


由于环境因素的限制, 不同物种只能在一定范围内栖息生存(Daniel et al., 2019)。这些环境因素通常包括温度、降水、地形、土壤理化性质和人为干扰因子(Roberts & Wuest, 1999; Chi et al., 2014; Feng et al., 2014)。温度和降水通过影响物种生态幅, 决定植物物种的空间分布(Chuine, 2010; Bykova & Sage, 2012; Porfirio et al., 2014)。海拔直接反映地表起伏、地形等因素, 通过影响太阳辐射及降水在空间上的再分配, 影响物种的空间更替(沈泽昊, 2002)。土壤空间异质性与植物水分、养分获取直接相关, 是驱动物种周转的重要因素(Paoli et al., 2006; Ulrich et al., 2014)。人类活动造成的土地利用变化, 改变了物种原生生境, 抑制物种空间更替, 导致区域内生物同质化(Maestre & Cortina, 2004; 陈圣宾等, 2010)。
我国中亚热带北部, 地处亚热带湿润气候区, 长期受人类活动干扰, 地带性常绿阔叶林大面积退化丧失, 形成了大面积的次生灌丛(杨一, 2015)。次生灌丛已成为我国中亚热带北部地区主要的植被类型(Su et al., 2020)。目前, 对亚热带灌丛的研究主要集中在群落特征和物种多样性特征(张亚茹等, 2013; 刘梦等, 2017)、物种多样性影响因素(张慧等, 2017;王飞等, 2018)等方面, 掌握了亚热带灌丛群落物种基本组成及其空间分布。但是, 灌丛物种组成在空间上的周转格局如何? 是什么因素驱动了物种周转? 到目前为止, 该方面的研究鲜有报道。本研究拟探究环境因子驱动下中亚热带北部灌丛群落物种空间周转格局, 以期为区域生物多样性保护和生态系统管理提供科学依据。
1 材料和方法
1.1 研究区概况
中亚热带北部位于25.96°-31.46° N, 106.14°- 121.19° E, 包括安徽、浙江、江西、湖南、湖北、重庆等省市(图1)。属亚热带季风气候, 年平均气温16-21 ℃, 最冷月平均气温5-12 ℃, 最热月平均气温25-30 ℃, 全年无霜期为270-300天, ≥10 ℃的年积温4 000-6 500 ℃, 年降水量为1 000-2 000 mm。土壤类型以红壤、黄壤为主(中国植被编辑委员会, 1980), 地带性植被为常绿阔叶林。主要灌丛类型有枹栎(Quercus serrata)灌丛、茅栗(Castanea seguinii)灌丛、杜鹃(Rhododendron simsii)灌丛、白栎(Quercus fabri)灌丛、盐肤木(Rhus chinensis)灌丛、黄荆(Vitex negundo)灌丛、檵木(Loropetalum chinense)灌丛、马桑(Coriaria nepalensis)灌丛等(中国科学院中国植被图编辑委员会, 2007)。
图1
图1
中国中亚热带北部灌丛植物群落样点分布。
Fig. 1
Geographical distribution of samples of shrubland communities in northern mid-subtropical China.
1.2 样地调查
1.2.1 样点布设和样方设置
依据《中国植被》关于灌丛群系种类、分布及面积的描述(中国植被编辑委员会, 1980), 基于中国植被区划(中国科学院中国植被图编辑委员会, 2007), 在中亚热带北部以县为单位, 布设灌丛群落样地。布设灌丛样地时, 在满足抽样调查数量和面积的基础上, 兼顾类型, 选择灌丛群落物种组成、群落结构和生境相对均匀, 且群落连片分布、斑块面积大于100 m × 100 m的区域, 设置3个重复样方。如果灌丛群落不连片分布, 3个样方需分开布置, 每个样方连片群落面积需大于25 m × 25 m, 且3个样方的分布距离不超过250 m, 并保证样地四周有10 m以上的缓冲区。本研究共调查200个样地(图1)。每个样地沿对角线设置3个5 m × 5 m的样方。3个样方边缘两两之间最小距离为5 m, 最大距离不超过50 m, 每个样方内沿对角线布置3个1 m × 1 m小样方, 进行草本层植物群落调查。
1.2.2 灌丛群落调查
本研究于2017年6月至2019年11月开展, 按灌木层、草本层分层完成群落样方调查。记录5 m × 5 m灌木层样方内每株灌木种名、高度、基围(离地表3 cm)、冠幅(长短轴)、物候期。同时, 记录1 m × 1 m草本层小样方全部草本植物种名、平均高度、盖度。
1.2.3 土壤取样及测试
在5 m × 5 m样方对角线, 选择3个土壤采样点, 去除未分解及半分解凋落物层, 采集0-20 cm土壤样品, 混合装入自封袋或布口袋, 带回实验室进行土壤理化性质分析。采用电位法(LY/T 1239—1999)测定土壤pH, 采用重铬酸钾氧化-外加热法(LY/T 1237—1999)测定土壤有机碳含量, 采用半微量凯氏法(LY/T 1228—2015)测定土壤氮(N)含量, 采用酸溶法(LY/T 1232—2015)测定土壤磷(P)含量。
1.3 自然环境和人为干扰因子数据
依据灌丛群落样点地理坐标, 从全国≥0 ℃积温(AAT0)、≥10 ℃积温(AAT10)、年平均气温(Ta)、年降水量(Pa)和湿润指数(IM)等气候栅格数据集(中国科学院资源环境科学与数据中心,
从中国国内生产总值(GDP)空间分布和人口空间(POP)分布千米网格数据集(中国科学院资源环境科学与数据中心,
1.4 数据处理与统计分析
1.4.1 样地环境因子的描述性统计
使用R 4.2.1软件对样地环境因子进行描述性统计, 通过Pearson相关系数评估各个环境因子间的相关关系。
1.4.2 广义相异性模型(GDM)分析
使用Bray-Curtis相异性指数, 计算样点间群落成分的相异性(dij), 代表物种空间周转差异(Koleff et al., 2003)。
式中, A表示样点i和样点j所共有的物种数, B表示样点i独有的物种数, C表示样点j独有的物种数。
式中, 截距b是一个常数, k为预测变量, P为预测变量的数量, xki和xkj分别表示样点i、j的k变量的实际观测值。函数fk通过变换对每个预测变量的拟合方式, 以满足随着地理距离的增加, 群落物种相异性越大的特点(Poulin, 2003)。
将基于群落调查的物种数据与年平均气温、年降水量、地形、土壤等环境数据, 以及国内生产总值)、人口分布情况、距道路距离(dRoad)、距居民点距离(dRespt)等干扰数据, 代入GDM, 计算各因子对物种空间相异性的影响效应。采用偏回归方法, 绘制偏响应图, 区分以上因素对物种更替的单独或共同影响, 偏响应图中的曲线形状变化可指示不同梯度下预测变量对β多样性的影响, 斜率越大, 表明预测变量在此梯度上每单位变化差异越大, 物种更替速率越快(Mokany et al., 2022)。GDM在运行过程中会自动剔除对物种周转没有显著影响的预测变量(Ashcroft et al., 2010), 此时将不再有偏响应图输出(戴美霞等, 2017)。使用基于矩阵置换的gdm::varImp方法(Mokany et al., 2022), 检测预测变量的相对重要性, 预测变量的重要性被量化为置换变量与未置换之间解释的偏差变化百分比, 以上分析均在R 4.2.1软件中完成。
2 结果
2.1 地形差异与灌丛植物群落物种空间更替
表1 中国中亚热带北部灌丛样地环境因子的描述性统计
Table 1
| 环境因子 Environmental variable | 简写 Abbreviation | 平均值±标准误 Mean ± SE | 最小值 Minimum | 最大值 Maximum | 变异系数 CV (%) |
|---|---|---|---|---|---|
| ≥0 ℃积温 Accumulated temperature (≥0 °C) (°C) | AAT0 | 5 725.40 ± 54.49 | 2 045.40 | 6 715.90 | 13.46 |
| ≥10 ℃积温 Accumulated temperature (≥10 °C) (°C) | AAT10 | 5 100.49 ± 55.66 | 1 216.10 | 5 947.00 | 15.43 |
| 年平均气温 Mean annual air temperature (°C) | Ta | 15.65 ± 0.16 | 3.80 | 18.40 | 14.38 |
| 年降水量 Mean annual precipitation (mm) | Pa | 1 467.53 ± 15.78 | 1 047.50 | 1 892.90 | 15.21 |
| 湿润指数 Moisture index | IM | 67.28 ± 1.89 | 14.85 | 174.01 | 39.70 |
| 海拔 Altitude (m) | Alt | 258.98 ± 1.95 | 7.00 | 2 818.00 | 128.16 |
| 坡度 Slope (°) | Slope | 27.32 ± 1.03 | 0.00 | 80.00 | 53.18 |
| 坡向 Aspect (°) | Aspect | 159.91 ± 7.51 | 0.00 | 351.00 | 66.42 |
| 土壤有机碳含量 Soil organic carbon content (mg·g-1) | SOC | 13.99 ± 1.41 | 0.00 | 118.55 | 142.46 |
| 土壤总氮含量 Soil total nitrogen content (mg·g-1) | TN | 1.35 ± 0.10 | 0.09 | 9.07 | 100.74 |
| 土壤总磷含量 Soil total phosphorus content (mg·g-1) | TP | 0.59 ± 0.03 | 0.09 | 3.34 | 74.57 |
| 土壤pH Soil pH | pH | 5.84 ± 0.11 | 3.87 | 9.34 | 27.05 |
| 国内生产总值(104元·km-2) Gross domestic product (104 yuan·km-2) | GDP | 1 130.89 ± 135.31 | 60.00 | 21 627.00 | 169.21 |
| 人口空间分布(人·km-2) Population spatial distribution (person·km-2) | POP | 261.14 ± 15.34 | 22.30 | 2 048.36 | 83.09 |
| 到居民点的距离 Distance to resident (m) | dRespt | 630.39 ± 58.72 | 0.00 | 3 727.12 | 131.74 |
| 到道路的距离 Distance to road (m) | dRoad | 153.86 ± 45.20 | 0.00 | 2 797.37 | 415.54 |
图2
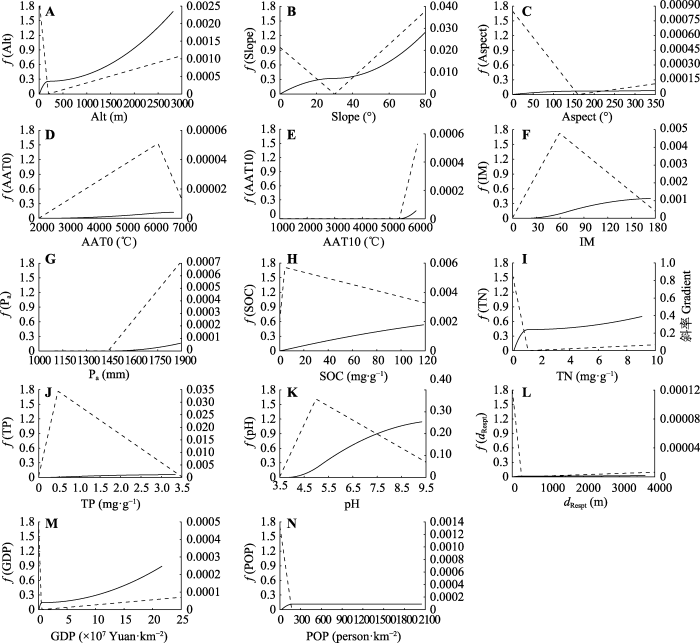
图2
中国中亚热带北部灌丛地形变量(A-C)、气候分异(D-G)、土壤理化性质(H-K)、人为干扰(L-N)与植物群落β多样性的关系。AAT0, ≥0 ℃积温; AAT10, ≥10 ℃积温; Alt, 海拔; Aspect, 坡向; dRespt, 距居民点的距离; GDP, 国内生产总值; IM, 湿润指数; Pa, 年降水量; POP, 人口空间分布; Slope, 坡度; SOC, 土壤有机碳含量; TN, 土壤总氮含量; TP, 土壤总磷含量。实线为偏响应曲线, 虚线为偏响应曲线的变化斜率, y轴f (变量)指示I-spline转换函数, 单位为群落相异性连接单位-ln(1 - dij), 其中dij指站点i和j之间的相异性。
Fig. 2
Relationship between the gradient of topographic variables (A-C), climate variables (D-G), soil physical and chemical properties (H-K), human disturbance (L-N) and β diversity of shrubland communities in northern mid-subtropical China. AAT0, accumulated temperature (≥0 °C); AAT10, accumulated temperature (≥10 °C); Alt, altitude; dRespt, distance to resident; GDP, gross domestic product; IM, moisture index; Pa, mean annual precipitation; POP, population spatial distribution; SOC, soil organic carbon content; TN, soil total nitrogen content; TP, soil total phosphorus content. Solid line is the partial response curve, and the dashed line is the change slope of the partial response curve. The function f (variable) in y axis indicated the I-spline-transformed function, and its unit was linking unit of community dissimilarity -ln(1 - dij), dij represents the dissimilarity between sites i and j.
2.2 气候分异与灌丛植物群落物种空间更替
2.3 土壤理化性质分异与灌丛植物群落物种空间更替
2.4 干扰因素与灌丛植物群落物种空间更替
图3
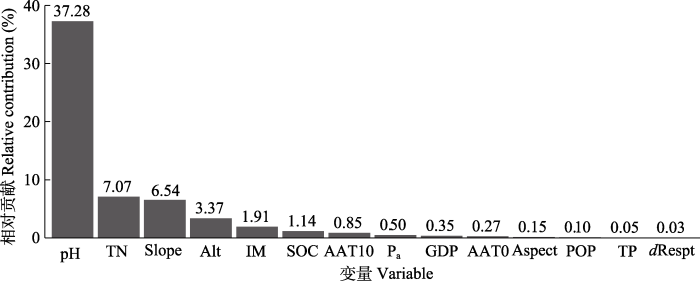
图3
预测变量对中国中亚热带北部灌丛群落间β多样性的相对贡献。AAT0, ≥0 ℃积温; AAT10, ≥10 ℃积温; Alt, 海拔; Aspect, 坡向; dRespt, 距居民点的距离; GDP, 国内生产总值; IM, 湿润指数; Pa, 年降水量; POP, 人口空间分布; Slope, 坡度; SOC, 土壤有机碳含量; TN, 土壤总氮含量; TP, 土壤总磷含量。
Fig. 3
Relative contribution of predictive variables to inter-community β diversity of shrubland communities in northern mid-subtropical China. AAT0, accumulated temperature (≥0 °C); AAT10, accumulated temperature (≥10 °C); Alt, altitude; GDP, gross domestic product; IM, moisture index; Pa, mean annual precipitation; POP, population spatial distribution; dRespt, distance to resident; SOC, soil organic carbon content; TN, soil total nitrogen content; TP, soil total phosphorus content.
2.5 环境因素和人为干扰因素对物种空间周转的解释
图4
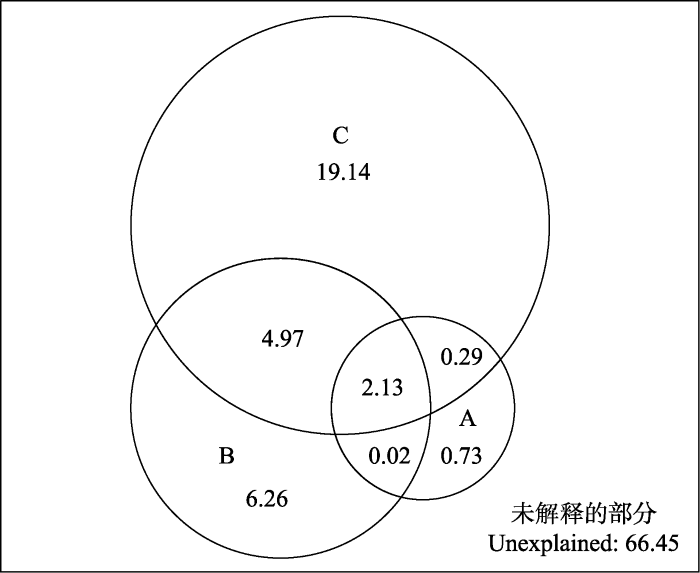
图4
人为干扰(A)、气候与地形分异(B)及土壤异质性(C)对中亚热带北部灌丛群落之间物种更替的解释度(%)。
Fig. 4
Partitional effects (%) of human disturbance (A), climate and topography difference (B) and soil difference (C) on species turnover of shruband communities in northern mid-subtropical China.
3 讨论
土壤作为植物主要的养分来源, 为植物的生长和发育提供营养物质和水分, 影响着植被群落结构与多样性特征(Xing & He, 2019; 罗巧玉等, 2021)。土壤pH通过调节土壤养分元素有效释放和转化以及根系微生物的活动, 影响物种分布(Shen et al., 2013; Zhang et al., 2020)。Zhang等(2020)在研究中发现, 土壤pH对西南草原藤本植物群落β多样性和功能多样性具有较大的影响。本研究发现, 土壤pH显著影响物种的空间更替, 当pH = 5 (弱酸性)时, 群落间物种更替速率达到最大值。这可能是因为亚热带地区土壤pH多以酸性为主(占区域取样点的65%), 植被更适宜在此种土壤中生存, 相较于过酸过碱的严苛土壤环境中趋于相似、单一的植被组成, 物种更替速率相应加快。
土壤有机碳是微生物活动的碳源和植物可利用养分的主要来源(伍旖旎等, 2022)。本研究发现, 土壤有机碳含量与土壤总氮含量显著相关(r = 0.88, 附录)。而氮、磷是植物生长发育过程中所需的重要营养元素, 土壤营养元素含量变化改变植物可利用的生态位空间, 从而影响灌丛植物群落间物种相异性(Stein et al., 2014)。Wang等(2015)研究发现, 土壤氮含量异质性是中国东北科尔沁草原植物多样性变化的主要影响因素。Zhang等(2022)研究发现, 土壤总氮含量独立解释了植物β多样性的16%。本研究发现土壤总氮含量显著影响灌丛物种周转, 与Zhang等(2022)研究结果一致。Jones等(2016)发现土壤总磷含量对澳大利亚西南部原生植被物种周转解释度最大, 但当土壤磷含量大于0.05 mg·g-1时, 其对物种更替影响则不显著。本研究发现的土壤总磷含量对物种相异性几乎没有影响的结果与之相一致。本研究发现中亚热带北部土壤总磷含量平均为(0.59 ± 0.03) mg·g-1 (表1), 远高于0.05 mg·g-1, 土壤总磷含量对灌丛物种更替影响相对较小。
海拔作为独特的地形因子, 能反映水热条件差异和微生境的共同作用, 对群落β多样性及物种更替具有显著影响(杨阳等, 2016; 赵鸣飞等, 2017)。Oishi (2021)在探究植物多样性与海拔变化的关系时发现, 亚高山交错带的灌木、草本和苔藓植物的β多样性随海拔升高而增加。Sanchez-Gonzalez和Lopez-Mata (2005)在探究墨西哥内华达山脉北部环境变量与物种多样性关系时发现, 物种β多样性沿海拔升高呈稳定增长趋势, 中间海拔相邻站点间β多样性较低, 而高海拔站点间β多样性较高(物种替换率接近100%)。本研究发现, 随海拔升高, 物种空间更替速率加快。200 m以下的低海拔地区, 能够维持较高的物种更替速率, 可能原因为人为经济生产活动增加了生境的异质性。在200-500 m的中海拔地区, 物种更替速率几乎为0, 群落结构较为稳定。随海拔的增加, 常绿灌丛向落叶灌丛加速更替, 群落间物种相异性增加。不同坡向的光照强度与光照时间存在差异, 会改变太阳辐射强度的空间分布(佘丹琦等, 2022), 坡度则影响土壤养分淋溶, 坡度、坡向等地形因素共同影响物种多样性的格局(李添甜, 2018)。Wang等(2021)研究发现, 坡度、坡向等地形因素对灌丛β多样性均具有较大的影响, 本研究结果与之相一致。此外, 本研究还发现当坡度>40°时, 物种周转速率随坡度增加逐渐增快, 这可能是因为坡度大的地形条件更容易受到干扰, 比如土壤侵蚀或大风, 通过改变资源的可利用性, 导致物种周转速率加快(Chapman & McEwan, 2013)。
气候因素是决定群落间物种多样性空间格局的重要因素(dos Santos et al., 2015; Hamid et al., 2019)。Liu等(2015)研究发现, 随着气温升高, 中国温带落叶阔叶林物种β多样性增加。Zhang等(2014)发现草原物种β多样性随降水量增加而显著增加。本研究发现≥0 ℃积温、≥10 ℃积温、年降水量、湿润指数等气候因素均与灌丛群落物种相异性呈显著的相关关系, 但气候因素差异对物种周转的解释度没有占主导地位, 可能是因为亚热带地区并没有明显的生境限制, 如湿润指数平均值为67.28, 表明中亚热带北部地区为潮湿区, 水热条件较为充足(李家湘等, 2017)。本研究同时发现, 气候与地形因子和土壤因子交互作用, 共同解释了中亚热带北部灌丛物种相异性的32.82%。这可能是因为水热要素影响土壤微生物活性, 改变土壤养分状态, 进而影响物种更替(Qian et al., 2017)。
亚热带地区高强度的农业生产活动及经济建设, 使森林遭到频繁的砍伐, 土地利用类型和土地覆盖度发生了显著改变, 物种生境退化或破碎化, 物种丰富度下降甚至灭绝, 物种相异性降低(中国植被编辑委员会, 1980; 陈圣宾等, 2010; 段语凤等, 2020)。王应刚等(2016)在探究晋中盆地人口聚集地与植物多样性关系时发现, 植物β多样性沿着村庄-乡镇-县市梯度降低。城镇化建设会对植被群落造成人为干扰, 改变原生植被生境(郭正刚等, 2004)。本研究发现, 随着GDP的增加, 物种周转速率加快, 表明人为干扰加剧了资源的异质性, 物种生境受土地利用类型的改变退化或丧失, 物种适生范围缩小而面临被淘汰的风险, 物种更替加快。
GDM模型的解释度范围为20%-50% (Mokany et al., 2022)。姜小蕾等(2020)使用GDM模型对崂山次生林群落β多样性格局及其驱动因素的研究发现, 11个环境变量共同解释了灌木植物β多样性的28.38%。本研究通过GDM模型发现, 14个环境变量解释了灌丛植物群落物种周转的33.55%。季节性气候影响物候, 进而决定物种的空间分布(Bykova et al., 2012)。Davidar等(2007)发现, 季节性气候是山地雨林树木β多样性的关键驱动因素。此外, 生物因素, 如群落内物种种间关系, 也是驱动物种周转的主要因素。佘丹琦等(2022)发现, 灌木高度和地径大小显著影响灌木层β多样性。增加最暖季平均气温和降水季节性等表征季节性气候的非生物因素和生物因素等环境变量, 预计可提高对灌丛群落物种周转的解释率, 这也是今后研究需关注的重点。
中亚热带作为重要的植被恢复区, 探究其灌丛群落物种空间周转的驱动因素, 能为该地区生物多样性保护和恢复提供依据。本研究发现环境因子在中亚热带北部灌丛植物空间周转过程中具有重要作用, 其中土壤pH等因子是决定中亚热带北部灌丛植物群落周转的主要因素。
附录 中国中亚热带北部灌丛群落不同环境因子之间的相关性
Supplement Correlation between different environmental factors of shrubland communities in northern mid-subtropical China
致谢
感谢植被与环境变化国家重点实验室研究群课题的资助。
参考文献
Using Generalised Dissimilarity Models and many small samples to improve the efficiency of regional and landscape scale invertebrate sampling
DOI:10.1016/j.ecoinf.2009.12.002 URL [本文引用: 1]
Linking global turnover of species and environments
DOI:10.1073/pnas.0803524105
PMID:19001274
[本文引用: 1]

Patterns of species turnover are central to the geography of biodiversity and resulting challenges for conservation, but at broad scales remain relatively little understood. Here, we take a first spatially-explicitly and global perspective to link the spatial turnover of species and environments. We compare how major groups of vertebrate ectotherms (amphibians) and endotherms (birds) respond to spatial environmental gradients. We find that high levels of species turnover occur regardless of environmental turnover rates, but environmental turnover provides a lower bound for species turnover. This lower bound increases more steeply with environmental turnover in tropical realms. While bird and amphibian turnover rates are correlated, the rate of amphibian turnover is four times steeper than bird rates. This is the same factor by which average geographic ranges of birds are larger than those of amphibians. Narrow-ranged birds exhibit rapid rates of species turnover similar to those for amphibians, while wide-ranged birds largely drive the aggregate patterns of avian turnover. We confirm a strong influence of the environment on species turnover that is mediated by range sizes and regional history. In contrast to geographic patterns of species richness, we find that the turnover in one group (amphibians) is a much better predictor for the turnover in another (birds) than is environment. This result confirms the role of amphibian sensitivity to environmental conditions for patterns of turnover and supports their value as a surrogate group. This spatially-explicit analysis of environmental turnover provides understanding for conservation planning in changing environments.
Temperature dependence of the reproduction niche and its relevance for plant species distributions
DOI:10.1111/j.1365-2699.2012.02764.x URL [本文引用: 1]
Winter cold tolerance and the geographic range separation of Bromus tectorum and Bromus rubens, two severe invasive species in North America
DOI:10.1111/gcb.12003 URL [本文引用: 1]
Spatiotemporal dynamics of α- and β-diversity across topographic gradients in the herbaceous layer of an old-growth deciduous forest
DOI:10.1111/j.1600-0706.2013.00544.x URL [本文引用: 1]
A review of beta diversity studies
DOI:10.3724/SP.J.1003.2010.323
[本文引用: 3]
<p>Beta diversity is an important component of biological diversity, measuring compositional change in species assemblages across temporal and spatial scales. Beta diversity concerns not only a number of ecological and evolutionary issues, but can also guide the selection of protected areas and help to optimize conservation networks. It has thus become a hot topic in biodiversity research in recent years. Researchers have used various measures and analytical methods to investigate patterns of beta diversity and its underlying mechanisms for various taxa and in different regions. Here, we reviewed literature from the past decade pertaining to the following aspects of beta diversity: metrics, temporal and spatial patterns, determinants and applications in biodiversity conservation. Whittaker introduced the term beta diversity in 1960, but defined it vaguely. As the concept of beta diversity evolved, a high variety of measures were developed to quantify the concept. The comparison of results from different studies may be hindered by the variety of measures used to quantify beta diversity. Presently, the most popular methods for measuring beta diversity are similarity/ dissimilarity coefficients such as Jaccard index and Sørensen index. In the last few years, several methods to quantify beta diversity have emerged, some of which are worth noting. Beta diversity depends on temporal scale, spatial scale and taxonomic scale, and decreases with increasing analytical grain size. There is no consensus among scientists that beta diversity decreases with latitude, i.e. that it is higher in tropics and lower near the poles. Beta diversity is high within mountain ranges and at the interface of biogeographic realms; thus, larger/more reserves are needed in these regions to cover the entire gradient of species turnover. Studies on beta diversity across temporal scales have shown that climatic change has resulted in shifts in species composition through time, and that the migration of species between different continents/regions has led to biotic homogenization. Based on a thorough review of beta diversity literature, we think the following questions might be the focus of future research: (1) the influence of evolutionary history and biological characteristics of different taxonomic groups on their beta diversity; (2) the influence of temporal/spatial scales on beta diversity and its determinants; and (3) the effect of anthropogenic activities on beta diversity.</p>
Beta多样性研究进展
DOI:10.3724/SP.J.1003.2010.323
[本文引用: 3]
Beta多样性度量时空尺度上物种组成的变化, 是生物多样性的重要组成部分, 与许多生态学和进化生物学问题密切相关, 并且其信息可用于保护区选址和布局规划, 因此在最近10年间成为生物多样性研究的热点问题之一。多年来, 学者们利用各种度量方式和分析方法, 在不同地理区域, 对许多生物类群beta多样性的时空格局和形成机制进行了大量研究。本文主要从beta多样性的度量方法、时空格局、形成机制及其在生物多样性保护中的应用等几个方面, 总结了最近10多年来相关研究的进展。Whittaker(1960)最初提出beta多样性概念时就缺乏严格的定义, 随着概念的不断演化, 度量方法也同样呈现出多样化, 而度量手段的多样化非常不利于不同研究之间的比较。目前应用最普遍的度量方法是采用相似性指数, 如Jaccard和Sørensen指数。最近几年, 新的度量方法还在不断出现, 其中一些方法非常值得注意。Beta多样性具有时空尺度和分类尺度依赖性, 一般随分析粒度(grain)的增加而降低。虽然有些研究表明beta多样性随纬度增加而降低, 但学者们并没有达成共识。山区和生物地理区的交界处beta多样性都比较高, 因而需要在这些地区增加保护区的面积或者数量以囊括物种变化梯度。对时间尺度上beta多样性的研究表明, 气候变化确实导致了物种组成在时间上的变化, 并且物种在不同大陆和地区间的迁移导致了生物同质化。扩散过程和生态位过程共同决定了beta多样性, 只是这两个过程的相对重要性依尺度、地理区域和物种类群的不同而有所差异。综上所述, 我们认为未来beta多样性研究的热点问题是:(1)不同生物类群的进化历史和生物学特征对beta多样性的影响; (2)不同的时空尺度对beta多样性及其维持机制的影响; (3)人类活动对beta多样性的影响。
Patterns of phylogenetic beta diversity in China’s grasslands in in relation to geographical and environmental distance
DOI:10.1016/j.baae.2014.07.003 URL [本文引用: 1]
Why does phenology drive species distribution?
DOI:10.1098/rstb.2010.0142 URL [本文引用: 1]
Interpretation of environmental factors affecting zooplanktonic beta diversity and its components in Xiangshan Bay.
象山港浮游动物β多样性及其成分变化的环境因子解释
Stochastic and deterministic processes drive wetland community assembly across a gradient of environmental filtering
DOI:10.1111/oik.05987
[本文引用: 1]
The role of deterministic and stochastic processes in community assembly is a key question in community ecology. We evaluated the effect of an abiotic filter (hydroperiod) on the partitioned diversity of three taxonomic groups (birds, vegetation, macroinvertebrates) from prairie pothole wetlands in Alberta, Canada, which naturally vary in water permanence. We observed that alpha and gamma diversity were higher in permanent than temporary wetlands (16-25% and 34-47% respectively, depending on the taxon). This suggests an influence of deterministic constraints on the number of species a wetland can support. Taxa which cannot persist in shallow, temporary wetlands are excluded by the deterministic constraints that a shortened hydroperiod imposes. In contrast, we observed that beta diversity was significantly higher (2-12%) in temporary wetlands than permanent ones, and temporary wetlands supported more unique combinations of community composition than permanent wetlands, despite having a smaller regional species pool. This observation contradicts prior mesocosm studies that found beta diversity mirrored the pattern in gamma diversity along an environmental filtering gradient. We conclude that deterministic processes are more influential in more stable permanent wetlands, whereas stochastic processes play a more important role in assembly in dynamic temporary wetlands that must disassemble and re-establish annually. Considering three distinct taxonomic groups differing in their relative mobility, our large-scale field study demonstrates that both stochastic and deterministic processes act together to influence the assembly of multiple communities and that the relative importance of the two processes varies consistently along a gradient of environmental filtering.
The effect of climatic gradients, topographic variation and species traits on the beta diversity of rain forest trees
DOI:10.1111/j.1466-8238.2007.00307.x URL [本文引用: 1]
Interactive effects of environmental filtering predict beta-diversity patterns in a subtropical forest metacommunity
DOI:10.1016/j.ppees.2015.01.002 URL [本文引用: 1]
Effects of the underground coal mining on the dynamic changes of vegetation in arid desert area
煤炭井工开采对干旱荒漠区植被动态变化的影响
Anthropogenic disturbance shapes phylogenetic and functional tree community structure in a subtropical forest
DOI:10.1016/j.foreco.2013.10.047 URL [本文引用: 1]
Using generalized dissimilarity modelling to analyse and predict patterns of beta diversity in regional biodiversity assessment
DOI:10.1111/j.1472-4642.2007.00341.x URL [本文引用: 1]
Distribution patterns and climate limitations of typical shrublands in northern China
中国北方典型灌丛的分布特征及气候限制
Effect of the Qinghai-Tibetan highway on the β diversity of grassland plant communities in the northern region of the Qinghai- Tibetan Plateau
人类工程对青藏高原北部草地群落β多样性的影响
Habitat fragmentation and its lasting impact on Earth’s ecosystems
DOI:10.1126/sciadv.1500052 [本文引用: 1]
Impact of climate change on the distribution range and niche dynamics of Himalayan birch, a typical treeline species in Himalayas
DOI:10.1007/s10531-018-1641-8 URL [本文引用: 1]
Interpretation of environmental factors affecting beta diversity and its components of secondary forest in Lao Mountain
崂山次生林群落β多样性格局及其组分的驱动因素
Underestimated effects of climate on plant species turnover in the Southwest Australian Floristic Region
DOI:10.1111/jbi.12628 URL [本文引用: 1]
Measuring beta diversity for presence-absence data
DOI:10.1046/j.1365-2656.2003.00710.x URL [本文引用: 1]
Partitioning beta diversity in a subtropical broad-leaved forest of China
DOI:10.1890/07-1880.1
PMID:19341137
[本文引用: 1]
The classical environmental control model assumes that species distribution is determined by the spatial variation of underlying habitat conditions. This niche-based model has recently been challenged by the neutral theory of biodiversity which assumes that ecological drift is a key process regulating species coexistence. Understanding the mechanisms that maintain biodiversity in communities critically depends on our ability to decompose the variation of diversity into the contributions of different processes affecting it. Here we investigated the effects of pure habitat, pure spatial, and spatially structured habitat processes on the distributions of species richness and species composition in a recently established 24-ha stem-mapping plot in the subtropical evergreen broad-leaved forest of Gutianshan National Nature Reserve in East China. We used the new spatial analysis method of principal coordinates of neighbor matrices (PCNM) to disentangle the contributions of these processes. The results showed that (1) habitat and space jointly explained approximately 53% of the variation in richness and approximately 65% of the variation in species composition, depending on the scale (sampling unit size); (2) tree diversity (richness and composition) in the Gutianshan forest was dominantly controlled by spatially structured habitat (24%) and habitat-independent spatial component (29%); the spatially independent habitat contributed a negligible effect (6%); (3) distributions of richness and species composition were strongly affected by altitude and terrain convexity, while the effects of slope and aspect were weak; (4) the spatial distribution of diversity in the forest was dominated by broad-scaled spatial variation; (5) environmental control on the one hand and unexplained spatial variation on the other (unmeasured environmental variables and neutral processes) corresponded to spatial structures with different scales in the Gutianshan forest plot; and (6) five habitat types were recognized; a few species were statistically significant indicators of three of these habitats, whereas two habitats had no significant indicator species. The results suggest that the diversity of the forest is equally governed by environmental control (30%) and neutral processes (29%). In the fine-scale analysis (10 x 10 m cells), neutral processes dominated (43%) over environmental control (20%).
The problem of pattern and scale in ecology
DOI:10.2307/1941447 URL [本文引用: 1]
Leaf nitrogen and phosphorus concentration and the empirical regulations in dominant woody plants of shrublands across southern China
中国南方灌丛优势木本植物叶的氮、磷含量及其影响因素
DOI:10.17521/cjpe.2016.0251
[本文引用: 1]
探索植物器官中氮(N)、磷(P)含量沿环境梯度的分异规律, 有助于揭示陆地生态系统物质循环和植物养分适应策略的过程和机制。该文采用分层随机抽样法, 在中国南方12个省市区布设462个灌丛样点, 对其中193种优势木本植物叶N、P含量进行调查取样。结果表明: 1)南方灌丛优势木本植物叶的N、P含量几何均值分别为16.57 mg·g<sup>-1</sup>和1.02 mg·g<sup>-1</sup>; 其中, 落叶木本植物(17.91 mg·g<sup>-1</sup>、1.14 mg·g<sup>-1</sup>)显著高于常绿木本植物(15.19 mg·g<sup>-1</sup>、0.89 mg·g<sup>-1</sup>); 叶P含量较N含量具有更大的变异性和环境依赖性。2)随年平均气温(MAT)的升高, 常绿木本植物叶N、P含量降低, 落叶木本植物叶N、P含量呈逐渐增加的趋势; 随年降水量(MAP)的增加, 常绿木本植物叶N含量降低, 落叶木本植物叶N含量增加, 两者P含量下降。3)土壤N含量的增加对两者叶的N含量无显著影响; 但随土壤P含量的增加, 常绿和落叶木本植物叶P含量均显著增加。4)广义线性回归显示, 植物生活型分别可解释叶N、P变异的7.6%和14.4%, MAP和土壤P含量分别解释了0.8%和16.4%的叶P变异。结果表明, 中国南方灌丛中优势植物叶的N含量主要受不同生活型植物生长需求所决定, 而P含量则受气候、土壤和植物生活型共同决定。
Species biodiversity of seven typical shrub communities in the middle of Guangxi Zhuang Autonomous Region
广西中部7种典型灌丛群落的物种多样性特征
Contribution of environmental filtering and dispersal limitation to species turnover of temperate deciduous broad-leaved forests in China
DOI:10.1111/avsc.12101 URL [本文引用: 1]
Plant community diversity and soil factor interpretation of adaptive region of Deschampsia caespitosa in the source region of the Yellow River
黄河源区发草适生地植物群落特征及其土壤因子解释
Insights into ecosystem composition and function in a sequence of degraded semiarid steppes
DOI:10.1111/j.1061-2971.2004.03106.x URL [本文引用: 1]
Disentangling the functional trait correlates of spatial aggregation in tropical forest trees
DOI:10.1002/ecy.2591 [本文引用: 1]
A working guide to harnessing generalized dissimilarity modelling for biodiversity analysis and conservation assessment
DOI:10.1111/geb.13459 URL [本文引用: 3]
Mechanisms of community assembly explaining beta- diversity patterns across biogeographic regions
DOI:10.1111/jvs.13032 [本文引用: 1]
Factors that shape the elevational patterns of plant diversity in the Yatsugatake Mountains, Japan
DOI:10.1002/ece3.7397
PMID:33976856
[本文引用: 1]
Elevation is involved in determining plant diversity in montane ecosystems. This study examined whether the distribution of plants in the Yatsugatake Mountains, central Japan, substantiated hypotheses associated with an elevational diversity gradient. Species richness of trees, shrubs, herbs, ferns, and bryophytes was investigated in study plots established at 200-m elevational intervals from 1,800 to 2,800 m. The changes in plant diversity (alpha and beta diversities, plant functional types, and elevational ranges) with elevation were analyzed in relation to climatic factors and elevational diversity gradient hypotheses, that is, mass effect, mid-domain effect, and Rapoport's elevational rule. In addition, the elevational patterns of dominance of plant functional types were also analyzed. A comparison of alpha and beta diversities revealed that different plant groups responded variably to elevation; the alpha diversity of trees and ferns decreased, that of herbs increased, whereas the alpha diversity of shrubs and bryophytes showed a U-shaped relationship and a hump-shaped pattern. The beta diversity of shrubs, herbs, and bryophytes increased above the subalpine-alpine ecotone. In accordance with these changes, the dominance of evergreen shrubs and graminoids increased above this ecotone, whereas that of evergreen trees and liverworts decreased. None of the plant groups showed a wide elevational range at higher elevations. These elevational patterns of plant groups were explained by climatic factors, and not by elevational diversity gradient hypotheses. Of note, the changes in the dominance of plant groups with elevation can be attributed to plant-plant interactions via competition for light and the changes in physical habitat. These interactions could alter the elevational diversity gradient shaped by climatic factors.© 2021 The Authors. Ecology and Evolution published by John Wiley & Sons Ltd.
Estimating and conserving patterns of invertebrate diversity: a test case of New Zealand land snails
DOI:10.1111/j.1472-4642.2009.00589.x URL [本文引用: 1]
Soil nutrients and beta diversity in the Bornean Dipterocarpaceae: evidence for niche partitioning by tropical rain forest trees
DOI:10.1111/j.1365-2745.2005.01077.x URL [本文引用: 1]
The carbon balance of terrestrial ecosystems in China
DOI:10.1038/nature07944 URL [本文引用: 1]
Improving the use of species distribution models in conservation planning and management under climate change
DOI:10.1371/journal.pone.0113749 [本文引用: 1]
The decay of similarity with geographical distance in parasite communities of vertebrate hosts
DOI:10.1046/j.1365-2699.2003.00949.x URL [本文引用: 1]
Phylogenetic diversity anomaly in angiosperms between eastern Asia and eastern North America
DOI:10.1073/pnas.1703985114
PMID:29073071
[本文引用: 1]
Although eastern Asia (EAS) and eastern North America (ENA) have similar climates, plant species richness in EAS greatly exceeds that in ENA. The degree to which this diversity difference reflects the ages of the floras or their rates of evolutionary diversification has not been quantified. Measures of species diversity that do not incorporate the ages of lineages disregard the evolutionary distinctiveness of species. In contrast, phylogenetic diversity integrates both the number of species and their history of evolutionary diversification. Here we compared species diversity and phylogenetic diversity in a large number of flowering plant (angiosperm) floras distributed across EAS and ENA, two regions with similar contemporary environments and broadly shared floristic history. After accounting for climate and sample area, we found both species diversity and phylogenetic diversity to be significantly higher in EAS than in ENA. When we controlled the number of species statistically, we found that phylogenetic diversity remained substantially higher in EAS than in ENA, although it tended to converge at high latitude. This pattern held independently for herbs, shrubs, and trees. The anomaly in species and phylogenetic diversity likely resulted from differences in regional processes, related in part to high climatic and topographic heterogeneity, and a strong monsoon climate, in EAS. The broad connection between tropical and temperate floras in southern Asia also might have played a role in creating the phylogenetic diversity anomaly.
Plant communities of New Brunswick in relation to environmental variation
DOI:10.2307/3237061 URL [本文引用: 1]
Plant species richness and diversity along an altitudinal gradient in the Sierra Nevada, Mexico
DOI:10.1111/j.1366-9516.2005.00186.x URL [本文引用: 1]
Plant beta diversity and its influence factors in the Liangshui National Nature Reserve in the central region of the Xiaoxing’an Mountains
小兴安岭凉水国家级自然保护区植物beta多样性及其影响因素
Soil pH drives the spatial distribution of bacterial communities along elevation on Changbai Mountain
DOI:10.1016/j.soilbio.2012.07.013 URL [本文引用: 1]
A multi-scale study on the vegetation-environment relationship of a mountain forest transect
山地森林样带植被-环境关系的多尺度研究
A multivariate analysis of beta diversity across organisms and environments
We examined variability in hierarchical beta diversity across ecosystems, geographical gradients, and organism groups using multivariate spatial mixed modeling analysis of two independent data sets. The larger data set comprised reported ratios of regional species richness (RSR) to local species richness (LSR) and the second data set consisted of RSR:LSR ratios derived from nested species-area relationships. There was a negative, albeit relatively weak, relationship between beta diversity and latitude. We found only relatively subtle differences in beta diversity among the realms, yet beta diversity was lower in marine systems than in terrestrial or freshwater realms. Beta diversity varied significantly among organisms' major characteristics such as body mass, trophic position, and dispersal type in the larger data set. Organisms that disperse via seeds had highest beta diversity, and passively dispersed organisms showed the lowest beta diversity. Furthermore, autotrophs had lower beta diversity than organisms higher up the food web; omnivores and carnivores had consistently higher beta diversity. This is evidence that beta diversity is simultaneously controlled by extrinsic factors related to geography and environment, and by intrinsic factors related to organism characteristics.
Environmental heterogeneity as a universal driver of species richness across taxa, biomes and spatial scales
DOI:10.1111/ele.12277
PMID:24751205
[本文引用: 1]
Environmental heterogeneity is regarded as one of the most important factors governing species richness gradients. An increase in available niche space, provision of refuges and opportunities for isolation and divergent adaptation are thought to enhance species coexistence, persistence and diversification. However, the extent and generality of positive heterogeneity-richness relationships are still debated. Apart from widespread evidence supporting positive relationships, negative and hump-shaped relationships have also been reported. In a meta-analysis of 1148 data points from 192 studies worldwide, we examine the strength and direction of the relationship between spatial environmental heterogeneity and species richness of terrestrial plants and animals. We find that separate effects of heterogeneity in land cover, vegetation, climate, soil and topography are significantly positive, with vegetation and topographic heterogeneity showing particularly strong associations with species richness. The use of equal-area study units, spatial grain and spatial extent emerge as key factors influencing the strength of heterogeneity-richness relationships, highlighting the pervasive influence of spatial scale in heterogeneity-richness studies. We provide the first quantitative support for the generality of positive heterogeneity-richness relationships across heterogeneity components, habitat types, taxa and spatial scales from landscape to global extents, and identify specific needs for future comparative heterogeneity-richness research.© 2014 John Wiley & Sons Ltd/CNRS.
An updated vegetation map of China (1:1000000)
DOI:10.1016/j.scib.2020.04.004
PMID:36659164
[本文引用: 2]
Vegetation maps are important sources of information for biodiversity conservation, ecological studies, vegetation management and restoration, and national strategic decision making. The current Vegetation Map of China (1:1000000) was generated by a team of more than 250 scientists in an effort that lasted over 20 years starting in the 1980s. However, the vegetation distribution of China has experienced drastic changes during the rapid development of China in the last three decades, and it urgently needs to be updated to better represent the distribution of current vegetation types. Here, we describe the process of updating the Vegetation Map of China (1:1000000) generated in the 1980s using a "crowdsourcing-change detection-classification-expert knowledge" vegetation mapping strategy. A total of 203,024 field samples were collected, and 50 taxonomists were involved in the updating process. The resulting updated map has 12 vegetation type groups, 55 vegetation types/subtypes, and 866 vegetation formation/sub-formation types. The overall accuracy and kappa coefficient of the updated map are 64.8% and 0.52 at the vegetation type group level, 61% and 0.55 at the vegetation type/subtype level and 40% and 0.38 at the vegetation formation/sub-formation level. When compared to the original map, the updated map showed that 3.3 million km of vegetated areas of China have changed their vegetation type group during the past three decades due to anthropogenic activities and climatic change. We expect this updated map to benefit the understanding and management of China's terrestrial ecosystems.Copyright © 2020 Science China Press. Published by Elsevier B.V. All rights reserved.
Vegetation of China
中国植被
Climate and soil attributes determine plant species turnover in global drylands
DOI:10.1111/jbi.12377
PMID:25914437
[本文引用: 1]
Geographic, climatic, and soil factors are major drivers of plant beta diversity, but their importance for dryland plant communities is poorly known. This study aims to: i) characterize patterns of beta diversity in global drylands, ii) detect common environmental drivers of beta diversity, and iii) test for thresholds in environmental conditions driving potential shifts in plant species composition.224 sites in diverse dryland plant communities from 22 geographical regions in six continents.Beta diversity was quantified with four complementary measures: the percentage of singletons (species occurring at only one site), Whittake's beta diversity (β(W)), a directional beta diversity metric based on the correlation in species occurrences among spatially contiguous sites (β(R)), and a multivariate abundance-based metric (β(MV)). We used linear modelling to quantify the relationships between these metrics of beta diversity and geographic, climatic, and soil variables.Soil fertility and variability in temperature and rainfall, and to a lesser extent latitude, were the most important environmental predictors of beta diversity. Metrics related to species identity (percentage of singletons and β(W)) were most sensitive to soil fertility, whereas those metrics related to environmental gradients and abundance ((β(R)) and β(MV)) were more associated with climate variability. Interactions among soil variables, climatic factors, and plant cover were not important determinants of beta diversity. Sites receiving less than 178 mm of annual rainfall differed sharply in species composition from more mesic sites (> 200 mm).Soil fertility and variability in temperature and rainfall are the most important environmental predictors of variation in plant beta diversity in global drylands. Our results suggest that those sites annually receiving ~ 178 mm of rainfall will be especially sensitive to future climate changes. These findings may help to define appropriate conservation strategies for mitigating effects of climate change on dryland vegetation.
The different altitude gradient change rules of the main shrub community in arid valleys of the Bailongjiang River with different slope
DOI:10.7525/j.issn.1673-5102.2018.01.004
[本文引用: 1]
The change rules of different slope on the structure characteristics and species diversity of the Main shrub community in arid valleys of the Bailongjiang River were studied with Quadrat survey method. Aim to understand the vegetation characteristic and Species diversity changes from different elevation gradient of arid valleys in bailongjiang river, It will provides the theory basis for vegetation restoration in this region.Results show that: (1)species are different in the same slope of different altitude gradient, as well as in the different slope of same altitude gradient. with the increase of altitude,the species of different slope show that increased after decreased first, The main vegetation types are also different in the same slope of different altitude gradient; (2)With the increase of altitude gradient, α-diversity of shrub community and α-diversity of herb community in different slope all showed that a trend from rise to decline. Correlation analysis was carried out on the α-diversity index, contribution rate of species diversity for the species diversity is the largest,it is showed that the richness index(D<sub>1</sub>, D<sub>2</sub>)> ecological dominance index(SN)> interspecific opportunity index(H)> evenness index of community(R); (3)With the change of altitude gradient, β-diversity Whittaker index of the main shrub community have less change in different slope, the maximum appeared at an altitude of 1 250-1 650 m,Routledge and Cody β-diversity index between an altitude of 1 450-1 650 m reache maximum, but generally presents the waveform changes. β-diversity of herb community have larger change, At an altitude of 1 050-1 250 m, β-diversity of herb community in the sunny slope reaches maximum. At an altitude of 1 250-1 450 m,β-diversity index of herb community in the semi-shady slope and semi-sunny slope reaches maximum, β-diversity index of herb community in the semi-shady slope and semi-sunny slope were higher than sunny slope. Species diversity α and diversity β are different that in Different slope directions and different altitudinal gradients in dry valleys of Bai Long River and different slope directions have a certain correlation with the change of altitudinal gradient, species diversity α and diversity β indicating that elevation and slope directions are one of the main factors affecting biodiversity.
白龙江干旱河谷不同坡向主要灌丛群落随海拔梯度变化的物种多样性研究
DOI:10.7525/j.issn.1673-5102.2018.01.004
[本文引用: 1]
采用样方调查法,研究了白龙江干旱河谷不同坡向主要灌丛群落沿着海拔梯度的结构特征、物种多样性的变化规律,旨在了解白龙江干旱河谷不同海拔梯度植被特征和物种多样性变化,为白龙江干旱河谷区域不同海拔植被恢复提供理论依据。研究结果表明:(1)不同海拔梯度同一坡向物种数不同,同一海拔不同坡向物种数也不同,随着海拔的升高不同坡向物种数表现为先增加后减少的趋势,同一海拔梯度内不同坡向主要植被类型也不同。(2)主要灌木群落α多样性在不同坡向随着海拔梯度的升高,表现出先升高后减小的趋势。不同坡向草本群落α多样性随着海拔的升高,也表现出先升高后减小的趋势。对主要灌丛α多样性指数进行相关性分析得物种丰富度指数对物种多样性贡献率最大,表现为丰富度指数(D<sub>1</sub>、D<sub>2</sub>)> 生态优势度指数(SN)> 种间机遇指数(H)> 群落均匀度指数(R)。(3)不同坡向主要灌丛群落β多样性Whittaker指数沿着不同海拔梯度变化不大,最大值出现在海拔1250~1650m;Routledge和Codyβ多样性指数在海拔1450~1650m出现最大值,但是大体呈现出波形变化。草本β多样性随着海拔的升高变化较大,阳坡植物的β多样性指数在海拔1050~1250m达到最大,阴坡和半阴半阳坡在海拔区间1250~1450m达到最大,半阴半阳坡的β多样性指数均大于阳坡。白龙江干旱河谷不同坡向、不同海拔梯度物种α多样性和β多样性都不同,且不同坡向随着海拔梯度的变化物种α多样性和β多样性呈一定的相关性,说明海拔和坡向是影响生物多样性主要因子之一。
Divergent roles of environmental and spatial factors in shaping plant β-diversity of different growth forms in drylands
DOI:10.1016/ j.gecco.2021.e01487 [本文引用: 1]
Plant diversity is associated with the amount and spatial structure of soil heterogeneity in meadow steppe of China
DOI:10.1007/s10980-013-9955-0 URL [本文引用: 1]
Patterns of plant diversity and population distribution in the Jinzhong Basin
晋中盆地人类聚居地植物多样性分异规律
Effects of nitrogen application on organic carbon mineralization in rhizosphere soil of Miscanthus spp. in the barren red soil
施氮对贫瘠红壤定植芒草根际土有机碳矿化过程的影响
DOI:10.11733/j.issn.1007-0435.2022.04.004
[本文引用: 1]
为了探究不同施氮量对长期定植芒属植物后酸性红壤中固定的土壤有机碳矿化过程的影响,本研究以长期定植芒草的根际红壤为试验材料,采用控制变量法开展土壤有机碳矿化培养试验。结果表明:施氮极显著降低了土壤有机碳矿化量,随着施氮量的升高而降低,呈现极显著的负激发效应(P-1</sup>,42.59 mg·kg<sup>-1</sup>和33.14 mg·kg<sup>-1</sup>。本研究发现,高水平施氮会降低土壤pH值、可溶性有机碳,从而抑制红壤有机碳矿化过程,进一步稳定芒属植物种植对酸性红壤有机碳的积累效应。
Environmental filtering explains a U-shape latitudinal pattern in regional beta-deviation for eastern North American trees
DOI:10.1111/ele.13188 URL [本文引用: 1]
A comparison of the altitudinal patterns in plant species diversity within the dry valleys of the Three Parallel Rivers region, northwestern Yunnan
DOI:10.17520/biods.2015361
[本文引用: 1]
<p>In the typical dry valleys of the Three Parallel Rivers region, northwest Yunnan Province, we investigated vegetation using six sampling transects, each comprised of ten standard plots, along altitudinal gradients on the eastern and the western aspects of Nu River, Langcang River, and Jinsha River. With these data, we compared the elevation-related distribution of plant species richness and species turnover rates along the six transects, and explained the patterns using geography and vegetation variables. The dry-warm vegetation zone was dominated by shrubs and herbs and located below the altitude of 3,000 m a.s.l. At higher elevations, shrub and herbs were replaced with a forest zone. The spatial distribution of plant species richness increased with elevation and latitude, especially for herb and shrub species, and was also related to river, vegetation zones, and longitude. The species richness of shrubs also increased significantly across the region, from west to east. Species richness of herbs and shrubs in the Nu River were higher than those in the Lancang River and the Jinsha River, whereas the difference of species richness among three rivers was not significant for trees. Herb species richness in the forest zone was less than that in the shrub and herb zones. Species turnover rate of different zones presented inconsistent altitudinal gradient patterns, but all peak values appeared in the ecotones between shrub communities, in the lower altitudes of the transect, and forest communities, in the higher altitudes. The forest-shrub ecotone is located at an altitude range of 1,900-2,100 m in the Nu River valley, at an altitude range of 2,300-2,400 m in the Lancang River valley and at an altitude range of 2,700-2,900 m in the Jinsha River valley. The mean species turnover rates between shrub & herb section and forest section within each transect were less than the mean turnover rates of the same vegetation section between different transects within the same basin, and also less than the mean turnover rates for same vegetation section in all six transects. Spatial isolation could explain 34.2% of the variation in species turnover rate among the 12 vegetation sections of the six altitudinal transects, while vegetation differences explained less than 0.5% of the variation. These results show the primary role of environment difference in determining the species richness between vegetation types, whereas geographical isolation between the rivers as a dominant factor in the assembly (e.g. species composition) of plant communities.</p>
三江并流地区干旱河谷植物物种多样性海拔梯度格局比较
DOI:10.17520/biods.2015361
[本文引用: 1]
在滇西北三江并流地区典型干旱河谷段, 在怒江、澜沧江和金沙江的东、西坡共设置了6条海拔梯度样带, 通过标准样地的植物群落调查, 分析各条样带植物的物种丰富度、物种更替率的海拔梯度格局, 并比较了地理和植被变量对分布格局的解释。干旱河谷植被带位于海拔3,000 m以下, 以灌丛和灌草丛为主, 其在各河谷的分布上限自西向东依次升高。植物物种丰富度的分布主要与海拔、流域、经纬度和植被带有关, 沿纬度和海拔梯度升高而显著增加的格局主要表现在草本层和灌木层, 灌木物种丰富度还呈现自西向东显著增加的趋势。怒江的灌木和草本种物种丰富度显著高于金沙江和澜沧江, 三条江的乔木种丰富度差异则不显著。森林带的样方草本物种丰富度显著低于灌草丛带样方, 并且还拥有后者没有的乔木种。不同样带的植物物种更替速率呈现了不一致的海拔梯度格局, 但均在样带海拔下部的灌草丛群落与海拔上部森林群落之间的交错带出现峰值。森林-灌草丛植被交错带在怒江样带处于海拔1,900-2,100 m处, 在澜沧江河谷位于海拔2,300-2,400 m, 在金沙江河谷位于海拔2,700-2,900 m。所有海拔样带的森林段或灌草丛段相对于同一样带不同植被段之间的物种更替程度为最小, 不仅小于同一流域不同样带相同植被段之间物种更替率的均值, 更小于所有样带相同植被段之间的更替率均值。在三条河流6条海拔样带的12个植被带段之间的物种更替变化中, 空间隔离因素可以解释34.2%, 而植被类型差异仅能解释不到0.5%。本研究结果显示了环境差异对不同植被类型物种丰富度的首要影响, 和各河流之间的空间隔离对植物群落构建和物种构成的主要作用。
Biodiversity, environmental context and structural attributes as drivers of aboveground biomass in shrublands at the middle and lower reaches of the Yellow River basin
DOI:10.1016/j.scitotenv.2021.145198 [本文引用: 1]
Effects of different human disturbances on soil water conversation and fertility of natural secondary shrub
不同人为干扰措施对天然次生灌丛土壤肥力及蓄水能力的影响
Climate-induced spatial mismatch may intensify giant panda habitat loss and fragmentation
DOI:10.1016/j. biocon.2019. 108392 [本文引用: 1]
Effects of target tree tending on community structure and diversity in subtropical natural secondary shrubs
DOI:10.13287/j.1001-9332.201705.037
[本文引用: 1]
The typical natural secondary shrub community was chosen in Lin’an of Zhejiang Pro-vince to discover its possibility of restoration to arbor forest with three kinds of forest management models being taken, <i>i.e</i>., no care as control, closed forest management and target tree tending. Over four years growth, compared with control, closed forest management significantly increased average DBH and height by 130% and 50%, respectively, while 260% and 110% for target tree tending. In target tree tending plots, larger trees had been emerging with 4.5-8.5 cm diameter class and 4.5-8.5 m height class and formed a new storey of 4 m compared with control. The species biodiversity indexes at shrub layer were significantly increased in closed management plots, and did not decrease in target tree tending plots. Closed forest management did not change the tree species composition, following its previous succession direction. However, target tree tending increased the importance value of target species with the high potential succession direction of mixed coniferous-broadleaved forest. The results revealed that the secondary shrub community with target tree tending achieved more desired goals on DBH and height growth of dominant trees and species composition improvement compared with closed management. If the secondary shrub community could be managed when the operational conditions existed, target tree tending model should be selected to accelerate the restoration of shrub toward arbor forest.
目标树抚育对亚热带天然次生灌丛群落结构和多样性的影响
DOI:10.13287/j.1001-9332.201705.037
[本文引用: 1]
在浙江省临安市选择典型天然次生灌丛,分别进行封禁和目标树抚育,探讨灌丛恢复为乔木林的可能性.结果表明: 4年后,与未管护(对照)相比,封禁和目标树抚育后群落平均胸径分别提高1.3和2.6倍,平均高度分别提高0.5和1.1倍;目标树抚育林木出现了对照林分没有的4.5~8.5 cm径阶和4.5~8.5 m树高阶,形成了4 m高的新林层;灌木层物种丰富度和多样性指数没有因抚育而下降;封禁管理维持了群落的树种组成,遵循原有的演替方向;目标树抚育显著改变了群落的树种组成,提高了目的树种的重要值,近期有可能恢复成为针阔混交林群落.与封禁相比,目标树抚育在优势林木胸径和高度生长、树种组成改善等方面更能达到预想的目标.在有条件经营的情况下,可以选择目标树抚育模式对天然次生灌丛进行管理,从而达到加快群落恢复演替形成乔木林的目的.
The relative and combined effects of herbivore assemblage and soil nitrogen on plant diversity
DOI:10.1007/s11427-021-1963-2 URL [本文引用: 2]
Alpha, beta and gamma diversity differ in response to precipitation in the Inner Mongolia grassland
DOI:10.1371/journal.pone.0093518 [本文引用: 1]
Effects of nitrogen addition on litter production and leaf decomposition in Rhododendron simsii shrubland in the mid-subtropical of China
氮添加对中亚热带杜鹃灌丛凋落物生产和叶分解的影响
Environmental filtering and spatial processes shape the beta diversity of liana communities in a valley savanna in southwest China
DOI:10.1111/avsc.12514 URL [本文引用: 2]
Shrub community characteristics and quantitative calculation of theirs biomass in southern China
我国南亚热带灌丛群落特征及生物量的定量计算
Patterns and determinants of species similarity decay of forest communities in the western Qinling Mountains
DOI:10.17520/biods.2016220
[本文引用: 1]
Although much research has focused on biodiversity in mountain regions, elevational patterns of community similarity (i.e. species turnover) and the underlying processes are still rarely discussed. In this study, based on field investigations of 28 forest communities in the western Qinling Mountains, we evaluated the combined effects of habitat divergence and geographical distance on patterns of similarity in species composition using Mantel tests and multiple regressions on distance matrices (MRM). We also compared the rates of similarity decay of different life forms with geographical distance using permutation tests. Results indicated that, for all three life forms, community similarity decreased significantly with elevational, geographical, and habitat distances. Geographical distance and habitat heterogeneity together explained 41.1%, 59.0% and 47.4% of variance in species composition of trees, shrubs and herbs, respectively, with geographical distance uniformly explaining more variance than habitat heterogeneity. Woody plants showed a faster decay in similarity with geographical distance than herbaceous species. These results suggest that, at the scale of our study, dispersal limitations and habitat filtering influence species composition together, and highlighted that dispersal limitation may play a more important role in structuring the forest communities of the western Qinling Mountains.
秦岭西部森林群落相似性递减格局及其影响因素
DOI:10.17520/biods.2016220
[本文引用: 1]
群落间物种组成的相似性递减格局与生境过滤、空间过程密切相关, 探索其成因涉及到群落构建机制等生态学核心问题。本研究以秦岭西部森林群落为研究对象, 运用Mantel检验和基于距离矩阵的多元回归方法, 探究了不同生态因子对群落相似性变异的贡献, 并通过置换检验比较不同生活型之间相似性的空间递减率差异。结果显示: (1) 3种生活型物种组成相似性与地理距离、海拔距离、局地生境异质性差异均呈显著递减趋势; (2)地理距离与生境差异作为主导因子, 分别共同解释乔木、灌木和草本群落相似性变异的41.1%、59.0%和47.4%, 且地理距离的单独解释率均大于生境过滤的单独解释率; (3)不同生活型相似性在空间上的递减率大小关系为: 乔木 > 灌木 > 草本, 表明物种因扩散能力的不同而对空间阻隔效应的响应存在差异。本研究初步确定扩散限制与生境过滤共同主导了秦岭西部森林群落的构建过程, 且扩散限制所起作用更为显著。

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}