1的速率攀升(Wang et al., 2019), 目前全国地表O3浓度每日8 h滑动平均最大值(MDA8)的年平均值已达(41.2 ± 6.3) μg·kg-1, 尤其华北平原、长江三角洲和珠江三角洲等地(MDA8峰值年平均值已处于60-70 μg·kg-1), 地表O3污染形势十分严峻(Lu et al., 2018).地表O3是具有植物毒性的气体污染物, 具有强氧化性.高浓度地表O3或长期低浓度O3能诱导植物叶片出现可见损伤, 加速植物衰老, 降低光合作用, 改变碳分配, 抑制植物生长和生物量积累, 影响作物品质和产量, 最终降低生态系统碳汇能力, 改变生态系统结构和功能(Ashmore, 2005; Ainsworth, 2017; 高峰, 2018). ...
Effects of oxidant air pollutants on western pine beetle (Coleoptera: Scolytidae) populations in southern California
1
1997
... 植物长期过量摄取O3还可以引起物种组成、冠层结构改变, 影响生态系统种群均匀度和丰富度, 威胁生态系统多样性(Fuhrer et al., 2016; Bergmann et al., 2017), 最终影响生态系统碳、氮和水循环(McLaughlin et al., 2007; Sun et al., 2012; Grulke & Heath, 2020).基于温室或OTCs控制实验下有关草本植物的多项研究都发现, 随着O3暴露时间的延长, 对O3相对敏感的物种/种群将会被对O3相对耐受的物种/种群替代(Fuhrer et al., 1994; Hayes et al., 2009), 非禾本科杂草与禾本科杂草的比例也在发生改变(Hayes et al., 2010).有关Aspen FACE森林生态系统的研究也表明, 尽管整个森林林分结构没有受到明显影响, 但敏感型欧洲山杨树种在O3熏蒸7年后几乎从森林冠层中消失(Kubiske et al., 2007).然而, 由于种内及种间复杂的竞争关系, O3对生态系统群落结构的影响有时候并不是统一的, 高浓度O3暴露环境下O3敏感型种群首先达到较高覆盖度的情况也时有发生(Evans & Ashmore, 1992; Barbo et al., 1998).此外, 由于生态系统各营养级之间的交互作用, 高浓度O3影响植物群落多样性的同时也潜在影响植物组织、化学组成、二次代谢产物(如酚苷类化合物、植物源挥发性有机化合物等)释放, 进而改变植物种群周边与之相关的动植物、真菌、细菌和昆虫行为或活动(ICP Vegetation, 2013; Fuhrer et al., 2016).例如, O3暴露使得西黄松(Pinus ponderosa)更易受到小蠹虫的袭击(Dahlsten et al., 1997), 但O3诱导红云杉(Picea rubens)萜烯物质释放增加的同时减少了其上蚜虫幼虫的数量(Cannon, 1990); 而有关欧洲山杨的研究则发现O3暴露降低柳皮苷和特里杨苷含量, 增加了缩合单宁含量, 引起林分中蚜虫和森林天幕毛虫幼虫数量剧增, 但蚜虫捕食者或天敌(如瓢虫、草蛉和蜘蛛)的数量显著减少(Percy et al., 2002; Kopper & Lindroth, 2003). ...
Increase of apoplastic ascorbate induced by ozone is insufficient to remove the negative effects in tobacco, soybean and poplar
1
2019
... 气孔防御机制失效后, 进入细胞的O3将诱导叶片内部抗氧化系统启动解毒和修复的非气孔防御响应(冯兆忠等, 2018).非气孔防御响应主要是指“质外体-共质体系统”内一些抗氧化组分对O3解毒和修复的防御过程.具体来说, O3进入植物叶片的细胞间隙后会溶解在细胞壁的结合水里, 经过一系列的化学反应形成活性氧分子(ROS), ROS能够破坏细胞壁并与细胞膜上的不饱和脂肪酸发生反应, 从而对叶片造成损伤(Tausz et al., 2007; Pellegrini et al., 2018).植物质外体的抗氧化物质如还原性抗坏血酸(ASC)、抗氧化酶(如超氧化物歧化酶)可与ROS反应进行初级解毒, 成为抵御O3的第二道防线(Kangasjärvi et al., 2005; Dai et al., 2019), 而剩余的ROS会经过质膜到达共质体, 此时共质体内抗氧化成分如过氧化物酶(POD)、过氧化氢酶(CAT)、抗坏血酸过氧化物酶(APX)及抗氧化剂(如酚类化合物-谷胱甘肽(GSH)、脯氨酸和多胺等)等抗氧化酶类物质被诱导作为次级解毒响应机制参与诸如“抗坏血酸-谷胱甘肽(ASA-GSH)循环”等氧化还原过程, 进而对ROS进行进一步解毒与消除(Luwe, 1996).气孔闭合除了会减少O3的吸收, 也会抑制CO2吸收和水汽交换, 而非气孔防御响应中的抗氧化系统解毒过程也会消耗大量的能量并对光合反应系统造成不同程度的损伤, 降低羧化速率和电子传递速率, 进而抑制光合和蒸腾作用, 损害植物生长(Gao et al., 2016).然而, 最新研究表明持续整个生长季的长期O3暴露会影响植物叶片的气孔密度、开度和大小等表观属性(李品等, 2018), 而O3从气孔进入到细胞间隙后, 也可以通过影响植物的叶肉导度降低植物的光合能力(Xu et al., 2019).此外, 有关29种落叶和常绿树种应对O3胁迫的研究还发现不同木本植物对O3响应的差异主要来自植物功能叶片比叶质量的大小(即叶片比叶质量越大的树种对O3越不敏感), 与气孔导度无明显相关性(Li et al., 2016). ...
Enhanced tolerance to ozone and drought stresses in transgenic tobacco overexpressing dehydroascorbate reductase in cytosol
1
2006
... 除单因子O3胁迫外, 由于全球气候变化和极端气候常态化, 植物经常受到O3和其他环境因子交互作用的影响(如O3与CO2、升温、干旱或氮沉降等)(Schmadel-Hagebölling et al., 1998; Ollinger et al., 2002; Eltayeb et al., 2006; Ghimire et al., 2018).然而, 植物应对O3与其他因子交互作用的响应并不是两者或单独作用的简单叠加, 往往取决于植物种类、生长时期、对特定环境因子的敏感性、胁迫浓度及时间, 以及具体的微气候条件(列淦文和薛立, 2014).一方面, 通过诱导气孔关闭、限制叶片O3吸收(CO2浓度升高/干旱胁迫), 或提高碳同化速率、促进植物光合作用(CO2浓度升高/氮增加), CO2浓度升高、干旱胁迫和氮沉降经常被报道可增强植物抵御O3的能力, 减缓O3伤害(白月明等, 2005; Handley & Grulke, 2008; Gao et al., 2017).另一方面, 与之相反, 也有研究发现长期高浓度CO2或干旱降低气孔导度的同时, 会引起植物蒸腾速率、光合产物合成和积累的降低(McDonald et al., 2002; Pollastrini et al., 2010); 而超过植物承载阈值的高氮负荷也会对植物的光合生理活性产生负效应(Yamaguchi et al., 2010), 进而加重植物在复合环境下的碳损失、水分蒸散损失和自由基积累, 对植物产生协同伤害(Karnosky et al., 2005; 列淦文和薛立, 2014).高峰(2018)认为O3与其他环境胁迫因子出现的先后次序也会影响植物对复合作用的响应. ...
The effects of ambient air on a seminatural grassland community
1
1992
... 植物长期过量摄取O3还可以引起物种组成、冠层结构改变, 影响生态系统种群均匀度和丰富度, 威胁生态系统多样性(Fuhrer et al., 2016; Bergmann et al., 2017), 最终影响生态系统碳、氮和水循环(McLaughlin et al., 2007; Sun et al., 2012; Grulke & Heath, 2020).基于温室或OTCs控制实验下有关草本植物的多项研究都发现, 随着O3暴露时间的延长, 对O3相对敏感的物种/种群将会被对O3相对耐受的物种/种群替代(Fuhrer et al., 1994; Hayes et al., 2009), 非禾本科杂草与禾本科杂草的比例也在发生改变(Hayes et al., 2010).有关Aspen FACE森林生态系统的研究也表明, 尽管整个森林林分结构没有受到明显影响, 但敏感型欧洲山杨树种在O3熏蒸7年后几乎从森林冠层中消失(Kubiske et al., 2007).然而, 由于种内及种间复杂的竞争关系, O3对生态系统群落结构的影响有时候并不是统一的, 高浓度O3暴露环境下O3敏感型种群首先达到较高覆盖度的情况也时有发生(Evans & Ashmore, 1992; Barbo et al., 1998).此外, 由于生态系统各营养级之间的交互作用, 高浓度O3影响植物群落多样性的同时也潜在影响植物组织、化学组成、二次代谢产物(如酚苷类化合物、植物源挥发性有机化合物等)释放, 进而改变植物种群周边与之相关的动植物、真菌、细菌和昆虫行为或活动(ICP Vegetation, 2013; Fuhrer et al., 2016).例如, O3暴露使得西黄松(Pinus ponderosa)更易受到小蠹虫的袭击(Dahlsten et al., 1997), 但O3诱导红云杉(Picea rubens)萜烯物质释放增加的同时减少了其上蚜虫幼虫的数量(Cannon, 1990); 而有关欧洲山杨的研究则发现O3暴露降低柳皮苷和特里杨苷含量, 增加了缩合单宁含量, 引起林分中蚜虫和森林天幕毛虫幼虫数量剧增, 但蚜虫捕食者或天敌(如瓢虫、草蛉和蜘蛛)的数量显著减少(Percy et al., 2002; Kopper & Lindroth, 2003). ...
Effects of ozone on net primary production and carbon sequestration in the conterminous United States using a biogeochemistry model
1
2004
... 区域尺度上, 地表O3污染对陆地生态系统固碳能力的影响主要依靠模式算法的估算.早在20世纪90年代, 基于AOT40指标的研究就发现高浓度O3降低美国东北部阔叶林净初级生产力(NPP) 3%-16%, 对地上生物量的降低每年为0-10% (Chappelka & Samuelson, 1998).另有研究表明O3污染造成美国植被碳固定降低18-38 Tg·a-1, NPP降低至少9 g·m-2·a-1, 且高浓度O3导致植物-土壤系统碳积累和固定能力下降(Felzer et al., 2004).此外, 运用自主开发的基于过程的陆地生态系统动态模型(DLEM), 国内学者发现近40年来O3浓度升高使中国森林净固碳量减少7.7%, 且不同的森林类型降低幅度不同: 如北方落叶阔叶林降低21.8%, 北方落叶针叶林降低5.2%, 温带落叶阔叶林降低21.1%, 温带常绿阔叶林降低7.6%, 温带常绿针叶林降低0.4%, 温带落叶针叶林降低6.2%, 热带落叶阔叶林降低43.1%, 热带常绿阔叶林降低9.6% (Ren et al., 2011).最近, Yue等(2017)用地球系统模式(NASA ModelE2-YIBs)得出O3污染造成中国陆地植被NPP降低600 Tg·a-1, 且对于不同功能类型的植物, O3对总初级生产力(GPP)的降低都随着O3浓度的升高而加大.总体来看, 模型驱动的结果与大量控制实验的发现相吻合, 均表明O3污染在不同尺度都对自然生态系统碳固定有显著的负效应, 未来O3浓度升高有可能抵消因大气CO2浓度升高所增加的GPP (18%-34%)(Sitch et al., 2007). ...
Future effects of ozone on carbon sequestration and climate change policy using a global biogeochemical model
1
2005
... 地表O3浓度升高对植物光合作用、生长和生物量积累、物质分配等过程的连锁负效应导致陆地生态系统生产力和固碳能力降低(Reich, 1987; Wittig et al., 2009; Ainsworth et al., 2012), 削弱了自然生态系统的碳汇能力(Felzer et al., 2005; Sitch et al., 2007; Collins et al., 2010).基于大量OTCs控制实验下北半球中纬度北方温带树种的meta分析发现, O3浓度升高导致树木的净光合作用和总生物量分别降低11%和7% (Wittig et al., 2009).最近, 有关中国热带、亚热带和温带树种的meta分析则发现, O3浓度升高导致中国木本植物的净光合作用和总生物量分别降低28%和14% (Li et al., 2017). ...
A unifying explanation for variation in ozone sensitivity among woody plants
1
2018
... (1)开展O3-FACE研究: 现有研究多为OTC系统下的实验, OTC气室内“微气候” (如温度、水分、光照和风力等生态因素)与外界环境状态存在明显差异, 这些生态因子的变化可能影响植物应对O3的响应, 缓解或加重O3污染对农作物产量或森林植被生产力的损伤(Long et al., 2005; Feng et al., 2018a).全球模型评估结果显示, OTC实验结果低估了O3对作物产量损失的影响(van Dingenen et al., 2009).最近, Feng等(2018b)也发现, 同样浓度的O3对OTC实验下小麦和水稻产量的影响要低于FACE实验; 而对大豆产量的影响则相反.因此, OTC实验结果并不能完全应用于实际O3区域风险的评估.O3-FACE平台是未来研究实际大田环境下O3浓度升高对农林生态系统影响的主要依托平台.为减少区域O3风险评估的不确定性, 未来应加大O3-FACE平台的实验研究.考虑到目前已开展的FACE实验多坐落在温带地区, 主要探究温带植被或作物对高浓度O3的响应, 较少涉及热带和寒带植物或其他区域典型生态系统(Ainsworth et al., 2020).最理想的情况是, 不同区域、不同生态系统类型共同开展O3-FACE实验研究(Ainsworth et al., 2012). ...
Ground-level O3 pollution and its impacts on food crops in China: a review
1
2015
... O3不仅抑制植物生长, 还导致农作物大量减产(Fuhrer, 2009; Ainsworth et al., 2012; Feng et al., 2015a; Singh & Agrawal, 2017).有关O3对作物产量的影响已经在北美、欧洲和亚洲多地的几十种作物上有所研究报道, 包括对O3敏感的小麦、西瓜(Citrullus lanatus)、豆类、棉花(Gossypium hirsutum)、萝卜(Raphanus sativus)、番茄(Lycopersicon esclentum)、洋葱(Allium cepa)、大豆(Glycine max)和莴苣(Lactuca sativa), 对O3适度敏感的甜菜(Beta vulgaris)、马铃薯(Solanum tuberosum)、油菜(Brassica napus)、烟草(Nicotiana tabacum)、水稻、玉米(Zea mays)、葡萄和西兰花(Brassica oleracea), 和对O3耐受的作物大麦(Hordeum vulgare)、李(Prunus salicina)和草莓(Fragaria ananassa)等(Mills et al., 2007; Booker et al., 2009; Feng & Kobayashi, 2009; Avnery et al., 2011; Broberg et al., 2015).通过汇总406个控制实验的数据进行meta分析发现, 与活性炭过滤环境(≤26 μg·kg-1)相比, 当前环境O3浓度(平均值40 μg·kg-1)可导致主要粮食作物(小麦、水稻、大豆和马铃薯等)减产10%左右; 未来地表O3浓度升高情景下(51-71 μg·kg-1), 作物产量将进一步降低10%- 20% (Feng & Kobayashi, 2009).最近, 有关美国、法国、印度、中国和俄罗斯等小麦主产国的模型评估数据显示, 2010-2012年间高浓度O3浓度造成北半球和南半球小麦平均减产9.9%和6.2%, 抵消了因水肥管理或改良品种等措施对其增产所做的努力(Mills et al., 2018).我国这方面的研究起步较晚, 但发展迅速.从20世纪90年代末开始, 学者先后利用OTC和O3-FACE系统研究了地表O3浓度升高对我国主要粮食作物和蔬菜的影响, 涉及品种包括冬小麦、水稻、大豆、玉米、油菜、菠菜(Spinacia oleracea)和青菜(Brassica rapa)等(冯兆忠等, 2018).研究显示, 高浓度O3暴露下农作物均出现不同程度减产, 不同浓度O3对水稻、小麦、大豆等不同品种作物的减产幅度为10%-60% (贾一磊, 2016; 曹嘉晨等, 2017), 对油菜、菠菜和青菜等蔬菜生物产量的降幅为4.0%-89% (白月明等, 2002; 郭建平等, 2003). ...
Assessing the impacts of current and future concentrations of surface ozone on crop yield with meta-analysis
2
2009
... O3不仅抑制植物生长, 还导致农作物大量减产(Fuhrer, 2009; Ainsworth et al., 2012; Feng et al., 2015a; Singh & Agrawal, 2017).有关O3对作物产量的影响已经在北美、欧洲和亚洲多地的几十种作物上有所研究报道, 包括对O3敏感的小麦、西瓜(Citrullus lanatus)、豆类、棉花(Gossypium hirsutum)、萝卜(Raphanus sativus)、番茄(Lycopersicon esclentum)、洋葱(Allium cepa)、大豆(Glycine max)和莴苣(Lactuca sativa), 对O3适度敏感的甜菜(Beta vulgaris)、马铃薯(Solanum tuberosum)、油菜(Brassica napus)、烟草(Nicotiana tabacum)、水稻、玉米(Zea mays)、葡萄和西兰花(Brassica oleracea), 和对O3耐受的作物大麦(Hordeum vulgare)、李(Prunus salicina)和草莓(Fragaria ananassa)等(Mills et al., 2007; Booker et al., 2009; Feng & Kobayashi, 2009; Avnery et al., 2011; Broberg et al., 2015).通过汇总406个控制实验的数据进行meta分析发现, 与活性炭过滤环境(≤26 μg·kg-1)相比, 当前环境O3浓度(平均值40 μg·kg-1)可导致主要粮食作物(小麦、水稻、大豆和马铃薯等)减产10%左右; 未来地表O3浓度升高情景下(51-71 μg·kg-1), 作物产量将进一步降低10%- 20% (Feng & Kobayashi, 2009).最近, 有关美国、法国、印度、中国和俄罗斯等小麦主产国的模型评估数据显示, 2010-2012年间高浓度O3浓度造成北半球和南半球小麦平均减产9.9%和6.2%, 抵消了因水肥管理或改良品种等措施对其增产所做的努力(Mills et al., 2018).我国这方面的研究起步较晚, 但发展迅速.从20世纪90年代末开始, 学者先后利用OTC和O3-FACE系统研究了地表O3浓度升高对我国主要粮食作物和蔬菜的影响, 涉及品种包括冬小麦、水稻、大豆、玉米、油菜、菠菜(Spinacia oleracea)和青菜(Brassica rapa)等(冯兆忠等, 2018).研究显示, 高浓度O3暴露下农作物均出现不同程度减产, 不同浓度O3对水稻、小麦、大豆等不同品种作物的减产幅度为10%-60% (贾一磊, 2016; 曹嘉晨等, 2017), 对油菜、菠菜和青菜等蔬菜生物产量的降幅为4.0%-89% (白月明等, 2002; 郭建平等, 2003). ...
... ), 作物产量将进一步降低10%- 20% (Feng & Kobayashi, 2009).最近, 有关美国、法国、印度、中国和俄罗斯等小麦主产国的模型评估数据显示, 2010-2012年间高浓度O3浓度造成北半球和南半球小麦平均减产9.9%和6.2%, 抵消了因水肥管理或改良品种等措施对其增产所做的努力(Mills et al., 2018).我国这方面的研究起步较晚, 但发展迅速.从20世纪90年代末开始, 学者先后利用OTC和O3-FACE系统研究了地表O3浓度升高对我国主要粮食作物和蔬菜的影响, 涉及品种包括冬小麦、水稻、大豆、玉米、油菜、菠菜(Spinacia oleracea)和青菜(Brassica rapa)等(冯兆忠等, 2018).研究显示, 高浓度O3暴露下农作物均出现不同程度减产, 不同浓度O3对水稻、小麦、大豆等不同品种作物的减产幅度为10%-60% (贾一磊, 2016; 曹嘉晨等, 2017), 对油菜、菠菜和青菜等蔬菜生物产量的降幅为4.0%-89% (白月明等, 2002; 郭建平等, 2003). ...
我国地表臭氧的生态环境效应研究进展
6
2018
... 地表臭氧(O3)特指距离地球表面15 km范围内的近地层O3, 除少量来自平流层大气传输外, 其余大部分是氮氧化物(NOx)、非甲烷类挥发性有机化合物(NMVOCs)、一氧化碳(CO)和甲烷(CH4)等前体物在强烈光照下发生光化学反应而产生(冯兆忠等, 2018).自工业革命以来, 伴随城市化进程加快和化石燃料的过度燃烧, 地表O3浓度在世界范围内普遍升高.近年来, 模型和观测数据都表明地表O3浓度小时峰值在欧洲和北美等地已有所下降, 但亚洲地区仍呈不断上升的趋势(Cooper et al., 2014).中国国家环境监测网络2013-2017年的数据表明, 过去5年中国地表O3浓度以每年3 μg·kg-1的速率攀升(Wang et al., 2019), 目前全国地表O3浓度每日8 h滑动平均最大值(MDA8)的年平均值已达(41.2 ± 6.3) μg·kg-1, 尤其华北平原、长江三角洲和珠江三角洲等地(MDA8峰值年平均值已处于60-70 μg·kg-1), 地表O3污染形势十分严峻(Lu et al., 2018).地表O3是具有植物毒性的气体污染物, 具有强氧化性.高浓度地表O3或长期低浓度O3能诱导植物叶片出现可见损伤, 加速植物衰老, 降低光合作用, 改变碳分配, 抑制植物生长和生物量积累, 影响作物品质和产量, 最终降低生态系统碳汇能力, 改变生态系统结构和功能(Ashmore, 2005; Ainsworth, 2017; 高峰, 2018). ...
... 气孔防御机制失效后, 进入细胞的O3将诱导叶片内部抗氧化系统启动解毒和修复的非气孔防御响应(冯兆忠等, 2018).非气孔防御响应主要是指“质外体-共质体系统”内一些抗氧化组分对O3解毒和修复的防御过程.具体来说, O3进入植物叶片的细胞间隙后会溶解在细胞壁的结合水里, 经过一系列的化学反应形成活性氧分子(ROS), ROS能够破坏细胞壁并与细胞膜上的不饱和脂肪酸发生反应, 从而对叶片造成损伤(Tausz et al., 2007; Pellegrini et al., 2018).植物质外体的抗氧化物质如还原性抗坏血酸(ASC)、抗氧化酶(如超氧化物歧化酶)可与ROS反应进行初级解毒, 成为抵御O3的第二道防线(Kangasjärvi et al., 2005; Dai et al., 2019), 而剩余的ROS会经过质膜到达共质体, 此时共质体内抗氧化成分如过氧化物酶(POD)、过氧化氢酶(CAT)、抗坏血酸过氧化物酶(APX)及抗氧化剂(如酚类化合物-谷胱甘肽(GSH)、脯氨酸和多胺等)等抗氧化酶类物质被诱导作为次级解毒响应机制参与诸如“抗坏血酸-谷胱甘肽(ASA-GSH)循环”等氧化还原过程, 进而对ROS进行进一步解毒与消除(Luwe, 1996).气孔闭合除了会减少O3的吸收, 也会抑制CO2吸收和水汽交换, 而非气孔防御响应中的抗氧化系统解毒过程也会消耗大量的能量并对光合反应系统造成不同程度的损伤, 降低羧化速率和电子传递速率, 进而抑制光合和蒸腾作用, 损害植物生长(Gao et al., 2016).然而, 最新研究表明持续整个生长季的长期O3暴露会影响植物叶片的气孔密度、开度和大小等表观属性(李品等, 2018), 而O3从气孔进入到细胞间隙后, 也可以通过影响植物的叶肉导度降低植物的光合能力(Xu et al., 2019).此外, 有关29种落叶和常绿树种应对O3胁迫的研究还发现不同木本植物对O3响应的差异主要来自植物功能叶片比叶质量的大小(即叶片比叶质量越大的树种对O3越不敏感), 与气孔导度无明显相关性(Li et al., 2016). ...
... O3不仅抑制植物生长, 还导致农作物大量减产(Fuhrer, 2009; Ainsworth et al., 2012; Feng et al., 2015a; Singh & Agrawal, 2017).有关O3对作物产量的影响已经在北美、欧洲和亚洲多地的几十种作物上有所研究报道, 包括对O3敏感的小麦、西瓜(Citrullus lanatus)、豆类、棉花(Gossypium hirsutum)、萝卜(Raphanus sativus)、番茄(Lycopersicon esclentum)、洋葱(Allium cepa)、大豆(Glycine max)和莴苣(Lactuca sativa), 对O3适度敏感的甜菜(Beta vulgaris)、马铃薯(Solanum tuberosum)、油菜(Brassica napus)、烟草(Nicotiana tabacum)、水稻、玉米(Zea mays)、葡萄和西兰花(Brassica oleracea), 和对O3耐受的作物大麦(Hordeum vulgare)、李(Prunus salicina)和草莓(Fragaria ananassa)等(Mills et al., 2007; Booker et al., 2009; Feng & Kobayashi, 2009; Avnery et al., 2011; Broberg et al., 2015).通过汇总406个控制实验的数据进行meta分析发现, 与活性炭过滤环境(≤26 μg·kg-1)相比, 当前环境O3浓度(平均值40 μg·kg-1)可导致主要粮食作物(小麦、水稻、大豆和马铃薯等)减产10%左右; 未来地表O3浓度升高情景下(51-71 μg·kg-1), 作物产量将进一步降低10%- 20% (Feng & Kobayashi, 2009).最近, 有关美国、法国、印度、中国和俄罗斯等小麦主产国的模型评估数据显示, 2010-2012年间高浓度O3浓度造成北半球和南半球小麦平均减产9.9%和6.2%, 抵消了因水肥管理或改良品种等措施对其增产所做的努力(Mills et al., 2018).我国这方面的研究起步较晚, 但发展迅速.从20世纪90年代末开始, 学者先后利用OTC和O3-FACE系统研究了地表O3浓度升高对我国主要粮食作物和蔬菜的影响, 涉及品种包括冬小麦、水稻、大豆、玉米、油菜、菠菜(Spinacia oleracea)和青菜(Brassica rapa)等(冯兆忠等, 2018).研究显示, 高浓度O3暴露下农作物均出现不同程度减产, 不同浓度O3对水稻、小麦、大豆等不同品种作物的减产幅度为10%-60% (贾一磊, 2016; 曹嘉晨等, 2017), 对油菜、菠菜和青菜等蔬菜生物产量的降幅为4.0%-89% (白月明等, 2002; 郭建平等, 2003). ...
... 相对于农作物产量, O3对粮食品质影响的研究集中在几种主要农作物(如小麦、水稻和大豆)(冯兆忠等, 2018).基于北美、欧洲和亚洲已有的O3对小麦影响的meta分析发现, O3显著降低小麦淀粉浓度, 但显著增加蛋白质和一些营养元素浓度(如钾、镁、钙、磷、锌、锰和铜)(Broberg et al., 2015).印度北部热带地区有关小麦的研究则显示, O3显著降低小麦籽粒糖含量, 但没有改变籽粒蛋白质含量(Tomer et al., 2015).有关水稻的研究还发现, O3增加粳稻直链淀粉含量, 但对籼稻直链淀粉含量无显著影响(Sawada et al., 2016).国内研究则表明, 尽管高浓度O3显著降低冬小麦面粉中支链淀粉和淀粉含量, 但是增加直链淀粉含量(Zhang et al., 2013).O3显著降低水稻稻米直链淀粉浓度、分解率, 增加黏度、起糊温度、蛋白质和微量矿物质浓度, 但蛋白质总量没有改变(Wang et al., 2012).此外, 研究还发现O3熏蒸显著降低了小麦籽粒出粉率和硬度, 但增加冬小麦面粉峰值黏度、低谷黏度、最终黏度和反弹值(贾一磊, 2016). ...
... (2)加强地表O3污染对地下生态过程的影响研究: 地下部分是生态系统水分供给、养分循环、碳分配的核心环节, 探究地下生态系统对地表O3浓度升高的响应对于准确理解地表O3浓度升高对整个生态系统的影响具有重要意义.事实上, 高浓度O3对地下部分的干扰比地上部分更明显且更早出现(Hofstra et al., 1981).然而, 目前有关O3对地下生态过程动态变化过程的研究还十分缺乏, 其响应机制尚不明确.已有的研究多集中在地表O3浓度升高对地下碳分配、根系生长和生物量的影响上, 少部分涉及凋落物、根系分泌物、土壤呼吸、微生物活性和结构等(Agathokleous et al., 2016), 而地表O3浓度升高对土壤微食物网的影响及其反馈几乎都没有涉及(Grulke & Heath, 2020), 更无法将土壤微生物过程(如微生物活性、生物量、群落组成、多样性和功能)与土壤关键生物地球化学循环过程, 以及与植物地上生态过程关联起来(Andersen, 2003; 冯兆忠等, 2018).未来的研究应加强O3对地下生态过程的长期定位研究, 特别是O3对地下碳分配动态、土壤微生物动态及根际氮循环的研究, 量化地下生态过程对O3响应的程度及其对生态系统长期稳定的反馈. ...
... (4)关注O3对作物品质及食品安全的危害: 世界上主要粮食作物要么对O3敏感(小麦、大豆等), 要么对O3适度敏感(玉米、水稻和马铃薯等).到2050年, 世界人口预计增加到90亿, 粮食供应需增加至少50%才能满足人口增长的需求, 如何保证粮食供应安全将成为21世纪最重要的一个挑战(Tai et al., 2014).目前为止, 除作物产量之外, 有关地表O3浓度升高对作物品质的影响研究多集中在蔬菜作物或粮食作物可食部位糖分、蛋白质和淀粉含量等食用口感方面(Booker et al., 2009), 很少涉及其背后对人体健康的直接影响.随着生活水平的提高, 人们更加关注食品的营养与安全(如微量元素含量或健康风险等), 而长期高浓度O3暴露可改变粮食作物籽粒微量元素及营养物质含量(Booker et al., 2009; 冯兆忠等, 2018).此外, 也有研究显示O3暴露导致马铃薯的糖苷生物碱含量增加, 糖苷生物碱不仅导致味苦而且浓度较高时具有毒性(Pell & Pearson, 1984).因此, 在全球环境变化大背景下, 长期食用暴露在O3污染下的粮食作物是否影响人体营养平衡或危及健康值得关注.未来的研究应关注高浓度O3对农作物外观品质(如籽粒形态、发芽率和成活率等)、加工特性、口感(如黏弹性、柔软性等)、烹饪特性和营养特性等外在和内在价值的综合影响. ...
我国地表臭氧的生态环境效应研究进展
6
2018
... 地表臭氧(O3)特指距离地球表面15 km范围内的近地层O3, 除少量来自平流层大气传输外, 其余大部分是氮氧化物(NOx)、非甲烷类挥发性有机化合物(NMVOCs)、一氧化碳(CO)和甲烷(CH4)等前体物在强烈光照下发生光化学反应而产生(冯兆忠等, 2018).自工业革命以来, 伴随城市化进程加快和化石燃料的过度燃烧, 地表O3浓度在世界范围内普遍升高.近年来, 模型和观测数据都表明地表O3浓度小时峰值在欧洲和北美等地已有所下降, 但亚洲地区仍呈不断上升的趋势(Cooper et al., 2014).中国国家环境监测网络2013-2017年的数据表明, 过去5年中国地表O3浓度以每年3 μg·kg-1的速率攀升(Wang et al., 2019), 目前全国地表O3浓度每日8 h滑动平均最大值(MDA8)的年平均值已达(41.2 ± 6.3) μg·kg-1, 尤其华北平原、长江三角洲和珠江三角洲等地(MDA8峰值年平均值已处于60-70 μg·kg-1), 地表O3污染形势十分严峻(Lu et al., 2018).地表O3是具有植物毒性的气体污染物, 具有强氧化性.高浓度地表O3或长期低浓度O3能诱导植物叶片出现可见损伤, 加速植物衰老, 降低光合作用, 改变碳分配, 抑制植物生长和生物量积累, 影响作物品质和产量, 最终降低生态系统碳汇能力, 改变生态系统结构和功能(Ashmore, 2005; Ainsworth, 2017; 高峰, 2018). ...
... 气孔防御机制失效后, 进入细胞的O3将诱导叶片内部抗氧化系统启动解毒和修复的非气孔防御响应(冯兆忠等, 2018).非气孔防御响应主要是指“质外体-共质体系统”内一些抗氧化组分对O3解毒和修复的防御过程.具体来说, O3进入植物叶片的细胞间隙后会溶解在细胞壁的结合水里, 经过一系列的化学反应形成活性氧分子(ROS), ROS能够破坏细胞壁并与细胞膜上的不饱和脂肪酸发生反应, 从而对叶片造成损伤(Tausz et al., 2007; Pellegrini et al., 2018).植物质外体的抗氧化物质如还原性抗坏血酸(ASC)、抗氧化酶(如超氧化物歧化酶)可与ROS反应进行初级解毒, 成为抵御O3的第二道防线(Kangasjärvi et al., 2005; Dai et al., 2019), 而剩余的ROS会经过质膜到达共质体, 此时共质体内抗氧化成分如过氧化物酶(POD)、过氧化氢酶(CAT)、抗坏血酸过氧化物酶(APX)及抗氧化剂(如酚类化合物-谷胱甘肽(GSH)、脯氨酸和多胺等)等抗氧化酶类物质被诱导作为次级解毒响应机制参与诸如“抗坏血酸-谷胱甘肽(ASA-GSH)循环”等氧化还原过程, 进而对ROS进行进一步解毒与消除(Luwe, 1996).气孔闭合除了会减少O3的吸收, 也会抑制CO2吸收和水汽交换, 而非气孔防御响应中的抗氧化系统解毒过程也会消耗大量的能量并对光合反应系统造成不同程度的损伤, 降低羧化速率和电子传递速率, 进而抑制光合和蒸腾作用, 损害植物生长(Gao et al., 2016).然而, 最新研究表明持续整个生长季的长期O3暴露会影响植物叶片的气孔密度、开度和大小等表观属性(李品等, 2018), 而O3从气孔进入到细胞间隙后, 也可以通过影响植物的叶肉导度降低植物的光合能力(Xu et al., 2019).此外, 有关29种落叶和常绿树种应对O3胁迫的研究还发现不同木本植物对O3响应的差异主要来自植物功能叶片比叶质量的大小(即叶片比叶质量越大的树种对O3越不敏感), 与气孔导度无明显相关性(Li et al., 2016). ...
... O3不仅抑制植物生长, 还导致农作物大量减产(Fuhrer, 2009; Ainsworth et al., 2012; Feng et al., 2015a; Singh & Agrawal, 2017).有关O3对作物产量的影响已经在北美、欧洲和亚洲多地的几十种作物上有所研究报道, 包括对O3敏感的小麦、西瓜(Citrullus lanatus)、豆类、棉花(Gossypium hirsutum)、萝卜(Raphanus sativus)、番茄(Lycopersicon esclentum)、洋葱(Allium cepa)、大豆(Glycine max)和莴苣(Lactuca sativa), 对O3适度敏感的甜菜(Beta vulgaris)、马铃薯(Solanum tuberosum)、油菜(Brassica napus)、烟草(Nicotiana tabacum)、水稻、玉米(Zea mays)、葡萄和西兰花(Brassica oleracea), 和对O3耐受的作物大麦(Hordeum vulgare)、李(Prunus salicina)和草莓(Fragaria ananassa)等(Mills et al., 2007; Booker et al., 2009; Feng & Kobayashi, 2009; Avnery et al., 2011; Broberg et al., 2015).通过汇总406个控制实验的数据进行meta分析发现, 与活性炭过滤环境(≤26 μg·kg-1)相比, 当前环境O3浓度(平均值40 μg·kg-1)可导致主要粮食作物(小麦、水稻、大豆和马铃薯等)减产10%左右; 未来地表O3浓度升高情景下(51-71 μg·kg-1), 作物产量将进一步降低10%- 20% (Feng & Kobayashi, 2009).最近, 有关美国、法国、印度、中国和俄罗斯等小麦主产国的模型评估数据显示, 2010-2012年间高浓度O3浓度造成北半球和南半球小麦平均减产9.9%和6.2%, 抵消了因水肥管理或改良品种等措施对其增产所做的努力(Mills et al., 2018).我国这方面的研究起步较晚, 但发展迅速.从20世纪90年代末开始, 学者先后利用OTC和O3-FACE系统研究了地表O3浓度升高对我国主要粮食作物和蔬菜的影响, 涉及品种包括冬小麦、水稻、大豆、玉米、油菜、菠菜(Spinacia oleracea)和青菜(Brassica rapa)等(冯兆忠等, 2018).研究显示, 高浓度O3暴露下农作物均出现不同程度减产, 不同浓度O3对水稻、小麦、大豆等不同品种作物的减产幅度为10%-60% (贾一磊, 2016; 曹嘉晨等, 2017), 对油菜、菠菜和青菜等蔬菜生物产量的降幅为4.0%-89% (白月明等, 2002; 郭建平等, 2003). ...
... 相对于农作物产量, O3对粮食品质影响的研究集中在几种主要农作物(如小麦、水稻和大豆)(冯兆忠等, 2018).基于北美、欧洲和亚洲已有的O3对小麦影响的meta分析发现, O3显著降低小麦淀粉浓度, 但显著增加蛋白质和一些营养元素浓度(如钾、镁、钙、磷、锌、锰和铜)(Broberg et al., 2015).印度北部热带地区有关小麦的研究则显示, O3显著降低小麦籽粒糖含量, 但没有改变籽粒蛋白质含量(Tomer et al., 2015).有关水稻的研究还发现, O3增加粳稻直链淀粉含量, 但对籼稻直链淀粉含量无显著影响(Sawada et al., 2016).国内研究则表明, 尽管高浓度O3显著降低冬小麦面粉中支链淀粉和淀粉含量, 但是增加直链淀粉含量(Zhang et al., 2013).O3显著降低水稻稻米直链淀粉浓度、分解率, 增加黏度、起糊温度、蛋白质和微量矿物质浓度, 但蛋白质总量没有改变(Wang et al., 2012).此外, 研究还发现O3熏蒸显著降低了小麦籽粒出粉率和硬度, 但增加冬小麦面粉峰值黏度、低谷黏度、最终黏度和反弹值(贾一磊, 2016). ...
... (2)加强地表O3污染对地下生态过程的影响研究: 地下部分是生态系统水分供给、养分循环、碳分配的核心环节, 探究地下生态系统对地表O3浓度升高的响应对于准确理解地表O3浓度升高对整个生态系统的影响具有重要意义.事实上, 高浓度O3对地下部分的干扰比地上部分更明显且更早出现(Hofstra et al., 1981).然而, 目前有关O3对地下生态过程动态变化过程的研究还十分缺乏, 其响应机制尚不明确.已有的研究多集中在地表O3浓度升高对地下碳分配、根系生长和生物量的影响上, 少部分涉及凋落物、根系分泌物、土壤呼吸、微生物活性和结构等(Agathokleous et al., 2016), 而地表O3浓度升高对土壤微食物网的影响及其反馈几乎都没有涉及(Grulke & Heath, 2020), 更无法将土壤微生物过程(如微生物活性、生物量、群落组成、多样性和功能)与土壤关键生物地球化学循环过程, 以及与植物地上生态过程关联起来(Andersen, 2003; 冯兆忠等, 2018).未来的研究应加强O3对地下生态过程的长期定位研究, 特别是O3对地下碳分配动态、土壤微生物动态及根际氮循环的研究, 量化地下生态过程对O3响应的程度及其对生态系统长期稳定的反馈. ...
... (4)关注O3对作物品质及食品安全的危害: 世界上主要粮食作物要么对O3敏感(小麦、大豆等), 要么对O3适度敏感(玉米、水稻和马铃薯等).到2050年, 世界人口预计增加到90亿, 粮食供应需增加至少50%才能满足人口增长的需求, 如何保证粮食供应安全将成为21世纪最重要的一个挑战(Tai et al., 2014).目前为止, 除作物产量之外, 有关地表O3浓度升高对作物品质的影响研究多集中在蔬菜作物或粮食作物可食部位糖分、蛋白质和淀粉含量等食用口感方面(Booker et al., 2009), 很少涉及其背后对人体健康的直接影响.随着生活水平的提高, 人们更加关注食品的营养与安全(如微量元素含量或健康风险等), 而长期高浓度O3暴露可改变粮食作物籽粒微量元素及营养物质含量(Booker et al., 2009; 冯兆忠等, 2018).此外, 也有研究显示O3暴露导致马铃薯的糖苷生物碱含量增加, 糖苷生物碱不仅导致味苦而且浓度较高时具有毒性(Pell & Pearson, 1984).因此, 在全球环境变化大背景下, 长期食用暴露在O3污染下的粮食作物是否影响人体营养平衡或危及健康值得关注.未来的研究应关注高浓度O3对农作物外观品质(如籽粒形态、发芽率和成活率等)、加工特性、口感(如黏弹性、柔软性等)、烹饪特性和营养特性等外在和内在价值的综合影响. ...
Air pollution affects food security in China: taking ozone as an example
1
2015
... (6)探究地表O3污染的生态控制措施: 欧洲和北美等发达国家治理大气污染的经验告诉我们, 严格的O3前体物NOx和挥发性有机化合物(VOCs)减排措施是降低地表O3浓度的有效手段(Feng et al., 2015b).然而, 以完全牺牲经济发展为代价的环境保护措施一般并不被发展中国家所接受.事实上, 自然植被与地表O3存在复杂的交互作用.一方面, 植物释放的VOCs有助于地表O3的形成; 另一方面, 植物在进行光合作用的同时也可以通过气孔、非气孔途径吸收或沉降大气O3 (Paoletti, 2009).考虑到植被VOCs释放和O3移除能力都具有树种特异性, 因此, 在一定条件下通过合理的树种筛选达到减少大气O3污染的目的将是完全可行的(Sicard et al., 2018).另外, 除植物自身遗传基因之外, 植物对O3胁迫的响应受土壤水分和营养元素如氮可利用性的影响(列淦文和薛立, 2014; 高峰, 2018).例如古巴农民通过提前1-2天灌溉的方式预防夏季高浓度O3对莴苣和烟草叶面的损伤(ICP Vegetation, 2011); 马来西亚研究者通过氮肥管理, 减轻O3污染对热带Elaeis guineensis产量和质量的伤害(Hewitt et al., 2009).因此通过合理的田间水肥管理措施(如改变灌溉模式、施肥剂量和方式)调控植物生长, 增强植物对O3的抗性, 进而减轻O3污染的生态效应是未来大气O3污染防治研究中值得思考的问题. ...
A stomatal ozone flux-response relationship to assess ozone-induced yield loss of winter wheat in subtropical China
1
2012
... 基于OTCs和O3-FACE平台下单个站点的控制实验可探究O3对植物个体、群落或生态系统的影响, 但将站点或样地尺度的研究外推到区域或全球, 评估O3污染区域风险的研究主要通过响应关系和模型.目前, 其研究领域主要集中在地表O3污染对农作物产量和森林生产力等方面.早在1994年, Chameides等(1994)就利用经济模型初步预测O3浓度升高可能导致2025年全球30%-75%的作物面临产量降低的风险.基于O3前体物排放、化学反应传输模型和M12、AOT40指标的评估显示, 2000年全球O3污染引起全球小麦、玉米和大豆产量平均降低2.2%-15%, 每年因上述作物产量降低所导致的经济损失达110-180亿美元(Avnery et al., 2011).最近, 基于PODY评估指标的研究则表明, O3对小麦造成的产量损失北半球(9.9%)明显高于南半球(6.2%), 平均产量损失最高的地区主要位于北半球暖温带湿润地区(12.5%)、热带湿热(12.5%)和热带潮湿气候区(17.2%)(Mills et al., 2018).在区域尺度上, 基于1990年O3浓度数据, Wang和Mauzerall (2004)利用M7和M12指标评估了O3对中国、日本和韩国主要农作物产量的影响, 结果显示O3污染造成小麦、玉米、棉花减产1%-9%, 大豆减产23%-27%, 造成2.4-35亿美元的经济损失, 在维持现有的污染物排放、管理措施和农业生产技术不变的情况下, 预计到2020年小麦、玉米、棉花减产幅度为2%-15%, 大豆为28%-35%.最近, 利用AOT40指标的研究显示, 2014年当前O3浓度导致中国农作物产量损失7 840万t, 小麦、水稻和玉米相对减产量分别为8.5%-14%、3.9%-15%和2.2%-5.5% (Lin et al., 2018).而只聚焦于长江三角洲地区的研究则发现, 2015年区域O3污染分别造成小麦和水稻减产9.9%-36.1%和7.3%- 23.9% (Zhao et al., 2018).虽然国内也有少量研究建立了PODY与个别树种或作物相对生物量或相对产量之间的通量响应关系(Feng et al., 2012; Hu et al., 2015; Gao et al., 2017), 但并没有研究利用PODY指标评估区域O3污染对农作物产量损失的影响. ...
Comparison of crop yield sensitivity to ozone between open-top chamber and free-air experiments
1
2018
... (1)开展O3-FACE研究: 现有研究多为OTC系统下的实验, OTC气室内“微气候” (如温度、水分、光照和风力等生态因素)与外界环境状态存在明显差异, 这些生态因子的变化可能影响植物应对O3的响应, 缓解或加重O3污染对农作物产量或森林植被生产力的损伤(Long et al., 2005; Feng et al., 2018a).全球模型评估结果显示, OTC实验结果低估了O3对作物产量损失的影响(van Dingenen et al., 2009).最近, Feng等(2018b)也发现, 同样浓度的O3对OTC实验下小麦和水稻产量的影响要低于FACE实验; 而对大豆产量的影响则相反.因此, OTC实验结果并不能完全应用于实际O3区域风险的评估.O3-FACE平台是未来研究实际大田环境下O3浓度升高对农林生态系统影响的主要依托平台.为减少区域O3风险评估的不确定性, 未来应加大O3-FACE平台的实验研究.考虑到目前已开展的FACE实验多坐落在温带地区, 主要探究温带植被或作物对高浓度O3的响应, 较少涉及热带和寒带植物或其他区域典型生态系统(Ainsworth et al., 2020).最理想的情况是, 不同区域、不同生态系统类型共同开展O3-FACE实验研究(Ainsworth et al., 2012). ...
Ozone risk for crops and pastures in present and future climates
1
2009
... O3不仅抑制植物生长, 还导致农作物大量减产(Fuhrer, 2009; Ainsworth et al., 2012; Feng et al., 2015a; Singh & Agrawal, 2017).有关O3对作物产量的影响已经在北美、欧洲和亚洲多地的几十种作物上有所研究报道, 包括对O3敏感的小麦、西瓜(Citrullus lanatus)、豆类、棉花(Gossypium hirsutum)、萝卜(Raphanus sativus)、番茄(Lycopersicon esclentum)、洋葱(Allium cepa)、大豆(Glycine max)和莴苣(Lactuca sativa), 对O3适度敏感的甜菜(Beta vulgaris)、马铃薯(Solanum tuberosum)、油菜(Brassica napus)、烟草(Nicotiana tabacum)、水稻、玉米(Zea mays)、葡萄和西兰花(Brassica oleracea), 和对O3耐受的作物大麦(Hordeum vulgare)、李(Prunus salicina)和草莓(Fragaria ananassa)等(Mills et al., 2007; Booker et al., 2009; Feng & Kobayashi, 2009; Avnery et al., 2011; Broberg et al., 2015).通过汇总406个控制实验的数据进行meta分析发现, 与活性炭过滤环境(≤26 μg·kg-1)相比, 当前环境O3浓度(平均值40 μg·kg-1)可导致主要粮食作物(小麦、水稻、大豆和马铃薯等)减产10%左右; 未来地表O3浓度升高情景下(51-71 μg·kg-1), 作物产量将进一步降低10%- 20% (Feng & Kobayashi, 2009).最近, 有关美国、法国、印度、中国和俄罗斯等小麦主产国的模型评估数据显示, 2010-2012年间高浓度O3浓度造成北半球和南半球小麦平均减产9.9%和6.2%, 抵消了因水肥管理或改良品种等措施对其增产所做的努力(Mills et al., 2018).我国这方面的研究起步较晚, 但发展迅速.从20世纪90年代末开始, 学者先后利用OTC和O3-FACE系统研究了地表O3浓度升高对我国主要粮食作物和蔬菜的影响, 涉及品种包括冬小麦、水稻、大豆、玉米、油菜、菠菜(Spinacia oleracea)和青菜(Brassica rapa)等(冯兆忠等, 2018).研究显示, 高浓度O3暴露下农作物均出现不同程度减产, 不同浓度O3对水稻、小麦、大豆等不同品种作物的减产幅度为10%-60% (贾一磊, 2016; 曹嘉晨等, 2017), 对油菜、菠菜和青菜等蔬菜生物产量的降幅为4.0%-89% (白月明等, 2002; 郭建平等, 2003). ...
Effects of ozone on managed pasture: II. Yield, species composition, canopy structure, and forage quality
1
1994
... 植物长期过量摄取O3还可以引起物种组成、冠层结构改变, 影响生态系统种群均匀度和丰富度, 威胁生态系统多样性(Fuhrer et al., 2016; Bergmann et al., 2017), 最终影响生态系统碳、氮和水循环(McLaughlin et al., 2007; Sun et al., 2012; Grulke & Heath, 2020).基于温室或OTCs控制实验下有关草本植物的多项研究都发现, 随着O3暴露时间的延长, 对O3相对敏感的物种/种群将会被对O3相对耐受的物种/种群替代(Fuhrer et al., 1994; Hayes et al., 2009), 非禾本科杂草与禾本科杂草的比例也在发生改变(Hayes et al., 2010).有关Aspen FACE森林生态系统的研究也表明, 尽管整个森林林分结构没有受到明显影响, 但敏感型欧洲山杨树种在O3熏蒸7年后几乎从森林冠层中消失(Kubiske et al., 2007).然而, 由于种内及种间复杂的竞争关系, O3对生态系统群落结构的影响有时候并不是统一的, 高浓度O3暴露环境下O3敏感型种群首先达到较高覆盖度的情况也时有发生(Evans & Ashmore, 1992; Barbo et al., 1998).此外, 由于生态系统各营养级之间的交互作用, 高浓度O3影响植物群落多样性的同时也潜在影响植物组织、化学组成、二次代谢产物(如酚苷类化合物、植物源挥发性有机化合物等)释放, 进而改变植物种群周边与之相关的动植物、真菌、细菌和昆虫行为或活动(ICP Vegetation, 2013; Fuhrer et al., 2016).例如, O3暴露使得西黄松(Pinus ponderosa)更易受到小蠹虫的袭击(Dahlsten et al., 1997), 但O3诱导红云杉(Picea rubens)萜烯物质释放增加的同时减少了其上蚜虫幼虫的数量(Cannon, 1990); 而有关欧洲山杨的研究则发现O3暴露降低柳皮苷和特里杨苷含量, 增加了缩合单宁含量, 引起林分中蚜虫和森林天幕毛虫幼虫数量剧增, 但蚜虫捕食者或天敌(如瓢虫、草蛉和蜘蛛)的数量显著减少(Percy et al., 2002; Kopper & Lindroth, 2003). ...
Critical levels for ozone effects on vegetation in Europe
1
1997
... 不管研究方法如何更新, 由于空间和时间上的限制, 模拟地表O3浓度升高的控制实验主要探究高浓度O3对植物损伤的机理, 而区域尺度上探究O3对陆地生态系统的影响却很难通过模拟实验获得.为评价区域O3风险和评估O3对生态系统影响带来的损失, 学者主要通过不同评估指标评估区域、国家或全球尺度上地表O3污染的生态效应.总体来说, 评估指标的研究主要经历了三个阶段, 分别是浓度响应关系、剂量响应关系和通量响应关系评估指标(Musselman et al., 2006).浓度响应关系评估指标主要包括M7/M12 (白天7 h (9:00-16:00) O3浓度平均值或白天12 h (8:00-20:00) O3浓度平均值), 早期曾广泛应用于表征O3浓度暴露与农作物产量损失相关性的研究中(Legge et al., 1995).剂量响应关系评估指标主要有SUM06 (O3小时浓度高于或等于60 μg·kg-1的累积值)(Lefohn & Foley, 1992), W126 (O3小时浓度在规定时段内的加权求和值)和AOTX (整个生长季太阳辐射>50 W·m-2时段内O3小时浓度超过X μg·kg-1的累计值, X普遍设为40 μg·kg-1)(Fuhrer et al., 1997; LRTAR Convention, 2015).剂量响应指标综合考虑O3浓度和暴露时间对植物生长的影响, 在世界范围内得到广泛应用. ...
Current and future ozone risks to global terrestrial biodiversity and ecosystem processes
5
2016
... 地表O3的发现最早可追溯到1839年(Leeds, 1880), 但直到1945年“洛杉矶烟雾”爆发后, 地表O3生态效应的研究才逐渐进入学者的视野(Rogers et al., 1956).经过近百年的发展, 涉及地表O3对植物、土壤及生态系统影响的研究, 已经遍及世界上大多数国家, 发表了2 500多篇期刊论文和十余本英文专著.到目前为止, O3表观伤害症状的描述及症状等级的鉴定(如叶片萎黄、褪绿和坏死等典型症状分类), O3伤害机理的探究(如O3降低光合速率的气孔与非气孔因素等), O3对植物叶片(光合作用、气孔导度及抗氧化系统等), 个体(生物量、产量或品质), 种群(种间关系、群落构成)及生态系统(生产力和固碳能力等)的影响, 及O3污染防治对策的探讨(前体物减排措施、化学防护剂的应用和作物栽培管理模式的筛选)等方面都进行了大量相关研究(Ashmore, 2005; Grulke & Heath, 2020).然而地表O3浓度升高对陆地生态系统水平碳、氮和水循环、营养交互、群落更替及地下生态系统动态变化过程(如根系功能变化、根际过程机制、土壤微生物活性及反馈等)方面的研究仍鲜有报道(Fuhrer et al., 2016).通过系统梳理和总结目前国内外研究成果, 本文对地表O3浓度升高对生态系统影响这一领域的研究历史、研究方法、评估指标、重大进展和存在的主要问题进行了回顾分析, 并对未来研究进行了展望. ...
... 植物长期过量摄取O3还可以引起物种组成、冠层结构改变, 影响生态系统种群均匀度和丰富度, 威胁生态系统多样性(Fuhrer et al., 2016; Bergmann et al., 2017), 最终影响生态系统碳、氮和水循环(McLaughlin et al., 2007; Sun et al., 2012; Grulke & Heath, 2020).基于温室或OTCs控制实验下有关草本植物的多项研究都发现, 随着O3暴露时间的延长, 对O3相对敏感的物种/种群将会被对O3相对耐受的物种/种群替代(Fuhrer et al., 1994; Hayes et al., 2009), 非禾本科杂草与禾本科杂草的比例也在发生改变(Hayes et al., 2010).有关Aspen FACE森林生态系统的研究也表明, 尽管整个森林林分结构没有受到明显影响, 但敏感型欧洲山杨树种在O3熏蒸7年后几乎从森林冠层中消失(Kubiske et al., 2007).然而, 由于种内及种间复杂的竞争关系, O3对生态系统群落结构的影响有时候并不是统一的, 高浓度O3暴露环境下O3敏感型种群首先达到较高覆盖度的情况也时有发生(Evans & Ashmore, 1992; Barbo et al., 1998).此外, 由于生态系统各营养级之间的交互作用, 高浓度O3影响植物群落多样性的同时也潜在影响植物组织、化学组成、二次代谢产物(如酚苷类化合物、植物源挥发性有机化合物等)释放, 进而改变植物种群周边与之相关的动植物、真菌、细菌和昆虫行为或活动(ICP Vegetation, 2013; Fuhrer et al., 2016).例如, O3暴露使得西黄松(Pinus ponderosa)更易受到小蠹虫的袭击(Dahlsten et al., 1997), 但O3诱导红云杉(Picea rubens)萜烯物质释放增加的同时减少了其上蚜虫幼虫的数量(Cannon, 1990); 而有关欧洲山杨的研究则发现O3暴露降低柳皮苷和特里杨苷含量, 增加了缩合单宁含量, 引起林分中蚜虫和森林天幕毛虫幼虫数量剧增, 但蚜虫捕食者或天敌(如瓢虫、草蛉和蜘蛛)的数量显著减少(Percy et al., 2002; Kopper & Lindroth, 2003). ...
... ; Fuhrer et al., 2016).例如, O3暴露使得西黄松(Pinus ponderosa)更易受到小蠹虫的袭击(Dahlsten et al., 1997), 但O3诱导红云杉(Picea rubens)萜烯物质释放增加的同时减少了其上蚜虫幼虫的数量(Cannon, 1990); 而有关欧洲山杨的研究则发现O3暴露降低柳皮苷和特里杨苷含量, 增加了缩合单宁含量, 引起林分中蚜虫和森林天幕毛虫幼虫数量剧增, 但蚜虫捕食者或天敌(如瓢虫、草蛉和蜘蛛)的数量显著减少(Percy et al., 2002; Kopper & Lindroth, 2003). ...
... 此外, 虽然地下生态系统并不是直接暴露在O3环境下, 但大量证据表明O3对地上生态系统的影响将会转移至地下(Andersen, 2003; Lindroth, 2010).如高浓度O3环境下, 植物分配到根的生物量将会大大减少, 基于100多组实验数据的meta分析研究表明O3暴露导致生物量分配到地下的量平均减少5.6% (Grantz et al., 2006).另外, 凋落物分解是生态系统营养循环的关键过程.通过影响凋落物的数量、质量和分解速率, 高浓度地表O3会间接改变土壤微生物群落活性、组成和结构, 土壤化学属性及土壤营养循环(Larson et al., 2002; Fuhrer et al., 2016).然而, 比较遗憾的是, 目前有关O3对地下生态过程影响的研究仅集中在农田生态系统, 仅有少部分研究涉及森林土壤(主要来自Aspen FACE)或草地生态系统(ICP Vegetation, 2013).而且, 由于凋落物种类、化学成分、分解者种群、土壤和大气环境条件的不同, 高浓度O3对凋落物分解动态的影响还存在很大争议(Gessner et al., 2010; Fuhrer et al., 2016; Grulke & Heath, 2020).例如, 研究发现O3对欧洲白桦(Betula pendula)叶片凋落物质量和损失率有明显影响(Kasurinen et al., 2006, 2007), 但不影响欧洲山毛榉凋落物的分解速率(Schloter et al., 2005).有关地表O3浓度升高对地下生态系统结构和功能的影响还有待进一步探究. ...
... ; Fuhrer et al., 2016; Grulke & Heath, 2020).例如, 研究发现O3对欧洲白桦(Betula pendula)叶片凋落物质量和损失率有明显影响(Kasurinen et al., 2006, 2007), 但不影响欧洲山毛榉凋落物的分解速率(Schloter et al., 2005).有关地表O3浓度升高对地下生态系统结构和功能的影响还有待进一步探究. ...
臭氧污染和干旱胁迫对杨树幼苗生长的影响机制研究
4
2018
... 地表臭氧(O3)特指距离地球表面15 km范围内的近地层O3, 除少量来自平流层大气传输外, 其余大部分是氮氧化物(NOx)、非甲烷类挥发性有机化合物(NMVOCs)、一氧化碳(CO)和甲烷(CH4)等前体物在强烈光照下发生光化学反应而产生(冯兆忠等, 2018).自工业革命以来, 伴随城市化进程加快和化石燃料的过度燃烧, 地表O3浓度在世界范围内普遍升高.近年来, 模型和观测数据都表明地表O3浓度小时峰值在欧洲和北美等地已有所下降, 但亚洲地区仍呈不断上升的趋势(Cooper et al., 2014).中国国家环境监测网络2013-2017年的数据表明, 过去5年中国地表O3浓度以每年3 μg·kg-1的速率攀升(Wang et al., 2019), 目前全国地表O3浓度每日8 h滑动平均最大值(MDA8)的年平均值已达(41.2 ± 6.3) μg·kg-1, 尤其华北平原、长江三角洲和珠江三角洲等地(MDA8峰值年平均值已处于60-70 μg·kg-1), 地表O3污染形势十分严峻(Lu et al., 2018).地表O3是具有植物毒性的气体污染物, 具有强氧化性.高浓度地表O3或长期低浓度O3能诱导植物叶片出现可见损伤, 加速植物衰老, 降低光合作用, 改变碳分配, 抑制植物生长和生物量积累, 影响作物品质和产量, 最终降低生态系统碳汇能力, 改变生态系统结构和功能(Ashmore, 2005; Ainsworth, 2017; 高峰, 2018). ...
... 除单因子O3胁迫外, 由于全球气候变化和极端气候常态化, 植物经常受到O3和其他环境因子交互作用的影响(如O3与CO2、升温、干旱或氮沉降等)(Schmadel-Hagebölling et al., 1998; Ollinger et al., 2002; Eltayeb et al., 2006; Ghimire et al., 2018).然而, 植物应对O3与其他因子交互作用的响应并不是两者或单独作用的简单叠加, 往往取决于植物种类、生长时期、对特定环境因子的敏感性、胁迫浓度及时间, 以及具体的微气候条件(列淦文和薛立, 2014).一方面, 通过诱导气孔关闭、限制叶片O3吸收(CO2浓度升高/干旱胁迫), 或提高碳同化速率、促进植物光合作用(CO2浓度升高/氮增加), CO2浓度升高、干旱胁迫和氮沉降经常被报道可增强植物抵御O3的能力, 减缓O3伤害(白月明等, 2005; Handley & Grulke, 2008; Gao et al., 2017).另一方面, 与之相反, 也有研究发现长期高浓度CO2或干旱降低气孔导度的同时, 会引起植物蒸腾速率、光合产物合成和积累的降低(McDonald et al., 2002; Pollastrini et al., 2010); 而超过植物承载阈值的高氮负荷也会对植物的光合生理活性产生负效应(Yamaguchi et al., 2010), 进而加重植物在复合环境下的碳损失、水分蒸散损失和自由基积累, 对植物产生协同伤害(Karnosky et al., 2005; 列淦文和薛立, 2014).高峰(2018)认为O3与其他环境胁迫因子出现的先后次序也会影响植物对复合作用的响应. ...
... (3)开展O3与其他环境因子复合研究: 全球气候变化、极端气候常态化和空气污染加重使两个或两个以上环境因子的交互作用大大增加, 复合作用下的实验条件也更接近植物生长的自然环境.现有的研究多集中在O3与CO2对植物或生态系统的共同影响, 并发现O3污染引起的负面作用经常被高浓度CO2对植物叶片光合作用或个体生物量累积的“肥效”所削弱(Ollinger et al., 2002).然而, 目前还无法确定这种结果是否与生态系统水平的响应相匹配(Grulke & Heath, 2020).植物应对O3胁迫的响应与土壤水分、氮有效性等因素密切相关(列淦文和薛立, 2014; 高峰, 2018).控制实验下的研究也发现, O3浓度的升高可能削弱植物对干旱或其他环境胁迫的抵抗力, 使生态系统更易受到病虫害的侵袭(Grulke et al., 2009).但是迄今为止, 国内外大部分研究多集中在温带农作物和森林树种上, 较少涉及亚热带地区乡土树种.由于生态系统类型差异, 各级营养交互及种间竞争的复杂性, 不同生态系统下O3与生物及非生物因子之间的多重交互作用机制还需进一步探究.此外, 已有研究多针对植物地上部分, 对植物地下部分及其功能变化与根际过程相互影响的机制、土壤关键过程研究较少(列淦文和薛立, 2014).因此, 在开展O3单因子实验研究的同时, 亟须考虑多个因子(如O3、CO2、水分和氮沉降等)共存对整个生态系统结构和功能的复合影响研究. ...
... (6)探究地表O3污染的生态控制措施: 欧洲和北美等发达国家治理大气污染的经验告诉我们, 严格的O3前体物NOx和挥发性有机化合物(VOCs)减排措施是降低地表O3浓度的有效手段(Feng et al., 2015b).然而, 以完全牺牲经济发展为代价的环境保护措施一般并不被发展中国家所接受.事实上, 自然植被与地表O3存在复杂的交互作用.一方面, 植物释放的VOCs有助于地表O3的形成; 另一方面, 植物在进行光合作用的同时也可以通过气孔、非气孔途径吸收或沉降大气O3 (Paoletti, 2009).考虑到植被VOCs释放和O3移除能力都具有树种特异性, 因此, 在一定条件下通过合理的树种筛选达到减少大气O3污染的目的将是完全可行的(Sicard et al., 2018).另外, 除植物自身遗传基因之外, 植物对O3胁迫的响应受土壤水分和营养元素如氮可利用性的影响(列淦文和薛立, 2014; 高峰, 2018).例如古巴农民通过提前1-2天灌溉的方式预防夏季高浓度O3对莴苣和烟草叶面的损伤(ICP Vegetation, 2011); 马来西亚研究者通过氮肥管理, 减轻O3污染对热带Elaeis guineensis产量和质量的伤害(Hewitt et al., 2009).因此通过合理的田间水肥管理措施(如改变灌溉模式、施肥剂量和方式)调控植物生长, 增强植物对O3的抗性, 进而减轻O3污染的生态效应是未来大气O3污染防治研究中值得思考的问题. ...
臭氧污染和干旱胁迫对杨树幼苗生长的影响机制研究
4
2018
... 地表臭氧(O3)特指距离地球表面15 km范围内的近地层O3, 除少量来自平流层大气传输外, 其余大部分是氮氧化物(NOx)、非甲烷类挥发性有机化合物(NMVOCs)、一氧化碳(CO)和甲烷(CH4)等前体物在强烈光照下发生光化学反应而产生(冯兆忠等, 2018).自工业革命以来, 伴随城市化进程加快和化石燃料的过度燃烧, 地表O3浓度在世界范围内普遍升高.近年来, 模型和观测数据都表明地表O3浓度小时峰值在欧洲和北美等地已有所下降, 但亚洲地区仍呈不断上升的趋势(Cooper et al., 2014).中国国家环境监测网络2013-2017年的数据表明, 过去5年中国地表O3浓度以每年3 μg·kg-1的速率攀升(Wang et al., 2019), 目前全国地表O3浓度每日8 h滑动平均最大值(MDA8)的年平均值已达(41.2 ± 6.3) μg·kg-1, 尤其华北平原、长江三角洲和珠江三角洲等地(MDA8峰值年平均值已处于60-70 μg·kg-1), 地表O3污染形势十分严峻(Lu et al., 2018).地表O3是具有植物毒性的气体污染物, 具有强氧化性.高浓度地表O3或长期低浓度O3能诱导植物叶片出现可见损伤, 加速植物衰老, 降低光合作用, 改变碳分配, 抑制植物生长和生物量积累, 影响作物品质和产量, 最终降低生态系统碳汇能力, 改变生态系统结构和功能(Ashmore, 2005; Ainsworth, 2017; 高峰, 2018). ...
... 除单因子O3胁迫外, 由于全球气候变化和极端气候常态化, 植物经常受到O3和其他环境因子交互作用的影响(如O3与CO2、升温、干旱或氮沉降等)(Schmadel-Hagebölling et al., 1998; Ollinger et al., 2002; Eltayeb et al., 2006; Ghimire et al., 2018).然而, 植物应对O3与其他因子交互作用的响应并不是两者或单独作用的简单叠加, 往往取决于植物种类、生长时期、对特定环境因子的敏感性、胁迫浓度及时间, 以及具体的微气候条件(列淦文和薛立, 2014).一方面, 通过诱导气孔关闭、限制叶片O3吸收(CO2浓度升高/干旱胁迫), 或提高碳同化速率、促进植物光合作用(CO2浓度升高/氮增加), CO2浓度升高、干旱胁迫和氮沉降经常被报道可增强植物抵御O3的能力, 减缓O3伤害(白月明等, 2005; Handley & Grulke, 2008; Gao et al., 2017).另一方面, 与之相反, 也有研究发现长期高浓度CO2或干旱降低气孔导度的同时, 会引起植物蒸腾速率、光合产物合成和积累的降低(McDonald et al., 2002; Pollastrini et al., 2010); 而超过植物承载阈值的高氮负荷也会对植物的光合生理活性产生负效应(Yamaguchi et al., 2010), 进而加重植物在复合环境下的碳损失、水分蒸散损失和自由基积累, 对植物产生协同伤害(Karnosky et al., 2005; 列淦文和薛立, 2014).高峰(2018)认为O3与其他环境胁迫因子出现的先后次序也会影响植物对复合作用的响应. ...
... (3)开展O3与其他环境因子复合研究: 全球气候变化、极端气候常态化和空气污染加重使两个或两个以上环境因子的交互作用大大增加, 复合作用下的实验条件也更接近植物生长的自然环境.现有的研究多集中在O3与CO2对植物或生态系统的共同影响, 并发现O3污染引起的负面作用经常被高浓度CO2对植物叶片光合作用或个体生物量累积的“肥效”所削弱(Ollinger et al., 2002).然而, 目前还无法确定这种结果是否与生态系统水平的响应相匹配(Grulke & Heath, 2020).植物应对O3胁迫的响应与土壤水分、氮有效性等因素密切相关(列淦文和薛立, 2014; 高峰, 2018).控制实验下的研究也发现, O3浓度的升高可能削弱植物对干旱或其他环境胁迫的抵抗力, 使生态系统更易受到病虫害的侵袭(Grulke et al., 2009).但是迄今为止, 国内外大部分研究多集中在温带农作物和森林树种上, 较少涉及亚热带地区乡土树种.由于生态系统类型差异, 各级营养交互及种间竞争的复杂性, 不同生态系统下O3与生物及非生物因子之间的多重交互作用机制还需进一步探究.此外, 已有研究多针对植物地上部分, 对植物地下部分及其功能变化与根际过程相互影响的机制、土壤关键过程研究较少(列淦文和薛立, 2014).因此, 在开展O3单因子实验研究的同时, 亟须考虑多个因子(如O3、CO2、水分和氮沉降等)共存对整个生态系统结构和功能的复合影响研究. ...
... (6)探究地表O3污染的生态控制措施: 欧洲和北美等发达国家治理大气污染的经验告诉我们, 严格的O3前体物NOx和挥发性有机化合物(VOCs)减排措施是降低地表O3浓度的有效手段(Feng et al., 2015b).然而, 以完全牺牲经济发展为代价的环境保护措施一般并不被发展中国家所接受.事实上, 自然植被与地表O3存在复杂的交互作用.一方面, 植物释放的VOCs有助于地表O3的形成; 另一方面, 植物在进行光合作用的同时也可以通过气孔、非气孔途径吸收或沉降大气O3 (Paoletti, 2009).考虑到植被VOCs释放和O3移除能力都具有树种特异性, 因此, 在一定条件下通过合理的树种筛选达到减少大气O3污染的目的将是完全可行的(Sicard et al., 2018).另外, 除植物自身遗传基因之外, 植物对O3胁迫的响应受土壤水分和营养元素如氮可利用性的影响(列淦文和薛立, 2014; 高峰, 2018).例如古巴农民通过提前1-2天灌溉的方式预防夏季高浓度O3对莴苣和烟草叶面的损伤(ICP Vegetation, 2011); 马来西亚研究者通过氮肥管理, 减轻O3污染对热带Elaeis guineensis产量和质量的伤害(Hewitt et al., 2009).因此通过合理的田间水肥管理措施(如改变灌溉模式、施肥剂量和方式)调控植物生长, 增强植物对O3的抗性, 进而减轻O3污染的生态效应是未来大气O3污染防治研究中值得思考的问题. ...
Effects of elevated ozone on physiological, anatomical and ultrastructural characteristics of four common urban tree species in China
1
2016
... 气孔防御机制失效后, 进入细胞的O3将诱导叶片内部抗氧化系统启动解毒和修复的非气孔防御响应(冯兆忠等, 2018).非气孔防御响应主要是指“质外体-共质体系统”内一些抗氧化组分对O3解毒和修复的防御过程.具体来说, O3进入植物叶片的细胞间隙后会溶解在细胞壁的结合水里, 经过一系列的化学反应形成活性氧分子(ROS), ROS能够破坏细胞壁并与细胞膜上的不饱和脂肪酸发生反应, 从而对叶片造成损伤(Tausz et al., 2007; Pellegrini et al., 2018).植物质外体的抗氧化物质如还原性抗坏血酸(ASC)、抗氧化酶(如超氧化物歧化酶)可与ROS反应进行初级解毒, 成为抵御O3的第二道防线(Kangasjärvi et al., 2005; Dai et al., 2019), 而剩余的ROS会经过质膜到达共质体, 此时共质体内抗氧化成分如过氧化物酶(POD)、过氧化氢酶(CAT)、抗坏血酸过氧化物酶(APX)及抗氧化剂(如酚类化合物-谷胱甘肽(GSH)、脯氨酸和多胺等)等抗氧化酶类物质被诱导作为次级解毒响应机制参与诸如“抗坏血酸-谷胱甘肽(ASA-GSH)循环”等氧化还原过程, 进而对ROS进行进一步解毒与消除(Luwe, 1996).气孔闭合除了会减少O3的吸收, 也会抑制CO2吸收和水汽交换, 而非气孔防御响应中的抗氧化系统解毒过程也会消耗大量的能量并对光合反应系统造成不同程度的损伤, 降低羧化速率和电子传递速率, 进而抑制光合和蒸腾作用, 损害植物生长(Gao et al., 2016).然而, 最新研究表明持续整个生长季的长期O3暴露会影响植物叶片的气孔密度、开度和大小等表观属性(李品等, 2018), 而O3从气孔进入到细胞间隙后, 也可以通过影响植物的叶肉导度降低植物的光合能力(Xu et al., 2019).此外, 有关29种落叶和常绿树种应对O3胁迫的研究还发现不同木本植物对O3响应的差异主要来自植物功能叶片比叶质量的大小(即叶片比叶质量越大的树种对O3越不敏感), 与气孔导度无明显相关性(Li et al., 2016). ...
Water stress mitigates the negative effects of ozone on photosynthesis and biomass in poplar plants
3
2017
... 然而, O3对植物的影响不仅依赖于大气O3浓度同时也依赖于叶片气孔导度(O3的吸收)和植物自身抗氧化能力(Matyssek et al., 2007).近年来, 基于气孔O3吸收量的评估指标PODY (植物毒性O3剂量, 整个生长季单位面积上气孔O3吸收通量超过临界值Y nmol m-2·s-1的积累量)被开发并逐步替代浓度暴露剂量指标, 用于区域或全球O3对农作物和森林树种生长影响的评估(Büker et al., 2015; LRTAR Convention, 2015).目前, 哪个指标更适合区域O3风险评估的争论一直存在, 但欧洲和亚洲地区多项O3对农作物或植被影响的研究均表明: 考虑水分供应的情况下, O3暴露造成的生物量或作物产量的降低与PODY的拟合度比AOTX更好(Mills et al., 2011; Gao et al., 2017), 能更好表征O3对生态系统的影响.整体来看, 各个评估指标的计算方式和所需参数从简单到复杂, 从单一考虑O3浓度到综合考虑环境因子对植物O3吸收通量的影响, 逐渐可以精确评估地表O3污染对生态系统农林生产力的影响(表3). ...
... 除单因子O3胁迫外, 由于全球气候变化和极端气候常态化, 植物经常受到O3和其他环境因子交互作用的影响(如O3与CO2、升温、干旱或氮沉降等)(Schmadel-Hagebölling et al., 1998; Ollinger et al., 2002; Eltayeb et al., 2006; Ghimire et al., 2018).然而, 植物应对O3与其他因子交互作用的响应并不是两者或单独作用的简单叠加, 往往取决于植物种类、生长时期、对特定环境因子的敏感性、胁迫浓度及时间, 以及具体的微气候条件(列淦文和薛立, 2014).一方面, 通过诱导气孔关闭、限制叶片O3吸收(CO2浓度升高/干旱胁迫), 或提高碳同化速率、促进植物光合作用(CO2浓度升高/氮增加), CO2浓度升高、干旱胁迫和氮沉降经常被报道可增强植物抵御O3的能力, 减缓O3伤害(白月明等, 2005; Handley & Grulke, 2008; Gao et al., 2017).另一方面, 与之相反, 也有研究发现长期高浓度CO2或干旱降低气孔导度的同时, 会引起植物蒸腾速率、光合产物合成和积累的降低(McDonald et al., 2002; Pollastrini et al., 2010); 而超过植物承载阈值的高氮负荷也会对植物的光合生理活性产生负效应(Yamaguchi et al., 2010), 进而加重植物在复合环境下的碳损失、水分蒸散损失和自由基积累, 对植物产生协同伤害(Karnosky et al., 2005; 列淦文和薛立, 2014).高峰(2018)认为O3与其他环境胁迫因子出现的先后次序也会影响植物对复合作用的响应. ...
... 基于OTCs和O3-FACE平台下单个站点的控制实验可探究O3对植物个体、群落或生态系统的影响, 但将站点或样地尺度的研究外推到区域或全球, 评估O3污染区域风险的研究主要通过响应关系和模型.目前, 其研究领域主要集中在地表O3污染对农作物产量和森林生产力等方面.早在1994年, Chameides等(1994)就利用经济模型初步预测O3浓度升高可能导致2025年全球30%-75%的作物面临产量降低的风险.基于O3前体物排放、化学反应传输模型和M12、AOT40指标的评估显示, 2000年全球O3污染引起全球小麦、玉米和大豆产量平均降低2.2%-15%, 每年因上述作物产量降低所导致的经济损失达110-180亿美元(Avnery et al., 2011).最近, 基于PODY评估指标的研究则表明, O3对小麦造成的产量损失北半球(9.9%)明显高于南半球(6.2%), 平均产量损失最高的地区主要位于北半球暖温带湿润地区(12.5%)、热带湿热(12.5%)和热带潮湿气候区(17.2%)(Mills et al., 2018).在区域尺度上, 基于1990年O3浓度数据, Wang和Mauzerall (2004)利用M7和M12指标评估了O3对中国、日本和韩国主要农作物产量的影响, 结果显示O3污染造成小麦、玉米、棉花减产1%-9%, 大豆减产23%-27%, 造成2.4-35亿美元的经济损失, 在维持现有的污染物排放、管理措施和农业生产技术不变的情况下, 预计到2020年小麦、玉米、棉花减产幅度为2%-15%, 大豆为28%-35%.最近, 利用AOT40指标的研究显示, 2014年当前O3浓度导致中国农作物产量损失7 840万t, 小麦、水稻和玉米相对减产量分别为8.5%-14%、3.9%-15%和2.2%-5.5% (Lin et al., 2018).而只聚焦于长江三角洲地区的研究则发现, 2015年区域O3污染分别造成小麦和水稻减产9.9%-36.1%和7.3%- 23.9% (Zhao et al., 2018).虽然国内也有少量研究建立了PODY与个别树种或作物相对生物量或相对产量之间的通量响应关系(Feng et al., 2012; Hu et al., 2015; Gao et al., 2017), 但并没有研究利用PODY指标评估区域O3污染对农作物产量损失的影响. ...
Diversity meets decompositon
1
2010
... 此外, 虽然地下生态系统并不是直接暴露在O3环境下, 但大量证据表明O3对地上生态系统的影响将会转移至地下(Andersen, 2003; Lindroth, 2010).如高浓度O3环境下, 植物分配到根的生物量将会大大减少, 基于100多组实验数据的meta分析研究表明O3暴露导致生物量分配到地下的量平均减少5.6% (Grantz et al., 2006).另外, 凋落物分解是生态系统营养循环的关键过程.通过影响凋落物的数量、质量和分解速率, 高浓度地表O3会间接改变土壤微生物群落活性、组成和结构, 土壤化学属性及土壤营养循环(Larson et al., 2002; Fuhrer et al., 2016).然而, 比较遗憾的是, 目前有关O3对地下生态过程影响的研究仅集中在农田生态系统, 仅有少部分研究涉及森林土壤(主要来自Aspen FACE)或草地生态系统(ICP Vegetation, 2013).而且, 由于凋落物种类、化学成分、分解者种群、土壤和大气环境条件的不同, 高浓度O3对凋落物分解动态的影响还存在很大争议(Gessner et al., 2010; Fuhrer et al., 2016; Grulke & Heath, 2020).例如, 研究发现O3对欧洲白桦(Betula pendula)叶片凋落物质量和损失率有明显影响(Kasurinen et al., 2006, 2007), 但不影响欧洲山毛榉凋落物的分解速率(Schloter et al., 2005).有关地表O3浓度升高对地下生态系统结构和功能的影响还有待进一步探究. ...
Combined effects of elevated ozone, temperature, and nitrogen on stem phenolic concentrations of Scots pine (Pinus sylvestris) seedlings
1
2018
... 除单因子O3胁迫外, 由于全球气候变化和极端气候常态化, 植物经常受到O3和其他环境因子交互作用的影响(如O3与CO2、升温、干旱或氮沉降等)(Schmadel-Hagebölling et al., 1998; Ollinger et al., 2002; Eltayeb et al., 2006; Ghimire et al., 2018).然而, 植物应对O3与其他因子交互作用的响应并不是两者或单独作用的简单叠加, 往往取决于植物种类、生长时期、对特定环境因子的敏感性、胁迫浓度及时间, 以及具体的微气候条件(列淦文和薛立, 2014).一方面, 通过诱导气孔关闭、限制叶片O3吸收(CO2浓度升高/干旱胁迫), 或提高碳同化速率、促进植物光合作用(CO2浓度升高/氮增加), CO2浓度升高、干旱胁迫和氮沉降经常被报道可增强植物抵御O3的能力, 减缓O3伤害(白月明等, 2005; Handley & Grulke, 2008; Gao et al., 2017).另一方面, 与之相反, 也有研究发现长期高浓度CO2或干旱降低气孔导度的同时, 会引起植物蒸腾速率、光合产物合成和积累的降低(McDonald et al., 2002; Pollastrini et al., 2010); 而超过植物承载阈值的高氮负荷也会对植物的光合生理活性产生负效应(Yamaguchi et al., 2010), 进而加重植物在复合环境下的碳损失、水分蒸散损失和自由基积累, 对植物产生协同伤害(Karnosky et al., 2005; 列淦文和薛立, 2014).高峰(2018)认为O3与其他环境胁迫因子出现的先后次序也会影响植物对复合作用的响应. ...
O3 impacts on plant development: a meta-analysis of root/shoot allocation and growth
1
2006
... 此外, 虽然地下生态系统并不是直接暴露在O3环境下, 但大量证据表明O3对地上生态系统的影响将会转移至地下(Andersen, 2003; Lindroth, 2010).如高浓度O3环境下, 植物分配到根的生物量将会大大减少, 基于100多组实验数据的meta分析研究表明O3暴露导致生物量分配到地下的量平均减少5.6% (Grantz et al., 2006).另外, 凋落物分解是生态系统营养循环的关键过程.通过影响凋落物的数量、质量和分解速率, 高浓度地表O3会间接改变土壤微生物群落活性、组成和结构, 土壤化学属性及土壤营养循环(Larson et al., 2002; Fuhrer et al., 2016).然而, 比较遗憾的是, 目前有关O3对地下生态过程影响的研究仅集中在农田生态系统, 仅有少部分研究涉及森林土壤(主要来自Aspen FACE)或草地生态系统(ICP Vegetation, 2013).而且, 由于凋落物种类、化学成分、分解者种群、土壤和大气环境条件的不同, 高浓度O3对凋落物分解动态的影响还存在很大争议(Gessner et al., 2010; Fuhrer et al., 2016; Grulke & Heath, 2020).例如, 研究发现O3对欧洲白桦(Betula pendula)叶片凋落物质量和损失率有明显影响(Kasurinen et al., 2006, 2007), 但不影响欧洲山毛榉凋落物的分解速率(Schloter et al., 2005).有关地表O3浓度升高对地下生态系统结构和功能的影响还有待进一步探究. ...
Ozone effects on plants in natural ecosystems
6
2020
... 地表O3的发现最早可追溯到1839年(Leeds, 1880), 但直到1945年“洛杉矶烟雾”爆发后, 地表O3生态效应的研究才逐渐进入学者的视野(Rogers et al., 1956).经过近百年的发展, 涉及地表O3对植物、土壤及生态系统影响的研究, 已经遍及世界上大多数国家, 发表了2 500多篇期刊论文和十余本英文专著.到目前为止, O3表观伤害症状的描述及症状等级的鉴定(如叶片萎黄、褪绿和坏死等典型症状分类), O3伤害机理的探究(如O3降低光合速率的气孔与非气孔因素等), O3对植物叶片(光合作用、气孔导度及抗氧化系统等), 个体(生物量、产量或品质), 种群(种间关系、群落构成)及生态系统(生产力和固碳能力等)的影响, 及O3污染防治对策的探讨(前体物减排措施、化学防护剂的应用和作物栽培管理模式的筛选)等方面都进行了大量相关研究(Ashmore, 2005; Grulke & Heath, 2020).然而地表O3浓度升高对陆地生态系统水平碳、氮和水循环、营养交互、群落更替及地下生态系统动态变化过程(如根系功能变化、根际过程机制、土壤微生物活性及反馈等)方面的研究仍鲜有报道(Fuhrer et al., 2016).通过系统梳理和总结目前国内外研究成果, 本文对地表O3浓度升高对生态系统影响这一领域的研究历史、研究方法、评估指标、重大进展和存在的主要问题进行了回顾分析, 并对未来研究进行了展望. ...
... 在此期间, 尽管社会经济不断发展、实验设备和研究方法不断更新, 但地表O3污染对植物及生态系统影响的研究主要还是以模拟O3浓度升高的控制实验为主.纵观发展历程, 试验设备与手段的改进和更新大致可分为三个阶段(王春乙, 1995): (1) 1970年之前, 研究主要利用封闭式静态或动态气室或室内生长箱进行模拟实验(Heagle et al., 1973). (2) 1973年之后, 美国学者Heagle等(1979)和Mandle (1973)研制开顶式气室(OTC)模拟高浓度O3对植物的损伤和影响机制, 并最终在世界范围内推广, 成为研究地表O3浓度升高对植物及生态系统影响的主流手段.(3)由于OTCs气室内外较大的环境差异, OTCs模拟实验下的熏蒸结果是否适用于大田研究一直备受争议.为解决OTCs内微气候环境带来的影响, 1986年Hendrey和Kimball (1994)提出建立自由空气中气体浓度增加(FACE)系统的想法, 并逐步用于大田实验.由于与外界大气几乎一致的气体流速、温度、光照和风速等条件(Matyssek et al., 2010), FACE系统被普遍认为是当前研究地表O3浓度升高对植物及生态系统影响最为理想的实验平台, 但由于造价昂贵, 技术要求高, 只有少数国家和实验室开展相关研究(有关上述研究方法的优缺点见表2).O3熏蒸的FACE平台仅有10个左右且多数已经停止运行(Paoletti et al., 2017), 主要包括美国威斯康星州Aspen FACE (停止运行; Karnosky et al., 2003), 美国伊利诺伊州Soybean FACE (停止运行; Morgan et al., 2004), 瑞典苏尔草地FACE (停止运行; Volk et al., 2003), 芬兰库奥皮奥FACE (停止运行; Oksanen, 2003), 德国克兰兹堡森林FACE (停止运行; Matyssek et al., 2010), 中国江都水稻(Oryza sativa)/小麦(Triticum aestivum)轮作FACE (停止运行; Tang et al., 2011), 日本札幌森林FACE (仍在运行; Watanabe et al., 2013), 意大利佛罗伦萨3D FACE (仍在运行; Paoletti et al., 2017), 以及刚刚建成的中国延庆唐家堡杨树人工林FACE.由于实验条件和FACE平台规模的限制, 有关地表O3浓度升高对生态系统水平的研究成果主要来自美国Aspen FACE和德国克兰兹堡森林FACE (Grulke & Heath, 2020). ...
... 植物长期过量摄取O3还可以引起物种组成、冠层结构改变, 影响生态系统种群均匀度和丰富度, 威胁生态系统多样性(Fuhrer et al., 2016; Bergmann et al., 2017), 最终影响生态系统碳、氮和水循环(McLaughlin et al., 2007; Sun et al., 2012; Grulke & Heath, 2020).基于温室或OTCs控制实验下有关草本植物的多项研究都发现, 随着O3暴露时间的延长, 对O3相对敏感的物种/种群将会被对O3相对耐受的物种/种群替代(Fuhrer et al., 1994; Hayes et al., 2009), 非禾本科杂草与禾本科杂草的比例也在发生改变(Hayes et al., 2010).有关Aspen FACE森林生态系统的研究也表明, 尽管整个森林林分结构没有受到明显影响, 但敏感型欧洲山杨树种在O3熏蒸7年后几乎从森林冠层中消失(Kubiske et al., 2007).然而, 由于种内及种间复杂的竞争关系, O3对生态系统群落结构的影响有时候并不是统一的, 高浓度O3暴露环境下O3敏感型种群首先达到较高覆盖度的情况也时有发生(Evans & Ashmore, 1992; Barbo et al., 1998).此外, 由于生态系统各营养级之间的交互作用, 高浓度O3影响植物群落多样性的同时也潜在影响植物组织、化学组成、二次代谢产物(如酚苷类化合物、植物源挥发性有机化合物等)释放, 进而改变植物种群周边与之相关的动植物、真菌、细菌和昆虫行为或活动(ICP Vegetation, 2013; Fuhrer et al., 2016).例如, O3暴露使得西黄松(Pinus ponderosa)更易受到小蠹虫的袭击(Dahlsten et al., 1997), 但O3诱导红云杉(Picea rubens)萜烯物质释放增加的同时减少了其上蚜虫幼虫的数量(Cannon, 1990); 而有关欧洲山杨的研究则发现O3暴露降低柳皮苷和特里杨苷含量, 增加了缩合单宁含量, 引起林分中蚜虫和森林天幕毛虫幼虫数量剧增, 但蚜虫捕食者或天敌(如瓢虫、草蛉和蜘蛛)的数量显著减少(Percy et al., 2002; Kopper & Lindroth, 2003). ...
... 此外, 虽然地下生态系统并不是直接暴露在O3环境下, 但大量证据表明O3对地上生态系统的影响将会转移至地下(Andersen, 2003; Lindroth, 2010).如高浓度O3环境下, 植物分配到根的生物量将会大大减少, 基于100多组实验数据的meta分析研究表明O3暴露导致生物量分配到地下的量平均减少5.6% (Grantz et al., 2006).另外, 凋落物分解是生态系统营养循环的关键过程.通过影响凋落物的数量、质量和分解速率, 高浓度地表O3会间接改变土壤微生物群落活性、组成和结构, 土壤化学属性及土壤营养循环(Larson et al., 2002; Fuhrer et al., 2016).然而, 比较遗憾的是, 目前有关O3对地下生态过程影响的研究仅集中在农田生态系统, 仅有少部分研究涉及森林土壤(主要来自Aspen FACE)或草地生态系统(ICP Vegetation, 2013).而且, 由于凋落物种类、化学成分、分解者种群、土壤和大气环境条件的不同, 高浓度O3对凋落物分解动态的影响还存在很大争议(Gessner et al., 2010; Fuhrer et al., 2016; Grulke & Heath, 2020).例如, 研究发现O3对欧洲白桦(Betula pendula)叶片凋落物质量和损失率有明显影响(Kasurinen et al., 2006, 2007), 但不影响欧洲山毛榉凋落物的分解速率(Schloter et al., 2005).有关地表O3浓度升高对地下生态系统结构和功能的影响还有待进一步探究. ...
... (2)加强地表O3污染对地下生态过程的影响研究: 地下部分是生态系统水分供给、养分循环、碳分配的核心环节, 探究地下生态系统对地表O3浓度升高的响应对于准确理解地表O3浓度升高对整个生态系统的影响具有重要意义.事实上, 高浓度O3对地下部分的干扰比地上部分更明显且更早出现(Hofstra et al., 1981).然而, 目前有关O3对地下生态过程动态变化过程的研究还十分缺乏, 其响应机制尚不明确.已有的研究多集中在地表O3浓度升高对地下碳分配、根系生长和生物量的影响上, 少部分涉及凋落物、根系分泌物、土壤呼吸、微生物活性和结构等(Agathokleous et al., 2016), 而地表O3浓度升高对土壤微食物网的影响及其反馈几乎都没有涉及(Grulke & Heath, 2020), 更无法将土壤微生物过程(如微生物活性、生物量、群落组成、多样性和功能)与土壤关键生物地球化学循环过程, 以及与植物地上生态过程关联起来(Andersen, 2003; 冯兆忠等, 2018).未来的研究应加强O3对地下生态过程的长期定位研究, 特别是O3对地下碳分配动态、土壤微生物动态及根际氮循环的研究, 量化地下生态过程对O3响应的程度及其对生态系统长期稳定的反馈. ...
... (3)开展O3与其他环境因子复合研究: 全球气候变化、极端气候常态化和空气污染加重使两个或两个以上环境因子的交互作用大大增加, 复合作用下的实验条件也更接近植物生长的自然环境.现有的研究多集中在O3与CO2对植物或生态系统的共同影响, 并发现O3污染引起的负面作用经常被高浓度CO2对植物叶片光合作用或个体生物量累积的“肥效”所削弱(Ollinger et al., 2002).然而, 目前还无法确定这种结果是否与生态系统水平的响应相匹配(Grulke & Heath, 2020).植物应对O3胁迫的响应与土壤水分、氮有效性等因素密切相关(列淦文和薛立, 2014; 高峰, 2018).控制实验下的研究也发现, O3浓度的升高可能削弱植物对干旱或其他环境胁迫的抵抗力, 使生态系统更易受到病虫害的侵袭(Grulke et al., 2009).但是迄今为止, 国内外大部分研究多集中在温带农作物和森林树种上, 较少涉及亚热带地区乡土树种.由于生态系统类型差异, 各级营养交互及种间竞争的复杂性, 不同生态系统下O3与生物及非生物因子之间的多重交互作用机制还需进一步探究.此外, 已有研究多针对植物地上部分, 对植物地下部分及其功能变化与根际过程相互影响的机制、土壤关键过程研究较少(列淦文和薛立, 2014).因此, 在开展O3单因子实验研究的同时, 亟须考虑多个因子(如O3、CO2、水分和氮沉降等)共存对整个生态系统结构和功能的复合影响研究. ...
Chapter 17 Air pollution increases forest susceptibility to wildfires: a case study in the San Bernardino Mountains in southern California
1
2009
... (3)开展O3与其他环境因子复合研究: 全球气候变化、极端气候常态化和空气污染加重使两个或两个以上环境因子的交互作用大大增加, 复合作用下的实验条件也更接近植物生长的自然环境.现有的研究多集中在O3与CO2对植物或生态系统的共同影响, 并发现O3污染引起的负面作用经常被高浓度CO2对植物叶片光合作用或个体生物量累积的“肥效”所削弱(Ollinger et al., 2002).然而, 目前还无法确定这种结果是否与生态系统水平的响应相匹配(Grulke & Heath, 2020).植物应对O3胁迫的响应与土壤水分、氮有效性等因素密切相关(列淦文和薛立, 2014; 高峰, 2018).控制实验下的研究也发现, O3浓度的升高可能削弱植物对干旱或其他环境胁迫的抵抗力, 使生态系统更易受到病虫害的侵袭(Grulke et al., 2009).但是迄今为止, 国内外大部分研究多集中在温带农作物和森林树种上, 较少涉及亚热带地区乡土树种.由于生态系统类型差异, 各级营养交互及种间竞争的复杂性, 不同生态系统下O3与生物及非生物因子之间的多重交互作用机制还需进一步探究.此外, 已有研究多针对植物地上部分, 对植物地下部分及其功能变化与根际过程相互影响的机制、土壤关键过程研究较少(列淦文和薛立, 2014).因此, 在开展O3单因子实验研究的同时, 亟须考虑多个因子(如O3、CO2、水分和氮沉降等)共存对整个生态系统结构和功能的复合影响研究. ...
大气中臭氧浓度变化对蔬菜的影响研究
1
2003
... O3不仅抑制植物生长, 还导致农作物大量减产(Fuhrer, 2009; Ainsworth et al., 2012; Feng et al., 2015a; Singh & Agrawal, 2017).有关O3对作物产量的影响已经在北美、欧洲和亚洲多地的几十种作物上有所研究报道, 包括对O3敏感的小麦、西瓜(Citrullus lanatus)、豆类、棉花(Gossypium hirsutum)、萝卜(Raphanus sativus)、番茄(Lycopersicon esclentum)、洋葱(Allium cepa)、大豆(Glycine max)和莴苣(Lactuca sativa), 对O3适度敏感的甜菜(Beta vulgaris)、马铃薯(Solanum tuberosum)、油菜(Brassica napus)、烟草(Nicotiana tabacum)、水稻、玉米(Zea mays)、葡萄和西兰花(Brassica oleracea), 和对O3耐受的作物大麦(Hordeum vulgare)、李(Prunus salicina)和草莓(Fragaria ananassa)等(Mills et al., 2007; Booker et al., 2009; Feng & Kobayashi, 2009; Avnery et al., 2011; Broberg et al., 2015).通过汇总406个控制实验的数据进行meta分析发现, 与活性炭过滤环境(≤26 μg·kg-1)相比, 当前环境O3浓度(平均值40 μg·kg-1)可导致主要粮食作物(小麦、水稻、大豆和马铃薯等)减产10%左右; 未来地表O3浓度升高情景下(51-71 μg·kg-1), 作物产量将进一步降低10%- 20% (Feng & Kobayashi, 2009).最近, 有关美国、法国、印度、中国和俄罗斯等小麦主产国的模型评估数据显示, 2010-2012年间高浓度O3浓度造成北半球和南半球小麦平均减产9.9%和6.2%, 抵消了因水肥管理或改良品种等措施对其增产所做的努力(Mills et al., 2018).我国这方面的研究起步较晚, 但发展迅速.从20世纪90年代末开始, 学者先后利用OTC和O3-FACE系统研究了地表O3浓度升高对我国主要粮食作物和蔬菜的影响, 涉及品种包括冬小麦、水稻、大豆、玉米、油菜、菠菜(Spinacia oleracea)和青菜(Brassica rapa)等(冯兆忠等, 2018).研究显示, 高浓度O3暴露下农作物均出现不同程度减产, 不同浓度O3对水稻、小麦、大豆等不同品种作物的减产幅度为10%-60% (贾一磊, 2016; 曹嘉晨等, 2017), 对油菜、菠菜和青菜等蔬菜生物产量的降幅为4.0%-89% (白月明等, 2002; 郭建平等, 2003). ...
大气中臭氧浓度变化对蔬菜的影响研究
1
2003
... O3不仅抑制植物生长, 还导致农作物大量减产(Fuhrer, 2009; Ainsworth et al., 2012; Feng et al., 2015a; Singh & Agrawal, 2017).有关O3对作物产量的影响已经在北美、欧洲和亚洲多地的几十种作物上有所研究报道, 包括对O3敏感的小麦、西瓜(Citrullus lanatus)、豆类、棉花(Gossypium hirsutum)、萝卜(Raphanus sativus)、番茄(Lycopersicon esclentum)、洋葱(Allium cepa)、大豆(Glycine max)和莴苣(Lactuca sativa), 对O3适度敏感的甜菜(Beta vulgaris)、马铃薯(Solanum tuberosum)、油菜(Brassica napus)、烟草(Nicotiana tabacum)、水稻、玉米(Zea mays)、葡萄和西兰花(Brassica oleracea), 和对O3耐受的作物大麦(Hordeum vulgare)、李(Prunus salicina)和草莓(Fragaria ananassa)等(Mills et al., 2007; Booker et al., 2009; Feng & Kobayashi, 2009; Avnery et al., 2011; Broberg et al., 2015).通过汇总406个控制实验的数据进行meta分析发现, 与活性炭过滤环境(≤26 μg·kg-1)相比, 当前环境O3浓度(平均值40 μg·kg-1)可导致主要粮食作物(小麦、水稻、大豆和马铃薯等)减产10%左右; 未来地表O3浓度升高情景下(51-71 μg·kg-1), 作物产量将进一步降低10%- 20% (Feng & Kobayashi, 2009).最近, 有关美国、法国、印度、中国和俄罗斯等小麦主产国的模型评估数据显示, 2010-2012年间高浓度O3浓度造成北半球和南半球小麦平均减产9.9%和6.2%, 抵消了因水肥管理或改良品种等措施对其增产所做的努力(Mills et al., 2018).我国这方面的研究起步较晚, 但发展迅速.从20世纪90年代末开始, 学者先后利用OTC和O3-FACE系统研究了地表O3浓度升高对我国主要粮食作物和蔬菜的影响, 涉及品种包括冬小麦、水稻、大豆、玉米、油菜、菠菜(Spinacia oleracea)和青菜(Brassica rapa)等(冯兆忠等, 2018).研究显示, 高浓度O3暴露下农作物均出现不同程度减产, 不同浓度O3对水稻、小麦、大豆等不同品种作物的减产幅度为10%-60% (贾一磊, 2016; 曹嘉晨等, 2017), 对油菜、菠菜和青菜等蔬菜生物产量的降幅为4.0%-89% (白月明等, 2002; 郭建平等, 2003). ...
Interactive effects of O3 exposure on California black oak (Quercus kelloggii Newb.) seedlings with and without N amendment
1
2008
... 除单因子O3胁迫外, 由于全球气候变化和极端气候常态化, 植物经常受到O3和其他环境因子交互作用的影响(如O3与CO2、升温、干旱或氮沉降等)(Schmadel-Hagebölling et al., 1998; Ollinger et al., 2002; Eltayeb et al., 2006; Ghimire et al., 2018).然而, 植物应对O3与其他因子交互作用的响应并不是两者或单独作用的简单叠加, 往往取决于植物种类、生长时期、对特定环境因子的敏感性、胁迫浓度及时间, 以及具体的微气候条件(列淦文和薛立, 2014).一方面, 通过诱导气孔关闭、限制叶片O3吸收(CO2浓度升高/干旱胁迫), 或提高碳同化速率、促进植物光合作用(CO2浓度升高/氮增加), CO2浓度升高、干旱胁迫和氮沉降经常被报道可增强植物抵御O3的能力, 减缓O3伤害(白月明等, 2005; Handley & Grulke, 2008; Gao et al., 2017).另一方面, 与之相反, 也有研究发现长期高浓度CO2或干旱降低气孔导度的同时, 会引起植物蒸腾速率、光合产物合成和积累的降低(McDonald et al., 2002; Pollastrini et al., 2010); 而超过植物承载阈值的高氮负荷也会对植物的光合生理活性产生负效应(Yamaguchi et al., 2010), 进而加重植物在复合环境下的碳损失、水分蒸散损失和自由基积累, 对植物产生协同伤害(Karnosky et al., 2005; 列淦文和薛立, 2014).高峰(2018)认为O3与其他环境胁迫因子出现的先后次序也会影响植物对复合作用的响应. ...
Effects of ozone on inter- and intra-species competition and photosynthesis in mesocosms of Lolium perenne and Trifolium repens
1
2009
... 植物长期过量摄取O3还可以引起物种组成、冠层结构改变, 影响生态系统种群均匀度和丰富度, 威胁生态系统多样性(Fuhrer et al., 2016; Bergmann et al., 2017), 最终影响生态系统碳、氮和水循环(McLaughlin et al., 2007; Sun et al., 2012; Grulke & Heath, 2020).基于温室或OTCs控制实验下有关草本植物的多项研究都发现, 随着O3暴露时间的延长, 对O3相对敏感的物种/种群将会被对O3相对耐受的物种/种群替代(Fuhrer et al., 1994; Hayes et al., 2009), 非禾本科杂草与禾本科杂草的比例也在发生改变(Hayes et al., 2010).有关Aspen FACE森林生态系统的研究也表明, 尽管整个森林林分结构没有受到明显影响, 但敏感型欧洲山杨树种在O3熏蒸7年后几乎从森林冠层中消失(Kubiske et al., 2007).然而, 由于种内及种间复杂的竞争关系, O3对生态系统群落结构的影响有时候并不是统一的, 高浓度O3暴露环境下O3敏感型种群首先达到较高覆盖度的情况也时有发生(Evans & Ashmore, 1992; Barbo et al., 1998).此外, 由于生态系统各营养级之间的交互作用, 高浓度O3影响植物群落多样性的同时也潜在影响植物组织、化学组成、二次代谢产物(如酚苷类化合物、植物源挥发性有机化合物等)释放, 进而改变植物种群周边与之相关的动植物、真菌、细菌和昆虫行为或活动(ICP Vegetation, 2013; Fuhrer et al., 2016).例如, O3暴露使得西黄松(Pinus ponderosa)更易受到小蠹虫的袭击(Dahlsten et al., 1997), 但O3诱导红云杉(Picea rubens)萜烯物质释放增加的同时减少了其上蚜虫幼虫的数量(Cannon, 1990); 而有关欧洲山杨的研究则发现O3暴露降低柳皮苷和特里杨苷含量, 增加了缩合单宁含量, 引起林分中蚜虫和森林天幕毛虫幼虫数量剧增, 但蚜虫捕食者或天敌(如瓢虫、草蛉和蜘蛛)的数量显著减少(Percy et al., 2002; Kopper & Lindroth, 2003). ...
Does a simulated upland grassland community respond to increasing background, peak or accumulated exposure of ozone?
1
2010
... 植物长期过量摄取O3还可以引起物种组成、冠层结构改变, 影响生态系统种群均匀度和丰富度, 威胁生态系统多样性(Fuhrer et al., 2016; Bergmann et al., 2017), 最终影响生态系统碳、氮和水循环(McLaughlin et al., 2007; Sun et al., 2012; Grulke & Heath, 2020).基于温室或OTCs控制实验下有关草本植物的多项研究都发现, 随着O3暴露时间的延长, 对O3相对敏感的物种/种群将会被对O3相对耐受的物种/种群替代(Fuhrer et al., 1994; Hayes et al., 2009), 非禾本科杂草与禾本科杂草的比例也在发生改变(Hayes et al., 2010).有关Aspen FACE森林生态系统的研究也表明, 尽管整个森林林分结构没有受到明显影响, 但敏感型欧洲山杨树种在O3熏蒸7年后几乎从森林冠层中消失(Kubiske et al., 2007).然而, 由于种内及种间复杂的竞争关系, O3对生态系统群落结构的影响有时候并不是统一的, 高浓度O3暴露环境下O3敏感型种群首先达到较高覆盖度的情况也时有发生(Evans & Ashmore, 1992; Barbo et al., 1998).此外, 由于生态系统各营养级之间的交互作用, 高浓度O3影响植物群落多样性的同时也潜在影响植物组织、化学组成、二次代谢产物(如酚苷类化合物、植物源挥发性有机化合物等)释放, 进而改变植物种群周边与之相关的动植物、真菌、细菌和昆虫行为或活动(ICP Vegetation, 2013; Fuhrer et al., 2016).例如, O3暴露使得西黄松(Pinus ponderosa)更易受到小蠹虫的袭击(Dahlsten et al., 1997), 但O3诱导红云杉(Picea rubens)萜烯物质释放增加的同时减少了其上蚜虫幼虫的数量(Cannon, 1990); 而有关欧洲山杨的研究则发现O3暴露降低柳皮苷和特里杨苷含量, 增加了缩合单宁含量, 引起林分中蚜虫和森林天幕毛虫幼虫数量剧增, 但蚜虫捕食者或天敌(如瓢虫、草蛉和蜘蛛)的数量显著减少(Percy et al., 2002; Kopper & Lindroth, 2003). ...
An open-top field chamber to assess the impact of air pollution on plants
1
1973
... 在此期间, 尽管社会经济不断发展、实验设备和研究方法不断更新, 但地表O3污染对植物及生态系统影响的研究主要还是以模拟O3浓度升高的控制实验为主.纵观发展历程, 试验设备与手段的改进和更新大致可分为三个阶段(王春乙, 1995): (1) 1970年之前, 研究主要利用封闭式静态或动态气室或室内生长箱进行模拟实验(Heagle et al., 1973). (2) 1973年之后, 美国学者Heagle等(1979)和Mandle (1973)研制开顶式气室(OTC)模拟高浓度O3对植物的损伤和影响机制, 并最终在世界范围内推广, 成为研究地表O3浓度升高对植物及生态系统影响的主流手段.(3)由于OTCs气室内外较大的环境差异, OTCs模拟实验下的熏蒸结果是否适用于大田研究一直备受争议.为解决OTCs内微气候环境带来的影响, 1986年Hendrey和Kimball (1994)提出建立自由空气中气体浓度增加(FACE)系统的想法, 并逐步用于大田实验.由于与外界大气几乎一致的气体流速、温度、光照和风速等条件(Matyssek et al., 2010), FACE系统被普遍认为是当前研究地表O3浓度升高对植物及生态系统影响最为理想的实验平台, 但由于造价昂贵, 技术要求高, 只有少数国家和实验室开展相关研究(有关上述研究方法的优缺点见表2).O3熏蒸的FACE平台仅有10个左右且多数已经停止运行(Paoletti et al., 2017), 主要包括美国威斯康星州Aspen FACE (停止运行; Karnosky et al., 2003), 美国伊利诺伊州Soybean FACE (停止运行; Morgan et al., 2004), 瑞典苏尔草地FACE (停止运行; Volk et al., 2003), 芬兰库奥皮奥FACE (停止运行; Oksanen, 2003), 德国克兰兹堡森林FACE (停止运行; Matyssek et al., 2010), 中国江都水稻(Oryza sativa)/小麦(Triticum aestivum)轮作FACE (停止运行; Tang et al., 2011), 日本札幌森林FACE (仍在运行; Watanabe et al., 2013), 意大利佛罗伦萨3D FACE (仍在运行; Paoletti et al., 2017), 以及刚刚建成的中国延庆唐家堡杨树人工林FACE.由于实验条件和FACE平台规模的限制, 有关地表O3浓度升高对生态系统水平的研究成果主要来自美国Aspen FACE和德国克兰兹堡森林FACE (Grulke & Heath, 2020). ...
Dispensing and monitoring ozone in open-top field chambers for plant-effects studies
1
1979
... 在此期间, 尽管社会经济不断发展、实验设备和研究方法不断更新, 但地表O3污染对植物及生态系统影响的研究主要还是以模拟O3浓度升高的控制实验为主.纵观发展历程, 试验设备与手段的改进和更新大致可分为三个阶段(王春乙, 1995): (1) 1970年之前, 研究主要利用封闭式静态或动态气室或室内生长箱进行模拟实验(Heagle et al., 1973). (2) 1973年之后, 美国学者Heagle等(1979)和Mandle (1973)研制开顶式气室(OTC)模拟高浓度O3对植物的损伤和影响机制, 并最终在世界范围内推广, 成为研究地表O3浓度升高对植物及生态系统影响的主流手段.(3)由于OTCs气室内外较大的环境差异, OTCs模拟实验下的熏蒸结果是否适用于大田研究一直备受争议.为解决OTCs内微气候环境带来的影响, 1986年Hendrey和Kimball (1994)提出建立自由空气中气体浓度增加(FACE)系统的想法, 并逐步用于大田实验.由于与外界大气几乎一致的气体流速、温度、光照和风速等条件(Matyssek et al., 2010), FACE系统被普遍认为是当前研究地表O3浓度升高对植物及生态系统影响最为理想的实验平台, 但由于造价昂贵, 技术要求高, 只有少数国家和实验室开展相关研究(有关上述研究方法的优缺点见表2).O3熏蒸的FACE平台仅有10个左右且多数已经停止运行(Paoletti et al., 2017), 主要包括美国威斯康星州Aspen FACE (停止运行; Karnosky et al., 2003), 美国伊利诺伊州Soybean FACE (停止运行; Morgan et al., 2004), 瑞典苏尔草地FACE (停止运行; Volk et al., 2003), 芬兰库奥皮奥FACE (停止运行; Oksanen, 2003), 德国克兰兹堡森林FACE (停止运行; Matyssek et al., 2010), 中国江都水稻(Oryza sativa)/小麦(Triticum aestivum)轮作FACE (停止运行; Tang et al., 2011), 日本札幌森林FACE (仍在运行; Watanabe et al., 2013), 意大利佛罗伦萨3D FACE (仍在运行; Paoletti et al., 2017), 以及刚刚建成的中国延庆唐家堡杨树人工林FACE.由于实验条件和FACE平台规模的限制, 有关地表O3浓度升高对生态系统水平的研究成果主要来自美国Aspen FACE和德国克兰兹堡森林FACE (Grulke & Heath, 2020). ...
The FACE program
1
1994
... 在此期间, 尽管社会经济不断发展、实验设备和研究方法不断更新, 但地表O3污染对植物及生态系统影响的研究主要还是以模拟O3浓度升高的控制实验为主.纵观发展历程, 试验设备与手段的改进和更新大致可分为三个阶段(王春乙, 1995): (1) 1970年之前, 研究主要利用封闭式静态或动态气室或室内生长箱进行模拟实验(Heagle et al., 1973). (2) 1973年之后, 美国学者Heagle等(1979)和Mandle (1973)研制开顶式气室(OTC)模拟高浓度O3对植物的损伤和影响机制, 并最终在世界范围内推广, 成为研究地表O3浓度升高对植物及生态系统影响的主流手段.(3)由于OTCs气室内外较大的环境差异, OTCs模拟实验下的熏蒸结果是否适用于大田研究一直备受争议.为解决OTCs内微气候环境带来的影响, 1986年Hendrey和Kimball (1994)提出建立自由空气中气体浓度增加(FACE)系统的想法, 并逐步用于大田实验.由于与外界大气几乎一致的气体流速、温度、光照和风速等条件(Matyssek et al., 2010), FACE系统被普遍认为是当前研究地表O3浓度升高对植物及生态系统影响最为理想的实验平台, 但由于造价昂贵, 技术要求高, 只有少数国家和实验室开展相关研究(有关上述研究方法的优缺点见表2).O3熏蒸的FACE平台仅有10个左右且多数已经停止运行(Paoletti et al., 2017), 主要包括美国威斯康星州Aspen FACE (停止运行; Karnosky et al., 2003), 美国伊利诺伊州Soybean FACE (停止运行; Morgan et al., 2004), 瑞典苏尔草地FACE (停止运行; Volk et al., 2003), 芬兰库奥皮奥FACE (停止运行; Oksanen, 2003), 德国克兰兹堡森林FACE (停止运行; Matyssek et al., 2010), 中国江都水稻(Oryza sativa)/小麦(Triticum aestivum)轮作FACE (停止运行; Tang et al., 2011), 日本札幌森林FACE (仍在运行; Watanabe et al., 2013), 意大利佛罗伦萨3D FACE (仍在运行; Paoletti et al., 2017), 以及刚刚建成的中国延庆唐家堡杨树人工林FACE.由于实验条件和FACE平台规模的限制, 有关地表O3浓度升高对生态系统水平的研究成果主要来自美国Aspen FACE和德国克兰兹堡森林FACE (Grulke & Heath, 2020). ...
Nitrogen management is essential to prevent tropical oil palm plantations from causing ground-level ozone pollution
1
2009
... (6)探究地表O3污染的生态控制措施: 欧洲和北美等发达国家治理大气污染的经验告诉我们, 严格的O3前体物NOx和挥发性有机化合物(VOCs)减排措施是降低地表O3浓度的有效手段(Feng et al., 2015b).然而, 以完全牺牲经济发展为代价的环境保护措施一般并不被发展中国家所接受.事实上, 自然植被与地表O3存在复杂的交互作用.一方面, 植物释放的VOCs有助于地表O3的形成; 另一方面, 植物在进行光合作用的同时也可以通过气孔、非气孔途径吸收或沉降大气O3 (Paoletti, 2009).考虑到植被VOCs释放和O3移除能力都具有树种特异性, 因此, 在一定条件下通过合理的树种筛选达到减少大气O3污染的目的将是完全可行的(Sicard et al., 2018).另外, 除植物自身遗传基因之外, 植物对O3胁迫的响应受土壤水分和营养元素如氮可利用性的影响(列淦文和薛立, 2014; 高峰, 2018).例如古巴农民通过提前1-2天灌溉的方式预防夏季高浓度O3对莴苣和烟草叶面的损伤(ICP Vegetation, 2011); 马来西亚研究者通过氮肥管理, 减轻O3污染对热带Elaeis guineensis产量和质量的伤害(Hewitt et al., 2009).因此通过合理的田间水肥管理措施(如改变灌溉模式、施肥剂量和方式)调控植物生长, 增强植物对O3的抗性, 进而减轻O3污染的生态效应是未来大气O3污染防治研究中值得思考的问题. ...
The rapid inhibition of root respiration after exposure of bean (Phaseolus vulgaris L.) plants to ozone
1
1981
... (2)加强地表O3污染对地下生态过程的影响研究: 地下部分是生态系统水分供给、养分循环、碳分配的核心环节, 探究地下生态系统对地表O3浓度升高的响应对于准确理解地表O3浓度升高对整个生态系统的影响具有重要意义.事实上, 高浓度O3对地下部分的干扰比地上部分更明显且更早出现(Hofstra et al., 1981).然而, 目前有关O3对地下生态过程动态变化过程的研究还十分缺乏, 其响应机制尚不明确.已有的研究多集中在地表O3浓度升高对地下碳分配、根系生长和生物量的影响上, 少部分涉及凋落物、根系分泌物、土壤呼吸、微生物活性和结构等(Agathokleous et al., 2016), 而地表O3浓度升高对土壤微食物网的影响及其反馈几乎都没有涉及(Grulke & Heath, 2020), 更无法将土壤微生物过程(如微生物活性、生物量、群落组成、多样性和功能)与土壤关键生物地球化学循环过程, 以及与植物地上生态过程关联起来(Andersen, 2003; 冯兆忠等, 2018).未来的研究应加强O3对地下生态过程的长期定位研究, 特别是O3对地下碳分配动态、土壤微生物动态及根际氮循环的研究, 量化地下生态过程对O3响应的程度及其对生态系统长期稳定的反馈. ...
Ozone-induced stomatal sluggishness develops progressively in Siebold’s beech (Fagus crenata)
1
2012
... 通常认为, 植物通过气孔防御和非气孔防御对O3胁迫做出响应.气孔作为叶片与外界气体交换的“门户”, 是O3进入植物体内的主要通道, 控制着植物蒸腾、呼吸和光合等重要生理过程.正常光热条件下, 气孔处于打开状态, 但高浓度O3可以直接影响调节气孔开张的保卫细胞上钾离子浓度, 改变细胞膨压, 诱导气孔关闭, 引发避逆性的防御响应(Wieser et al., 2002).因此, 气孔闭合被认为是植物抵御O3胁迫的第一道防线(Ainsworth, 2017).然而, 气孔对O3的响应(关闭)也并不是始终如一的.研究发现随着O3暴露时间的延长, O3诱导的气孔关闭会出现滞后甚至失灵的现象(Hoshika et al., 2012), 从而导致大量O3进入细胞间隙, 气孔防御机制 失效. ...
Concentration- and flux-based ozone dose-response relationships for five poplar clones grown in North China
1
2015
... 基于OTCs和O3-FACE平台下单个站点的控制实验可探究O3对植物个体、群落或生态系统的影响, 但将站点或样地尺度的研究外推到区域或全球, 评估O3污染区域风险的研究主要通过响应关系和模型.目前, 其研究领域主要集中在地表O3污染对农作物产量和森林生产力等方面.早在1994年, Chameides等(1994)就利用经济模型初步预测O3浓度升高可能导致2025年全球30%-75%的作物面临产量降低的风险.基于O3前体物排放、化学反应传输模型和M12、AOT40指标的评估显示, 2000年全球O3污染引起全球小麦、玉米和大豆产量平均降低2.2%-15%, 每年因上述作物产量降低所导致的经济损失达110-180亿美元(Avnery et al., 2011).最近, 基于PODY评估指标的研究则表明, O3对小麦造成的产量损失北半球(9.9%)明显高于南半球(6.2%), 平均产量损失最高的地区主要位于北半球暖温带湿润地区(12.5%)、热带湿热(12.5%)和热带潮湿气候区(17.2%)(Mills et al., 2018).在区域尺度上, 基于1990年O3浓度数据, Wang和Mauzerall (2004)利用M7和M12指标评估了O3对中国、日本和韩国主要农作物产量的影响, 结果显示O3污染造成小麦、玉米、棉花减产1%-9%, 大豆减产23%-27%, 造成2.4-35亿美元的经济损失, 在维持现有的污染物排放、管理措施和农业生产技术不变的情况下, 预计到2020年小麦、玉米、棉花减产幅度为2%-15%, 大豆为28%-35%.最近, 利用AOT40指标的研究显示, 2014年当前O3浓度导致中国农作物产量损失7 840万t, 小麦、水稻和玉米相对减产量分别为8.5%-14%、3.9%-15%和2.2%-5.5% (Lin et al., 2018).而只聚焦于长江三角洲地区的研究则发现, 2015年区域O3污染分别造成小麦和水稻减产9.9%-36.1%和7.3%- 23.9% (Zhao et al., 2018).虽然国内也有少量研究建立了PODY与个别树种或作物相对生物量或相对产量之间的通量响应关系(Feng et al., 2012; Hu et al., 2015; Gao et al., 2017), 但并没有研究利用PODY指标评估区域O3污染对农作物产量损失的影响. ...
Ozone pollution: a hidden threat to food security. Programme Coordination Centre for the ICP Vegetation//Mills G, Harmens H. Programme Coordination Centre for the ICP Vegetation
1
2011
... (6)探究地表O3污染的生态控制措施: 欧洲和北美等发达国家治理大气污染的经验告诉我们, 严格的O3前体物NOx和挥发性有机化合物(VOCs)减排措施是降低地表O3浓度的有效手段(Feng et al., 2015b).然而, 以完全牺牲经济发展为代价的环境保护措施一般并不被发展中国家所接受.事实上, 自然植被与地表O3存在复杂的交互作用.一方面, 植物释放的VOCs有助于地表O3的形成; 另一方面, 植物在进行光合作用的同时也可以通过气孔、非气孔途径吸收或沉降大气O3 (Paoletti, 2009).考虑到植被VOCs释放和O3移除能力都具有树种特异性, 因此, 在一定条件下通过合理的树种筛选达到减少大气O3污染的目的将是完全可行的(Sicard et al., 2018).另外, 除植物自身遗传基因之外, 植物对O3胁迫的响应受土壤水分和营养元素如氮可利用性的影响(列淦文和薛立, 2014; 高峰, 2018).例如古巴农民通过提前1-2天灌溉的方式预防夏季高浓度O3对莴苣和烟草叶面的损伤(ICP Vegetation, 2011); 马来西亚研究者通过氮肥管理, 减轻O3污染对热带Elaeis guineensis产量和质量的伤害(Hewitt et al., 2009).因此通过合理的田间水肥管理措施(如改变灌溉模式、施肥剂量和方式)调控植物生长, 增强植物对O3的抗性, 进而减轻O3污染的生态效应是未来大气O3污染防治研究中值得思考的问题. ...
Ozone pollution: impacts on ecosystem services and biodiversity. Programme Coordination Centre for the ICP Vegetation//Mills G, Wagg S, Harmens H. Programme Coordination Centre for the ICP Vegetation
2
2013
... 植物长期过量摄取O3还可以引起物种组成、冠层结构改变, 影响生态系统种群均匀度和丰富度, 威胁生态系统多样性(Fuhrer et al., 2016; Bergmann et al., 2017), 最终影响生态系统碳、氮和水循环(McLaughlin et al., 2007; Sun et al., 2012; Grulke & Heath, 2020).基于温室或OTCs控制实验下有关草本植物的多项研究都发现, 随着O3暴露时间的延长, 对O3相对敏感的物种/种群将会被对O3相对耐受的物种/种群替代(Fuhrer et al., 1994; Hayes et al., 2009), 非禾本科杂草与禾本科杂草的比例也在发生改变(Hayes et al., 2010).有关Aspen FACE森林生态系统的研究也表明, 尽管整个森林林分结构没有受到明显影响, 但敏感型欧洲山杨树种在O3熏蒸7年后几乎从森林冠层中消失(Kubiske et al., 2007).然而, 由于种内及种间复杂的竞争关系, O3对生态系统群落结构的影响有时候并不是统一的, 高浓度O3暴露环境下O3敏感型种群首先达到较高覆盖度的情况也时有发生(Evans & Ashmore, 1992; Barbo et al., 1998).此外, 由于生态系统各营养级之间的交互作用, 高浓度O3影响植物群落多样性的同时也潜在影响植物组织、化学组成、二次代谢产物(如酚苷类化合物、植物源挥发性有机化合物等)释放, 进而改变植物种群周边与之相关的动植物、真菌、细菌和昆虫行为或活动(ICP Vegetation, 2013; Fuhrer et al., 2016).例如, O3暴露使得西黄松(Pinus ponderosa)更易受到小蠹虫的袭击(Dahlsten et al., 1997), 但O3诱导红云杉(Picea rubens)萜烯物质释放增加的同时减少了其上蚜虫幼虫的数量(Cannon, 1990); 而有关欧洲山杨的研究则发现O3暴露降低柳皮苷和特里杨苷含量, 增加了缩合单宁含量, 引起林分中蚜虫和森林天幕毛虫幼虫数量剧增, 但蚜虫捕食者或天敌(如瓢虫、草蛉和蜘蛛)的数量显著减少(Percy et al., 2002; Kopper & Lindroth, 2003). ...
... 此外, 虽然地下生态系统并不是直接暴露在O3环境下, 但大量证据表明O3对地上生态系统的影响将会转移至地下(Andersen, 2003; Lindroth, 2010).如高浓度O3环境下, 植物分配到根的生物量将会大大减少, 基于100多组实验数据的meta分析研究表明O3暴露导致生物量分配到地下的量平均减少5.6% (Grantz et al., 2006).另外, 凋落物分解是生态系统营养循环的关键过程.通过影响凋落物的数量、质量和分解速率, 高浓度地表O3会间接改变土壤微生物群落活性、组成和结构, 土壤化学属性及土壤营养循环(Larson et al., 2002; Fuhrer et al., 2016).然而, 比较遗憾的是, 目前有关O3对地下生态过程影响的研究仅集中在农田生态系统, 仅有少部分研究涉及森林土壤(主要来自Aspen FACE)或草地生态系统(ICP Vegetation, 2013).而且, 由于凋落物种类、化学成分、分解者种群、土壤和大气环境条件的不同, 高浓度O3对凋落物分解动态的影响还存在很大争议(Gessner et al., 2010; Fuhrer et al., 2016; Grulke & Heath, 2020).例如, 研究发现O3对欧洲白桦(Betula pendula)叶片凋落物质量和损失率有明显影响(Kasurinen et al., 2006, 2007), 但不影响欧洲山毛榉凋落物的分解速率(Schloter et al., 2005).有关地表O3浓度升高对地下生态系统结构和功能的影响还有待进一步探究. ...
臭氧胁迫对不同小麦品种产量、品质和抗倒性的影响
2
2016
... O3不仅抑制植物生长, 还导致农作物大量减产(Fuhrer, 2009; Ainsworth et al., 2012; Feng et al., 2015a; Singh & Agrawal, 2017).有关O3对作物产量的影响已经在北美、欧洲和亚洲多地的几十种作物上有所研究报道, 包括对O3敏感的小麦、西瓜(Citrullus lanatus)、豆类、棉花(Gossypium hirsutum)、萝卜(Raphanus sativus)、番茄(Lycopersicon esclentum)、洋葱(Allium cepa)、大豆(Glycine max)和莴苣(Lactuca sativa), 对O3适度敏感的甜菜(Beta vulgaris)、马铃薯(Solanum tuberosum)、油菜(Brassica napus)、烟草(Nicotiana tabacum)、水稻、玉米(Zea mays)、葡萄和西兰花(Brassica oleracea), 和对O3耐受的作物大麦(Hordeum vulgare)、李(Prunus salicina)和草莓(Fragaria ananassa)等(Mills et al., 2007; Booker et al., 2009; Feng & Kobayashi, 2009; Avnery et al., 2011; Broberg et al., 2015).通过汇总406个控制实验的数据进行meta分析发现, 与活性炭过滤环境(≤26 μg·kg-1)相比, 当前环境O3浓度(平均值40 μg·kg-1)可导致主要粮食作物(小麦、水稻、大豆和马铃薯等)减产10%左右; 未来地表O3浓度升高情景下(51-71 μg·kg-1), 作物产量将进一步降低10%- 20% (Feng & Kobayashi, 2009).最近, 有关美国、法国、印度、中国和俄罗斯等小麦主产国的模型评估数据显示, 2010-2012年间高浓度O3浓度造成北半球和南半球小麦平均减产9.9%和6.2%, 抵消了因水肥管理或改良品种等措施对其增产所做的努力(Mills et al., 2018).我国这方面的研究起步较晚, 但发展迅速.从20世纪90年代末开始, 学者先后利用OTC和O3-FACE系统研究了地表O3浓度升高对我国主要粮食作物和蔬菜的影响, 涉及品种包括冬小麦、水稻、大豆、玉米、油菜、菠菜(Spinacia oleracea)和青菜(Brassica rapa)等(冯兆忠等, 2018).研究显示, 高浓度O3暴露下农作物均出现不同程度减产, 不同浓度O3对水稻、小麦、大豆等不同品种作物的减产幅度为10%-60% (贾一磊, 2016; 曹嘉晨等, 2017), 对油菜、菠菜和青菜等蔬菜生物产量的降幅为4.0%-89% (白月明等, 2002; 郭建平等, 2003). ...
... 相对于农作物产量, O3对粮食品质影响的研究集中在几种主要农作物(如小麦、水稻和大豆)(冯兆忠等, 2018).基于北美、欧洲和亚洲已有的O3对小麦影响的meta分析发现, O3显著降低小麦淀粉浓度, 但显著增加蛋白质和一些营养元素浓度(如钾、镁、钙、磷、锌、锰和铜)(Broberg et al., 2015).印度北部热带地区有关小麦的研究则显示, O3显著降低小麦籽粒糖含量, 但没有改变籽粒蛋白质含量(Tomer et al., 2015).有关水稻的研究还发现, O3增加粳稻直链淀粉含量, 但对籼稻直链淀粉含量无显著影响(Sawada et al., 2016).国内研究则表明, 尽管高浓度O3显著降低冬小麦面粉中支链淀粉和淀粉含量, 但是增加直链淀粉含量(Zhang et al., 2013).O3显著降低水稻稻米直链淀粉浓度、分解率, 增加黏度、起糊温度、蛋白质和微量矿物质浓度, 但蛋白质总量没有改变(Wang et al., 2012).此外, 研究还发现O3熏蒸显著降低了小麦籽粒出粉率和硬度, 但增加冬小麦面粉峰值黏度、低谷黏度、最终黏度和反弹值(贾一磊, 2016). ...
臭氧胁迫对不同小麦品种产量、品质和抗倒性的影响
2
2016
... O3不仅抑制植物生长, 还导致农作物大量减产(Fuhrer, 2009; Ainsworth et al., 2012; Feng et al., 2015a; Singh & Agrawal, 2017).有关O3对作物产量的影响已经在北美、欧洲和亚洲多地的几十种作物上有所研究报道, 包括对O3敏感的小麦、西瓜(Citrullus lanatus)、豆类、棉花(Gossypium hirsutum)、萝卜(Raphanus sativus)、番茄(Lycopersicon esclentum)、洋葱(Allium cepa)、大豆(Glycine max)和莴苣(Lactuca sativa), 对O3适度敏感的甜菜(Beta vulgaris)、马铃薯(Solanum tuberosum)、油菜(Brassica napus)、烟草(Nicotiana tabacum)、水稻、玉米(Zea mays)、葡萄和西兰花(Brassica oleracea), 和对O3耐受的作物大麦(Hordeum vulgare)、李(Prunus salicina)和草莓(Fragaria ananassa)等(Mills et al., 2007; Booker et al., 2009; Feng & Kobayashi, 2009; Avnery et al., 2011; Broberg et al., 2015).通过汇总406个控制实验的数据进行meta分析发现, 与活性炭过滤环境(≤26 μg·kg-1)相比, 当前环境O3浓度(平均值40 μg·kg-1)可导致主要粮食作物(小麦、水稻、大豆和马铃薯等)减产10%左右; 未来地表O3浓度升高情景下(51-71 μg·kg-1), 作物产量将进一步降低10%- 20% (Feng & Kobayashi, 2009).最近, 有关美国、法国、印度、中国和俄罗斯等小麦主产国的模型评估数据显示, 2010-2012年间高浓度O3浓度造成北半球和南半球小麦平均减产9.9%和6.2%, 抵消了因水肥管理或改良品种等措施对其增产所做的努力(Mills et al., 2018).我国这方面的研究起步较晚, 但发展迅速.从20世纪90年代末开始, 学者先后利用OTC和O3-FACE系统研究了地表O3浓度升高对我国主要粮食作物和蔬菜的影响, 涉及品种包括冬小麦、水稻、大豆、玉米、油菜、菠菜(Spinacia oleracea)和青菜(Brassica rapa)等(冯兆忠等, 2018).研究显示, 高浓度O3暴露下农作物均出现不同程度减产, 不同浓度O3对水稻、小麦、大豆等不同品种作物的减产幅度为10%-60% (贾一磊, 2016; 曹嘉晨等, 2017), 对油菜、菠菜和青菜等蔬菜生物产量的降幅为4.0%-89% (白月明等, 2002; 郭建平等, 2003). ...
... 相对于农作物产量, O3对粮食品质影响的研究集中在几种主要农作物(如小麦、水稻和大豆)(冯兆忠等, 2018).基于北美、欧洲和亚洲已有的O3对小麦影响的meta分析发现, O3显著降低小麦淀粉浓度, 但显著增加蛋白质和一些营养元素浓度(如钾、镁、钙、磷、锌、锰和铜)(Broberg et al., 2015).印度北部热带地区有关小麦的研究则显示, O3显著降低小麦籽粒糖含量, 但没有改变籽粒蛋白质含量(Tomer et al., 2015).有关水稻的研究还发现, O3增加粳稻直链淀粉含量, 但对籼稻直链淀粉含量无显著影响(Sawada et al., 2016).国内研究则表明, 尽管高浓度O3显著降低冬小麦面粉中支链淀粉和淀粉含量, 但是增加直链淀粉含量(Zhang et al., 2013).O3显著降低水稻稻米直链淀粉浓度、分解率, 增加黏度、起糊温度、蛋白质和微量矿物质浓度, 但蛋白质总量没有改变(Wang et al., 2012).此外, 研究还发现O3熏蒸显著降低了小麦籽粒出粉率和硬度, 但增加冬小麦面粉峰值黏度、低谷黏度、最终黏度和反弹值(贾一磊, 2016). ...
Signalling and cell death in ozone-exposed plants
1
2005
... 气孔防御机制失效后, 进入细胞的O3将诱导叶片内部抗氧化系统启动解毒和修复的非气孔防御响应(冯兆忠等, 2018).非气孔防御响应主要是指“质外体-共质体系统”内一些抗氧化组分对O3解毒和修复的防御过程.具体来说, O3进入植物叶片的细胞间隙后会溶解在细胞壁的结合水里, 经过一系列的化学反应形成活性氧分子(ROS), ROS能够破坏细胞壁并与细胞膜上的不饱和脂肪酸发生反应, 从而对叶片造成损伤(Tausz et al., 2007; Pellegrini et al., 2018).植物质外体的抗氧化物质如还原性抗坏血酸(ASC)、抗氧化酶(如超氧化物歧化酶)可与ROS反应进行初级解毒, 成为抵御O3的第二道防线(Kangasjärvi et al., 2005; Dai et al., 2019), 而剩余的ROS会经过质膜到达共质体, 此时共质体内抗氧化成分如过氧化物酶(POD)、过氧化氢酶(CAT)、抗坏血酸过氧化物酶(APX)及抗氧化剂(如酚类化合物-谷胱甘肽(GSH)、脯氨酸和多胺等)等抗氧化酶类物质被诱导作为次级解毒响应机制参与诸如“抗坏血酸-谷胱甘肽(ASA-GSH)循环”等氧化还原过程, 进而对ROS进行进一步解毒与消除(Luwe, 1996).气孔闭合除了会减少O3的吸收, 也会抑制CO2吸收和水汽交换, 而非气孔防御响应中的抗氧化系统解毒过程也会消耗大量的能量并对光合反应系统造成不同程度的损伤, 降低羧化速率和电子传递速率, 进而抑制光合和蒸腾作用, 损害植物生长(Gao et al., 2016).然而, 最新研究表明持续整个生长季的长期O3暴露会影响植物叶片的气孔密度、开度和大小等表观属性(李品等, 2018), 而O3从气孔进入到细胞间隙后, 也可以通过影响植物的叶肉导度降低植物的光合能力(Xu et al., 2019).此外, 有关29种落叶和常绿树种应对O3胁迫的研究还发现不同木本植物对O3响应的差异主要来自植物功能叶片比叶质量的大小(即叶片比叶质量越大的树种对O3越不敏感), 与气孔导度无明显相关性(Li et al., 2016). ...
Scaling ozone responses of forest trees to the ecosystem level in a changing climate
2
2005
... 除单因子O3胁迫外, 由于全球气候变化和极端气候常态化, 植物经常受到O3和其他环境因子交互作用的影响(如O3与CO2、升温、干旱或氮沉降等)(Schmadel-Hagebölling et al., 1998; Ollinger et al., 2002; Eltayeb et al., 2006; Ghimire et al., 2018).然而, 植物应对O3与其他因子交互作用的响应并不是两者或单独作用的简单叠加, 往往取决于植物种类、生长时期、对特定环境因子的敏感性、胁迫浓度及时间, 以及具体的微气候条件(列淦文和薛立, 2014).一方面, 通过诱导气孔关闭、限制叶片O3吸收(CO2浓度升高/干旱胁迫), 或提高碳同化速率、促进植物光合作用(CO2浓度升高/氮增加), CO2浓度升高、干旱胁迫和氮沉降经常被报道可增强植物抵御O3的能力, 减缓O3伤害(白月明等, 2005; Handley & Grulke, 2008; Gao et al., 2017).另一方面, 与之相反, 也有研究发现长期高浓度CO2或干旱降低气孔导度的同时, 会引起植物蒸腾速率、光合产物合成和积累的降低(McDonald et al., 2002; Pollastrini et al., 2010); 而超过植物承载阈值的高氮负荷也会对植物的光合生理活性产生负效应(Yamaguchi et al., 2010), 进而加重植物在复合环境下的碳损失、水分蒸散损失和自由基积累, 对植物产生协同伤害(Karnosky et al., 2005; 列淦文和薛立, 2014).高峰(2018)认为O3与其他环境胁迫因子出现的先后次序也会影响植物对复合作用的响应. ...
... 有关自然环境下O3浓度升高对陆地生态系统影响的研究较为稀缺.但是现有开放式O3浓度增强系统(O3-FACE)观测平台下的长期研究也有着相似结果, Aspen FACE实验发现, O3浓度升高3-4年后, 欧洲山杨(Populus tremuloides)纯林、欧洲山杨-纸皮桦(Betula papyrifera)和欧洲山杨-糖枫(Acer saccharum)混交林的总生物量分别降低23%、13%和14% (Karnosky et al., 2003, 2005); 虽然在最初的7年内生物量的分配格局并未受O3影响(King et al., 2005), 但在实验11年后, 敏感型欧洲山杨几乎从森林冠层中消失(Kubiske et al., 2007).德国克兰兹堡郁闭冠层的成熟挪威云杉(Picea abies)和欧洲山毛榉(Fagus sylvatica)混交林(60年)对O3浓度升高长期响应的研究也表明, 8年O3暴露导致欧洲山毛榉的树干生产力降低了44% (Pretzsch et al., 2010).目前, 国内还没有研究涉及O3-FACE平台下O3浓度升高对森林生态系统碳汇能力的影响. ...
Tropospheric O3 moderates responses of temperate hardwood forests to elevated CO2: a synthesis of molecular to ecosystem results from the Aspen FACE project
2
2003
... 在此期间, 尽管社会经济不断发展、实验设备和研究方法不断更新, 但地表O3污染对植物及生态系统影响的研究主要还是以模拟O3浓度升高的控制实验为主.纵观发展历程, 试验设备与手段的改进和更新大致可分为三个阶段(王春乙, 1995): (1) 1970年之前, 研究主要利用封闭式静态或动态气室或室内生长箱进行模拟实验(Heagle et al., 1973). (2) 1973年之后, 美国学者Heagle等(1979)和Mandle (1973)研制开顶式气室(OTC)模拟高浓度O3对植物的损伤和影响机制, 并最终在世界范围内推广, 成为研究地表O3浓度升高对植物及生态系统影响的主流手段.(3)由于OTCs气室内外较大的环境差异, OTCs模拟实验下的熏蒸结果是否适用于大田研究一直备受争议.为解决OTCs内微气候环境带来的影响, 1986年Hendrey和Kimball (1994)提出建立自由空气中气体浓度增加(FACE)系统的想法, 并逐步用于大田实验.由于与外界大气几乎一致的气体流速、温度、光照和风速等条件(Matyssek et al., 2010), FACE系统被普遍认为是当前研究地表O3浓度升高对植物及生态系统影响最为理想的实验平台, 但由于造价昂贵, 技术要求高, 只有少数国家和实验室开展相关研究(有关上述研究方法的优缺点见表2).O3熏蒸的FACE平台仅有10个左右且多数已经停止运行(Paoletti et al., 2017), 主要包括美国威斯康星州Aspen FACE (停止运行; Karnosky et al., 2003), 美国伊利诺伊州Soybean FACE (停止运行; Morgan et al., 2004), 瑞典苏尔草地FACE (停止运行; Volk et al., 2003), 芬兰库奥皮奥FACE (停止运行; Oksanen, 2003), 德国克兰兹堡森林FACE (停止运行; Matyssek et al., 2010), 中国江都水稻(Oryza sativa)/小麦(Triticum aestivum)轮作FACE (停止运行; Tang et al., 2011), 日本札幌森林FACE (仍在运行; Watanabe et al., 2013), 意大利佛罗伦萨3D FACE (仍在运行; Paoletti et al., 2017), 以及刚刚建成的中国延庆唐家堡杨树人工林FACE.由于实验条件和FACE平台规模的限制, 有关地表O3浓度升高对生态系统水平的研究成果主要来自美国Aspen FACE和德国克兰兹堡森林FACE (Grulke & Heath, 2020). ...
... 有关自然环境下O3浓度升高对陆地生态系统影响的研究较为稀缺.但是现有开放式O3浓度增强系统(O3-FACE)观测平台下的长期研究也有着相似结果, Aspen FACE实验发现, O3浓度升高3-4年后, 欧洲山杨(Populus tremuloides)纯林、欧洲山杨-纸皮桦(Betula papyrifera)和欧洲山杨-糖枫(Acer saccharum)混交林的总生物量分别降低23%、13%和14% (Karnosky et al., 2003, 2005); 虽然在最初的7年内生物量的分配格局并未受O3影响(King et al., 2005), 但在实验11年后, 敏感型欧洲山杨几乎从森林冠层中消失(Kubiske et al., 2007).德国克兰兹堡郁闭冠层的成熟挪威云杉(Picea abies)和欧洲山毛榉(Fagus sylvatica)混交林(60年)对O3浓度升高长期响应的研究也表明, 8年O3暴露导致欧洲山毛榉的树干生产力降低了44% (Pretzsch et al., 2010).目前, 国内还没有研究涉及O3-FACE平台下O3浓度升高对森林生态系统碳汇能力的影响. ...
Leaf litter under changing climate: Will increasing levels of CO2 and O3 affect decomposition and nutrient cycling processes?
1
2007
... 此外, 虽然地下生态系统并不是直接暴露在O3环境下, 但大量证据表明O3对地上生态系统的影响将会转移至地下(Andersen, 2003; Lindroth, 2010).如高浓度O3环境下, 植物分配到根的生物量将会大大减少, 基于100多组实验数据的meta分析研究表明O3暴露导致生物量分配到地下的量平均减少5.6% (Grantz et al., 2006).另外, 凋落物分解是生态系统营养循环的关键过程.通过影响凋落物的数量、质量和分解速率, 高浓度地表O3会间接改变土壤微生物群落活性、组成和结构, 土壤化学属性及土壤营养循环(Larson et al., 2002; Fuhrer et al., 2016).然而, 比较遗憾的是, 目前有关O3对地下生态过程影响的研究仅集中在农田生态系统, 仅有少部分研究涉及森林土壤(主要来自Aspen FACE)或草地生态系统(ICP Vegetation, 2013).而且, 由于凋落物种类、化学成分、分解者种群、土壤和大气环境条件的不同, 高浓度O3对凋落物分解动态的影响还存在很大争议(Gessner et al., 2010; Fuhrer et al., 2016; Grulke & Heath, 2020).例如, 研究发现O3对欧洲白桦(Betula pendula)叶片凋落物质量和损失率有明显影响(Kasurinen et al., 2006, 2007), 但不影响欧洲山毛榉凋落物的分解速率(Schloter et al., 2005).有关地表O3浓度升高对地下生态系统结构和功能的影响还有待进一步探究. ...
Chemical composition and decomposition of silver birch leaf litter produced under elevated CO2 and O3
1
2006
... 此外, 虽然地下生态系统并不是直接暴露在O3环境下, 但大量证据表明O3对地上生态系统的影响将会转移至地下(Andersen, 2003; Lindroth, 2010).如高浓度O3环境下, 植物分配到根的生物量将会大大减少, 基于100多组实验数据的meta分析研究表明O3暴露导致生物量分配到地下的量平均减少5.6% (Grantz et al., 2006).另外, 凋落物分解是生态系统营养循环的关键过程.通过影响凋落物的数量、质量和分解速率, 高浓度地表O3会间接改变土壤微生物群落活性、组成和结构, 土壤化学属性及土壤营养循环(Larson et al., 2002; Fuhrer et al., 2016).然而, 比较遗憾的是, 目前有关O3对地下生态过程影响的研究仅集中在农田生态系统, 仅有少部分研究涉及森林土壤(主要来自Aspen FACE)或草地生态系统(ICP Vegetation, 2013).而且, 由于凋落物种类、化学成分、分解者种群、土壤和大气环境条件的不同, 高浓度O3对凋落物分解动态的影响还存在很大争议(Gessner et al., 2010; Fuhrer et al., 2016; Grulke & Heath, 2020).例如, 研究发现O3对欧洲白桦(Betula pendula)叶片凋落物质量和损失率有明显影响(Kasurinen et al., 2006, 2007), 但不影响欧洲山毛榉凋落物的分解速率(Schloter et al., 2005).有关地表O3浓度升高对地下生态系统结构和功能的影响还有待进一步探究. ...
Tropospheric O3 compromises net primary production in young stands of trembling aspen, paper birch and sugar maple in response to elevated atmospheric CO2
1
2005
... 有关自然环境下O3浓度升高对陆地生态系统影响的研究较为稀缺.但是现有开放式O3浓度增强系统(O3-FACE)观测平台下的长期研究也有着相似结果, Aspen FACE实验发现, O3浓度升高3-4年后, 欧洲山杨(Populus tremuloides)纯林、欧洲山杨-纸皮桦(Betula papyrifera)和欧洲山杨-糖枫(Acer saccharum)混交林的总生物量分别降低23%、13%和14% (Karnosky et al., 2003, 2005); 虽然在最初的7年内生物量的分配格局并未受O3影响(King et al., 2005), 但在实验11年后, 敏感型欧洲山杨几乎从森林冠层中消失(Kubiske et al., 2007).德国克兰兹堡郁闭冠层的成熟挪威云杉(Picea abies)和欧洲山毛榉(Fagus sylvatica)混交林(60年)对O3浓度升高长期响应的研究也表明, 8年O3暴露导致欧洲山毛榉的树干生产力降低了44% (Pretzsch et al., 2010).目前, 国内还没有研究涉及O3-FACE平台下O3浓度升高对森林生态系统碳汇能力的影响. ...
Effects of elevated carbon dioxide and ozone on the phytochemistry of aspen and performance of an herbivore
1
2003
... 植物长期过量摄取O3还可以引起物种组成、冠层结构改变, 影响生态系统种群均匀度和丰富度, 威胁生态系统多样性(Fuhrer et al., 2016; Bergmann et al., 2017), 最终影响生态系统碳、氮和水循环(McLaughlin et al., 2007; Sun et al., 2012; Grulke & Heath, 2020).基于温室或OTCs控制实验下有关草本植物的多项研究都发现, 随着O3暴露时间的延长, 对O3相对敏感的物种/种群将会被对O3相对耐受的物种/种群替代(Fuhrer et al., 1994; Hayes et al., 2009), 非禾本科杂草与禾本科杂草的比例也在发生改变(Hayes et al., 2010).有关Aspen FACE森林生态系统的研究也表明, 尽管整个森林林分结构没有受到明显影响, 但敏感型欧洲山杨树种在O3熏蒸7年后几乎从森林冠层中消失(Kubiske et al., 2007).然而, 由于种内及种间复杂的竞争关系, O3对生态系统群落结构的影响有时候并不是统一的, 高浓度O3暴露环境下O3敏感型种群首先达到较高覆盖度的情况也时有发生(Evans & Ashmore, 1992; Barbo et al., 1998).此外, 由于生态系统各营养级之间的交互作用, 高浓度O3影响植物群落多样性的同时也潜在影响植物组织、化学组成、二次代谢产物(如酚苷类化合物、植物源挥发性有机化合物等)释放, 进而改变植物种群周边与之相关的动植物、真菌、细菌和昆虫行为或活动(ICP Vegetation, 2013; Fuhrer et al., 2016).例如, O3暴露使得西黄松(Pinus ponderosa)更易受到小蠹虫的袭击(Dahlsten et al., 1997), 但O3诱导红云杉(Picea rubens)萜烯物质释放增加的同时减少了其上蚜虫幼虫的数量(Cannon, 1990); 而有关欧洲山杨的研究则发现O3暴露降低柳皮苷和特里杨苷含量, 增加了缩合单宁含量, 引起林分中蚜虫和森林天幕毛虫幼虫数量剧增, 但蚜虫捕食者或天敌(如瓢虫、草蛉和蜘蛛)的数量显著减少(Percy et al., 2002; Kopper & Lindroth, 2003). ...
Effects of elevated atmospheric CO2 and/or O3 on intra- and interspecific competitive ability of aspen
2
2007
... 有关自然环境下O3浓度升高对陆地生态系统影响的研究较为稀缺.但是现有开放式O3浓度增强系统(O3-FACE)观测平台下的长期研究也有着相似结果, Aspen FACE实验发现, O3浓度升高3-4年后, 欧洲山杨(Populus tremuloides)纯林、欧洲山杨-纸皮桦(Betula papyrifera)和欧洲山杨-糖枫(Acer saccharum)混交林的总生物量分别降低23%、13%和14% (Karnosky et al., 2003, 2005); 虽然在最初的7年内生物量的分配格局并未受O3影响(King et al., 2005), 但在实验11年后, 敏感型欧洲山杨几乎从森林冠层中消失(Kubiske et al., 2007).德国克兰兹堡郁闭冠层的成熟挪威云杉(Picea abies)和欧洲山毛榉(Fagus sylvatica)混交林(60年)对O3浓度升高长期响应的研究也表明, 8年O3暴露导致欧洲山毛榉的树干生产力降低了44% (Pretzsch et al., 2010).目前, 国内还没有研究涉及O3-FACE平台下O3浓度升高对森林生态系统碳汇能力的影响. ...
... 植物长期过量摄取O3还可以引起物种组成、冠层结构改变, 影响生态系统种群均匀度和丰富度, 威胁生态系统多样性(Fuhrer et al., 2016; Bergmann et al., 2017), 最终影响生态系统碳、氮和水循环(McLaughlin et al., 2007; Sun et al., 2012; Grulke & Heath, 2020).基于温室或OTCs控制实验下有关草本植物的多项研究都发现, 随着O3暴露时间的延长, 对O3相对敏感的物种/种群将会被对O3相对耐受的物种/种群替代(Fuhrer et al., 1994; Hayes et al., 2009), 非禾本科杂草与禾本科杂草的比例也在发生改变(Hayes et al., 2010).有关Aspen FACE森林生态系统的研究也表明, 尽管整个森林林分结构没有受到明显影响, 但敏感型欧洲山杨树种在O3熏蒸7年后几乎从森林冠层中消失(Kubiske et al., 2007).然而, 由于种内及种间复杂的竞争关系, O3对生态系统群落结构的影响有时候并不是统一的, 高浓度O3暴露环境下O3敏感型种群首先达到较高覆盖度的情况也时有发生(Evans & Ashmore, 1992; Barbo et al., 1998).此外, 由于生态系统各营养级之间的交互作用, 高浓度O3影响植物群落多样性的同时也潜在影响植物组织、化学组成、二次代谢产物(如酚苷类化合物、植物源挥发性有机化合物等)释放, 进而改变植物种群周边与之相关的动植物、真菌、细菌和昆虫行为或活动(ICP Vegetation, 2013; Fuhrer et al., 2016).例如, O3暴露使得西黄松(Pinus ponderosa)更易受到小蠹虫的袭击(Dahlsten et al., 1997), 但O3诱导红云杉(Picea rubens)萜烯物质释放增加的同时减少了其上蚜虫幼虫的数量(Cannon, 1990); 而有关欧洲山杨的研究则发现O3暴露降低柳皮苷和特里杨苷含量, 增加了缩合单宁含量, 引起林分中蚜虫和森林天幕毛虫幼虫数量剧增, 但蚜虫捕食者或天敌(如瓢虫、草蛉和蜘蛛)的数量显著减少(Percy et al., 2002; Kopper & Lindroth, 2003). ...
Extracellular enzyme activity beneath temperate trees growing under elevated carbon dioxide and ozone
1
2002
... 此外, 虽然地下生态系统并不是直接暴露在O3环境下, 但大量证据表明O3对地上生态系统的影响将会转移至地下(Andersen, 2003; Lindroth, 2010).如高浓度O3环境下, 植物分配到根的生物量将会大大减少, 基于100多组实验数据的meta分析研究表明O3暴露导致生物量分配到地下的量平均减少5.6% (Grantz et al., 2006).另外, 凋落物分解是生态系统营养循环的关键过程.通过影响凋落物的数量、质量和分解速率, 高浓度地表O3会间接改变土壤微生物群落活性、组成和结构, 土壤化学属性及土壤营养循环(Larson et al., 2002; Fuhrer et al., 2016).然而, 比较遗憾的是, 目前有关O3对地下生态过程影响的研究仅集中在农田生态系统, 仅有少部分研究涉及森林土壤(主要来自Aspen FACE)或草地生态系统(ICP Vegetation, 2013).而且, 由于凋落物种类、化学成分、分解者种群、土壤和大气环境条件的不同, 高浓度O3对凋落物分解动态的影响还存在很大争议(Gessner et al., 2010; Fuhrer et al., 2016; Grulke & Heath, 2020).例如, 研究发现O3对欧洲白桦(Betula pendula)叶片凋落物质量和损失率有明显影响(Kasurinen et al., 2006, 2007), 但不影响欧洲山毛榉凋落物的分解速率(Schloter et al., 2005).有关地表O3浓度升高对地下生态系统结构和功能的影响还有待进一步探究. ...
Lines of discovery in the history of ozone
1
1880
... 地表O3的发现最早可追溯到1839年(Leeds, 1880), 但直到1945年“洛杉矶烟雾”爆发后, 地表O3生态效应的研究才逐渐进入学者的视野(Rogers et al., 1956).经过近百年的发展, 涉及地表O3对植物、土壤及生态系统影响的研究, 已经遍及世界上大多数国家, 发表了2 500多篇期刊论文和十余本英文专著.到目前为止, O3表观伤害症状的描述及症状等级的鉴定(如叶片萎黄、褪绿和坏死等典型症状分类), O3伤害机理的探究(如O3降低光合速率的气孔与非气孔因素等), O3对植物叶片(光合作用、气孔导度及抗氧化系统等), 个体(生物量、产量或品质), 种群(种间关系、群落构成)及生态系统(生产力和固碳能力等)的影响, 及O3污染防治对策的探讨(前体物减排措施、化学防护剂的应用和作物栽培管理模式的筛选)等方面都进行了大量相关研究(Ashmore, 2005; Grulke & Heath, 2020).然而地表O3浓度升高对陆地生态系统水平碳、氮和水循环、营养交互、群落更替及地下生态系统动态变化过程(如根系功能变化、根际过程机制、土壤微生物活性及反馈等)方面的研究仍鲜有报道(Fuhrer et al., 2016).通过系统梳理和总结目前国内外研究成果, 本文对地表O3浓度升高对生态系统影响这一领域的研究历史、研究方法、评估指标、重大进展和存在的主要问题进行了回顾分析, 并对未来研究进行了展望. ...
NCLAN results and their application to the standard setting process: protecting vegetation from surface ozone exposures
1
1992
... 不管研究方法如何更新, 由于空间和时间上的限制, 模拟地表O3浓度升高的控制实验主要探究高浓度O3对植物损伤的机理, 而区域尺度上探究O3对陆地生态系统的影响却很难通过模拟实验获得.为评价区域O3风险和评估O3对生态系统影响带来的损失, 学者主要通过不同评估指标评估区域、国家或全球尺度上地表O3污染的生态效应.总体来说, 评估指标的研究主要经历了三个阶段, 分别是浓度响应关系、剂量响应关系和通量响应关系评估指标(Musselman et al., 2006).浓度响应关系评估指标主要包括M7/M12 (白天7 h (9:00-16:00) O3浓度平均值或白天12 h (8:00-20:00) O3浓度平均值), 早期曾广泛应用于表征O3浓度暴露与农作物产量损失相关性的研究中(Legge et al., 1995).剂量响应关系评估指标主要有SUM06 (O3小时浓度高于或等于60 μg·kg-1的累积值)(Lefohn & Foley, 1992), W126 (O3小时浓度在规定时段内的加权求和值)和AOTX (整个生长季太阳辐射>50 W·m-2时段内O3小时浓度超过X μg·kg-1的累计值, X普遍设为40 μg·kg-1)(Fuhrer et al., 1997; LRTAR Convention, 2015).剂量响应指标综合考虑O3浓度和暴露时间对植物生长的影响, 在世界范围内得到广泛应用. ...
Tropospheric ozone assessment report: global ozone metrics for climate change, human health, and crop/ecosystem research
1
2018
... (5)开展联网研究, 建立统一评价体系: 目前, 全球覆盖面最广的地表O3监测网络——TOAR (Tropospheric Ozone Assessment Report)已拥有大大小小近万个监测网点(Mills et al., 2018).但考虑到站点分布的不均(80%的监测网点分布在北美和欧洲地区, 亚洲地区的站点占据16%, 但主要集中在日本和韩国, 中国地区只有26个监测网点), 观测频率、时长、年份及评价标准或评价体系的差异(如部分站点为长期研究, 有些则为短期观测; 一些研究采用SUM06或W126作为评估指标, 另一些研究使用AOTX和PODY)(Lefohn et al., 2018), 不同地区、不同学者开展的实验研究结果差异较大, 无法进行统一对比.亚洲地区的研究网点及加入全球的联网实验仍然稀少.因此, 未来亟须扩展联网观测, 建立统一标准为区域或全球O3风险评估提供更具可比性的实测数据. ...
Ambient ozone and adverse crop response: an evaluation of north American and European data as they relate to exposure indices and critical levels
1
1995
... 不管研究方法如何更新, 由于空间和时间上的限制, 模拟地表O3浓度升高的控制实验主要探究高浓度O3对植物损伤的机理, 而区域尺度上探究O3对陆地生态系统的影响却很难通过模拟实验获得.为评价区域O3风险和评估O3对生态系统影响带来的损失, 学者主要通过不同评估指标评估区域、国家或全球尺度上地表O3污染的生态效应.总体来说, 评估指标的研究主要经历了三个阶段, 分别是浓度响应关系、剂量响应关系和通量响应关系评估指标(Musselman et al., 2006).浓度响应关系评估指标主要包括M7/M12 (白天7 h (9:00-16:00) O3浓度平均值或白天12 h (8:00-20:00) O3浓度平均值), 早期曾广泛应用于表征O3浓度暴露与农作物产量损失相关性的研究中(Legge et al., 1995).剂量响应关系评估指标主要有SUM06 (O3小时浓度高于或等于60 μg·kg-1的累积值)(Lefohn & Foley, 1992), W126 (O3小时浓度在规定时段内的加权求和值)和AOTX (整个生长季太阳辐射>50 W·m-2时段内O3小时浓度超过X μg·kg-1的累计值, X普遍设为40 μg·kg-1)(Fuhrer et al., 1997; LRTAR Convention, 2015).剂量响应指标综合考虑O3浓度和暴露时间对植物生长的影响, 在世界范围内得到广泛应用. ...
Differences in ozone sensitivity among woody species are related to leaf morphology and antioxidant levels
1
2016
... 气孔防御机制失效后, 进入细胞的O3将诱导叶片内部抗氧化系统启动解毒和修复的非气孔防御响应(冯兆忠等, 2018).非气孔防御响应主要是指“质外体-共质体系统”内一些抗氧化组分对O3解毒和修复的防御过程.具体来说, O3进入植物叶片的细胞间隙后会溶解在细胞壁的结合水里, 经过一系列的化学反应形成活性氧分子(ROS), ROS能够破坏细胞壁并与细胞膜上的不饱和脂肪酸发生反应, 从而对叶片造成损伤(Tausz et al., 2007; Pellegrini et al., 2018).植物质外体的抗氧化物质如还原性抗坏血酸(ASC)、抗氧化酶(如超氧化物歧化酶)可与ROS反应进行初级解毒, 成为抵御O3的第二道防线(Kangasjärvi et al., 2005; Dai et al., 2019), 而剩余的ROS会经过质膜到达共质体, 此时共质体内抗氧化成分如过氧化物酶(POD)、过氧化氢酶(CAT)、抗坏血酸过氧化物酶(APX)及抗氧化剂(如酚类化合物-谷胱甘肽(GSH)、脯氨酸和多胺等)等抗氧化酶类物质被诱导作为次级解毒响应机制参与诸如“抗坏血酸-谷胱甘肽(ASA-GSH)循环”等氧化还原过程, 进而对ROS进行进一步解毒与消除(Luwe, 1996).气孔闭合除了会减少O3的吸收, 也会抑制CO2吸收和水汽交换, 而非气孔防御响应中的抗氧化系统解毒过程也会消耗大量的能量并对光合反应系统造成不同程度的损伤, 降低羧化速率和电子传递速率, 进而抑制光合和蒸腾作用, 损害植物生长(Gao et al., 2016).然而, 最新研究表明持续整个生长季的长期O3暴露会影响植物叶片的气孔密度、开度和大小等表观属性(李品等, 2018), 而O3从气孔进入到细胞间隙后, 也可以通过影响植物的叶肉导度降低植物的光合能力(Xu et al., 2019).此外, 有关29种落叶和常绿树种应对O3胁迫的研究还发现不同木本植物对O3响应的差异主要来自植物功能叶片比叶质量的大小(即叶片比叶质量越大的树种对O3越不敏感), 与气孔导度无明显相关性(Li et al., 2016). ...
A meta-analysis on growth, physiological, and biochemical responses of woody species to ground-level ozone highlights the role of plant functional types
1
2017
... 地表O3浓度升高对植物光合作用、生长和生物量积累、物质分配等过程的连锁负效应导致陆地生态系统生产力和固碳能力降低(Reich, 1987; Wittig et al., 2009; Ainsworth et al., 2012), 削弱了自然生态系统的碳汇能力(Felzer et al., 2005; Sitch et al., 2007; Collins et al., 2010).基于大量OTCs控制实验下北半球中纬度北方温带树种的meta分析发现, O3浓度升高导致树木的净光合作用和总生物量分别降低11%和7% (Wittig et al., 2009).最近, 有关中国热带、亚热带和温带树种的meta分析则发现, O3浓度升高导致中国木本植物的净光合作用和总生物量分别降低28%和14% (Li et al., 2017). ...
6种绿化树种的气孔特性与臭氧剂量的响应关系
2018
6种绿化树种的气孔特性与臭氧剂量的响应关系
2018
臭氧与其他环境因子对植物的交互作用
6
2014
... 气孔防御机制失效后, 进入细胞的O3将诱导叶片内部抗氧化系统启动解毒和修复的非气孔防御响应(冯兆忠等, 2018).非气孔防御响应主要是指“质外体-共质体系统”内一些抗氧化组分对O3解毒和修复的防御过程.具体来说, O3进入植物叶片的细胞间隙后会溶解在细胞壁的结合水里, 经过一系列的化学反应形成活性氧分子(ROS), ROS能够破坏细胞壁并与细胞膜上的不饱和脂肪酸发生反应, 从而对叶片造成损伤(Tausz et al., 2007; Pellegrini et al., 2018).植物质外体的抗氧化物质如还原性抗坏血酸(ASC)、抗氧化酶(如超氧化物歧化酶)可与ROS反应进行初级解毒, 成为抵御O3的第二道防线(Kangasjärvi et al., 2005; Dai et al., 2019), 而剩余的ROS会经过质膜到达共质体, 此时共质体内抗氧化成分如过氧化物酶(POD)、过氧化氢酶(CAT)、抗坏血酸过氧化物酶(APX)及抗氧化剂(如酚类化合物-谷胱甘肽(GSH)、脯氨酸和多胺等)等抗氧化酶类物质被诱导作为次级解毒响应机制参与诸如“抗坏血酸-谷胱甘肽(ASA-GSH)循环”等氧化还原过程, 进而对ROS进行进一步解毒与消除(Luwe, 1996).气孔闭合除了会减少O3的吸收, 也会抑制CO2吸收和水汽交换, 而非气孔防御响应中的抗氧化系统解毒过程也会消耗大量的能量并对光合反应系统造成不同程度的损伤, 降低羧化速率和电子传递速率, 进而抑制光合和蒸腾作用, 损害植物生长(Gao et al., 2016).然而, 最新研究表明持续整个生长季的长期O3暴露会影响植物叶片的气孔密度、开度和大小等表观属性(李品等, 2018), 而O3从气孔进入到细胞间隙后, 也可以通过影响植物的叶肉导度降低植物的光合能力(Xu et al., 2019).此外, 有关29种落叶和常绿树种应对O3胁迫的研究还发现不同木本植物对O3响应的差异主要来自植物功能叶片比叶质量的大小(即叶片比叶质量越大的树种对O3越不敏感), 与气孔导度无明显相关性(Li et al., 2016). ...
... 除单因子O3胁迫外, 由于全球气候变化和极端气候常态化, 植物经常受到O3和其他环境因子交互作用的影响(如O3与CO2、升温、干旱或氮沉降等)(Schmadel-Hagebölling et al., 1998; Ollinger et al., 2002; Eltayeb et al., 2006; Ghimire et al., 2018).然而, 植物应对O3与其他因子交互作用的响应并不是两者或单独作用的简单叠加, 往往取决于植物种类、生长时期、对特定环境因子的敏感性、胁迫浓度及时间, 以及具体的微气候条件(列淦文和薛立, 2014).一方面, 通过诱导气孔关闭、限制叶片O3吸收(CO2浓度升高/干旱胁迫), 或提高碳同化速率、促进植物光合作用(CO2浓度升高/氮增加), CO2浓度升高、干旱胁迫和氮沉降经常被报道可增强植物抵御O3的能力, 减缓O3伤害(白月明等, 2005; Handley & Grulke, 2008; Gao et al., 2017).另一方面, 与之相反, 也有研究发现长期高浓度CO2或干旱降低气孔导度的同时, 会引起植物蒸腾速率、光合产物合成和积累的降低(McDonald et al., 2002; Pollastrini et al., 2010); 而超过植物承载阈值的高氮负荷也会对植物的光合生理活性产生负效应(Yamaguchi et al., 2010), 进而加重植物在复合环境下的碳损失、水分蒸散损失和自由基积累, 对植物产生协同伤害(Karnosky et al., 2005; 列淦文和薛立, 2014).高峰(2018)认为O3与其他环境胁迫因子出现的先后次序也会影响植物对复合作用的响应. ...
... ; 列淦文和薛立, 2014).高峰(2018)认为O3与其他环境胁迫因子出现的先后次序也会影响植物对复合作用的响应. ...
... (3)开展O3与其他环境因子复合研究: 全球气候变化、极端气候常态化和空气污染加重使两个或两个以上环境因子的交互作用大大增加, 复合作用下的实验条件也更接近植物生长的自然环境.现有的研究多集中在O3与CO2对植物或生态系统的共同影响, 并发现O3污染引起的负面作用经常被高浓度CO2对植物叶片光合作用或个体生物量累积的“肥效”所削弱(Ollinger et al., 2002).然而, 目前还无法确定这种结果是否与生态系统水平的响应相匹配(Grulke & Heath, 2020).植物应对O3胁迫的响应与土壤水分、氮有效性等因素密切相关(列淦文和薛立, 2014; 高峰, 2018).控制实验下的研究也发现, O3浓度的升高可能削弱植物对干旱或其他环境胁迫的抵抗力, 使生态系统更易受到病虫害的侵袭(Grulke et al., 2009).但是迄今为止, 国内外大部分研究多集中在温带农作物和森林树种上, 较少涉及亚热带地区乡土树种.由于生态系统类型差异, 各级营养交互及种间竞争的复杂性, 不同生态系统下O3与生物及非生物因子之间的多重交互作用机制还需进一步探究.此外, 已有研究多针对植物地上部分, 对植物地下部分及其功能变化与根际过程相互影响的机制、土壤关键过程研究较少(列淦文和薛立, 2014).因此, 在开展O3单因子实验研究的同时, 亟须考虑多个因子(如O3、CO2、水分和氮沉降等)共存对整个生态系统结构和功能的复合影响研究. ...
... 与生物及非生物因子之间的多重交互作用机制还需进一步探究.此外, 已有研究多针对植物地上部分, 对植物地下部分及其功能变化与根际过程相互影响的机制、土壤关键过程研究较少(列淦文和薛立, 2014).因此, 在开展O3单因子实验研究的同时, 亟须考虑多个因子(如O3、CO2、水分和氮沉降等)共存对整个生态系统结构和功能的复合影响研究. ...
... (6)探究地表O3污染的生态控制措施: 欧洲和北美等发达国家治理大气污染的经验告诉我们, 严格的O3前体物NOx和挥发性有机化合物(VOCs)减排措施是降低地表O3浓度的有效手段(Feng et al., 2015b).然而, 以完全牺牲经济发展为代价的环境保护措施一般并不被发展中国家所接受.事实上, 自然植被与地表O3存在复杂的交互作用.一方面, 植物释放的VOCs有助于地表O3的形成; 另一方面, 植物在进行光合作用的同时也可以通过气孔、非气孔途径吸收或沉降大气O3 (Paoletti, 2009).考虑到植被VOCs释放和O3移除能力都具有树种特异性, 因此, 在一定条件下通过合理的树种筛选达到减少大气O3污染的目的将是完全可行的(Sicard et al., 2018).另外, 除植物自身遗传基因之外, 植物对O3胁迫的响应受土壤水分和营养元素如氮可利用性的影响(列淦文和薛立, 2014; 高峰, 2018).例如古巴农民通过提前1-2天灌溉的方式预防夏季高浓度O3对莴苣和烟草叶面的损伤(ICP Vegetation, 2011); 马来西亚研究者通过氮肥管理, 减轻O3污染对热带Elaeis guineensis产量和质量的伤害(Hewitt et al., 2009).因此通过合理的田间水肥管理措施(如改变灌溉模式、施肥剂量和方式)调控植物生长, 增强植物对O3的抗性, 进而减轻O3污染的生态效应是未来大气O3污染防治研究中值得思考的问题. ...
臭氧与其他环境因子对植物的交互作用
6
2014
... 气孔防御机制失效后, 进入细胞的O3将诱导叶片内部抗氧化系统启动解毒和修复的非气孔防御响应(冯兆忠等, 2018).非气孔防御响应主要是指“质外体-共质体系统”内一些抗氧化组分对O3解毒和修复的防御过程.具体来说, O3进入植物叶片的细胞间隙后会溶解在细胞壁的结合水里, 经过一系列的化学反应形成活性氧分子(ROS), ROS能够破坏细胞壁并与细胞膜上的不饱和脂肪酸发生反应, 从而对叶片造成损伤(Tausz et al., 2007; Pellegrini et al., 2018).植物质外体的抗氧化物质如还原性抗坏血酸(ASC)、抗氧化酶(如超氧化物歧化酶)可与ROS反应进行初级解毒, 成为抵御O3的第二道防线(Kangasjärvi et al., 2005; Dai et al., 2019), 而剩余的ROS会经过质膜到达共质体, 此时共质体内抗氧化成分如过氧化物酶(POD)、过氧化氢酶(CAT)、抗坏血酸过氧化物酶(APX)及抗氧化剂(如酚类化合物-谷胱甘肽(GSH)、脯氨酸和多胺等)等抗氧化酶类物质被诱导作为次级解毒响应机制参与诸如“抗坏血酸-谷胱甘肽(ASA-GSH)循环”等氧化还原过程, 进而对ROS进行进一步解毒与消除(Luwe, 1996).气孔闭合除了会减少O3的吸收, 也会抑制CO2吸收和水汽交换, 而非气孔防御响应中的抗氧化系统解毒过程也会消耗大量的能量并对光合反应系统造成不同程度的损伤, 降低羧化速率和电子传递速率, 进而抑制光合和蒸腾作用, 损害植物生长(Gao et al., 2016).然而, 最新研究表明持续整个生长季的长期O3暴露会影响植物叶片的气孔密度、开度和大小等表观属性(李品等, 2018), 而O3从气孔进入到细胞间隙后, 也可以通过影响植物的叶肉导度降低植物的光合能力(Xu et al., 2019).此外, 有关29种落叶和常绿树种应对O3胁迫的研究还发现不同木本植物对O3响应的差异主要来自植物功能叶片比叶质量的大小(即叶片比叶质量越大的树种对O3越不敏感), 与气孔导度无明显相关性(Li et al., 2016). ...
... 除单因子O3胁迫外, 由于全球气候变化和极端气候常态化, 植物经常受到O3和其他环境因子交互作用的影响(如O3与CO2、升温、干旱或氮沉降等)(Schmadel-Hagebölling et al., 1998; Ollinger et al., 2002; Eltayeb et al., 2006; Ghimire et al., 2018).然而, 植物应对O3与其他因子交互作用的响应并不是两者或单独作用的简单叠加, 往往取决于植物种类、生长时期、对特定环境因子的敏感性、胁迫浓度及时间, 以及具体的微气候条件(列淦文和薛立, 2014).一方面, 通过诱导气孔关闭、限制叶片O3吸收(CO2浓度升高/干旱胁迫), 或提高碳同化速率、促进植物光合作用(CO2浓度升高/氮增加), CO2浓度升高、干旱胁迫和氮沉降经常被报道可增强植物抵御O3的能力, 减缓O3伤害(白月明等, 2005; Handley & Grulke, 2008; Gao et al., 2017).另一方面, 与之相反, 也有研究发现长期高浓度CO2或干旱降低气孔导度的同时, 会引起植物蒸腾速率、光合产物合成和积累的降低(McDonald et al., 2002; Pollastrini et al., 2010); 而超过植物承载阈值的高氮负荷也会对植物的光合生理活性产生负效应(Yamaguchi et al., 2010), 进而加重植物在复合环境下的碳损失、水分蒸散损失和自由基积累, 对植物产生协同伤害(Karnosky et al., 2005; 列淦文和薛立, 2014).高峰(2018)认为O3与其他环境胁迫因子出现的先后次序也会影响植物对复合作用的响应. ...
... ; 列淦文和薛立, 2014).高峰(2018)认为O3与其他环境胁迫因子出现的先后次序也会影响植物对复合作用的响应. ...
... (3)开展O3与其他环境因子复合研究: 全球气候变化、极端气候常态化和空气污染加重使两个或两个以上环境因子的交互作用大大增加, 复合作用下的实验条件也更接近植物生长的自然环境.现有的研究多集中在O3与CO2对植物或生态系统的共同影响, 并发现O3污染引起的负面作用经常被高浓度CO2对植物叶片光合作用或个体生物量累积的“肥效”所削弱(Ollinger et al., 2002).然而, 目前还无法确定这种结果是否与生态系统水平的响应相匹配(Grulke & Heath, 2020).植物应对O3胁迫的响应与土壤水分、氮有效性等因素密切相关(列淦文和薛立, 2014; 高峰, 2018).控制实验下的研究也发现, O3浓度的升高可能削弱植物对干旱或其他环境胁迫的抵抗力, 使生态系统更易受到病虫害的侵袭(Grulke et al., 2009).但是迄今为止, 国内外大部分研究多集中在温带农作物和森林树种上, 较少涉及亚热带地区乡土树种.由于生态系统类型差异, 各级营养交互及种间竞争的复杂性, 不同生态系统下O3与生物及非生物因子之间的多重交互作用机制还需进一步探究.此外, 已有研究多针对植物地上部分, 对植物地下部分及其功能变化与根际过程相互影响的机制、土壤关键过程研究较少(列淦文和薛立, 2014).因此, 在开展O3单因子实验研究的同时, 亟须考虑多个因子(如O3、CO2、水分和氮沉降等)共存对整个生态系统结构和功能的复合影响研究. ...
... 与生物及非生物因子之间的多重交互作用机制还需进一步探究.此外, 已有研究多针对植物地上部分, 对植物地下部分及其功能变化与根际过程相互影响的机制、土壤关键过程研究较少(列淦文和薛立, 2014).因此, 在开展O3单因子实验研究的同时, 亟须考虑多个因子(如O3、CO2、水分和氮沉降等)共存对整个生态系统结构和功能的复合影响研究. ...
... (6)探究地表O3污染的生态控制措施: 欧洲和北美等发达国家治理大气污染的经验告诉我们, 严格的O3前体物NOx和挥发性有机化合物(VOCs)减排措施是降低地表O3浓度的有效手段(Feng et al., 2015b).然而, 以完全牺牲经济发展为代价的环境保护措施一般并不被发展中国家所接受.事实上, 自然植被与地表O3存在复杂的交互作用.一方面, 植物释放的VOCs有助于地表O3的形成; 另一方面, 植物在进行光合作用的同时也可以通过气孔、非气孔途径吸收或沉降大气O3 (Paoletti, 2009).考虑到植被VOCs释放和O3移除能力都具有树种特异性, 因此, 在一定条件下通过合理的树种筛选达到减少大气O3污染的目的将是完全可行的(Sicard et al., 2018).另外, 除植物自身遗传基因之外, 植物对O3胁迫的响应受土壤水分和营养元素如氮可利用性的影响(列淦文和薛立, 2014; 高峰, 2018).例如古巴农民通过提前1-2天灌溉的方式预防夏季高浓度O3对莴苣和烟草叶面的损伤(ICP Vegetation, 2011); 马来西亚研究者通过氮肥管理, 减轻O3污染对热带Elaeis guineensis产量和质量的伤害(Hewitt et al., 2009).因此通过合理的田间水肥管理措施(如改变灌溉模式、施肥剂量和方式)调控植物生长, 增强植物对O3的抗性, 进而减轻O3污染的生态效应是未来大气O3污染防治研究中值得思考的问题. ...
Impacts of O3 on premature mortality and crop yield loss across China
1
2018
... 基于OTCs和O3-FACE平台下单个站点的控制实验可探究O3对植物个体、群落或生态系统的影响, 但将站点或样地尺度的研究外推到区域或全球, 评估O3污染区域风险的研究主要通过响应关系和模型.目前, 其研究领域主要集中在地表O3污染对农作物产量和森林生产力等方面.早在1994年, Chameides等(1994)就利用经济模型初步预测O3浓度升高可能导致2025年全球30%-75%的作物面临产量降低的风险.基于O3前体物排放、化学反应传输模型和M12、AOT40指标的评估显示, 2000年全球O3污染引起全球小麦、玉米和大豆产量平均降低2.2%-15%, 每年因上述作物产量降低所导致的经济损失达110-180亿美元(Avnery et al., 2011).最近, 基于PODY评估指标的研究则表明, O3对小麦造成的产量损失北半球(9.9%)明显高于南半球(6.2%), 平均产量损失最高的地区主要位于北半球暖温带湿润地区(12.5%)、热带湿热(12.5%)和热带潮湿气候区(17.2%)(Mills et al., 2018).在区域尺度上, 基于1990年O3浓度数据, Wang和Mauzerall (2004)利用M7和M12指标评估了O3对中国、日本和韩国主要农作物产量的影响, 结果显示O3污染造成小麦、玉米、棉花减产1%-9%, 大豆减产23%-27%, 造成2.4-35亿美元的经济损失, 在维持现有的污染物排放、管理措施和农业生产技术不变的情况下, 预计到2020年小麦、玉米、棉花减产幅度为2%-15%, 大豆为28%-35%.最近, 利用AOT40指标的研究显示, 2014年当前O3浓度导致中国农作物产量损失7 840万t, 小麦、水稻和玉米相对减产量分别为8.5%-14%、3.9%-15%和2.2%-5.5% (Lin et al., 2018).而只聚焦于长江三角洲地区的研究则发现, 2015年区域O3污染分别造成小麦和水稻减产9.9%-36.1%和7.3%- 23.9% (Zhao et al., 2018).虽然国内也有少量研究建立了PODY与个别树种或作物相对生物量或相对产量之间的通量响应关系(Feng et al., 2012; Hu et al., 2015; Gao et al., 2017), 但并没有研究利用PODY指标评估区域O3污染对农作物产量损失的影响. ...
Impacts of elevated atmospheric CO2 and O3 on forests: phytochemistry, trophic interactions, and ecosystem dynamics
1
2010
... 此外, 虽然地下生态系统并不是直接暴露在O3环境下, 但大量证据表明O3对地上生态系统的影响将会转移至地下(Andersen, 2003; Lindroth, 2010).如高浓度O3环境下, 植物分配到根的生物量将会大大减少, 基于100多组实验数据的meta分析研究表明O3暴露导致生物量分配到地下的量平均减少5.6% (Grantz et al., 2006).另外, 凋落物分解是生态系统营养循环的关键过程.通过影响凋落物的数量、质量和分解速率, 高浓度地表O3会间接改变土壤微生物群落活性、组成和结构, 土壤化学属性及土壤营养循环(Larson et al., 2002; Fuhrer et al., 2016).然而, 比较遗憾的是, 目前有关O3对地下生态过程影响的研究仅集中在农田生态系统, 仅有少部分研究涉及森林土壤(主要来自Aspen FACE)或草地生态系统(ICP Vegetation, 2013).而且, 由于凋落物种类、化学成分、分解者种群、土壤和大气环境条件的不同, 高浓度O3对凋落物分解动态的影响还存在很大争议(Gessner et al., 2010; Fuhrer et al., 2016; Grulke & Heath, 2020).例如, 研究发现O3对欧洲白桦(Betula pendula)叶片凋落物质量和损失率有明显影响(Kasurinen et al., 2006, 2007), 但不影响欧洲山毛榉凋落物的分解速率(Schloter et al., 2005).有关地表O3浓度升高对地下生态系统结构和功能的影响还有待进一步探究. ...
Global food insecurity. Treatment of major food crops with elevated carbon dioxide or ozone under large-scale fully open-air conditions suggests recent models may have overestimated future yields
1
2005
... (1)开展O3-FACE研究: 现有研究多为OTC系统下的实验, OTC气室内“微气候” (如温度、水分、光照和风力等生态因素)与外界环境状态存在明显差异, 这些生态因子的变化可能影响植物应对O3的响应, 缓解或加重O3污染对农作物产量或森林植被生产力的损伤(Long et al., 2005; Feng et al., 2018a).全球模型评估结果显示, OTC实验结果低估了O3对作物产量损失的影响(van Dingenen et al., 2009).最近, Feng等(2018b)也发现, 同样浓度的O3对OTC实验下小麦和水稻产量的影响要低于FACE实验; 而对大豆产量的影响则相反.因此, OTC实验结果并不能完全应用于实际O3区域风险的评估.O3-FACE平台是未来研究实际大田环境下O3浓度升高对农林生态系统影响的主要依托平台.为减少区域O3风险评估的不确定性, 未来应加大O3-FACE平台的实验研究.考虑到目前已开展的FACE实验多坐落在温带地区, 主要探究温带植被或作物对高浓度O3的响应, 较少涉及热带和寒带植物或其他区域典型生态系统(Ainsworth et al., 2020).最理想的情况是, 不同区域、不同生态系统类型共同开展O3-FACE实验研究(Ainsworth et al., 2012). ...
2
2015
... 不管研究方法如何更新, 由于空间和时间上的限制, 模拟地表O3浓度升高的控制实验主要探究高浓度O3对植物损伤的机理, 而区域尺度上探究O3对陆地生态系统的影响却很难通过模拟实验获得.为评价区域O3风险和评估O3对生态系统影响带来的损失, 学者主要通过不同评估指标评估区域、国家或全球尺度上地表O3污染的生态效应.总体来说, 评估指标的研究主要经历了三个阶段, 分别是浓度响应关系、剂量响应关系和通量响应关系评估指标(Musselman et al., 2006).浓度响应关系评估指标主要包括M7/M12 (白天7 h (9:00-16:00) O3浓度平均值或白天12 h (8:00-20:00) O3浓度平均值), 早期曾广泛应用于表征O3浓度暴露与农作物产量损失相关性的研究中(Legge et al., 1995).剂量响应关系评估指标主要有SUM06 (O3小时浓度高于或等于60 μg·kg-1的累积值)(Lefohn & Foley, 1992), W126 (O3小时浓度在规定时段内的加权求和值)和AOTX (整个生长季太阳辐射>50 W·m-2时段内O3小时浓度超过X μg·kg-1的累计值, X普遍设为40 μg·kg-1)(Fuhrer et al., 1997; LRTAR Convention, 2015).剂量响应指标综合考虑O3浓度和暴露时间对植物生长的影响, 在世界范围内得到广泛应用. ...
... 然而, O3对植物的影响不仅依赖于大气O3浓度同时也依赖于叶片气孔导度(O3的吸收)和植物自身抗氧化能力(Matyssek et al., 2007).近年来, 基于气孔O3吸收量的评估指标PODY (植物毒性O3剂量, 整个生长季单位面积上气孔O3吸收通量超过临界值Y nmol m-2·s-1的积累量)被开发并逐步替代浓度暴露剂量指标, 用于区域或全球O3对农作物和森林树种生长影响的评估(Büker et al., 2015; LRTAR Convention, 2015).目前, 哪个指标更适合区域O3风险评估的争论一直存在, 但欧洲和亚洲地区多项O3对农作物或植被影响的研究均表明: 考虑水分供应的情况下, O3暴露造成的生物量或作物产量的降低与PODY的拟合度比AOTX更好(Mills et al., 2011; Gao et al., 2017), 能更好表征O3对生态系统的影响.整体来看, 各个评估指标的计算方式和所需参数从简单到复杂, 从单一考虑O3浓度到综合考虑环境因子对植物O3吸收通量的影响, 逐渐可以精确评估地表O3污染对生态系统农林生产力的影响(表3). ...
Severe surface ozone pollution in China: a global perspective
1
2018
... 地表臭氧(O3)特指距离地球表面15 km范围内的近地层O3, 除少量来自平流层大气传输外, 其余大部分是氮氧化物(NOx)、非甲烷类挥发性有机化合物(NMVOCs)、一氧化碳(CO)和甲烷(CH4)等前体物在强烈光照下发生光化学反应而产生(冯兆忠等, 2018).自工业革命以来, 伴随城市化进程加快和化石燃料的过度燃烧, 地表O3浓度在世界范围内普遍升高.近年来, 模型和观测数据都表明地表O3浓度小时峰值在欧洲和北美等地已有所下降, 但亚洲地区仍呈不断上升的趋势(Cooper et al., 2014).中国国家环境监测网络2013-2017年的数据表明, 过去5年中国地表O3浓度以每年3 μg·kg-1的速率攀升(Wang et al., 2019), 目前全国地表O3浓度每日8 h滑动平均最大值(MDA8)的年平均值已达(41.2 ± 6.3) μg·kg-1, 尤其华北平原、长江三角洲和珠江三角洲等地(MDA8峰值年平均值已处于60-70 μg·kg-1), 地表O3污染形势十分严峻(Lu et al., 2018).地表O3是具有植物毒性的气体污染物, 具有强氧化性.高浓度地表O3或长期低浓度O3能诱导植物叶片出现可见损伤, 加速植物衰老, 降低光合作用, 改变碳分配, 抑制植物生长和生物量积累, 影响作物品质和产量, 最终降低生态系统碳汇能力, 改变生态系统结构和功能(Ashmore, 2005; Ainsworth, 2017; 高峰, 2018). ...
Antioxidants in the apoplast and symplast of beech (Fagus sylvatica L.) leaves: seasonal variations and responses to changing ozone concentrations in air
1
1996
... 气孔防御机制失效后, 进入细胞的O3将诱导叶片内部抗氧化系统启动解毒和修复的非气孔防御响应(冯兆忠等, 2018).非气孔防御响应主要是指“质外体-共质体系统”内一些抗氧化组分对O3解毒和修复的防御过程.具体来说, O3进入植物叶片的细胞间隙后会溶解在细胞壁的结合水里, 经过一系列的化学反应形成活性氧分子(ROS), ROS能够破坏细胞壁并与细胞膜上的不饱和脂肪酸发生反应, 从而对叶片造成损伤(Tausz et al., 2007; Pellegrini et al., 2018).植物质外体的抗氧化物质如还原性抗坏血酸(ASC)、抗氧化酶(如超氧化物歧化酶)可与ROS反应进行初级解毒, 成为抵御O3的第二道防线(Kangasjärvi et al., 2005; Dai et al., 2019), 而剩余的ROS会经过质膜到达共质体, 此时共质体内抗氧化成分如过氧化物酶(POD)、过氧化氢酶(CAT)、抗坏血酸过氧化物酶(APX)及抗氧化剂(如酚类化合物-谷胱甘肽(GSH)、脯氨酸和多胺等)等抗氧化酶类物质被诱导作为次级解毒响应机制参与诸如“抗坏血酸-谷胱甘肽(ASA-GSH)循环”等氧化还原过程, 进而对ROS进行进一步解毒与消除(Luwe, 1996).气孔闭合除了会减少O3的吸收, 也会抑制CO2吸收和水汽交换, 而非气孔防御响应中的抗氧化系统解毒过程也会消耗大量的能量并对光合反应系统造成不同程度的损伤, 降低羧化速率和电子传递速率, 进而抑制光合和蒸腾作用, 损害植物生长(Gao et al., 2016).然而, 最新研究表明持续整个生长季的长期O3暴露会影响植物叶片的气孔密度、开度和大小等表观属性(李品等, 2018), 而O3从气孔进入到细胞间隙后, 也可以通过影响植物的叶肉导度降低植物的光合能力(Xu et al., 2019).此外, 有关29种落叶和常绿树种应对O3胁迫的研究还发现不同木本植物对O3响应的差异主要来自植物功能叶片比叶质量的大小(即叶片比叶质量越大的树种对O3越不敏感), 与气孔导度无明显相关性(Li et al., 2016). ...
A cylindrical open top chamber for the exposure of plants to air pollutants in the field
1
1973
... 在此期间, 尽管社会经济不断发展、实验设备和研究方法不断更新, 但地表O3污染对植物及生态系统影响的研究主要还是以模拟O3浓度升高的控制实验为主.纵观发展历程, 试验设备与手段的改进和更新大致可分为三个阶段(王春乙, 1995): (1) 1970年之前, 研究主要利用封闭式静态或动态气室或室内生长箱进行模拟实验(Heagle et al., 1973). (2) 1973年之后, 美国学者Heagle等(1979)和Mandle (1973)研制开顶式气室(OTC)模拟高浓度O3对植物的损伤和影响机制, 并最终在世界范围内推广, 成为研究地表O3浓度升高对植物及生态系统影响的主流手段.(3)由于OTCs气室内外较大的环境差异, OTCs模拟实验下的熏蒸结果是否适用于大田研究一直备受争议.为解决OTCs内微气候环境带来的影响, 1986年Hendrey和Kimball (1994)提出建立自由空气中气体浓度增加(FACE)系统的想法, 并逐步用于大田实验.由于与外界大气几乎一致的气体流速、温度、光照和风速等条件(Matyssek et al., 2010), FACE系统被普遍认为是当前研究地表O3浓度升高对植物及生态系统影响最为理想的实验平台, 但由于造价昂贵, 技术要求高, 只有少数国家和实验室开展相关研究(有关上述研究方法的优缺点见表2).O3熏蒸的FACE平台仅有10个左右且多数已经停止运行(Paoletti et al., 2017), 主要包括美国威斯康星州Aspen FACE (停止运行; Karnosky et al., 2003), 美国伊利诺伊州Soybean FACE (停止运行; Morgan et al., 2004), 瑞典苏尔草地FACE (停止运行; Volk et al., 2003), 芬兰库奥皮奥FACE (停止运行; Oksanen, 2003), 德国克兰兹堡森林FACE (停止运行; Matyssek et al., 2010), 中国江都水稻(Oryza sativa)/小麦(Triticum aestivum)轮作FACE (停止运行; Tang et al., 2011), 日本札幌森林FACE (仍在运行; Watanabe et al., 2013), 意大利佛罗伦萨3D FACE (仍在运行; Paoletti et al., 2017), 以及刚刚建成的中国延庆唐家堡杨树人工林FACE.由于实验条件和FACE平台规模的限制, 有关地表O3浓度升高对生态系统水平的研究成果主要来自美国Aspen FACE和德国克兰兹堡森林FACE (Grulke & Heath, 2020). ...
Promoting the O3 flux concept for European forest trees
1
2007
... 然而, O3对植物的影响不仅依赖于大气O3浓度同时也依赖于叶片气孔导度(O3的吸收)和植物自身抗氧化能力(Matyssek et al., 2007).近年来, 基于气孔O3吸收量的评估指标PODY (植物毒性O3剂量, 整个生长季单位面积上气孔O3吸收通量超过临界值Y nmol m-2·s-1的积累量)被开发并逐步替代浓度暴露剂量指标, 用于区域或全球O3对农作物和森林树种生长影响的评估(Büker et al., 2015; LRTAR Convention, 2015).目前, 哪个指标更适合区域O3风险评估的争论一直存在, 但欧洲和亚洲地区多项O3对农作物或植被影响的研究均表明: 考虑水分供应的情况下, O3暴露造成的生物量或作物产量的降低与PODY的拟合度比AOTX更好(Mills et al., 2011; Gao et al., 2017), 能更好表征O3对生态系统的影响.整体来看, 各个评估指标的计算方式和所需参数从简单到复杂, 从单一考虑O3浓度到综合考虑环境因子对植物O3吸收通量的影响, 逐渐可以精确评估地表O3污染对生态系统农林生产力的影响(表3). ...
Enhanced ozone strongly reduces carbon sink strength of adult beech (Fagus sylvatica)—Resume from the free-air fumigation study at Kranzberg forest
2
2010
... 在此期间, 尽管社会经济不断发展、实验设备和研究方法不断更新, 但地表O3污染对植物及生态系统影响的研究主要还是以模拟O3浓度升高的控制实验为主.纵观发展历程, 试验设备与手段的改进和更新大致可分为三个阶段(王春乙, 1995): (1) 1970年之前, 研究主要利用封闭式静态或动态气室或室内生长箱进行模拟实验(Heagle et al., 1973). (2) 1973年之后, 美国学者Heagle等(1979)和Mandle (1973)研制开顶式气室(OTC)模拟高浓度O3对植物的损伤和影响机制, 并最终在世界范围内推广, 成为研究地表O3浓度升高对植物及生态系统影响的主流手段.(3)由于OTCs气室内外较大的环境差异, OTCs模拟实验下的熏蒸结果是否适用于大田研究一直备受争议.为解决OTCs内微气候环境带来的影响, 1986年Hendrey和Kimball (1994)提出建立自由空气中气体浓度增加(FACE)系统的想法, 并逐步用于大田实验.由于与外界大气几乎一致的气体流速、温度、光照和风速等条件(Matyssek et al., 2010), FACE系统被普遍认为是当前研究地表O3浓度升高对植物及生态系统影响最为理想的实验平台, 但由于造价昂贵, 技术要求高, 只有少数国家和实验室开展相关研究(有关上述研究方法的优缺点见表2).O3熏蒸的FACE平台仅有10个左右且多数已经停止运行(Paoletti et al., 2017), 主要包括美国威斯康星州Aspen FACE (停止运行; Karnosky et al., 2003), 美国伊利诺伊州Soybean FACE (停止运行; Morgan et al., 2004), 瑞典苏尔草地FACE (停止运行; Volk et al., 2003), 芬兰库奥皮奥FACE (停止运行; Oksanen, 2003), 德国克兰兹堡森林FACE (停止运行; Matyssek et al., 2010), 中国江都水稻(Oryza sativa)/小麦(Triticum aestivum)轮作FACE (停止运行; Tang et al., 2011), 日本札幌森林FACE (仍在运行; Watanabe et al., 2013), 意大利佛罗伦萨3D FACE (仍在运行; Paoletti et al., 2017), 以及刚刚建成的中国延庆唐家堡杨树人工林FACE.由于实验条件和FACE平台规模的限制, 有关地表O3浓度升高对生态系统水平的研究成果主要来自美国Aspen FACE和德国克兰兹堡森林FACE (Grulke & Heath, 2020). ...
... ), 德国克兰兹堡森林FACE (停止运行; Matyssek et al., 2010), 中国江都水稻(Oryza sativa)/小麦(Triticum aestivum)轮作FACE (停止运行; Tang et al., 2011), 日本札幌森林FACE (仍在运行; Watanabe et al., 2013), 意大利佛罗伦萨3D FACE (仍在运行; Paoletti et al., 2017), 以及刚刚建成的中国延庆唐家堡杨树人工林FACE.由于实验条件和FACE平台规模的限制, 有关地表O3浓度升高对生态系统水平的研究成果主要来自美国Aspen FACE和德国克兰兹堡森林FACE (Grulke & Heath, 2020). ...
Competitive status influences tree-growth responses to elevated CO2 and O3 in aggrading aspen stands
1
2002
... 除单因子O3胁迫外, 由于全球气候变化和极端气候常态化, 植物经常受到O3和其他环境因子交互作用的影响(如O3与CO2、升温、干旱或氮沉降等)(Schmadel-Hagebölling et al., 1998; Ollinger et al., 2002; Eltayeb et al., 2006; Ghimire et al., 2018).然而, 植物应对O3与其他因子交互作用的响应并不是两者或单独作用的简单叠加, 往往取决于植物种类、生长时期、对特定环境因子的敏感性、胁迫浓度及时间, 以及具体的微气候条件(列淦文和薛立, 2014).一方面, 通过诱导气孔关闭、限制叶片O3吸收(CO2浓度升高/干旱胁迫), 或提高碳同化速率、促进植物光合作用(CO2浓度升高/氮增加), CO2浓度升高、干旱胁迫和氮沉降经常被报道可增强植物抵御O3的能力, 减缓O3伤害(白月明等, 2005; Handley & Grulke, 2008; Gao et al., 2017).另一方面, 与之相反, 也有研究发现长期高浓度CO2或干旱降低气孔导度的同时, 会引起植物蒸腾速率、光合产物合成和积累的降低(McDonald et al., 2002; Pollastrini et al., 2010); 而超过植物承载阈值的高氮负荷也会对植物的光合生理活性产生负效应(Yamaguchi et al., 2010), 进而加重植物在复合环境下的碳损失、水分蒸散损失和自由基积累, 对植物产生协同伤害(Karnosky et al., 2005; 列淦文和薛立, 2014).高峰(2018)认为O3与其他环境胁迫因子出现的先后次序也会影响植物对复合作用的响应. ...
Interactive effects of ozone and climate on water use, soil moisture content and streamflow in a southern Appalachian forest in the USA
1
2007
... 植物长期过量摄取O3还可以引起物种组成、冠层结构改变, 影响生态系统种群均匀度和丰富度, 威胁生态系统多样性(Fuhrer et al., 2016; Bergmann et al., 2017), 最终影响生态系统碳、氮和水循环(McLaughlin et al., 2007; Sun et al., 2012; Grulke & Heath, 2020).基于温室或OTCs控制实验下有关草本植物的多项研究都发现, 随着O3暴露时间的延长, 对O3相对敏感的物种/种群将会被对O3相对耐受的物种/种群替代(Fuhrer et al., 1994; Hayes et al., 2009), 非禾本科杂草与禾本科杂草的比例也在发生改变(Hayes et al., 2010).有关Aspen FACE森林生态系统的研究也表明, 尽管整个森林林分结构没有受到明显影响, 但敏感型欧洲山杨树种在O3熏蒸7年后几乎从森林冠层中消失(Kubiske et al., 2007).然而, 由于种内及种间复杂的竞争关系, O3对生态系统群落结构的影响有时候并不是统一的, 高浓度O3暴露环境下O3敏感型种群首先达到较高覆盖度的情况也时有发生(Evans & Ashmore, 1992; Barbo et al., 1998).此外, 由于生态系统各营养级之间的交互作用, 高浓度O3影响植物群落多样性的同时也潜在影响植物组织、化学组成、二次代谢产物(如酚苷类化合物、植物源挥发性有机化合物等)释放, 进而改变植物种群周边与之相关的动植物、真菌、细菌和昆虫行为或活动(ICP Vegetation, 2013; Fuhrer et al., 2016).例如, O3暴露使得西黄松(Pinus ponderosa)更易受到小蠹虫的袭击(Dahlsten et al., 1997), 但O3诱导红云杉(Picea rubens)萜烯物质释放增加的同时减少了其上蚜虫幼虫的数量(Cannon, 1990); 而有关欧洲山杨的研究则发现O3暴露降低柳皮苷和特里杨苷含量, 增加了缩合单宁含量, 引起林分中蚜虫和森林天幕毛虫幼虫数量剧增, 但蚜虫捕食者或天敌(如瓢虫、草蛉和蜘蛛)的数量显著减少(Percy et al., 2002; Kopper & Lindroth, 2003). ...
A synthesis of AOT40-based response functions and critical levels of ozone for agricultural and horticultural crops
1
2007
... O3不仅抑制植物生长, 还导致农作物大量减产(Fuhrer, 2009; Ainsworth et al., 2012; Feng et al., 2015a; Singh & Agrawal, 2017).有关O3对作物产量的影响已经在北美、欧洲和亚洲多地的几十种作物上有所研究报道, 包括对O3敏感的小麦、西瓜(Citrullus lanatus)、豆类、棉花(Gossypium hirsutum)、萝卜(Raphanus sativus)、番茄(Lycopersicon esclentum)、洋葱(Allium cepa)、大豆(Glycine max)和莴苣(Lactuca sativa), 对O3适度敏感的甜菜(Beta vulgaris)、马铃薯(Solanum tuberosum)、油菜(Brassica napus)、烟草(Nicotiana tabacum)、水稻、玉米(Zea mays)、葡萄和西兰花(Brassica oleracea), 和对O3耐受的作物大麦(Hordeum vulgare)、李(Prunus salicina)和草莓(Fragaria ananassa)等(Mills et al., 2007; Booker et al., 2009; Feng & Kobayashi, 2009; Avnery et al., 2011; Broberg et al., 2015).通过汇总406个控制实验的数据进行meta分析发现, 与活性炭过滤环境(≤26 μg·kg-1)相比, 当前环境O3浓度(平均值40 μg·kg-1)可导致主要粮食作物(小麦、水稻、大豆和马铃薯等)减产10%左右; 未来地表O3浓度升高情景下(51-71 μg·kg-1), 作物产量将进一步降低10%- 20% (Feng & Kobayashi, 2009).最近, 有关美国、法国、印度、中国和俄罗斯等小麦主产国的模型评估数据显示, 2010-2012年间高浓度O3浓度造成北半球和南半球小麦平均减产9.9%和6.2%, 抵消了因水肥管理或改良品种等措施对其增产所做的努力(Mills et al., 2018).我国这方面的研究起步较晚, 但发展迅速.从20世纪90年代末开始, 学者先后利用OTC和O3-FACE系统研究了地表O3浓度升高对我国主要粮食作物和蔬菜的影响, 涉及品种包括冬小麦、水稻、大豆、玉米、油菜、菠菜(Spinacia oleracea)和青菜(Brassica rapa)等(冯兆忠等, 2018).研究显示, 高浓度O3暴露下农作物均出现不同程度减产, 不同浓度O3对水稻、小麦、大豆等不同品种作物的减产幅度为10%-60% (贾一磊, 2016; 曹嘉晨等, 2017), 对油菜、菠菜和青菜等蔬菜生物产量的降幅为4.0%-89% (白月明等, 2002; 郭建平等, 2003). ...
Evidence of widespread effects of ozone on crops and (semi-) natural vegetation in Europe (1990-2006) in relation to AOT40- and flux-based risk maps
1
2011
... 然而, O3对植物的影响不仅依赖于大气O3浓度同时也依赖于叶片气孔导度(O3的吸收)和植物自身抗氧化能力(Matyssek et al., 2007).近年来, 基于气孔O3吸收量的评估指标PODY (植物毒性O3剂量, 整个生长季单位面积上气孔O3吸收通量超过临界值Y nmol m-2·s-1的积累量)被开发并逐步替代浓度暴露剂量指标, 用于区域或全球O3对农作物和森林树种生长影响的评估(Büker et al., 2015; LRTAR Convention, 2015).目前, 哪个指标更适合区域O3风险评估的争论一直存在, 但欧洲和亚洲地区多项O3对农作物或植被影响的研究均表明: 考虑水分供应的情况下, O3暴露造成的生物量或作物产量的降低与PODY的拟合度比AOTX更好(Mills et al., 2011; Gao et al., 2017), 能更好表征O3对生态系统的影响.整体来看, 各个评估指标的计算方式和所需参数从简单到复杂, 从单一考虑O3浓度到综合考虑环境因子对植物O3吸收通量的影响, 逐渐可以精确评估地表O3污染对生态系统农林生产力的影响(表3). ...
Ozone pollution will compromise efforts to increase global wheat production
3
2018
... O3不仅抑制植物生长, 还导致农作物大量减产(Fuhrer, 2009; Ainsworth et al., 2012; Feng et al., 2015a; Singh & Agrawal, 2017).有关O3对作物产量的影响已经在北美、欧洲和亚洲多地的几十种作物上有所研究报道, 包括对O3敏感的小麦、西瓜(Citrullus lanatus)、豆类、棉花(Gossypium hirsutum)、萝卜(Raphanus sativus)、番茄(Lycopersicon esclentum)、洋葱(Allium cepa)、大豆(Glycine max)和莴苣(Lactuca sativa), 对O3适度敏感的甜菜(Beta vulgaris)、马铃薯(Solanum tuberosum)、油菜(Brassica napus)、烟草(Nicotiana tabacum)、水稻、玉米(Zea mays)、葡萄和西兰花(Brassica oleracea), 和对O3耐受的作物大麦(Hordeum vulgare)、李(Prunus salicina)和草莓(Fragaria ananassa)等(Mills et al., 2007; Booker et al., 2009; Feng & Kobayashi, 2009; Avnery et al., 2011; Broberg et al., 2015).通过汇总406个控制实验的数据进行meta分析发现, 与活性炭过滤环境(≤26 μg·kg-1)相比, 当前环境O3浓度(平均值40 μg·kg-1)可导致主要粮食作物(小麦、水稻、大豆和马铃薯等)减产10%左右; 未来地表O3浓度升高情景下(51-71 μg·kg-1), 作物产量将进一步降低10%- 20% (Feng & Kobayashi, 2009).最近, 有关美国、法国、印度、中国和俄罗斯等小麦主产国的模型评估数据显示, 2010-2012年间高浓度O3浓度造成北半球和南半球小麦平均减产9.9%和6.2%, 抵消了因水肥管理或改良品种等措施对其增产所做的努力(Mills et al., 2018).我国这方面的研究起步较晚, 但发展迅速.从20世纪90年代末开始, 学者先后利用OTC和O3-FACE系统研究了地表O3浓度升高对我国主要粮食作物和蔬菜的影响, 涉及品种包括冬小麦、水稻、大豆、玉米、油菜、菠菜(Spinacia oleracea)和青菜(Brassica rapa)等(冯兆忠等, 2018).研究显示, 高浓度O3暴露下农作物均出现不同程度减产, 不同浓度O3对水稻、小麦、大豆等不同品种作物的减产幅度为10%-60% (贾一磊, 2016; 曹嘉晨等, 2017), 对油菜、菠菜和青菜等蔬菜生物产量的降幅为4.0%-89% (白月明等, 2002; 郭建平等, 2003). ...
... 基于OTCs和O3-FACE平台下单个站点的控制实验可探究O3对植物个体、群落或生态系统的影响, 但将站点或样地尺度的研究外推到区域或全球, 评估O3污染区域风险的研究主要通过响应关系和模型.目前, 其研究领域主要集中在地表O3污染对农作物产量和森林生产力等方面.早在1994年, Chameides等(1994)就利用经济模型初步预测O3浓度升高可能导致2025年全球30%-75%的作物面临产量降低的风险.基于O3前体物排放、化学反应传输模型和M12、AOT40指标的评估显示, 2000年全球O3污染引起全球小麦、玉米和大豆产量平均降低2.2%-15%, 每年因上述作物产量降低所导致的经济损失达110-180亿美元(Avnery et al., 2011).最近, 基于PODY评估指标的研究则表明, O3对小麦造成的产量损失北半球(9.9%)明显高于南半球(6.2%), 平均产量损失最高的地区主要位于北半球暖温带湿润地区(12.5%)、热带湿热(12.5%)和热带潮湿气候区(17.2%)(Mills et al., 2018).在区域尺度上, 基于1990年O3浓度数据, Wang和Mauzerall (2004)利用M7和M12指标评估了O3对中国、日本和韩国主要农作物产量的影响, 结果显示O3污染造成小麦、玉米、棉花减产1%-9%, 大豆减产23%-27%, 造成2.4-35亿美元的经济损失, 在维持现有的污染物排放、管理措施和农业生产技术不变的情况下, 预计到2020年小麦、玉米、棉花减产幅度为2%-15%, 大豆为28%-35%.最近, 利用AOT40指标的研究显示, 2014年当前O3浓度导致中国农作物产量损失7 840万t, 小麦、水稻和玉米相对减产量分别为8.5%-14%、3.9%-15%和2.2%-5.5% (Lin et al., 2018).而只聚焦于长江三角洲地区的研究则发现, 2015年区域O3污染分别造成小麦和水稻减产9.9%-36.1%和7.3%- 23.9% (Zhao et al., 2018).虽然国内也有少量研究建立了PODY与个别树种或作物相对生物量或相对产量之间的通量响应关系(Feng et al., 2012; Hu et al., 2015; Gao et al., 2017), 但并没有研究利用PODY指标评估区域O3污染对农作物产量损失的影响. ...
... (5)开展联网研究, 建立统一评价体系: 目前, 全球覆盖面最广的地表O3监测网络——TOAR (Tropospheric Ozone Assessment Report)已拥有大大小小近万个监测网点(Mills et al., 2018).但考虑到站点分布的不均(80%的监测网点分布在北美和欧洲地区, 亚洲地区的站点占据16%, 但主要集中在日本和韩国, 中国地区只有26个监测网点), 观测频率、时长、年份及评价标准或评价体系的差异(如部分站点为长期研究, 有些则为短期观测; 一些研究采用SUM06或W126作为评估指标, 另一些研究使用AOTX和PODY)(Lefohn et al., 2018), 不同地区、不同学者开展的实验研究结果差异较大, 无法进行统一对比.亚洲地区的研究网点及加入全球的联网实验仍然稀少.因此, 未来亟须扩展联网观测, 建立统一标准为区域或全球O3风险评估提供更具可比性的实测数据. ...
An in vivo analysis of the effect of season-long open-air elevation of ozone to anticipated 2050 levels on photosynthesis in soybean
1
2004
... 在此期间, 尽管社会经济不断发展、实验设备和研究方法不断更新, 但地表O3污染对植物及生态系统影响的研究主要还是以模拟O3浓度升高的控制实验为主.纵观发展历程, 试验设备与手段的改进和更新大致可分为三个阶段(王春乙, 1995): (1) 1970年之前, 研究主要利用封闭式静态或动态气室或室内生长箱进行模拟实验(Heagle et al., 1973). (2) 1973年之后, 美国学者Heagle等(1979)和Mandle (1973)研制开顶式气室(OTC)模拟高浓度O3对植物的损伤和影响机制, 并最终在世界范围内推广, 成为研究地表O3浓度升高对植物及生态系统影响的主流手段.(3)由于OTCs气室内外较大的环境差异, OTCs模拟实验下的熏蒸结果是否适用于大田研究一直备受争议.为解决OTCs内微气候环境带来的影响, 1986年Hendrey和Kimball (1994)提出建立自由空气中气体浓度增加(FACE)系统的想法, 并逐步用于大田实验.由于与外界大气几乎一致的气体流速、温度、光照和风速等条件(Matyssek et al., 2010), FACE系统被普遍认为是当前研究地表O3浓度升高对植物及生态系统影响最为理想的实验平台, 但由于造价昂贵, 技术要求高, 只有少数国家和实验室开展相关研究(有关上述研究方法的优缺点见表2).O3熏蒸的FACE平台仅有10个左右且多数已经停止运行(Paoletti et al., 2017), 主要包括美国威斯康星州Aspen FACE (停止运行; Karnosky et al., 2003), 美国伊利诺伊州Soybean FACE (停止运行; Morgan et al., 2004), 瑞典苏尔草地FACE (停止运行; Volk et al., 2003), 芬兰库奥皮奥FACE (停止运行; Oksanen, 2003), 德国克兰兹堡森林FACE (停止运行; Matyssek et al., 2010), 中国江都水稻(Oryza sativa)/小麦(Triticum aestivum)轮作FACE (停止运行; Tang et al., 2011), 日本札幌森林FACE (仍在运行; Watanabe et al., 2013), 意大利佛罗伦萨3D FACE (仍在运行; Paoletti et al., 2017), 以及刚刚建成的中国延庆唐家堡杨树人工林FACE.由于实验条件和FACE平台规模的限制, 有关地表O3浓度升高对生态系统水平的研究成果主要来自美国Aspen FACE和德国克兰兹堡森林FACE (Grulke & Heath, 2020). ...
A critical review and analysis of the use of exposure- and flux-based ozone indices for predicting vegetation effects
1
2006
... 不管研究方法如何更新, 由于空间和时间上的限制, 模拟地表O3浓度升高的控制实验主要探究高浓度O3对植物损伤的机理, 而区域尺度上探究O3对陆地生态系统的影响却很难通过模拟实验获得.为评价区域O3风险和评估O3对生态系统影响带来的损失, 学者主要通过不同评估指标评估区域、国家或全球尺度上地表O3污染的生态效应.总体来说, 评估指标的研究主要经历了三个阶段, 分别是浓度响应关系、剂量响应关系和通量响应关系评估指标(Musselman et al., 2006).浓度响应关系评估指标主要包括M7/M12 (白天7 h (9:00-16:00) O3浓度平均值或白天12 h (8:00-20:00) O3浓度平均值), 早期曾广泛应用于表征O3浓度暴露与农作物产量损失相关性的研究中(Legge et al., 1995).剂量响应关系评估指标主要有SUM06 (O3小时浓度高于或等于60 μg·kg-1的累积值)(Lefohn & Foley, 1992), W126 (O3小时浓度在规定时段内的加权求和值)和AOTX (整个生长季太阳辐射>50 W·m-2时段内O3小时浓度超过X μg·kg-1的累计值, X普遍设为40 μg·kg-1)(Fuhrer et al., 1997; LRTAR Convention, 2015).剂量响应指标综合考虑O3浓度和暴露时间对植物生长的影响, 在世界范围内得到广泛应用. ...
Responses of selected birch (Betula pendula) clones to ozone change over time
1
2003
... 在此期间, 尽管社会经济不断发展、实验设备和研究方法不断更新, 但地表O3污染对植物及生态系统影响的研究主要还是以模拟O3浓度升高的控制实验为主.纵观发展历程, 试验设备与手段的改进和更新大致可分为三个阶段(王春乙, 1995): (1) 1970年之前, 研究主要利用封闭式静态或动态气室或室内生长箱进行模拟实验(Heagle et al., 1973). (2) 1973年之后, 美国学者Heagle等(1979)和Mandle (1973)研制开顶式气室(OTC)模拟高浓度O3对植物的损伤和影响机制, 并最终在世界范围内推广, 成为研究地表O3浓度升高对植物及生态系统影响的主流手段.(3)由于OTCs气室内外较大的环境差异, OTCs模拟实验下的熏蒸结果是否适用于大田研究一直备受争议.为解决OTCs内微气候环境带来的影响, 1986年Hendrey和Kimball (1994)提出建立自由空气中气体浓度增加(FACE)系统的想法, 并逐步用于大田实验.由于与外界大气几乎一致的气体流速、温度、光照和风速等条件(Matyssek et al., 2010), FACE系统被普遍认为是当前研究地表O3浓度升高对植物及生态系统影响最为理想的实验平台, 但由于造价昂贵, 技术要求高, 只有少数国家和实验室开展相关研究(有关上述研究方法的优缺点见表2).O3熏蒸的FACE平台仅有10个左右且多数已经停止运行(Paoletti et al., 2017), 主要包括美国威斯康星州Aspen FACE (停止运行; Karnosky et al., 2003), 美国伊利诺伊州Soybean FACE (停止运行; Morgan et al., 2004), 瑞典苏尔草地FACE (停止运行; Volk et al., 2003), 芬兰库奥皮奥FACE (停止运行; Oksanen, 2003), 德国克兰兹堡森林FACE (停止运行; Matyssek et al., 2010), 中国江都水稻(Oryza sativa)/小麦(Triticum aestivum)轮作FACE (停止运行; Tang et al., 2011), 日本札幌森林FACE (仍在运行; Watanabe et al., 2013), 意大利佛罗伦萨3D FACE (仍在运行; Paoletti et al., 2017), 以及刚刚建成的中国延庆唐家堡杨树人工林FACE.由于实验条件和FACE平台规模的限制, 有关地表O3浓度升高对生态系统水平的研究成果主要来自美国Aspen FACE和德国克兰兹堡森林FACE (Grulke & Heath, 2020). ...
Interactive effects of nitrogen deposition, tropospheric ozone, elevated CO2 and land use history on the carbon dynamics of northern hardwood forests
2
2002
... 除单因子O3胁迫外, 由于全球气候变化和极端气候常态化, 植物经常受到O3和其他环境因子交互作用的影响(如O3与CO2、升温、干旱或氮沉降等)(Schmadel-Hagebölling et al., 1998; Ollinger et al., 2002; Eltayeb et al., 2006; Ghimire et al., 2018).然而, 植物应对O3与其他因子交互作用的响应并不是两者或单独作用的简单叠加, 往往取决于植物种类、生长时期、对特定环境因子的敏感性、胁迫浓度及时间, 以及具体的微气候条件(列淦文和薛立, 2014).一方面, 通过诱导气孔关闭、限制叶片O3吸收(CO2浓度升高/干旱胁迫), 或提高碳同化速率、促进植物光合作用(CO2浓度升高/氮增加), CO2浓度升高、干旱胁迫和氮沉降经常被报道可增强植物抵御O3的能力, 减缓O3伤害(白月明等, 2005; Handley & Grulke, 2008; Gao et al., 2017).另一方面, 与之相反, 也有研究发现长期高浓度CO2或干旱降低气孔导度的同时, 会引起植物蒸腾速率、光合产物合成和积累的降低(McDonald et al., 2002; Pollastrini et al., 2010); 而超过植物承载阈值的高氮负荷也会对植物的光合生理活性产生负效应(Yamaguchi et al., 2010), 进而加重植物在复合环境下的碳损失、水分蒸散损失和自由基积累, 对植物产生协同伤害(Karnosky et al., 2005; 列淦文和薛立, 2014).高峰(2018)认为O3与其他环境胁迫因子出现的先后次序也会影响植物对复合作用的响应. ...
... (3)开展O3与其他环境因子复合研究: 全球气候变化、极端气候常态化和空气污染加重使两个或两个以上环境因子的交互作用大大增加, 复合作用下的实验条件也更接近植物生长的自然环境.现有的研究多集中在O3与CO2对植物或生态系统的共同影响, 并发现O3污染引起的负面作用经常被高浓度CO2对植物叶片光合作用或个体生物量累积的“肥效”所削弱(Ollinger et al., 2002).然而, 目前还无法确定这种结果是否与生态系统水平的响应相匹配(Grulke & Heath, 2020).植物应对O3胁迫的响应与土壤水分、氮有效性等因素密切相关(列淦文和薛立, 2014; 高峰, 2018).控制实验下的研究也发现, O3浓度的升高可能削弱植物对干旱或其他环境胁迫的抵抗力, 使生态系统更易受到病虫害的侵袭(Grulke et al., 2009).但是迄今为止, 国内外大部分研究多集中在温带农作物和森林树种上, 较少涉及亚热带地区乡土树种.由于生态系统类型差异, 各级营养交互及种间竞争的复杂性, 不同生态系统下O3与生物及非生物因子之间的多重交互作用机制还需进一步探究.此外, 已有研究多针对植物地上部分, 对植物地下部分及其功能变化与根际过程相互影响的机制、土壤关键过程研究较少(列淦文和薛立, 2014).因此, 在开展O3单因子实验研究的同时, 亟须考虑多个因子(如O3、CO2、水分和氮沉降等)共存对整个生态系统结构和功能的复合影响研究. ...
Ozone and urban forests in Italy
1
2009
... (6)探究地表O3污染的生态控制措施: 欧洲和北美等发达国家治理大气污染的经验告诉我们, 严格的O3前体物NOx和挥发性有机化合物(VOCs)减排措施是降低地表O3浓度的有效手段(Feng et al., 2015b).然而, 以完全牺牲经济发展为代价的环境保护措施一般并不被发展中国家所接受.事实上, 自然植被与地表O3存在复杂的交互作用.一方面, 植物释放的VOCs有助于地表O3的形成; 另一方面, 植物在进行光合作用的同时也可以通过气孔、非气孔途径吸收或沉降大气O3 (Paoletti, 2009).考虑到植被VOCs释放和O3移除能力都具有树种特异性, 因此, 在一定条件下通过合理的树种筛选达到减少大气O3污染的目的将是完全可行的(Sicard et al., 2018).另外, 除植物自身遗传基因之外, 植物对O3胁迫的响应受土壤水分和营养元素如氮可利用性的影响(列淦文和薛立, 2014; 高峰, 2018).例如古巴农民通过提前1-2天灌溉的方式预防夏季高浓度O3对莴苣和烟草叶面的损伤(ICP Vegetation, 2011); 马来西亚研究者通过氮肥管理, 减轻O3污染对热带Elaeis guineensis产量和质量的伤害(Hewitt et al., 2009).因此通过合理的田间水肥管理措施(如改变灌溉模式、施肥剂量和方式)调控植物生长, 增强植物对O3的抗性, 进而减轻O3污染的生态效应是未来大气O3污染防治研究中值得思考的问题. ...
A new-generation 3D ozone FACE (Free Air Controlled Exposure)
2
2017
... 在此期间, 尽管社会经济不断发展、实验设备和研究方法不断更新, 但地表O3污染对植物及生态系统影响的研究主要还是以模拟O3浓度升高的控制实验为主.纵观发展历程, 试验设备与手段的改进和更新大致可分为三个阶段(王春乙, 1995): (1) 1970年之前, 研究主要利用封闭式静态或动态气室或室内生长箱进行模拟实验(Heagle et al., 1973). (2) 1973年之后, 美国学者Heagle等(1979)和Mandle (1973)研制开顶式气室(OTC)模拟高浓度O3对植物的损伤和影响机制, 并最终在世界范围内推广, 成为研究地表O3浓度升高对植物及生态系统影响的主流手段.(3)由于OTCs气室内外较大的环境差异, OTCs模拟实验下的熏蒸结果是否适用于大田研究一直备受争议.为解决OTCs内微气候环境带来的影响, 1986年Hendrey和Kimball (1994)提出建立自由空气中气体浓度增加(FACE)系统的想法, 并逐步用于大田实验.由于与外界大气几乎一致的气体流速、温度、光照和风速等条件(Matyssek et al., 2010), FACE系统被普遍认为是当前研究地表O3浓度升高对植物及生态系统影响最为理想的实验平台, 但由于造价昂贵, 技术要求高, 只有少数国家和实验室开展相关研究(有关上述研究方法的优缺点见表2).O3熏蒸的FACE平台仅有10个左右且多数已经停止运行(Paoletti et al., 2017), 主要包括美国威斯康星州Aspen FACE (停止运行; Karnosky et al., 2003), 美国伊利诺伊州Soybean FACE (停止运行; Morgan et al., 2004), 瑞典苏尔草地FACE (停止运行; Volk et al., 2003), 芬兰库奥皮奥FACE (停止运行; Oksanen, 2003), 德国克兰兹堡森林FACE (停止运行; Matyssek et al., 2010), 中国江都水稻(Oryza sativa)/小麦(Triticum aestivum)轮作FACE (停止运行; Tang et al., 2011), 日本札幌森林FACE (仍在运行; Watanabe et al., 2013), 意大利佛罗伦萨3D FACE (仍在运行; Paoletti et al., 2017), 以及刚刚建成的中国延庆唐家堡杨树人工林FACE.由于实验条件和FACE平台规模的限制, 有关地表O3浓度升高对生态系统水平的研究成果主要来自美国Aspen FACE和德国克兰兹堡森林FACE (Grulke & Heath, 2020). ...
... ), 意大利佛罗伦萨3D FACE (仍在运行; Paoletti et al., 2017), 以及刚刚建成的中国延庆唐家堡杨树人工林FACE.由于实验条件和FACE平台规模的限制, 有关地表O3浓度升高对生态系统水平的研究成果主要来自美国Aspen FACE和德国克兰兹堡森林FACE (Grulke & Heath, 2020). ...
Ozone-induced reduction in quantity and quality of two potato cultivars
1
1984
... (4)关注O3对作物品质及食品安全的危害: 世界上主要粮食作物要么对O3敏感(小麦、大豆等), 要么对O3适度敏感(玉米、水稻和马铃薯等).到2050年, 世界人口预计增加到90亿, 粮食供应需增加至少50%才能满足人口增长的需求, 如何保证粮食供应安全将成为21世纪最重要的一个挑战(Tai et al., 2014).目前为止, 除作物产量之外, 有关地表O3浓度升高对作物品质的影响研究多集中在蔬菜作物或粮食作物可食部位糖分、蛋白质和淀粉含量等食用口感方面(Booker et al., 2009), 很少涉及其背后对人体健康的直接影响.随着生活水平的提高, 人们更加关注食品的营养与安全(如微量元素含量或健康风险等), 而长期高浓度O3暴露可改变粮食作物籽粒微量元素及营养物质含量(Booker et al., 2009; 冯兆忠等, 2018).此外, 也有研究显示O3暴露导致马铃薯的糖苷生物碱含量增加, 糖苷生物碱不仅导致味苦而且浓度较高时具有毒性(Pell & Pearson, 1984).因此, 在全球环境变化大背景下, 长期食用暴露在O3污染下的粮食作物是否影响人体营养平衡或危及健康值得关注.未来的研究应关注高浓度O3对农作物外观品质(如籽粒形态、发芽率和成活率等)、加工特性、口感(如黏弹性、柔软性等)、烹饪特性和营养特性等外在和内在价值的综合影响. ...
What about the detoxification mechanisms underlying ozone sensitivity in Liriodendron tulipifera
1
2018
... 气孔防御机制失效后, 进入细胞的O3将诱导叶片内部抗氧化系统启动解毒和修复的非气孔防御响应(冯兆忠等, 2018).非气孔防御响应主要是指“质外体-共质体系统”内一些抗氧化组分对O3解毒和修复的防御过程.具体来说, O3进入植物叶片的细胞间隙后会溶解在细胞壁的结合水里, 经过一系列的化学反应形成活性氧分子(ROS), ROS能够破坏细胞壁并与细胞膜上的不饱和脂肪酸发生反应, 从而对叶片造成损伤(Tausz et al., 2007; Pellegrini et al., 2018).植物质外体的抗氧化物质如还原性抗坏血酸(ASC)、抗氧化酶(如超氧化物歧化酶)可与ROS反应进行初级解毒, 成为抵御O3的第二道防线(Kangasjärvi et al., 2005; Dai et al., 2019), 而剩余的ROS会经过质膜到达共质体, 此时共质体内抗氧化成分如过氧化物酶(POD)、过氧化氢酶(CAT)、抗坏血酸过氧化物酶(APX)及抗氧化剂(如酚类化合物-谷胱甘肽(GSH)、脯氨酸和多胺等)等抗氧化酶类物质被诱导作为次级解毒响应机制参与诸如“抗坏血酸-谷胱甘肽(ASA-GSH)循环”等氧化还原过程, 进而对ROS进行进一步解毒与消除(Luwe, 1996).气孔闭合除了会减少O3的吸收, 也会抑制CO2吸收和水汽交换, 而非气孔防御响应中的抗氧化系统解毒过程也会消耗大量的能量并对光合反应系统造成不同程度的损伤, 降低羧化速率和电子传递速率, 进而抑制光合和蒸腾作用, 损害植物生长(Gao et al., 2016).然而, 最新研究表明持续整个生长季的长期O3暴露会影响植物叶片的气孔密度、开度和大小等表观属性(李品等, 2018), 而O3从气孔进入到细胞间隙后, 也可以通过影响植物的叶肉导度降低植物的光合能力(Xu et al., 2019).此外, 有关29种落叶和常绿树种应对O3胁迫的研究还发现不同木本植物对O3响应的差异主要来自植物功能叶片比叶质量的大小(即叶片比叶质量越大的树种对O3越不敏感), 与气孔导度无明显相关性(Li et al., 2016). ...
Altered performance of forest pests under CO2- and O3-enriched atmospheres
1
2002
... 植物长期过量摄取O3还可以引起物种组成、冠层结构改变, 影响生态系统种群均匀度和丰富度, 威胁生态系统多样性(Fuhrer et al., 2016; Bergmann et al., 2017), 最终影响生态系统碳、氮和水循环(McLaughlin et al., 2007; Sun et al., 2012; Grulke & Heath, 2020).基于温室或OTCs控制实验下有关草本植物的多项研究都发现, 随着O3暴露时间的延长, 对O3相对敏感的物种/种群将会被对O3相对耐受的物种/种群替代(Fuhrer et al., 1994; Hayes et al., 2009), 非禾本科杂草与禾本科杂草的比例也在发生改变(Hayes et al., 2010).有关Aspen FACE森林生态系统的研究也表明, 尽管整个森林林分结构没有受到明显影响, 但敏感型欧洲山杨树种在O3熏蒸7年后几乎从森林冠层中消失(Kubiske et al., 2007).然而, 由于种内及种间复杂的竞争关系, O3对生态系统群落结构的影响有时候并不是统一的, 高浓度O3暴露环境下O3敏感型种群首先达到较高覆盖度的情况也时有发生(Evans & Ashmore, 1992; Barbo et al., 1998).此外, 由于生态系统各营养级之间的交互作用, 高浓度O3影响植物群落多样性的同时也潜在影响植物组织、化学组成、二次代谢产物(如酚苷类化合物、植物源挥发性有机化合物等)释放, 进而改变植物种群周边与之相关的动植物、真菌、细菌和昆虫行为或活动(ICP Vegetation, 2013; Fuhrer et al., 2016).例如, O3暴露使得西黄松(Pinus ponderosa)更易受到小蠹虫的袭击(Dahlsten et al., 1997), 但O3诱导红云杉(Picea rubens)萜烯物质释放增加的同时减少了其上蚜虫幼虫的数量(Cannon, 1990); 而有关欧洲山杨的研究则发现O3暴露降低柳皮苷和特里杨苷含量, 增加了缩合单宁含量, 引起林分中蚜虫和森林天幕毛虫幼虫数量剧增, 但蚜虫捕食者或天敌(如瓢虫、草蛉和蜘蛛)的数量显著减少(Percy et al., 2002; Kopper & Lindroth, 2003). ...
Growth and physiological responses to ozone and mild drought stress of tree species with different ecological requirements
1
2010
... 除单因子O3胁迫外, 由于全球气候变化和极端气候常态化, 植物经常受到O3和其他环境因子交互作用的影响(如O3与CO2、升温、干旱或氮沉降等)(Schmadel-Hagebölling et al., 1998; Ollinger et al., 2002; Eltayeb et al., 2006; Ghimire et al., 2018).然而, 植物应对O3与其他因子交互作用的响应并不是两者或单独作用的简单叠加, 往往取决于植物种类、生长时期、对特定环境因子的敏感性、胁迫浓度及时间, 以及具体的微气候条件(列淦文和薛立, 2014).一方面, 通过诱导气孔关闭、限制叶片O3吸收(CO2浓度升高/干旱胁迫), 或提高碳同化速率、促进植物光合作用(CO2浓度升高/氮增加), CO2浓度升高、干旱胁迫和氮沉降经常被报道可增强植物抵御O3的能力, 减缓O3伤害(白月明等, 2005; Handley & Grulke, 2008; Gao et al., 2017).另一方面, 与之相反, 也有研究发现长期高浓度CO2或干旱降低气孔导度的同时, 会引起植物蒸腾速率、光合产物合成和积累的降低(McDonald et al., 2002; Pollastrini et al., 2010); 而超过植物承载阈值的高氮负荷也会对植物的光合生理活性产生负效应(Yamaguchi et al., 2010), 进而加重植物在复合环境下的碳损失、水分蒸散损失和自由基积累, 对植物产生协同伤害(Karnosky et al., 2005; 列淦文和薛立, 2014).高峰(2018)认为O3与其他环境胁迫因子出现的先后次序也会影响植物对复合作用的响应. ...
Tree and stand growth of mature Norway spruce and European beech under long-term ozone fumigation
1
2010
... 有关自然环境下O3浓度升高对陆地生态系统影响的研究较为稀缺.但是现有开放式O3浓度增强系统(O3-FACE)观测平台下的长期研究也有着相似结果, Aspen FACE实验发现, O3浓度升高3-4年后, 欧洲山杨(Populus tremuloides)纯林、欧洲山杨-纸皮桦(Betula papyrifera)和欧洲山杨-糖枫(Acer saccharum)混交林的总生物量分别降低23%、13%和14% (Karnosky et al., 2003, 2005); 虽然在最初的7年内生物量的分配格局并未受O3影响(King et al., 2005), 但在实验11年后, 敏感型欧洲山杨几乎从森林冠层中消失(Kubiske et al., 2007).德国克兰兹堡郁闭冠层的成熟挪威云杉(Picea abies)和欧洲山毛榉(Fagus sylvatica)混交林(60年)对O3浓度升高长期响应的研究也表明, 8年O3暴露导致欧洲山毛榉的树干生产力降低了44% (Pretzsch et al., 2010).目前, 国内还没有研究涉及O3-FACE平台下O3浓度升高对森林生态系统碳汇能力的影响. ...
Quantifying plant response to ozone: a unifying theory
1
1987
... 地表O3浓度升高对植物光合作用、生长和生物量积累、物质分配等过程的连锁负效应导致陆地生态系统生产力和固碳能力降低(Reich, 1987; Wittig et al., 2009; Ainsworth et al., 2012), 削弱了自然生态系统的碳汇能力(Felzer et al., 2005; Sitch et al., 2007; Collins et al., 2010).基于大量OTCs控制实验下北半球中纬度北方温带树种的meta分析发现, O3浓度升高导致树木的净光合作用和总生物量分别降低11%和7% (Wittig et al., 2009).最近, 有关中国热带、亚热带和温带树种的meta分析则发现, O3浓度升高导致中国木本植物的净光合作用和总生物量分别降低28%和14% (Li et al., 2017). ...
Impacts of tropospheric ozone and climate change on net primary productivity and net carbon exchange of China’s forest ecosystems
1
2011
... 区域尺度上, 地表O3污染对陆地生态系统固碳能力的影响主要依靠模式算法的估算.早在20世纪90年代, 基于AOT40指标的研究就发现高浓度O3降低美国东北部阔叶林净初级生产力(NPP) 3%-16%, 对地上生物量的降低每年为0-10% (Chappelka & Samuelson, 1998).另有研究表明O3污染造成美国植被碳固定降低18-38 Tg·a-1, NPP降低至少9 g·m-2·a-1, 且高浓度O3导致植物-土壤系统碳积累和固定能力下降(Felzer et al., 2004).此外, 运用自主开发的基于过程的陆地生态系统动态模型(DLEM), 国内学者发现近40年来O3浓度升高使中国森林净固碳量减少7.7%, 且不同的森林类型降低幅度不同: 如北方落叶阔叶林降低21.8%, 北方落叶针叶林降低5.2%, 温带落叶阔叶林降低21.1%, 温带常绿阔叶林降低7.6%, 温带常绿针叶林降低0.4%, 温带落叶针叶林降低6.2%, 热带落叶阔叶林降低43.1%, 热带常绿阔叶林降低9.6% (Ren et al., 2011).最近, Yue等(2017)用地球系统模式(NASA ModelE2-YIBs)得出O3污染造成中国陆地植被NPP降低600 Tg·a-1, 且对于不同功能类型的植物, O3对总初级生产力(GPP)的降低都随着O3浓度的升高而加大.总体来看, 模型驱动的结果与大量控制实验的发现相吻合, 均表明O3污染在不同尺度都对自然生态系统碳固定有显著的负效应, 未来O3浓度升高有可能抵消因大气CO2浓度升高所增加的GPP (18%-34%)(Sitch et al., 2007). ...
Air pollution with relation to agronomic crops: V. Oxidant stipple of grape
1
1958
... 1958年美国加利福尼亚Richards等在期刊文献中首次报道高浓度地表O3引起加利福尼亚城市周边葡萄(Vitis vinifera)叶片出现坏死病斑.随后, 高浓度O3对植物的毒害作用逐渐受到社会的关注, 相关研究也逐步活跃起来.历经几十年发展, 其研究对象逐渐从作物、草地转向森林树木, 从地上生态系统逐步向地下生态系统转移; 从探讨单一物种在O3熏蒸下的响应, 转向多因子或多物种种内或种间关系; 研究尺度也从叶片光合生理, 上升到整株生物量, 并逐渐转向生态系统碳氮循环; 从短期实验向长期可控实验发展. ...
Smog effects and chemical analysis of the Los Angeles atmosphere
1
1956
... 地表O3的发现最早可追溯到1839年(Leeds, 1880), 但直到1945年“洛杉矶烟雾”爆发后, 地表O3生态效应的研究才逐渐进入学者的视野(Rogers et al., 1956).经过近百年的发展, 涉及地表O3对植物、土壤及生态系统影响的研究, 已经遍及世界上大多数国家, 发表了2 500多篇期刊论文和十余本英文专著.到目前为止, O3表观伤害症状的描述及症状等级的鉴定(如叶片萎黄、褪绿和坏死等典型症状分类), O3伤害机理的探究(如O3降低光合速率的气孔与非气孔因素等), O3对植物叶片(光合作用、气孔导度及抗氧化系统等), 个体(生物量、产量或品质), 种群(种间关系、群落构成)及生态系统(生产力和固碳能力等)的影响, 及O3污染防治对策的探讨(前体物减排措施、化学防护剂的应用和作物栽培管理模式的筛选)等方面都进行了大量相关研究(Ashmore, 2005; Grulke & Heath, 2020).然而地表O3浓度升高对陆地生态系统水平碳、氮和水循环、营养交互、群落更替及地下生态系统动态变化过程(如根系功能变化、根际过程机制、土壤微生物活性及反馈等)方面的研究仍鲜有报道(Fuhrer et al., 2016).通过系统梳理和总结目前国内外研究成果, 本文对地表O3浓度升高对生态系统影响这一领域的研究历史、研究方法、评估指标、重大进展和存在的主要问题进行了回顾分析, 并对未来研究进行了展望. ...
Elevated ozone deteriorates grain quality of Japonica Rice cv. Koshihikari, even if it does not cause yield reduction
1
2016
... 相对于农作物产量, O3对粮食品质影响的研究集中在几种主要农作物(如小麦、水稻和大豆)(冯兆忠等, 2018).基于北美、欧洲和亚洲已有的O3对小麦影响的meta分析发现, O3显著降低小麦淀粉浓度, 但显著增加蛋白质和一些营养元素浓度(如钾、镁、钙、磷、锌、锰和铜)(Broberg et al., 2015).印度北部热带地区有关小麦的研究则显示, O3显著降低小麦籽粒糖含量, 但没有改变籽粒蛋白质含量(Tomer et al., 2015).有关水稻的研究还发现, O3增加粳稻直链淀粉含量, 但对籼稻直链淀粉含量无显著影响(Sawada et al., 2016).国内研究则表明, 尽管高浓度O3显著降低冬小麦面粉中支链淀粉和淀粉含量, 但是增加直链淀粉含量(Zhang et al., 2013).O3显著降低水稻稻米直链淀粉浓度、分解率, 增加黏度、起糊温度、蛋白质和微量矿物质浓度, 但蛋白质总量没有改变(Wang et al., 2012).此外, 研究还发现O3熏蒸显著降低了小麦籽粒出粉率和硬度, 但增加冬小麦面粉峰值黏度、低谷黏度、最终黏度和反弹值(贾一磊, 2016). ...
Short term effects of ozone on the plant-rhizosphere-bulk soil system of young beech trees
1
2005
... 此外, 虽然地下生态系统并不是直接暴露在O3环境下, 但大量证据表明O3对地上生态系统的影响将会转移至地下(Andersen, 2003; Lindroth, 2010).如高浓度O3环境下, 植物分配到根的生物量将会大大减少, 基于100多组实验数据的meta分析研究表明O3暴露导致生物量分配到地下的量平均减少5.6% (Grantz et al., 2006).另外, 凋落物分解是生态系统营养循环的关键过程.通过影响凋落物的数量、质量和分解速率, 高浓度地表O3会间接改变土壤微生物群落活性、组成和结构, 土壤化学属性及土壤营养循环(Larson et al., 2002; Fuhrer et al., 2016).然而, 比较遗憾的是, 目前有关O3对地下生态过程影响的研究仅集中在农田生态系统, 仅有少部分研究涉及森林土壤(主要来自Aspen FACE)或草地生态系统(ICP Vegetation, 2013).而且, 由于凋落物种类、化学成分、分解者种群、土壤和大气环境条件的不同, 高浓度O3对凋落物分解动态的影响还存在很大争议(Gessner et al., 2010; Fuhrer et al., 2016; Grulke & Heath, 2020).例如, 研究发现O3对欧洲白桦(Betula pendula)叶片凋落物质量和损失率有明显影响(Kasurinen et al., 2006, 2007), 但不影响欧洲山毛榉凋落物的分解速率(Schloter et al., 2005).有关地表O3浓度升高对地下生态系统结构和功能的影响还有待进一步探究. ...
The combined effects of CO2, ozone and drought on rubisco and nitrogen metabolism of young oak trees (Quercus petraea)—A phytotron study
1
1998
... 除单因子O3胁迫外, 由于全球气候变化和极端气候常态化, 植物经常受到O3和其他环境因子交互作用的影响(如O3与CO2、升温、干旱或氮沉降等)(Schmadel-Hagebölling et al., 1998; Ollinger et al., 2002; Eltayeb et al., 2006; Ghimire et al., 2018).然而, 植物应对O3与其他因子交互作用的响应并不是两者或单独作用的简单叠加, 往往取决于植物种类、生长时期、对特定环境因子的敏感性、胁迫浓度及时间, 以及具体的微气候条件(列淦文和薛立, 2014).一方面, 通过诱导气孔关闭、限制叶片O3吸收(CO2浓度升高/干旱胁迫), 或提高碳同化速率、促进植物光合作用(CO2浓度升高/氮增加), CO2浓度升高、干旱胁迫和氮沉降经常被报道可增强植物抵御O3的能力, 减缓O3伤害(白月明等, 2005; Handley & Grulke, 2008; Gao et al., 2017).另一方面, 与之相反, 也有研究发现长期高浓度CO2或干旱降低气孔导度的同时, 会引起植物蒸腾速率、光合产物合成和积累的降低(McDonald et al., 2002; Pollastrini et al., 2010); 而超过植物承载阈值的高氮负荷也会对植物的光合生理活性产生负效应(Yamaguchi et al., 2010), 进而加重植物在复合环境下的碳损失、水分蒸散损失和自由基积累, 对植物产生协同伤害(Karnosky et al., 2005; 列淦文和薛立, 2014).高峰(2018)认为O3与其他环境胁迫因子出现的先后次序也会影响植物对复合作用的响应. ...
Should we see urban trees as effective solutions to reduce increasing ozone levels in cities?
1
2018
... (6)探究地表O3污染的生态控制措施: 欧洲和北美等发达国家治理大气污染的经验告诉我们, 严格的O3前体物NOx和挥发性有机化合物(VOCs)减排措施是降低地表O3浓度的有效手段(Feng et al., 2015b).然而, 以完全牺牲经济发展为代价的环境保护措施一般并不被发展中国家所接受.事实上, 自然植被与地表O3存在复杂的交互作用.一方面, 植物释放的VOCs有助于地表O3的形成; 另一方面, 植物在进行光合作用的同时也可以通过气孔、非气孔途径吸收或沉降大气O3 (Paoletti, 2009).考虑到植被VOCs释放和O3移除能力都具有树种特异性, 因此, 在一定条件下通过合理的树种筛选达到减少大气O3污染的目的将是完全可行的(Sicard et al., 2018).另外, 除植物自身遗传基因之外, 植物对O3胁迫的响应受土壤水分和营养元素如氮可利用性的影响(列淦文和薛立, 2014; 高峰, 2018).例如古巴农民通过提前1-2天灌溉的方式预防夏季高浓度O3对莴苣和烟草叶面的损伤(ICP Vegetation, 2011); 马来西亚研究者通过氮肥管理, 减轻O3污染对热带Elaeis guineensis产量和质量的伤害(Hewitt et al., 2009).因此通过合理的田间水肥管理措施(如改变灌溉模式、施肥剂量和方式)调控植物生长, 增强植物对O3的抗性, 进而减轻O3污染的生态效应是未来大气O3污染防治研究中值得思考的问题. ...
Tropospheric ozone pollution in India: effects on crop yield and product quality
1
2017
... O3不仅抑制植物生长, 还导致农作物大量减产(Fuhrer, 2009; Ainsworth et al., 2012; Feng et al., 2015a; Singh & Agrawal, 2017).有关O3对作物产量的影响已经在北美、欧洲和亚洲多地的几十种作物上有所研究报道, 包括对O3敏感的小麦、西瓜(Citrullus lanatus)、豆类、棉花(Gossypium hirsutum)、萝卜(Raphanus sativus)、番茄(Lycopersicon esclentum)、洋葱(Allium cepa)、大豆(Glycine max)和莴苣(Lactuca sativa), 对O3适度敏感的甜菜(Beta vulgaris)、马铃薯(Solanum tuberosum)、油菜(Brassica napus)、烟草(Nicotiana tabacum)、水稻、玉米(Zea mays)、葡萄和西兰花(Brassica oleracea), 和对O3耐受的作物大麦(Hordeum vulgare)、李(Prunus salicina)和草莓(Fragaria ananassa)等(Mills et al., 2007; Booker et al., 2009; Feng & Kobayashi, 2009; Avnery et al., 2011; Broberg et al., 2015).通过汇总406个控制实验的数据进行meta分析发现, 与活性炭过滤环境(≤26 μg·kg-1)相比, 当前环境O3浓度(平均值40 μg·kg-1)可导致主要粮食作物(小麦、水稻、大豆和马铃薯等)减产10%左右; 未来地表O3浓度升高情景下(51-71 μg·kg-1), 作物产量将进一步降低10%- 20% (Feng & Kobayashi, 2009).最近, 有关美国、法国、印度、中国和俄罗斯等小麦主产国的模型评估数据显示, 2010-2012年间高浓度O3浓度造成北半球和南半球小麦平均减产9.9%和6.2%, 抵消了因水肥管理或改良品种等措施对其增产所做的努力(Mills et al., 2018).我国这方面的研究起步较晚, 但发展迅速.从20世纪90年代末开始, 学者先后利用OTC和O3-FACE系统研究了地表O3浓度升高对我国主要粮食作物和蔬菜的影响, 涉及品种包括冬小麦、水稻、大豆、玉米、油菜、菠菜(Spinacia oleracea)和青菜(Brassica rapa)等(冯兆忠等, 2018).研究显示, 高浓度O3暴露下农作物均出现不同程度减产, 不同浓度O3对水稻、小麦、大豆等不同品种作物的减产幅度为10%-60% (贾一磊, 2016; 曹嘉晨等, 2017), 对油菜、菠菜和青菜等蔬菜生物产量的降幅为4.0%-89% (白月明等, 2002; 郭建平等, 2003). ...
Indirect radiative forcing of climate change through ozone effects on the land-carbon sink
2
2007
... 地表O3浓度升高对植物光合作用、生长和生物量积累、物质分配等过程的连锁负效应导致陆地生态系统生产力和固碳能力降低(Reich, 1987; Wittig et al., 2009; Ainsworth et al., 2012), 削弱了自然生态系统的碳汇能力(Felzer et al., 2005; Sitch et al., 2007; Collins et al., 2010).基于大量OTCs控制实验下北半球中纬度北方温带树种的meta分析发现, O3浓度升高导致树木的净光合作用和总生物量分别降低11%和7% (Wittig et al., 2009).最近, 有关中国热带、亚热带和温带树种的meta分析则发现, O3浓度升高导致中国木本植物的净光合作用和总生物量分别降低28%和14% (Li et al., 2017). ...
... 区域尺度上, 地表O3污染对陆地生态系统固碳能力的影响主要依靠模式算法的估算.早在20世纪90年代, 基于AOT40指标的研究就发现高浓度O3降低美国东北部阔叶林净初级生产力(NPP) 3%-16%, 对地上生物量的降低每年为0-10% (Chappelka & Samuelson, 1998).另有研究表明O3污染造成美国植被碳固定降低18-38 Tg·a-1, NPP降低至少9 g·m-2·a-1, 且高浓度O3导致植物-土壤系统碳积累和固定能力下降(Felzer et al., 2004).此外, 运用自主开发的基于过程的陆地生态系统动态模型(DLEM), 国内学者发现近40年来O3浓度升高使中国森林净固碳量减少7.7%, 且不同的森林类型降低幅度不同: 如北方落叶阔叶林降低21.8%, 北方落叶针叶林降低5.2%, 温带落叶阔叶林降低21.1%, 温带常绿阔叶林降低7.6%, 温带常绿针叶林降低0.4%, 温带落叶针叶林降低6.2%, 热带落叶阔叶林降低43.1%, 热带常绿阔叶林降低9.6% (Ren et al., 2011).最近, Yue等(2017)用地球系统模式(NASA ModelE2-YIBs)得出O3污染造成中国陆地植被NPP降低600 Tg·a-1, 且对于不同功能类型的植物, O3对总初级生产力(GPP)的降低都随着O3浓度的升高而加大.总体来看, 模型驱动的结果与大量控制实验的发现相吻合, 均表明O3污染在不同尺度都对自然生态系统碳固定有显著的负效应, 未来O3浓度升高有可能抵消因大气CO2浓度升高所增加的GPP (18%-34%)(Sitch et al., 2007). ...
Interactive influences of ozone and climate on streamflow of forested watersheds
1
2012
... 植物长期过量摄取O3还可以引起物种组成、冠层结构改变, 影响生态系统种群均匀度和丰富度, 威胁生态系统多样性(Fuhrer et al., 2016; Bergmann et al., 2017), 最终影响生态系统碳、氮和水循环(McLaughlin et al., 2007; Sun et al., 2012; Grulke & Heath, 2020).基于温室或OTCs控制实验下有关草本植物的多项研究都发现, 随着O3暴露时间的延长, 对O3相对敏感的物种/种群将会被对O3相对耐受的物种/种群替代(Fuhrer et al., 1994; Hayes et al., 2009), 非禾本科杂草与禾本科杂草的比例也在发生改变(Hayes et al., 2010).有关Aspen FACE森林生态系统的研究也表明, 尽管整个森林林分结构没有受到明显影响, 但敏感型欧洲山杨树种在O3熏蒸7年后几乎从森林冠层中消失(Kubiske et al., 2007).然而, 由于种内及种间复杂的竞争关系, O3对生态系统群落结构的影响有时候并不是统一的, 高浓度O3暴露环境下O3敏感型种群首先达到较高覆盖度的情况也时有发生(Evans & Ashmore, 1992; Barbo et al., 1998).此外, 由于生态系统各营养级之间的交互作用, 高浓度O3影响植物群落多样性的同时也潜在影响植物组织、化学组成、二次代谢产物(如酚苷类化合物、植物源挥发性有机化合物等)释放, 进而改变植物种群周边与之相关的动植物、真菌、细菌和昆虫行为或活动(ICP Vegetation, 2013; Fuhrer et al., 2016).例如, O3暴露使得西黄松(Pinus ponderosa)更易受到小蠹虫的袭击(Dahlsten et al., 1997), 但O3诱导红云杉(Picea rubens)萜烯物质释放增加的同时减少了其上蚜虫幼虫的数量(Cannon, 1990); 而有关欧洲山杨的研究则发现O3暴露降低柳皮苷和特里杨苷含量, 增加了缩合单宁含量, 引起林分中蚜虫和森林天幕毛虫幼虫数量剧增, 但蚜虫捕食者或天敌(如瓢虫、草蛉和蜘蛛)的数量显著减少(Percy et al., 2002; Kopper & Lindroth, 2003). ...
Threat to future global food security from climate change and ozone air pollution
1
2014
... (4)关注O3对作物品质及食品安全的危害: 世界上主要粮食作物要么对O3敏感(小麦、大豆等), 要么对O3适度敏感(玉米、水稻和马铃薯等).到2050年, 世界人口预计增加到90亿, 粮食供应需增加至少50%才能满足人口增长的需求, 如何保证粮食供应安全将成为21世纪最重要的一个挑战(Tai et al., 2014).目前为止, 除作物产量之外, 有关地表O3浓度升高对作物品质的影响研究多集中在蔬菜作物或粮食作物可食部位糖分、蛋白质和淀粉含量等食用口感方面(Booker et al., 2009), 很少涉及其背后对人体健康的直接影响.随着生活水平的提高, 人们更加关注食品的营养与安全(如微量元素含量或健康风险等), 而长期高浓度O3暴露可改变粮食作物籽粒微量元素及营养物质含量(Booker et al., 2009; 冯兆忠等, 2018).此外, 也有研究显示O3暴露导致马铃薯的糖苷生物碱含量增加, 糖苷生物碱不仅导致味苦而且浓度较高时具有毒性(Pell & Pearson, 1984).因此, 在全球环境变化大背景下, 长期食用暴露在O3污染下的粮食作物是否影响人体营养平衡或危及健康值得关注.未来的研究应关注高浓度O3对农作物外观品质(如籽粒形态、发芽率和成活率等)、加工特性、口感(如黏弹性、柔软性等)、烹饪特性和营养特性等外在和内在价值的综合影响. ...
A system for free-air ozone concentration elevation with rice and wheat: control performance and ozone exposure regime
1
2011
... 在此期间, 尽管社会经济不断发展、实验设备和研究方法不断更新, 但地表O3污染对植物及生态系统影响的研究主要还是以模拟O3浓度升高的控制实验为主.纵观发展历程, 试验设备与手段的改进和更新大致可分为三个阶段(王春乙, 1995): (1) 1970年之前, 研究主要利用封闭式静态或动态气室或室内生长箱进行模拟实验(Heagle et al., 1973). (2) 1973年之后, 美国学者Heagle等(1979)和Mandle (1973)研制开顶式气室(OTC)模拟高浓度O3对植物的损伤和影响机制, 并最终在世界范围内推广, 成为研究地表O3浓度升高对植物及生态系统影响的主流手段.(3)由于OTCs气室内外较大的环境差异, OTCs模拟实验下的熏蒸结果是否适用于大田研究一直备受争议.为解决OTCs内微气候环境带来的影响, 1986年Hendrey和Kimball (1994)提出建立自由空气中气体浓度增加(FACE)系统的想法, 并逐步用于大田实验.由于与外界大气几乎一致的气体流速、温度、光照和风速等条件(Matyssek et al., 2010), FACE系统被普遍认为是当前研究地表O3浓度升高对植物及生态系统影响最为理想的实验平台, 但由于造价昂贵, 技术要求高, 只有少数国家和实验室开展相关研究(有关上述研究方法的优缺点见表2).O3熏蒸的FACE平台仅有10个左右且多数已经停止运行(Paoletti et al., 2017), 主要包括美国威斯康星州Aspen FACE (停止运行; Karnosky et al., 2003), 美国伊利诺伊州Soybean FACE (停止运行; Morgan et al., 2004), 瑞典苏尔草地FACE (停止运行; Volk et al., 2003), 芬兰库奥皮奥FACE (停止运行; Oksanen, 2003), 德国克兰兹堡森林FACE (停止运行; Matyssek et al., 2010), 中国江都水稻(Oryza sativa)/小麦(Triticum aestivum)轮作FACE (停止运行; Tang et al., 2011), 日本札幌森林FACE (仍在运行; Watanabe et al., 2013), 意大利佛罗伦萨3D FACE (仍在运行; Paoletti et al., 2017), 以及刚刚建成的中国延庆唐家堡杨树人工林FACE.由于实验条件和FACE平台规模的限制, 有关地表O3浓度升高对生态系统水平的研究成果主要来自美国Aspen FACE和德国克兰兹堡森林FACE (Grulke & Heath, 2020). ...
Defense and avoidance of ozone under global change
1
2007
... 气孔防御机制失效后, 进入细胞的O3将诱导叶片内部抗氧化系统启动解毒和修复的非气孔防御响应(冯兆忠等, 2018).非气孔防御响应主要是指“质外体-共质体系统”内一些抗氧化组分对O3解毒和修复的防御过程.具体来说, O3进入植物叶片的细胞间隙后会溶解在细胞壁的结合水里, 经过一系列的化学反应形成活性氧分子(ROS), ROS能够破坏细胞壁并与细胞膜上的不饱和脂肪酸发生反应, 从而对叶片造成损伤(Tausz et al., 2007; Pellegrini et al., 2018).植物质外体的抗氧化物质如还原性抗坏血酸(ASC)、抗氧化酶(如超氧化物歧化酶)可与ROS反应进行初级解毒, 成为抵御O3的第二道防线(Kangasjärvi et al., 2005; Dai et al., 2019), 而剩余的ROS会经过质膜到达共质体, 此时共质体内抗氧化成分如过氧化物酶(POD)、过氧化氢酶(CAT)、抗坏血酸过氧化物酶(APX)及抗氧化剂(如酚类化合物-谷胱甘肽(GSH)、脯氨酸和多胺等)等抗氧化酶类物质被诱导作为次级解毒响应机制参与诸如“抗坏血酸-谷胱甘肽(ASA-GSH)循环”等氧化还原过程, 进而对ROS进行进一步解毒与消除(Luwe, 1996).气孔闭合除了会减少O3的吸收, 也会抑制CO2吸收和水汽交换, 而非气孔防御响应中的抗氧化系统解毒过程也会消耗大量的能量并对光合反应系统造成不同程度的损伤, 降低羧化速率和电子传递速率, 进而抑制光合和蒸腾作用, 损害植物生长(Gao et al., 2016).然而, 最新研究表明持续整个生长季的长期O3暴露会影响植物叶片的气孔密度、开度和大小等表观属性(李品等, 2018), 而O3从气孔进入到细胞间隙后, 也可以通过影响植物的叶肉导度降低植物的光合能力(Xu et al., 2019).此外, 有关29种落叶和常绿树种应对O3胁迫的研究还发现不同木本植物对O3响应的差异主要来自植物功能叶片比叶质量的大小(即叶片比叶质量越大的树种对O3越不敏感), 与气孔导度无明显相关性(Li et al., 2016). ...
Impact of elevated ozone on growth, yield and nutritional quality of two wheat species in Northern India
2
2015
... 相对于农作物产量, O3对粮食品质影响的研究集中在几种主要农作物(如小麦、水稻和大豆)(冯兆忠等, 2018).基于北美、欧洲和亚洲已有的O3对小麦影响的meta分析发现, O3显著降低小麦淀粉浓度, 但显著增加蛋白质和一些营养元素浓度(如钾、镁、钙、磷、锌、锰和铜)(Broberg et al., 2015).印度北部热带地区有关小麦的研究则显示, O3显著降低小麦籽粒糖含量, 但没有改变籽粒蛋白质含量(Tomer et al., 2015).有关水稻的研究还发现, O3增加粳稻直链淀粉含量, 但对籼稻直链淀粉含量无显著影响(Sawada et al., 2016).国内研究则表明, 尽管高浓度O3显著降低冬小麦面粉中支链淀粉和淀粉含量, 但是增加直链淀粉含量(Zhang et al., 2013).O3显著降低水稻稻米直链淀粉浓度、分解率, 增加黏度、起糊温度、蛋白质和微量矿物质浓度, 但蛋白质总量没有改变(Wang et al., 2012).此外, 研究还发现O3熏蒸显著降低了小麦籽粒出粉率和硬度, 但增加冬小麦面粉峰值黏度、低谷黏度、最终黏度和反弹值(贾一磊, 2016). ...
... (1)开展O3-FACE研究: 现有研究多为OTC系统下的实验, OTC气室内“微气候” (如温度、水分、光照和风力等生态因素)与外界环境状态存在明显差异, 这些生态因子的变化可能影响植物应对O3的响应, 缓解或加重O3污染对农作物产量或森林植被生产力的损伤(Long et al., 2005; Feng et al., 2018a).全球模型评估结果显示, OTC实验结果低估了O3对作物产量损失的影响(van Dingenen et al., 2009).最近, Feng等(2018b)也发现, 同样浓度的O3对OTC实验下小麦和水稻产量的影响要低于FACE实验; 而对大豆产量的影响则相反.因此, OTC实验结果并不能完全应用于实际O3区域风险的评估.O3-FACE平台是未来研究实际大田环境下O3浓度升高对农林生态系统影响的主要依托平台.为减少区域O3风险评估的不确定性, 未来应加大O3-FACE平台的实验研究.考虑到目前已开展的FACE实验多坐落在温带地区, 主要探究温带植被或作物对高浓度O3的响应, 较少涉及热带和寒带植物或其他区域典型生态系统(Ainsworth et al., 2020).最理想的情况是, 不同区域、不同生态系统类型共同开展O3-FACE实验研究(Ainsworth et al., 2012). ...
The global impact of ozone on agricultural crop yields under current and future air quality legislation
2009
Design and performance of a free-air exposure system to study long-term effects of ozone on grasslands
1
2003
... 在此期间, 尽管社会经济不断发展、实验设备和研究方法不断更新, 但地表O3污染对植物及生态系统影响的研究主要还是以模拟O3浓度升高的控制实验为主.纵观发展历程, 试验设备与手段的改进和更新大致可分为三个阶段(王春乙, 1995): (1) 1970年之前, 研究主要利用封闭式静态或动态气室或室内生长箱进行模拟实验(Heagle et al., 1973). (2) 1973年之后, 美国学者Heagle等(1979)和Mandle (1973)研制开顶式气室(OTC)模拟高浓度O3对植物的损伤和影响机制, 并最终在世界范围内推广, 成为研究地表O3浓度升高对植物及生态系统影响的主流手段.(3)由于OTCs气室内外较大的环境差异, OTCs模拟实验下的熏蒸结果是否适用于大田研究一直备受争议.为解决OTCs内微气候环境带来的影响, 1986年Hendrey和Kimball (1994)提出建立自由空气中气体浓度增加(FACE)系统的想法, 并逐步用于大田实验.由于与外界大气几乎一致的气体流速、温度、光照和风速等条件(Matyssek et al., 2010), FACE系统被普遍认为是当前研究地表O3浓度升高对植物及生态系统影响最为理想的实验平台, 但由于造价昂贵, 技术要求高, 只有少数国家和实验室开展相关研究(有关上述研究方法的优缺点见表2).O3熏蒸的FACE平台仅有10个左右且多数已经停止运行(Paoletti et al., 2017), 主要包括美国威斯康星州Aspen FACE (停止运行; Karnosky et al., 2003), 美国伊利诺伊州Soybean FACE (停止运行; Morgan et al., 2004), 瑞典苏尔草地FACE (停止运行; Volk et al., 2003), 芬兰库奥皮奥FACE (停止运行; Oksanen, 2003), 德国克兰兹堡森林FACE (停止运行; Matyssek et al., 2010), 中国江都水稻(Oryza sativa)/小麦(Triticum aestivum)轮作FACE (停止运行; Tang et al., 2011), 日本札幌森林FACE (仍在运行; Watanabe et al., 2013), 意大利佛罗伦萨3D FACE (仍在运行; Paoletti et al., 2017), 以及刚刚建成的中国延庆唐家堡杨树人工林FACE.由于实验条件和FACE平台规模的限制, 有关地表O3浓度升高对生态系统水平的研究成果主要来自美国Aspen FACE和德国克兰兹堡森林FACE (Grulke & Heath, 2020). ...
臭氧对农作物的影响研究
1
1995
... 在此期间, 尽管社会经济不断发展、实验设备和研究方法不断更新, 但地表O3污染对植物及生态系统影响的研究主要还是以模拟O3浓度升高的控制实验为主.纵观发展历程, 试验设备与手段的改进和更新大致可分为三个阶段(王春乙, 1995): (1) 1970年之前, 研究主要利用封闭式静态或动态气室或室内生长箱进行模拟实验(Heagle et al., 1973). (2) 1973年之后, 美国学者Heagle等(1979)和Mandle (1973)研制开顶式气室(OTC)模拟高浓度O3对植物的损伤和影响机制, 并最终在世界范围内推广, 成为研究地表O3浓度升高对植物及生态系统影响的主流手段.(3)由于OTCs气室内外较大的环境差异, OTCs模拟实验下的熏蒸结果是否适用于大田研究一直备受争议.为解决OTCs内微气候环境带来的影响, 1986年Hendrey和Kimball (1994)提出建立自由空气中气体浓度增加(FACE)系统的想法, 并逐步用于大田实验.由于与外界大气几乎一致的气体流速、温度、光照和风速等条件(Matyssek et al., 2010), FACE系统被普遍认为是当前研究地表O3浓度升高对植物及生态系统影响最为理想的实验平台, 但由于造价昂贵, 技术要求高, 只有少数国家和实验室开展相关研究(有关上述研究方法的优缺点见表2).O3熏蒸的FACE平台仅有10个左右且多数已经停止运行(Paoletti et al., 2017), 主要包括美国威斯康星州Aspen FACE (停止运行; Karnosky et al., 2003), 美国伊利诺伊州Soybean FACE (停止运行; Morgan et al., 2004), 瑞典苏尔草地FACE (停止运行; Volk et al., 2003), 芬兰库奥皮奥FACE (停止运行; Oksanen, 2003), 德国克兰兹堡森林FACE (停止运行; Matyssek et al., 2010), 中国江都水稻(Oryza sativa)/小麦(Triticum aestivum)轮作FACE (停止运行; Tang et al., 2011), 日本札幌森林FACE (仍在运行; Watanabe et al., 2013), 意大利佛罗伦萨3D FACE (仍在运行; Paoletti et al., 2017), 以及刚刚建成的中国延庆唐家堡杨树人工林FACE.由于实验条件和FACE平台规模的限制, 有关地表O3浓度升高对生态系统水平的研究成果主要来自美国Aspen FACE和德国克兰兹堡森林FACE (Grulke & Heath, 2020). ...
臭氧对农作物的影响研究
1
1995
... 在此期间, 尽管社会经济不断发展、实验设备和研究方法不断更新, 但地表O3污染对植物及生态系统影响的研究主要还是以模拟O3浓度升高的控制实验为主.纵观发展历程, 试验设备与手段的改进和更新大致可分为三个阶段(王春乙, 1995): (1) 1970年之前, 研究主要利用封闭式静态或动态气室或室内生长箱进行模拟实验(Heagle et al., 1973). (2) 1973年之后, 美国学者Heagle等(1979)和Mandle (1973)研制开顶式气室(OTC)模拟高浓度O3对植物的损伤和影响机制, 并最终在世界范围内推广, 成为研究地表O3浓度升高对植物及生态系统影响的主流手段.(3)由于OTCs气室内外较大的环境差异, OTCs模拟实验下的熏蒸结果是否适用于大田研究一直备受争议.为解决OTCs内微气候环境带来的影响, 1986年Hendrey和Kimball (1994)提出建立自由空气中气体浓度增加(FACE)系统的想法, 并逐步用于大田实验.由于与外界大气几乎一致的气体流速、温度、光照和风速等条件(Matyssek et al., 2010), FACE系统被普遍认为是当前研究地表O3浓度升高对植物及生态系统影响最为理想的实验平台, 但由于造价昂贵, 技术要求高, 只有少数国家和实验室开展相关研究(有关上述研究方法的优缺点见表2).O3熏蒸的FACE平台仅有10个左右且多数已经停止运行(Paoletti et al., 2017), 主要包括美国威斯康星州Aspen FACE (停止运行; Karnosky et al., 2003), 美国伊利诺伊州Soybean FACE (停止运行; Morgan et al., 2004), 瑞典苏尔草地FACE (停止运行; Volk et al., 2003), 芬兰库奥皮奥FACE (停止运行; Oksanen, 2003), 德国克兰兹堡森林FACE (停止运行; Matyssek et al., 2010), 中国江都水稻(Oryza sativa)/小麦(Triticum aestivum)轮作FACE (停止运行; Tang et al., 2011), 日本札幌森林FACE (仍在运行; Watanabe et al., 2013), 意大利佛罗伦萨3D FACE (仍在运行; Paoletti et al., 2017), 以及刚刚建成的中国延庆唐家堡杨树人工林FACE.由于实验条件和FACE平台规模的限制, 有关地表O3浓度升高对生态系统水平的研究成果主要来自美国Aspen FACE和德国克兰兹堡森林FACE (Grulke & Heath, 2020). ...
Aggravating O3 pollution due to NOx emission control in eastern China
1
2019
... 地表臭氧(O3)特指距离地球表面15 km范围内的近地层O3, 除少量来自平流层大气传输外, 其余大部分是氮氧化物(NOx)、非甲烷类挥发性有机化合物(NMVOCs)、一氧化碳(CO)和甲烷(CH4)等前体物在强烈光照下发生光化学反应而产生(冯兆忠等, 2018).自工业革命以来, 伴随城市化进程加快和化石燃料的过度燃烧, 地表O3浓度在世界范围内普遍升高.近年来, 模型和观测数据都表明地表O3浓度小时峰值在欧洲和北美等地已有所下降, 但亚洲地区仍呈不断上升的趋势(Cooper et al., 2014).中国国家环境监测网络2013-2017年的数据表明, 过去5年中国地表O3浓度以每年3 μg·kg-1的速率攀升(Wang et al., 2019), 目前全国地表O3浓度每日8 h滑动平均最大值(MDA8)的年平均值已达(41.2 ± 6.3) μg·kg-1, 尤其华北平原、长江三角洲和珠江三角洲等地(MDA8峰值年平均值已处于60-70 μg·kg-1), 地表O3污染形势十分严峻(Lu et al., 2018).地表O3是具有植物毒性的气体污染物, 具有强氧化性.高浓度地表O3或长期低浓度O3能诱导植物叶片出现可见损伤, 加速植物衰老, 降低光合作用, 改变碳分配, 抑制植物生长和生物量积累, 影响作物品质和产量, 最终降低生态系统碳汇能力, 改变生态系统结构和功能(Ashmore, 2005; Ainsworth, 2017; 高峰, 2018). ...
Characterizing distributions of surface ozone and its impact on grain production in China, Japan and South Korea: 1990 and 2020
1
2004
... 基于OTCs和O3-FACE平台下单个站点的控制实验可探究O3对植物个体、群落或生态系统的影响, 但将站点或样地尺度的研究外推到区域或全球, 评估O3污染区域风险的研究主要通过响应关系和模型.目前, 其研究领域主要集中在地表O3污染对农作物产量和森林生产力等方面.早在1994年, Chameides等(1994)就利用经济模型初步预测O3浓度升高可能导致2025年全球30%-75%的作物面临产量降低的风险.基于O3前体物排放、化学反应传输模型和M12、AOT40指标的评估显示, 2000年全球O3污染引起全球小麦、玉米和大豆产量平均降低2.2%-15%, 每年因上述作物产量降低所导致的经济损失达110-180亿美元(Avnery et al., 2011).最近, 基于PODY评估指标的研究则表明, O3对小麦造成的产量损失北半球(9.9%)明显高于南半球(6.2%), 平均产量损失最高的地区主要位于北半球暖温带湿润地区(12.5%)、热带湿热(12.5%)和热带潮湿气候区(17.2%)(Mills et al., 2018).在区域尺度上, 基于1990年O3浓度数据, Wang和Mauzerall (2004)利用M7和M12指标评估了O3对中国、日本和韩国主要农作物产量的影响, 结果显示O3污染造成小麦、玉米、棉花减产1%-9%, 大豆减产23%-27%, 造成2.4-35亿美元的经济损失, 在维持现有的污染物排放、管理措施和农业生产技术不变的情况下, 预计到2020年小麦、玉米、棉花减产幅度为2%-15%, 大豆为28%-35%.最近, 利用AOT40指标的研究显示, 2014年当前O3浓度导致中国农作物产量损失7 840万t, 小麦、水稻和玉米相对减产量分别为8.5%-14%、3.9%-15%和2.2%-5.5% (Lin et al., 2018).而只聚焦于长江三角洲地区的研究则发现, 2015年区域O3污染分别造成小麦和水稻减产9.9%-36.1%和7.3%- 23.9% (Zhao et al., 2018).虽然国内也有少量研究建立了PODY与个别树种或作物相对生物量或相对产量之间的通量响应关系(Feng et al., 2012; Hu et al., 2015; Gao et al., 2017), 但并没有研究利用PODY指标评估区域O3污染对农作物产量损失的影响. ...
The impact of elevated tropospheric ozone on grain quality of hybrid rice: a free-air gas concentration enrichment (FACE) experiment
1
2012
... 相对于农作物产量, O3对粮食品质影响的研究集中在几种主要农作物(如小麦、水稻和大豆)(冯兆忠等, 2018).基于北美、欧洲和亚洲已有的O3对小麦影响的meta分析发现, O3显著降低小麦淀粉浓度, 但显著增加蛋白质和一些营养元素浓度(如钾、镁、钙、磷、锌、锰和铜)(Broberg et al., 2015).印度北部热带地区有关小麦的研究则显示, O3显著降低小麦籽粒糖含量, 但没有改变籽粒蛋白质含量(Tomer et al., 2015).有关水稻的研究还发现, O3增加粳稻直链淀粉含量, 但对籼稻直链淀粉含量无显著影响(Sawada et al., 2016).国内研究则表明, 尽管高浓度O3显著降低冬小麦面粉中支链淀粉和淀粉含量, 但是增加直链淀粉含量(Zhang et al., 2013).O3显著降低水稻稻米直链淀粉浓度、分解率, 增加黏度、起糊温度、蛋白质和微量矿物质浓度, 但蛋白质总量没有改变(Wang et al., 2012).此外, 研究还发现O3熏蒸显著降低了小麦籽粒出粉率和硬度, 但增加冬小麦面粉峰值黏度、低谷黏度、最终黏度和反弹值(贾一磊, 2016). ...
Photosynthetic traits of Siebold’s beech and oak saplings grown under free air ozone exposure
1
2013
... 在此期间, 尽管社会经济不断发展、实验设备和研究方法不断更新, 但地表O3污染对植物及生态系统影响的研究主要还是以模拟O3浓度升高的控制实验为主.纵观发展历程, 试验设备与手段的改进和更新大致可分为三个阶段(王春乙, 1995): (1) 1970年之前, 研究主要利用封闭式静态或动态气室或室内生长箱进行模拟实验(Heagle et al., 1973). (2) 1973年之后, 美国学者Heagle等(1979)和Mandle (1973)研制开顶式气室(OTC)模拟高浓度O3对植物的损伤和影响机制, 并最终在世界范围内推广, 成为研究地表O3浓度升高对植物及生态系统影响的主流手段.(3)由于OTCs气室内外较大的环境差异, OTCs模拟实验下的熏蒸结果是否适用于大田研究一直备受争议.为解决OTCs内微气候环境带来的影响, 1986年Hendrey和Kimball (1994)提出建立自由空气中气体浓度增加(FACE)系统的想法, 并逐步用于大田实验.由于与外界大气几乎一致的气体流速、温度、光照和风速等条件(Matyssek et al., 2010), FACE系统被普遍认为是当前研究地表O3浓度升高对植物及生态系统影响最为理想的实验平台, 但由于造价昂贵, 技术要求高, 只有少数国家和实验室开展相关研究(有关上述研究方法的优缺点见表2).O3熏蒸的FACE平台仅有10个左右且多数已经停止运行(Paoletti et al., 2017), 主要包括美国威斯康星州Aspen FACE (停止运行; Karnosky et al., 2003), 美国伊利诺伊州Soybean FACE (停止运行; Morgan et al., 2004), 瑞典苏尔草地FACE (停止运行; Volk et al., 2003), 芬兰库奥皮奥FACE (停止运行; Oksanen, 2003), 德国克兰兹堡森林FACE (停止运行; Matyssek et al., 2010), 中国江都水稻(Oryza sativa)/小麦(Triticum aestivum)轮作FACE (停止运行; Tang et al., 2011), 日本札幌森林FACE (仍在运行; Watanabe et al., 2013), 意大利佛罗伦萨3D FACE (仍在运行; Paoletti et al., 2017), 以及刚刚建成的中国延庆唐家堡杨树人工林FACE.由于实验条件和FACE平台规模的限制, 有关地表O3浓度升高对生态系统水平的研究成果主要来自美国Aspen FACE和德国克兰兹堡森林FACE (Grulke & Heath, 2020). ...
Age effects on Norway spruce (Picea abies) susceptibility to ozone uptake: a novel approach relating stress avoidance to defense
1
2002
... 通常认为, 植物通过气孔防御和非气孔防御对O3胁迫做出响应.气孔作为叶片与外界气体交换的“门户”, 是O3进入植物体内的主要通道, 控制着植物蒸腾、呼吸和光合等重要生理过程.正常光热条件下, 气孔处于打开状态, 但高浓度O3可以直接影响调节气孔开张的保卫细胞上钾离子浓度, 改变细胞膨压, 诱导气孔关闭, 引发避逆性的防御响应(Wieser et al., 2002).因此, 气孔闭合被认为是植物抵御O3胁迫的第一道防线(Ainsworth, 2017).然而, 气孔对O3的响应(关闭)也并不是始终如一的.研究发现随着O3暴露时间的延长, O3诱导的气孔关闭会出现滞后甚至失灵的现象(Hoshika et al., 2012), 从而导致大量O3进入细胞间隙, 气孔防御机制 失效. ...
Quantifying the impact of current and future tropospheric ozone on tree biomass, growth, physiology and biochemistry: a quantitative meta-analysis
2
2009
... 地表O3浓度升高对植物光合作用、生长和生物量积累、物质分配等过程的连锁负效应导致陆地生态系统生产力和固碳能力降低(Reich, 1987; Wittig et al., 2009; Ainsworth et al., 2012), 削弱了自然生态系统的碳汇能力(Felzer et al., 2005; Sitch et al., 2007; Collins et al., 2010).基于大量OTCs控制实验下北半球中纬度北方温带树种的meta分析发现, O3浓度升高导致树木的净光合作用和总生物量分别降低11%和7% (Wittig et al., 2009).最近, 有关中国热带、亚热带和温带树种的meta分析则发现, O3浓度升高导致中国木本植物的净光合作用和总生物量分别降低28%和14% (Li et al., 2017). ...
... 浓度升高导致树木的净光合作用和总生物量分别降低11%和7% (Wittig et al., 2009).最近, 有关中国热带、亚热带和温带树种的meta分析则发现, O3浓度升高导致中国木本植物的净光合作用和总生物量分别降低28%和14% (Li et al., 2017). ...
Mesophyll conductance limitation of photosynthesis in poplar under elevated ozone
1
2019
... 气孔防御机制失效后, 进入细胞的O3将诱导叶片内部抗氧化系统启动解毒和修复的非气孔防御响应(冯兆忠等, 2018).非气孔防御响应主要是指“质外体-共质体系统”内一些抗氧化组分对O3解毒和修复的防御过程.具体来说, O3进入植物叶片的细胞间隙后会溶解在细胞壁的结合水里, 经过一系列的化学反应形成活性氧分子(ROS), ROS能够破坏细胞壁并与细胞膜上的不饱和脂肪酸发生反应, 从而对叶片造成损伤(Tausz et al., 2007; Pellegrini et al., 2018).植物质外体的抗氧化物质如还原性抗坏血酸(ASC)、抗氧化酶(如超氧化物歧化酶)可与ROS反应进行初级解毒, 成为抵御O3的第二道防线(Kangasjärvi et al., 2005; Dai et al., 2019), 而剩余的ROS会经过质膜到达共质体, 此时共质体内抗氧化成分如过氧化物酶(POD)、过氧化氢酶(CAT)、抗坏血酸过氧化物酶(APX)及抗氧化剂(如酚类化合物-谷胱甘肽(GSH)、脯氨酸和多胺等)等抗氧化酶类物质被诱导作为次级解毒响应机制参与诸如“抗坏血酸-谷胱甘肽(ASA-GSH)循环”等氧化还原过程, 进而对ROS进行进一步解毒与消除(Luwe, 1996).气孔闭合除了会减少O3的吸收, 也会抑制CO2吸收和水汽交换, 而非气孔防御响应中的抗氧化系统解毒过程也会消耗大量的能量并对光合反应系统造成不同程度的损伤, 降低羧化速率和电子传递速率, 进而抑制光合和蒸腾作用, 损害植物生长(Gao et al., 2016).然而, 最新研究表明持续整个生长季的长期O3暴露会影响植物叶片的气孔密度、开度和大小等表观属性(李品等, 2018), 而O3从气孔进入到细胞间隙后, 也可以通过影响植物的叶肉导度降低植物的光合能力(Xu et al., 2019).此外, 有关29种落叶和常绿树种应对O3胁迫的研究还发现不同木本植物对O3响应的差异主要来自植物功能叶片比叶质量的大小(即叶片比叶质量越大的树种对O3越不敏感), 与气孔导度无明显相关性(Li et al., 2016). ...
Effects of ozone on nitrogen metabolism in the leaves of Fagus crenata seedlings under different soil nitrogen loads
1
2010
... 除单因子O3胁迫外, 由于全球气候变化和极端气候常态化, 植物经常受到O3和其他环境因子交互作用的影响(如O3与CO2、升温、干旱或氮沉降等)(Schmadel-Hagebölling et al., 1998; Ollinger et al., 2002; Eltayeb et al., 2006; Ghimire et al., 2018).然而, 植物应对O3与其他因子交互作用的响应并不是两者或单独作用的简单叠加, 往往取决于植物种类、生长时期、对特定环境因子的敏感性、胁迫浓度及时间, 以及具体的微气候条件(列淦文和薛立, 2014).一方面, 通过诱导气孔关闭、限制叶片O3吸收(CO2浓度升高/干旱胁迫), 或提高碳同化速率、促进植物光合作用(CO2浓度升高/氮增加), CO2浓度升高、干旱胁迫和氮沉降经常被报道可增强植物抵御O3的能力, 减缓O3伤害(白月明等, 2005; Handley & Grulke, 2008; Gao et al., 2017).另一方面, 与之相反, 也有研究发现长期高浓度CO2或干旱降低气孔导度的同时, 会引起植物蒸腾速率、光合产物合成和积累的降低(McDonald et al., 2002; Pollastrini et al., 2010); 而超过植物承载阈值的高氮负荷也会对植物的光合生理活性产生负效应(Yamaguchi et al., 2010), 进而加重植物在复合环境下的碳损失、水分蒸散损失和自由基积累, 对植物产生协同伤害(Karnosky et al., 2005; 列淦文和薛立, 2014).高峰(2018)认为O3与其他环境胁迫因子出现的先后次序也会影响植物对复合作用的响应. ...
Ozone and haze pollution weakens net primary productivity in China
1
2017
... 区域尺度上, 地表O3污染对陆地生态系统固碳能力的影响主要依靠模式算法的估算.早在20世纪90年代, 基于AOT40指标的研究就发现高浓度O3降低美国东北部阔叶林净初级生产力(NPP) 3%-16%, 对地上生物量的降低每年为0-10% (Chappelka & Samuelson, 1998).另有研究表明O3污染造成美国植被碳固定降低18-38 Tg·a-1, NPP降低至少9 g·m-2·a-1, 且高浓度O3导致植物-土壤系统碳积累和固定能力下降(Felzer et al., 2004).此外, 运用自主开发的基于过程的陆地生态系统动态模型(DLEM), 国内学者发现近40年来O3浓度升高使中国森林净固碳量减少7.7%, 且不同的森林类型降低幅度不同: 如北方落叶阔叶林降低21.8%, 北方落叶针叶林降低5.2%, 温带落叶阔叶林降低21.1%, 温带常绿阔叶林降低7.6%, 温带常绿针叶林降低0.4%, 温带落叶针叶林降低6.2%, 热带落叶阔叶林降低43.1%, 热带常绿阔叶林降低9.6% (Ren et al., 2011).最近, Yue等(2017)用地球系统模式(NASA ModelE2-YIBs)得出O3污染造成中国陆地植被NPP降低600 Tg·a-1, 且对于不同功能类型的植物, O3对总初级生产力(GPP)的降低都随着O3浓度的升高而加大.总体来看, 模型驱动的结果与大量控制实验的发现相吻合, 均表明O3污染在不同尺度都对自然生态系统碳固定有显著的负效应, 未来O3浓度升高有可能抵消因大气CO2浓度升高所增加的GPP (18%-34%)(Sitch et al., 2007). ...
Effects of elevated ozone concentration on starch and starch synthesis enzymes of yangmai 16 under fully open-air field conditions
1
2013
... 相对于农作物产量, O3对粮食品质影响的研究集中在几种主要农作物(如小麦、水稻和大豆)(冯兆忠等, 2018).基于北美、欧洲和亚洲已有的O3对小麦影响的meta分析发现, O3显著降低小麦淀粉浓度, 但显著增加蛋白质和一些营养元素浓度(如钾、镁、钙、磷、锌、锰和铜)(Broberg et al., 2015).印度北部热带地区有关小麦的研究则显示, O3显著降低小麦籽粒糖含量, 但没有改变籽粒蛋白质含量(Tomer et al., 2015).有关水稻的研究还发现, O3增加粳稻直链淀粉含量, 但对籼稻直链淀粉含量无显著影响(Sawada et al., 2016).国内研究则表明, 尽管高浓度O3显著降低冬小麦面粉中支链淀粉和淀粉含量, 但是增加直链淀粉含量(Zhang et al., 2013).O3显著降低水稻稻米直链淀粉浓度、分解率, 增加黏度、起糊温度、蛋白质和微量矿物质浓度, 但蛋白质总量没有改变(Wang et al., 2012).此外, 研究还发现O3熏蒸显著降低了小麦籽粒出粉率和硬度, 但增加冬小麦面粉峰值黏度、低谷黏度、最终黏度和反弹值(贾一磊, 2016). ...
Assessment of yield and economic losses for wheat and rice due to ground level O3 exposure in the Yangtze River Delta, China
1
2018
... 基于OTCs和O3-FACE平台下单个站点的控制实验可探究O3对植物个体、群落或生态系统的影响, 但将站点或样地尺度的研究外推到区域或全球, 评估O3污染区域风险的研究主要通过响应关系和模型.目前, 其研究领域主要集中在地表O3污染对农作物产量和森林生产力等方面.早在1994年, Chameides等(1994)就利用经济模型初步预测O3浓度升高可能导致2025年全球30%-75%的作物面临产量降低的风险.基于O3前体物排放、化学反应传输模型和M12、AOT40指标的评估显示, 2000年全球O3污染引起全球小麦、玉米和大豆产量平均降低2.2%-15%, 每年因上述作物产量降低所导致的经济损失达110-180亿美元(Avnery et al., 2011).最近, 基于PODY评估指标的研究则表明, O3对小麦造成的产量损失北半球(9.9%)明显高于南半球(6.2%), 平均产量损失最高的地区主要位于北半球暖温带湿润地区(12.5%)、热带湿热(12.5%)和热带潮湿气候区(17.2%)(Mills et al., 2018).在区域尺度上, 基于1990年O3浓度数据, Wang和Mauzerall (2004)利用M7和M12指标评估了O3对中国、日本和韩国主要农作物产量的影响, 结果显示O3污染造成小麦、玉米、棉花减产1%-9%, 大豆减产23%-27%, 造成2.4-35亿美元的经济损失, 在维持现有的污染物排放、管理措施和农业生产技术不变的情况下, 预计到2020年小麦、玉米、棉花减产幅度为2%-15%, 大豆为28%-35%.最近, 利用AOT40指标的研究显示, 2014年当前O3浓度导致中国农作物产量损失7 840万t, 小麦、水稻和玉米相对减产量分别为8.5%-14%、3.9%-15%和2.2%-5.5% (Lin et al., 2018).而只聚焦于长江三角洲地区的研究则发现, 2015年区域O3污染分别造成小麦和水稻减产9.9%-36.1%和7.3%- 23.9% (Zhao et al., 2018).虽然国内也有少量研究建立了PODY与个别树种或作物相对生物量或相对产量之间的通量响应关系(Feng et al., 2012; Hu et al., 2015; Gao et al., 2017), 但并没有研究利用PODY指标评估区域O3污染对农作物产量损失的影响. ...




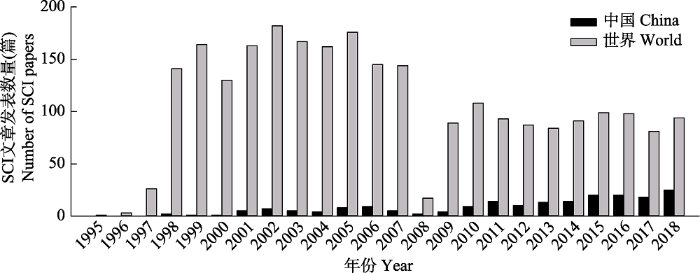
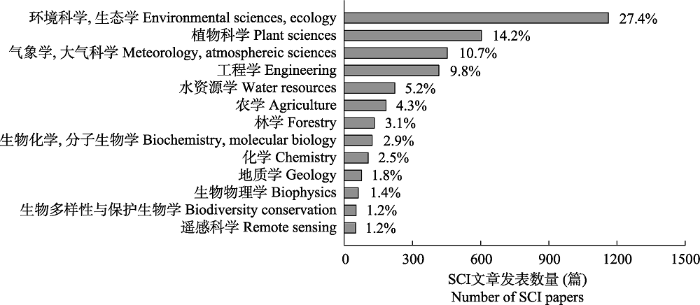
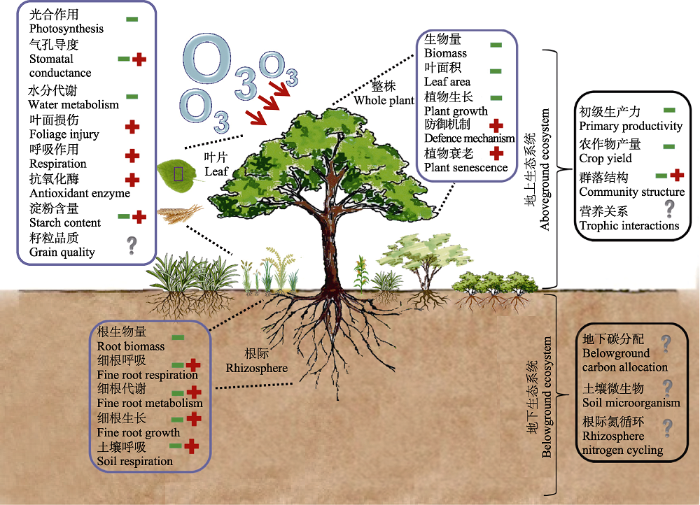



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}