



不同环境下植物功能性状变异由种间性状变异和种内性状变异组成(Díaz et al., 2004)。种间性状变异是指不同物种之间的功能性状差异, 主要由物种属性决定。种内性状变异指同一物种内不同个体之间的性状差异, 来源于表型可塑性和遗传多样性。然而, 沿环境梯度的植物功能性状研究通常以物种性状平均值表示, 忽略了种内个体之间的性状差异(Messier et al., 2010)。越来越多证据表明, 种内性状变异不可忽略。Siefert等(2015)对625个群落36个性状的研究表明, 种内性状变异解释了群落内总性状变异的25%和群落间性状变异的32%。Albert等(2010)对13种生活型物种的研究发现, 约30%的性状变异来自种内, 其中叶氮和叶碳含量的种内变异性最高。我国亚热带常绿阔叶林植物比茎密度的种内性状变异解释量高达51.50% (唐青青等, 2016)。种内性状变异除能解释群落性状变异外, 对群落构建和生态系统功能也具有重要影响(Violle et al., 2012; Benavides et al., 2021)。群落构建方面, Jung等(2010)发现种内性状变异有助于更多物种通过生物和非生物筛, 促进物种共存。Siefert (2012)使用零模型也得出, 包含种内性状变异能增强环境过滤的影响, 提供更为全面的群落构建信息。生态系统功能方面, Laforest-Lapointe等(2014)发现种内性状变异是决定植物能否应对气候变化和其他环境因子改变的重要因素; 对青藏高原草地群落的研究也发现, 种内性状变异主导了年际间气候波动引起的草地群落功能多样性变化, 缓冲了气候波动对群落的影响, 对群落稳定性有正向作用(Chen et al., 2019)。忽略种内性状变异, 只考虑种间功能性状, 会低估物种间生态位和功能性状重叠, 可能对基于性状的物种共存、群落功能与群落动态的理解产生偏差(Violle et al., 2012)。尽管各组织尺度的性状变异来源分析已有较多成果, 但受空间范围、物种类型、性状选择等因素影响, 功能性状变异在不同区域的相对大小仍存在争议(Albert et al., 2010; Siefert et al., 2015)。
林窗是森林群落中林冠层树木死亡或人为移除从而形成的林中空隙(Watt, 1947)。作为一种经常发生的小尺度干扰, 林窗是森林动态循环的关键环节(梁晓东和叶万辉, 2001), 林窗形成释放了林内生存空间与可获得性资源, 实现了林中生态资源的重新分配(龙翠玲, 2008; Zhu et al., 2014; 李强等, 2019)。林窗大小是林窗最基本特征之一, 随着林窗面积增大, 光照、温度、水分与土壤等环境因子呈现一定空间变异(He et al., 2012, 2015)。不同大小林窗即不同资源状态, 具有差异化的资源有效性(龙翠玲, 2006), 促使植物最大化利用资源, 提高个体适合度, 最终引起群落内物种生态位分化(Nishimura et al., 2010)。其分化结果主要表现在3个方面: 一是群落内物种组成变化, 尤其是喜光和耐阴树种变化。大林窗因具有更多可利用光资源易于喜光先锋树种生长, 小林窗则更利于耐阴树种建植(Zhu et al., 2014; 罗丝琼等, 2020)。另一方面, 林窗也促进了某些特定树种定居。Yan等(2010)研究发现, 林窗内土壤种子与地上植被的相似性与林窗面积成反比, 林窗内种子植物种类是林内的1.25倍。邻域种源物种丰富度是林窗内物种丰富度的决定因素之一(刘何铭等, 2015)。二是物种生态位宽度及生态位重叠变化。林窗与林下生态位宽度变化具有物种特异性, 如亚热带常绿阔叶林的格氏栲(Castanopsis kawakamii)、光叶山矾(Symplocos lancifolia)和千里香(Murraya paniculata)在林窗中生态位宽度要高于林下, 而红皮糙果茶(Camellia crapnelliana)、木荷(Schima superba)与矩叶鼠刺(Itea oblonga)的生态位宽度林下高于林窗(何中声等, 2012a)。尽管生态位宽度因物种而异, 但林窗有助于降低种间生态位重叠, 缓解激烈的种间竞争, 促进物种共存(龙翠玲, 2006; 何中声等, 2012b)。三是植物功能性状变异, 即植物通过表型可塑性响应环境变化。对寒温带落叶林的研究发现, 人为创造林窗导致物种幼苗的光饱和速率和相对生长速率显著增加(Oguchi et al., 2017)。对紫楠(Phoebe sheareri)的研究也发现, 林窗中紫楠幼苗的叶长、叶面积和叶周长显著低于林下生境(周俊宏等, 2015)。然而目前植物响应林窗变化研究以物种组成和更新生态位研究较多, 林窗特征对植物功能性状及其变异分解的相关研究则较少, 难以深入理解林窗在物种共存与更新中的重要作用。
福建三明格氏栲省级自然保护区分布着世界范围内面积较大、分布较集中的格氏栲天然群落, 有“世界格氏栲林”之称(何中声等, 2012a)。目前, 三明格氏栲种群面临衰退, 林冠层出现严重断层现象, 林窗数量增多(刘金福等, 2006)。前期研究发现, 林窗能有效提高群落物种多样性和系统发育多样性(陈博等, 2021), 降低物种间生态位重叠(何中声等, 2012b), 有利于群落物种共存和天然更新。但植物功能性状如何响应林窗变化, 尤其林窗大小影响植物功能性状变异的研究目前鲜有报道, 这限制了对森林动态循环中植物适应性的深入理解。为探究格氏栲天然林林窗大小对植物功能性状变异的影响, 分析变异来源及其相对贡献, 通过设置不同大小林窗固定样方测量植物功能性状, 以研究: (1)格氏栲天然林林窗环境下, 植物功能性状变异来源的相对贡献如何? (2)植物群落性状是否随林窗大小而改变? (3)不同大小林窗间, 种间和种内性状变异的相对重要性如何?
1 材料和方法
1.1 研究区概况
1.2 数据获取
林窗分为实际林窗和扩展林窗, 实际林窗指林冠层空隙向下投影的土地面积或空间(梁晓东和叶万辉, 2001), 是阳光垂直照射的部分, 而扩展林窗是由组成林冠空隙的边界木所围成的土地面积或空间(梁晓东和叶万辉, 2001), 其面积高于实际林窗, 包括了阳光照射及其向林下过渡的部分。一般而言, 林窗大小由实际林窗计算得到, 而林窗植被调查选择扩展林窗, 格氏栲林也采取相同方法, 即拍摄实际林窗计算林窗大小, 扩展林窗内进行物种调查。2018年3月, 对格氏栲自然保护区内林窗集中分布区域实地踏查后, 随机选择不同大小的林窗共9个(图1)。每个林窗定为一个样方, 根据林窗边界木确定样方范围。在样方中心, 利用尼康D7200搭配鱼眼镜头拍摄冠层林窗图片, 采用半球面影像法计算林窗面积(Hu & Zhu, 2009), 利用Gap Light Analyzer Version 2.0图像处理软件进行冠层半球影像分析, 输入样地经纬度和海拔等信息作为分析参数后, 可得到林冠开放度。林冠开放度指从林地中一点向上仰视, 视野范围内未被林木枝体遮挡的天空球面的比例(朱教君, 2003), 本研究以林冠开放度作为林窗大小的度量(表1)。样方确定后, 对样方内所有胸径大于1 cm植株进行编号挂牌并记录物种名。
图1

图1
格氏栲天然林林窗样方分布。
Fig. 1
Distribution of canopy gap plots in the Castanopsis kawakamii natural forest.
表1 格氏栲天然林林窗基本概况
Table 1
| 林窗编号 Gap number | 林窗面积 Gap area (m2) | 林冠开放度 Canopy openness (%) | 海拔 Altitude (m) | 物种数 Species number | 主要树种 Dominate species |
|---|---|---|---|---|---|
| L1 | 210.56 | 31.96 | 224 | 24 | 格氏栲 Castanopsis kawakamii 桂北木姜子 Litsea subcoriacea 狗骨柴 Diplospora dubia |
| L2 | 200.38 | 23.92 | 211 | 18 | 格氏栲 Castanopsis kawakamii 桂北木姜子 Litsea subcoriacea 木荷 Schima superba |
| L3 | 207.57 | 29.91 | 214 | 22 | 格氏栲 Castanopsis kawakamii 桂北木姜子 Litsea subcoriacea 新木姜子 Neolitsea aurata |
| M1 | 74.22 | 17.34 | 196 | 11 | 格氏栲 Castanopsis kawakamii 黄绒润楠 Machilus grijsii 木荷 Schima superba |
| M2 | 70.59 | 11.56 | 214 | 18 | 格氏栲 Castanopsis kawakamii 桂北木姜子 Litsea subcoriacea 光叶山矾 Symplocos lancifolia |
| M3 | 74.65 | 19.21 | 188 | 13 | 格氏栲 Castanopsis kawakamii 桂北木姜子 Litsea subcoriacea 狗骨柴 Diplospora dubia |
| S1 | 32.11 | 7.45 | 225 | 16 | 桂北木姜子 Litsea subcoriacea 黄绒润楠 Machilus grijsii 茜树 Aidia cochinchinensis |
| S2 | 31.59 | 7.32 | 203 | 10 | 格氏栲 Castanopsis kawakamii 桂北木姜子 Litsea subcoriacea 木荷 Schima superba |
| S3 | 36.78 | 8.32 | 214 | 19 | 格氏栲 Castanopsis kawakamii 桂北木姜子 Litsea subcoriacea 榕叶冬青 Ilex ficoidea |
植物功能性状选择6个常见叶片性状: 比叶面积、叶干物质含量、叶厚、叶绿素含量、叶氮含量和叶磷含量。比叶面积、叶绿素、叶氮和叶磷含量可直接或间接反映植物光合与碳获取能力, 叶干物质含量和叶厚能指示植物叶片的物理抵抗能力(Pérez-Harguindeguy et al., 2013)。每个林窗样方内每个物种随机选择5株个体, 每个个体采集完全展叶并充分接受阳光照射且无病虫害的叶片约30片, 若不足5株个体, 则所有个体均采集叶片。从中选取5片大小均匀的叶片(含叶柄)测定比叶面积、叶干物质含量、叶厚和叶绿素含量。使用便携式叶面积仪(Yaxin-1242, 北京雅欣理仪科技有限公司)测定植物叶面积。采用分析天平测定叶片饱和鲜质量。使用数显游标卡尺避开叶片主脉分别测量叶片前、中、末端厚度, 以平均值作为厚度。采用便携式叶绿素仪(SPAD-502 plus, Konica Minolta, Tokyo, Japan)测定叶绿素相对含量。以上指标测定后, 所有叶片放置105 ℃杀青10 min, 80 ℃烘干至恒质量, 取出测量叶片干质量。比叶面积为叶面积与干质量的比值, 采用靛蓝比色法测定植物叶片全氮含量, 钼锑抗比色法测定叶片全磷含量(Pérez-Harguindeguy et al., 2013)。本研究对格氏栲天然林林窗内共47种植物361株个体采集叶片样品。
林窗内选择海拔作为地形因子, 环境因子选择土壤含水量、土壤温度、土壤pH、水解氮和有效磷含量。海拔由手持GPS (G138BD, UniStrong, 北京)在林窗样方中心处测得。土壤温度监测使用iButton温度记录器(DS1922L-F50, Maxim, California, USA)测定。2018年3月将该温度记录器放置在各样方中心, 设置每2 h记录一次数据, 连续监测一年。计算一年数据的平均值作为每个样地的土壤温度。在林窗东、南、西、北、中5个方位采集土样, 带回实验室测量土壤理化性质。土壤含水量用烘干法测定, 土壤pH用电位法测定, 土壤水解氮含量利用碱解-扩散法测定, 有效磷含量用钼锑抗比色法测定(国家林业局, 1999)。
1.3 数据分析
图2
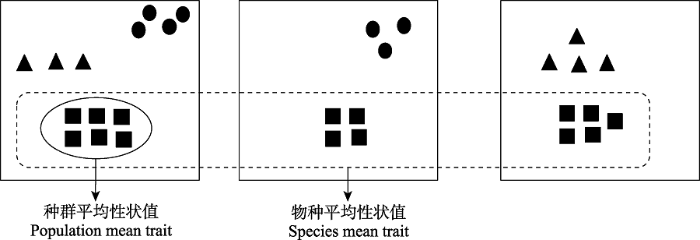
图2
物种平均性状值与种群平均性状值概念示意图(参考Jung et al., 2010)。图中粗实线方框代表一个群落或样方, 某一物种的种群平均性状值是群落内该种群不同个体的性状平均值, 即图中实线椭圆形所示部分。某一物种的物种平均性状值为分布于所有群落的所有个体的性状均值, 即图中虚线矩形所示部分。
Fig. 2
Conceptual illustration of species mean trait and population mean trait (developed based on Jung et al., 2010). The thick solid rectangle represents a community or plot. The population mean trait value of a species is the average of different individual trait values in the plot, as shown by solid ellipse. The species mean trait value is the average of all individual trait values distributed in all plots, as shown by the dotted rectangle.
为了解整个格氏栲林窗群落植物叶功能性状变异来源及其相对贡献, 采用线性混合模型分解3个嵌套尺度(林窗、物种和个体)对性状变异的解释方差(Messier et al., 2010)。格氏栲天然林共设有9个不同大小的林窗样方, 每个样方内包含不同的物种, 每个物种又由多个个体组成, 由此构成林窗-物种-个体的嵌套尺度。林窗尺度代表群落水平由林窗大小差异引起的局部微生境异质性对功能性状的影响, 可以指示非生物因素对性状变异的作用; 物种尺度用于揭示种间性状变异的影响; 个体尺度反映的是种内性状变异。由于功能性状数据不符合正态分布, 故对数转换后进行后续分析。使用R 4.0.3软件“nlme”包基于限制性最大似然估计(REML)的“lme”函数拟合线性混合模型(Pinheiro et al., 2020)。然后通过ape包对林窗、物种和个体3个嵌套尺度的功能性状进行方差分解(Paradis & Schliep, 2019)。方差分解构建的模型形式如下, 以比叶面积为例:
varcomp.SLA < -varcomp (lme (SLA~1, random = ~1|Gap/Species/Individual, data = GSK, na.action = na. omit), 1)。
式中, SLA表示比叶面积, Gap、Species和Individual分别为林窗、物种和个体, GSK为格氏栲林功能性状数据集, 通过“varcomp”函数分离林窗、物种和个体对功能性状变异的相对贡献。
以样方内所有物种为对象, 以物种在样方内的相对多度为权重计算群落加权平均性状值(CWMpopulation)作为该样方的群落功能性状。CWMpopulation计算公式如下:
式中, S为每个林窗样方的物种数, pi为物种i在每个林窗样方中的相对多度, traiti为物种i在每个林窗样方内所有个体的性状均值(即种群平均性状值)。每个样方物种数与优势种情况见表1。采用线性回归拟合群落功能性状CWMpopulation与林冠开放度之间的关系, 分析林窗大小如何影响CWMpopulation。为明确驱动CWMpopulation变化的主要环境因素, 首先对所有因子(海拔、土壤含水量、土壤温度、土壤pH、土壤水解氮和土壤有效磷含量)进行正态性检验, 由于数据不符合正态分布, 对数据进行对数转换。之后, 利用逐步回归结合赤池信息准则(AIC)筛选最优模型确定影响因子(附录I), 并采用方差膨胀因子(VIF)分析变量间的共线性, 剔除VIF > 3的影响因子以保证因子间无明显共线性(附录II)。最后, 采用线性模型分析影响CWMpopulation的关键影响因子。采用R 4.0.3进行正态性检验、逐步回归和线性回归, “vegan”包实现VIF去除共线性(Oksanen et al., 2020)。
为了解沿环境梯度群落平均性状变化的种间和种内相对贡献, 根据Cornwell和Ackerly (2009)的研究, 还计算了基于物种平均性状值的群落加权平均性状CWMspecies。CWMspecies与CWMpopulation的不同之处在于, CWMpopulation同时包含了物种组成变化和种内性状变异, 而CWMspecies沿环境梯度的变化完全由物种组成改变引起。因此, 二者与林冠开放度的回归斜率比值为种间性状变异的相对贡献, 回归斜率相对减少量为种内性状变异贡献。
2 结果
2.1 林窗环境下植物功能性状变异系数
格氏栲天然林林窗环境下, 叶磷含量和叶干物质含量的种间性状变异系数较高(表2), 分别为0.28和0.25, 叶绿素含量和比叶面积较低, 分别为0.04和0.08。平均种内性状变异系数最高的是叶磷含量, 比叶面积最低。其中格氏栲、狗骨柴(Diplospora dubia)、桂北木姜子(Litsea subcoriacea)、虎皮楠(Daphniphyllum oldhami)和黄绒润楠(Machilus grijsii)在不同大小林窗的种内性状变异系数较大。
表2 格氏栲天然林林窗环境下植物种间与种内性状变异系数
Table 2
| 物种 Species | 种内性状变异系数 Coefficient of variation of intraspecific functional traits | |||||
|---|---|---|---|---|---|---|
| SLA | LDMC | Chl | LT | LNC | LPC | |
| 八角枫 Alangium chinense | - | - | - | - | - | - |
| 白花苦灯笼 Tarenna mollissima | 0.22 | 0.37 | 0.03 | 0.26 | 0.34 | 0.23 |
| 赤楠 Syzygium buxifolium | 0.04 | 0.23 | 0.05 | 0.16 | 0.21 | 0.44 |
| 冬青 Ilex chinensis | 0.07 | 0.13 | 0.02 | 0.09 | 0.13 | 0.21 |
| 短尾越桔 Vaccinium carlesii | 0.02 | 0.72 | 0.04 | 0.16 | 0.40 | 0.44 |
| 格氏栲 Castanopsis kawakamii | 0.04 | 0.66 | 0.75 | 0.61 | 0.71 | 0.72 |
| 狗骨柴 Diplospora dubia | 0.03 | 0.71 | 0.85 | 0.76 | 0.73 | 0.84 |
| 光叶山矾 Symplocos lancifolia | 0.04 | 0.23 | 0.03 | 0.08 | 0.10 | 0.37 |
| 广东冬青 Ilex kwangtungensis | 0.01 | 0.58 | 0.04 | 0.10 | 0.24 | 0.01 |
| 桂北木姜子 Litsea subcoriacea | 0.03 | 0.81 | 0.93 | 0.79 | 0.76 | 1.09 |
| 褐毛石楠 Photinia hirsuta | 0.01 | 0.10 | 0.04 | 0.08 | 0.12 | 0.62 |
| 红皮糙果茶 Camellia crapnelliana | 0.08 | 0.14 | 0.01 | 0.04 | 0.04 | 0.39 |
| 虎皮楠 Daphniphyllum oldhami | 0.04 | 0.57 | 0.68 | 0.54 | 0.63 | 0.69 |
| 华南桂 Cinnamomum austrosinense | 0.03 | 0.39 | 0.02 | 0.05 | 0.20 | 0.44 |
| 黄绒润楠 Machilus grijsii | 0.10 | 0.55 | 0.60 | 0.48 | 0.48 | 0.90 |
| 幌伞枫 Heteropanax fragrans | 0.05 | 0.15 | 0.01 | 0.42 | 0.04 | 0.01 |
| 矩叶鼠刺 Itea oblonga | 0.03 | 0.15 | 0.01 | 0.08 | 0.11 | 0.29 |
| 罗浮栲 Castanopsis faberi | - | - | - | - | - | - |
| 罗浮柿 Diospyros morrisiana | 0.16 | 0.36 | 0.00 | 0.27 | 0.23 | 0.59 |
| 毛冬青 Ilex pubescens | 0.04 | 0.30 | 0.03 | 0.17 | 0.26 | 0.41 |
| 米槠 Castanopsis carlesii | 0.03 | 0.11 | 0.03 | 0.07 | 0.03 | 0.16 |
| 闽楠 Phoebe bournei | 0.00 | 0.12 | 0.01 | 0.03 | 0.08 | 0.13 |
| 木荷 Schima superba | 0.06 | 0.12 | 0.02 | 0.13 | 0.12 | 0.54 |
| 木姜叶柯 Lithocarpus litseifolius | 0.01 | 0.20 | 0.00 | 0.03 | 0.54 | 0.31 |
| 茜树 Aidia cochinchinensis | 0.09 | 0.37 | 0.01 | 0.11 | 0.09 | 0.30 |
| 日本杜英 Elaeocarpus japonicus | 0.05 | 0.35 | 0.38 | 0.35 | 0.46 | 0.61 |
| 绒毛山胡椒 Lindera nacusua | - | - | - | - | - | - |
| 榕叶冬青 Ilex ficoidea | 0.00 | 0.49 | 0.01 | 0.13 | 0.02 | 0.14 |
| 山鸡椒 Litsea cubeba | - | - | - | - | - | |
| 山杜英 Elaeocarpus sylvestris | 0.06 | 0.18 | 0.03 | 0.06 | 0.51 | 0.07 |
| 山矾 Symplocos sumuntia | 0.02 | 0.06 | 0.02 | 0.08 | 0.18 | 0.42 |
| 千里香 Murraya paniculata | 0.04 | 0.30 | 0.03 | 0.08 | 0.22 | 0.16 |
| 石斑木 Rhaphiolepis indica | - | - | - | - | - | - |
| 石楠 Photinia serrulata | - | - | - | - | - | - |
| 树参 Dendropanax dentiger | - | - | - | - | - | - |
| 光亮山矾 Symplocos lucida | - | - | - | - | - | - |
| 酸味子 Antidesma japonicum | 0.13 | 0.20 | 0.04 | 0.27 | 0.11 | 0.55 |
| 台湾冬青 Ilex formosana | - | - | - | - | - | - |
| 甜槠 Castanopsis eyrei | - | - | - | - | - | - |
| 细枝柃 Eurya loquaiana | 0.06 | 0.07 | 0.02 | 0.19 | 0.06 | 0.57 |
| 香港新木姜子 Neolitsea cambodiana var. glabra | 0.02 | 0.17 | 0.02 | 0.07 | 0.06 | 0.54 |
| 香叶树 Lindera communis | - | - | - | - | - | - |
| 小叶蚊母树 Distylium buxifolium | - | - | - | - | - | - |
| 新木姜子 Neolitsea aurata | 0.02 | 0.13 | 0.01 | 0.08 | 0.07 | 0.31 |
| 杨桐 Adinandra millettii | - | - | - | - | - | - |
| 野含笑 Michelia skinneriana | - | - | - | - | - | - |
| 油茶 Camellia oleifera | 0.03 | 0.07 | 0.02 | 0.12 | 0.10 | 0.46 |
| 种间性状变异 Coefficient of variation of interspecific functional traits | 0.08 | 0.25 | 0.04 | 0.16 | 0.13 | 0.28 |
| 平均种内性状变异 Mean coefficient of variation of intraspecific functional trait | 0.05 | 0.31 | 0.14 | 0.21 | 0.25 | 0.42 |
Chl、LDMC、LNC、LPC、LT和SLA分别为叶绿素含量、叶干物质含量、叶氮含量、叶磷含量、叶厚和比叶面积。表中“-”表示该物种仅在一个样方分布, 无法计算种内性状变异系数。
Chl, LDMC, LNC, LPC, LT and SLA were leaf chlorophyll content, leaf dry matter content, leaf nitrogen content, leaf phosphorus content, leaf thickness and specific leaf area. The symbol “-” in the table means that the species only distributes in one plot, so that this species can not calculate intraspecific coefficient of variation.
2.2 格氏栲林窗环境下植物功能性状变异来源分析
格氏栲天然林林窗环境下植物功能性状变异来源因所选性状不同而有所差异(图3), 大致可分为3类。第一类, 环境主导。主要为叶磷含量, 其性状变异主要来自于林窗(62%)。第二类, 种间性状变异主导。包括比叶面积、叶干物质含量、叶厚和叶绿素含量, 种间性状变异对这4个性状的解释量在45%-65%。第三类, 种内性状变异主导。种内性状变异对叶氮含量的解释量高达72%, 是叶氮含量变异最主要来源。林窗、种间性状变异和种内性状变异平均解释量分别为23%、41%和30%。
图3
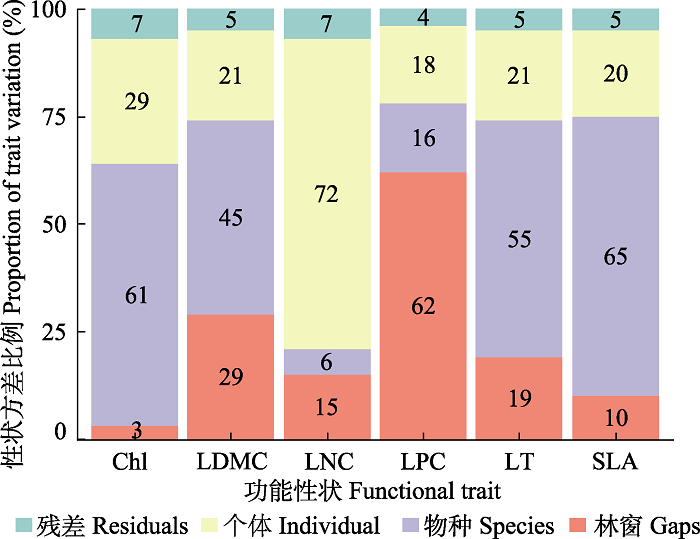
图3
格氏栲天然林林窗环境下植物功能性状变异来源分析。变异主要来源于嵌套的3个部分, 分别为林窗、物种和个体。图中数值为对应的方差解释量, 解释量越高, 表明该因素对性状变异的影响越大。Chl、LDMC、LNC、LPC、LT和SLA分别为叶绿素含量、叶干物质含量、叶氮含量、叶磷含量、叶厚和比叶面积。
Fig. 3
The variation partitioning of plant functional traits under canopy gaps in the Castanopsis kawakamii natural forest. The variation mainly comes from three nested parts, namely gap, species and individual. The value in the figure is the explanatory variance. The higher the variance explained, the greater the influence of the factor on trait variation. Chl, LDMC, LNC, LPC, LT and SLA were leaf chlorophyll content, leaf dry matter content, leaf nitrogen content, leaf phosphorus content, leaf thickness and specific leaf area.
2.3 林窗大小异质生境中群落功能性状变化
林冠开放度与群落功能性状的回归分析如图4所示。6个群落功能性状中, 仅群落叶磷含量与林冠开放度具有显著线性关系(R2 = 0.67, p < 0.05), 表现为随林冠开放度增加, 群落叶磷含量越高。
图4
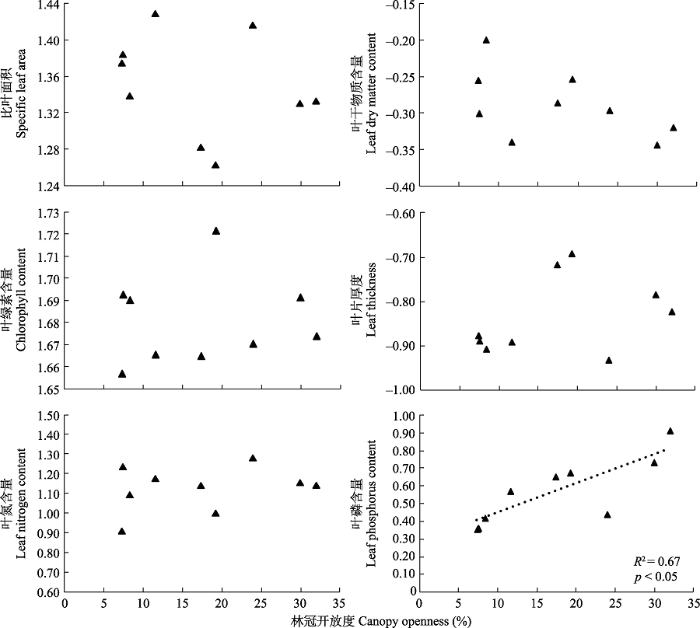
图4
不同大小林窗异质生境中群落加权平均性状变化。图中功能性状为对数转换后数值。
Fig. 4
Changes in community weight mean of leaf functional traits among different size of canopy gaps. Functional traits in the figure were log-transformed values.
2.4 群落功能性状变化影响因子分析
线性模型分析发现, 非生物因子对除叶绿素含量外的所有性状均具有显著影响(图5)。比叶面积受土壤温度的影响显著; 干物质含量受水解氮和有效磷含量影响显著; 土壤温度和海拔显著影响叶片厚度; 群落叶氮含量受土壤温度、土壤水解氮含量和土壤有效磷含量的影响显著; 对于群落叶磷含量, 土壤温度和土壤水解氮含量对其具有显著正效应, 土壤有效磷含量对其具有显著负效应。
图5
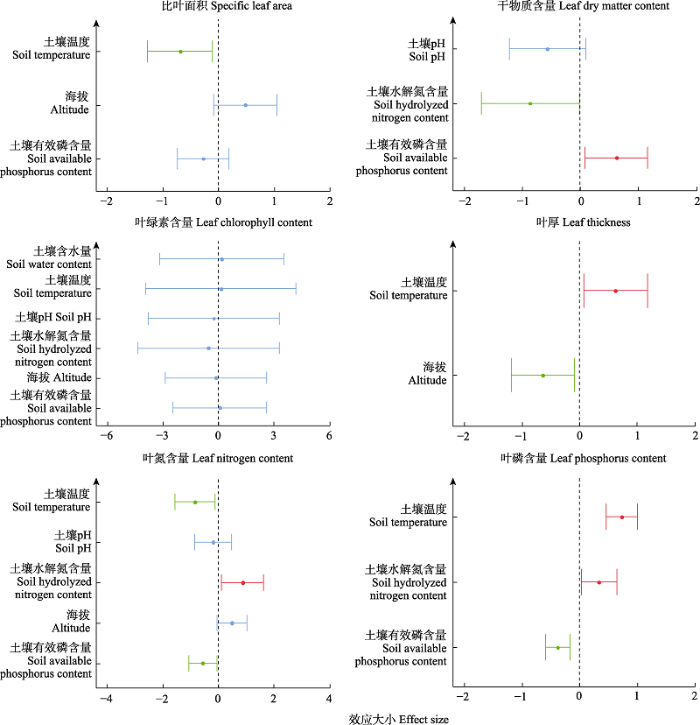
图5
非生物因子对群落加权平均性状的线性模型分析。图中蓝色线条表示该因子对群落性状无显著效应, 红色线条表示具有显著正效应, 绿色线条为显著负效应。显著水平为0.05。
Fig. 5
Linear regression analysis of abiotic factors for community weight mean of leaf functional traits. The blue line indicates no significant effect of the factor on CWM, the red line indicates a significant positive effect, and the green line indicates a significant negative effect. The significance level is 0.05.
2.5 不同林窗大小群落叶磷性状的变异来源分解
由于随林冠开放度增大, 仅群落叶磷含量(LPC)具有显著变化, 因此分析了基于种群平均性状值的CWMpopulation_LPC和物种平均性状值的CWMspecies_LPC与林冠开放度的线性回归关系(表3)。CWMpopulation_LPC拟合的回归斜率为0.016, CWMspecies_LPC拟合的回归斜率为0.003。种间性状变异对CWMpopulation_LPC的相对重要性为17.79%, 而种内性状变异的相对贡献为82.21%。
表3 群落叶磷含量的种间和种内来源分析
Table 3
| 变异来源 Source of variability | 响应变量 Predictive variable | 预测变量 Response variable | 斜率 Slope | R2 | p | 相对重要性 Relative importance (%) |
|---|---|---|---|---|---|---|
| 种间和种内 Inter- and intraspecific | CO | CWMpopulation_LPC | 0.016 | 0.67 | 0.007 | 100.00 |
| 种间 Interspecific | CO | CWMspecies_LPC | 0.003 | 0.62 | 0.012 | 17.79 |
| 种内 Intraspecific | - | - | - | - | - | 82.21 |
CWMpopulation_LPC和CWMspecies_LPC分别是基于种群平均性状值和物种平均性状值计算的群落叶磷含量。CO, 林冠开放度。
CWMpopulation_LPC and CWMspecies_LPC are community mean values of leaf phosphorus content calculated by population mean trait and species mean trait, respectively. CO, canopy openness.
3 讨论
3.1 格氏栲天然林林窗环境下植物功能性状的变异来源
格氏栲林林窗环境下植物功能性状变异来源依性状不同而有所差异。比叶面积、叶干物质含量、叶厚和叶绿素含量由种间性状变异主导, 种内性状变异解释了72%的叶氮含量变异, 而叶磷含量受林窗影响较大(图3)。叶片化学性状比形态性状具有更高的种内性状变异(Siefert et al., 2015), 加拿大老龄林叶氮含量也具有较大种内性状变异(Auger & Shipley, 2013), 本研究的叶氮含量变异与之相符, 但叶磷含量的种内性状变异低于其他性状。其原因可能在于林窗引起的生境异质性作为叶磷性状变异最主要来源, 降低了种内性状变异的相对贡献。回归分析也表明群落叶磷含量沿林窗大小具有显著变化(图4)。环境条件对叶磷含量的影响在星斗山(吴漫玲等, 2019)和延河流域(张莉等, 2013)均有报道, 尤其星斗山, 海拔显著影响了叶磷含量, 而对叶氮含量影响不显著。此外, 林窗样地较少, 样本量偏低, 也可能造成叶磷和叶氮含量变异来源的差异。总的来说, 种内、种间和环境对性状变异的相对大小具有性状特异性, 叶磷含量主要由林窗变化引起, 叶氮含量种内性状变异占优势, 经济谱性状变异更多来源于种间。
3.2 林窗环境下群落叶磷含量影响因素分析
林冠开放度变化引起光环境改善, 促进了格氏栲天然林植物群落叶磷含量的显著提高(图4)。磷是植物光合作用必不可少的元素, 是组成ATP及多种光合辅酶的重要成分, 直接参与光合磷酸化与碳同化(程滨等, 2010), 一定程度上增加叶磷含量可提高植物光合效率。根据生长速率假说(Elser et al., 2000), 有机体内磷含量直接影响生长率变化, 当有机体内磷含量丰富时, 能满足细胞合成核糖体和蛋白质的需求, 有机体具有较高生长率。因而, 随着林冠开放度增大, 林窗内植物通过提高叶磷含量, 增强植物光合能力, 以快速生长的策略适应林窗生境。植物叶磷含量主要来源于土壤磷吸收, 磷作为中国南方土壤主要限制元素, 格氏栲天然林土壤有效磷含量仅12.05 mg·kg-1 (附录III), 显著低于全国平均水平(27.9 mg·kg-1)(张俊伶等, 2020)。土壤磷限制充当一个环境筛, 导致群落叶磷含量受环境制约较大。线性模型发现, 格氏栲林窗中土壤磷含量对群落叶磷含量具有显著负效应。土壤磷主要来源于凋落物分解, 冯雪萍等(2017)对格氏栲林凋落物与土壤化学计量的研究表明, 大林窗和中林窗凋落物磷含量与土壤磷含量呈显著负相关关系, 说明土壤养分消耗高于凋落物分解回归, 加之南方土壤淋溶作用强烈, 磷限制严重, 植物对磷有较大需求, 促成了土壤磷含量对群落叶磷含量的显著负效应, 符合营养奢侈假说(Chapin III, 1980), 即养分冲刷较大生境中植物养分吸收超过即时生长所需, 有助于土壤养分储备耗尽时支持自身生长。除土壤磷含量影响外, 土壤温度对植物叶磷含量也有显著正效应, 生物地球化学假说为植物叶磷含量变化提供了重要解释(Reich & Oleksyn, 2004)。该假说指出, 温度影响土壤微生物活性和有机质分解与矿化, 进而改变植物对土壤磷的可用性, 温度升高能促进植物叶磷含量提升(Reich & Oleksyn, 2004)。格氏栲天然林植物叶磷含量对土壤温度的响应一定程度上支持生物地球化学假说, 但土壤温度对叶氮含量具有显著负效应, 这与Reich和Oleksyn (2004)基于全球尺度的研究结果一致。群落性状如比叶面积、叶干物质含量、叶厚、叶绿素和叶氮含量随着林冠开放度增大无显著变化, 这可能受本研究样本量太小影响(仅9个群落性状值), 从而降低了回归方程的稳定性, 今后研究需扩大林窗样地数量, 保证足够的样本量。总体而言, 群落叶磷含量是响应格氏栲天然林林窗大小最敏感的性状, 群落叶磷含量可能受到林冠开放度、土壤温度、水解氮和土壤磷含量的共同影响。林窗大小引起的微环境异质性是功能性状适应性变化的潜在原因。
3.3 群落叶磷含量种间和种内来源的相对重要性
不同大小林窗间, 群落叶磷含量变化的82.21%来源于种内性状变异, 17.79%来源于种间性状变异, 表明沿环境梯度的种内性状变异是群落叶磷含量变化的主体, 群落间物种组成差异对群落叶磷含量的影响相对较小。由表2可知, 种内性状变异系数在各物种之间变化很大, 群落内优势种格氏栲、桂北木姜子、狗骨柴、黄绒润楠等种内性状变异系数均较高, 表明优势种在林窗环境下具有较大的表型可塑性。对林窗群落物种β多样性研究发现(未发表数据), 优势种能在不同群落中稳定存在, 其对β多样性的周转和嵌套贡献都很小, 导致种间性状变异的相对贡献较低。随着林冠开放度增加, 群落叶磷含量变化主要来源于种内性状变异, 优势种在其中扮演着重要角色。优势种的广泛分布降低了物种组成变化, 而其较大的表型可塑性增大了种内性状变异。
然而对于整个格氏栲林窗群落来说, 种间性状变异是该群落植物功能性状变异最主要来源, 种间性状变异平均解释量41%, 种内性状变异平均解释量30% (图3)。而对于不同林窗大小的植物群落来讲, 种内性状变异是植物适应环境变化的最主要方式。二者产生差异的原因可能是嵌套方差分解纵向分离了不同生态尺度对局域群落植物功能性状的相对影响, 性状变异不只来源于个体和物种, 按分析要求还可上升至样方、站点、区域等, 加之群落叶磷含量受林窗大小影响显著, 促使林窗对性状变异的解释量有所提高, 而种间和种内性状变异解释量降低。基于群落平均性状的回归分析分离的是沿环境梯度的物种组成和种内性状变异的相对影响, 变异来源只区分物种组成变化和种内性状变异, 且与环境梯度密切相关。沿林窗大小梯度, 群落间物种组成变化较小, 种内个体间性状变异大, 提高了种内性状变异的相对贡献。此外, 嵌套方差分解分离的种间水平差异更多是物种本身性状差异, 由物种进化历史决定, 而回归分析的种间性状变异来源于沿环境梯度的物种组成变化, 通过群落间物种周转来实现, 由此造成不同分析方法结果的差异性。
格氏栲天然林林窗环境下, 随着林冠开放度增加, 群落叶磷含量发生显著变化, 种内性状变异是群落性状变化的最主要来源, 不同群落间物种组成虽有所改变, 但相对重要性较低。当关注的对象上升至整个格氏栲林窗植物群落时, 植物功能性状变异由物种主导, 林窗和个体变异也会引起功能性状变化。总的来说, 格氏栲林窗环境下植物功能性状变异主要来源于种间, 但随着林窗增大, 植物主要通过种内性状变异响应环境变化。
4 总结
格氏栲天然林林窗环境下植物功能性状仍以种间性状变异为主,植物叶磷含量是响应林窗大小变化最敏感的性状, 受林冠开放度、土壤温度、水解氮和有效磷含量的共同影响, 大林窗群落叶磷含量更高。群落叶磷含量随林冠开放度的变化主要来源于种内性状变异, 植物主要通过表型可塑性响应环境变化, 优势种在其中扮演着重要角色。
致谢
感谢福建农林大学林学院的谷新光、张子扬、卢尧舜、元云峰和林敏杰同学在野外样地设置、样品采集和实验测定工作中给予的帮助。
附录I 基于逐步回归的格氏栲天然林非生物因子模型筛选
Supplement I Model selection based on stepwise regression for abiotic factors in Castanopsis kawakamii natural forest
附录II 格氏栲天然林非生物因子的方差膨胀因子分析结果
Supplement II Results of variance inflation factor for abiotic factors in Castanopsis kawakamii natural forest
附录III 格氏栲天然林不同林窗大小地形与土壤因子
Supplement III Topography and soil factors of different size of canopy gaps in Castanopsis kawakamii natural forest
参考文献
A multi-trait approach reveals the structure and the relative importance of intra-vs. interspecific variability in plant traits
DOI:10.1111/j.1365-2435.2010.01727.x URL [本文引用: 2]
Inter-specific and intra-specific trait variation along short environmental gradients in an old-growth temperate forest
DOI:10.1111/j.1654-1103.2012.01473.x URL [本文引用: 1]
The GenTree Leaf Collection: inter- and intraspecific leaf variation in seven forest tree species in Europe
DOI:10.1111/geb.13239 URL [本文引用: 1]
The mineral nutrition of wild plants
DOI:10.1146/annurev.es.11.110180.001313 URL [本文引用: 1]
Taxonomic and phylogenetic diversity of plants in a Castanopsis kawakamii natural forest
DOI:10.17520/biods.2020399
[本文引用: 1]

<p id="p00005"><strong>Aims:</strong> Environmental heterogeneity of forest gaps leads to variation in taxonomic and phylogenetic diversity of trees in these areas. Studying tree diversity in different sizes of forest gap communities can help to reveal the mechanisms that drive the formation and maintenance of biodiversity. This study took <i>Castanopsis kawakamii</i>gaps as the research object, and aimed to reveal the relationship between the taxonomic and phylogenetic diversity of plants and its environmental influence factors. </p> <p id="p00010"><strong>Methods:</strong> We examined different sizes of forest gaps in a <i>Castanopsis kawakamii</i> natural forest as to study the taxonomic and phylogenetic diversity of plants, and used a generalized linear model (GLM) to explore the environmental factors driving the community assembly. </p> <p id="p00015"><strong>Results:</strong> We found that the plant species and plant abundance in large gaps (> 200 m<sup>2</sup>) were higher than those of medium gaps ([50 m<sup>2</sup>, 100 m<sup>2</sup>)), small gaps ([30 m<sup>2</sup>, 50 m<sup>2</sup>)) and non-gaps (100 m<sup>2</sup>). The phylogenetic community structure of the large gaps tends to diverge, while that of the medium gaps, small gaps and non-gaps were affected by the combined effect of habitat filtering and competitive exclusion. The phylogenetic community diversity index (PD) was significantly positively correlated with species richness (SR), Margalef index and Shannon-Wiener index, which is related to the higher species composition of sparse species than dense species in forest gaps. Overall, forest gap size had a significantly positive effect on species diversity, and the soil total nitrogen content had a significantly positive effect on community phylogenetic diversity and phylogenetic structure. </p> <p id="p00020"><strong>Conclusion:</strong> The formation of forest gaps increase the taxonomic and phylogenetic diversity of trees in natural forests, with gap size and soil total nitrogen jointly driving tree diversity in these natural forest gaps. </p>
格氏栲天然林林窗植物物种多样性与系统发育多样性
DOI:10.17520/biods.2020399
[本文引用: 1]
林窗环境异质性导致群落物种多样性与系统发育多样性(phylogenetic diversity, PD)存在差异, 研究不同大小的林窗中群落的物种多样性与系统发育多样性有助于揭示林下生物多样性的形成及维持机制。本文以格氏栲(Castanopsis kawakamii)天然林为研究对象, 通过Pearson相关性分析与广义线性模型探讨了林窗内物种多样性与系统发育多样性间的相互关系及其环境影响因素。结果表明: (1)大林窗(面积 > 200 m<sup>2</sup>)植物种类及多度均高于中林窗(50 m<sup>2</sup> ≤ 面积 < 100 m<sup>2</sup>)、小林窗(30 m<sup>2</sup> ≤ 面积 < 50 m<sup>2</sup>)和非林窗(面积 = 100 m<sup>2</sup>)。大林窗群落系统发育结构趋于发散, 中、小林窗和非林窗群落系统发育结构受到生境过滤和竞争排斥综合作用。(2)群落系统发育多样性指数与物种丰富度(species richness, SR)、Margalef丰富度指数和Shannon-Wiener指数均呈显著正相关, 这与林窗内稀有种种类组成多于优势种有关。(3)林窗面积对物种多样性存在显著正效应; 土壤全氮含量对系统发育多样性和系统发育结构存在显著正效应。林窗形成提高了格氏栲天然林群落物种多样性和系统发育多样性, 林窗面积与土壤全氮共同驱动了格氏栲天然林林窗物种多样性和系统发育多样性的变化。
The research advances and prospect of ecological stoichiometry
生态化学计量学研究进展
Temporal intraspecific trait variability drives responses of functional diversity to interannual aridity variation in grasslands
DOI:10.1002/ece3.5156 URL [本文引用: 1]
Community assembly and shifts in plant trait distributions across an environmental gradient in coastal California
DOI:10.1890/07-1134.1 URL [本文引用: 1]
The plant traits that drive ecosystems: evidence from three continents
DOI:10.1111/j.1654-1103.2004.tb02266.x URL [本文引用: 1]
Biological stoichiometry from genes to ecosystems
DOI:10.1111/j.1461-0248.2000.00185.x URL [本文引用: 1]
Ecological stoichiometric characteristics of litter-soil in gap of Castanopsis kawakamii natural forest in Sanming of Fujian
福建三明格氏栲天然林林窗凋落物-土壤的生态化学计量特征
Effects of forest gaps on some microclimate variables in Castanopsis kawakamii natural forest
DOI:10.1007/s11629-012-2304-y URL [本文引用: 1]
The influence of forest gap on species competition of Castanopsis kawakamii natural forest updated layer
林窗对格氏栲天然林更新层物种竞争的影响
Effects of forest gaps on species niche in regeneration layers of Castanopsis kawakamii natural forest
林窗对格氏栲天然林更新层物种生态位的影响
Effects of forest gaps on soil properties in Castanopsis kawakamii nature forest
DOI:10.1371/journal.pone.0141203 URL [本文引用: 1]
Determination of the tridimensional shape of canopy gaps using two hemispherical photographs
DOI:10.1016/j.agrformet.2008.11.008 URL [本文引用: 1]
Intraspecific variability and trait-based community assembly
DOI:10.1111/j.1365-2745.2010.01687.x URL [本文引用: 5]
Intraspecific variability in functional traits matters: case study of Scots pine
DOI:10.1007/s00442-014-2967-x
PMID:24850418
[本文引用: 1]
Although intraspecific trait variability is an important component of species ecological plasticity and niche breadth, its implications for community and functional ecology have not been thoroughly explored. We characterized the intraspecific functional trait variability of Scots pine (Pinus sylvestris) in Catalonia (NE Spain) in order to (1) compare it to the interspecific trait variability of trees in the same region, (2) explore the relationships among functional traits and the relationships between them and stand and climatic variables, and (3) study the role of functional trait variability as a determinant of radial growth. We considered five traits: wood density (WD), maximum tree height (H max), leaf nitrogen content (Nmass), specific leaf area (SLA), and leaf biomass-to-sapwood area ratio (B L:A S). A unique dataset was obtained from the Ecological and Forest Inventory of Catalonia (IEFC), including data from 406 plots. Intraspecific trait variation was substantial for all traits, with coefficients of variation ranging between 8% for WD and 24% for B L:A S. In some cases, correlations among functional traits differed from those reported across species (e.g., H max and WD were positively related, whereas SLA and Nmass were uncorrelated). Overall, our model accounted for 47% of the spatial variability in Scots pine radial growth. Our study emphasizes the hierarchy of factors that determine intraspecific variations in functional traits in Scots pine and their strong association with spatial variability in radial growth. We claim that intraspecific trait variation is an important determinant of responses of plants to changes in climate and other environmental factors, and should be included in predictive models of vegetation dynamics.
Correlation between spatial distribution of forest canopy gaps and plant diversity indices in Xishuangbanna tropical forests
DOI:10.17520/biods.2018258 URL [本文引用: 1]
西双版纳热带雨林林窗空间分布格局及其特征指数与林窗下植物多样性的相关性
DOI:10.17520/biods.2018258
[本文引用: 1]
林窗作为森林群落中一种重要的干扰方式, 对林下物种构成有着重要的影响。开展林窗空间格局及其特征指数与林下植物多样性关系研究对于探讨林窗对林下生物多样性的影响有重要意义, 有助于进一步了解群落动态, 在物种多样性保护方面也具有指导作用。本研究在西双版纳热带雨林地区随机选取3块大小为1 ha的热带雨林为研究样地, 采用轻小型六旋翼无人机搭载Sony ILCE-A7r可见光传感器, 分别获取各个样地的高清数字影像, 结合数字表面高程模型以及各个样地的地形数据用以确定各样区的林窗分布格局, 并进一步提取出各林窗的景观格局指数。结合地面样方基础调查数据, 对各样地各林窗下植物多样性情况进行统计, 旨在分析热带雨林林窗空间分布格局以及林窗下植物多样性对各林窗空间格局特征的响应情况。研究表明, 西双版纳州热带雨林林窗呈大而分散的空间分布, 林窗空间格局特征指数如林窗形状复杂性指数、林窗面积都与林下植物多样性呈显著正相关关系。在面积小的林窗下, 较之林窗形状复杂性因子, 林窗面积大小对林下植物多样性影响更显著; 在面积达到一定程度后, 相对于面积因子, 林窗形状复杂性指数对林下植物多样性影响更显著, 各样地林窗皆趋于向各自所处样地顶极群落发展。
Advances in study on forest gaps
林窗研究进展
Influences on gap species richness in a subtropical evergreen broadleaved forest
DOI:10.17520/biods.2014137 URL [本文引用: 1]
亚热带常绿阔叶林林窗物种丰富度的影响因素
DOI:10.17520/biods.2014137
[本文引用: 1]
林窗是森林群落物种多样性维持中十分重要的结构, 但其本身物种多样性的维持机制尚不清楚, 可能与相邻群落的物种丰富度、林窗面积以及土壤和地形因子等有关。本文选取浙江天童20 ha森林动态监测样地内的84个冠林窗作为研究对象, 分析了林窗内部以及不同距离邻域内的物种组成, 选取包含大部分林窗内物种(≥75%), 并且对林窗内物种丰富度影响最大的邻域作为相关邻域(林窗周围成熟植株通过种子传播影响林窗物种丰富度, 把距离冠林窗边缘5 m范围内的区域作为种源效应邻域; 由于距离林窗较近, 物种组成与林窗形成时仍存活个体的物种组成相似, 故把距离冠林窗边缘4 m范围内的区域作为距离效应邻域), 然后计算种源效应邻域内成熟植株的物种丰富度、距离效应邻域内所有植株的物种丰富度、冠林窗的面积、地形因子(海拔、坡度、坡向、凸度)和土壤因子(pH值、全氮、全碳、全磷), 最后对11个潜在的影响因素进行随机组合, 构建广义线性回归模型, 筛选最优模型(AIC<sub>c</sub>最小)进行分析。结果显示: 种源效应邻域内成熟植株的物种丰富度和冠林窗面积显著影响林窗内物种丰富度, 其他因素并未对冠林窗内物种丰富度产生显著影响。上述结果说明, 冠林窗面积的大小以及外部种源的丰富程度是决定冠林窗内物种丰富度的主要因素, 其他因素可能只产生间接影响。
Characteristics of gaps renewal in Castanopsis kawakamii forests
格氏栲林林窗更新特征的研究
Plant functional traits—Concepts, applications and future directions
植物功能性状研究进展
Study on the regeneration niche of major tree species in gaps in the karst forest in Maolan Nature Reserve
茂兰喀斯特森林林隙更新生态位的研究
Effects of gaps size on regeneration of karst forest in Maolan Natural Reserve of Guizhou Province
茂兰喀斯特森林林隙大小对树种更新的影响
Regeneration composition of woody plant species under forest gap in Maolan karst evergreen deciduous broad-leaved mixed forest
茂兰喀斯特常绿落叶阔叶混交林林窗下木本植物更新组成
How do traits vary across ecological scales? A case for trait-based ecology
DOI:10.1111/j.1461-0248.2010.01476.x
PMID:20482582
[本文引用: 2]
Despite the increasing importance of functional traits for the study of plant ecology, we do not know how variation in a given trait changes across ecological scales, which prevents us from assessing potential scale-dependent aspects of trait variation. To address this deficiency, we partitioned the variance in two key functional traits (leaf mass area and leaf dry matter content) across six nested ecological scales (site, plot, species, tree, strata and leaf) in lowland tropical rainforests. In both traits, the plot level shows virtually no variance despite high species turnover among plots and the size of within-species variation (leaf + strata + tree) is comparable with that of species level variation. The lack of variance at the plot level brings substantial support to the idea that trait-based environmental filtering plays a central role in plant community assembly. These results and the finding that the amount of within-species variation is comparable with interspecific variation support a shift of focus from species-based to trait-based ecology.
Architecture and growth of an annual plant Chenopodium album in different light climates
DOI:10.1007/s11284-009-0666-6 URL [本文引用: 1]
The effect of interspecific variation in photosynthetic plasticity on 4-year growth rate and 8-year survival of understorey tree seedlings in response to gap formations in a cool-temperate deciduous forest
DOI:10.1093/treephys/tpx042 URL [本文引用: 1]
vegan: Community ecology package
ape 5.0: An environment for modern phylogenetics and evolutionary analyses in R
DOI:10.1093/bioinformatics/bty633 URL [本文引用: 1]
New handbook for standardised measurement of plant functional traits worldwide
DOI:10.1071/BT12225 URL [本文引用: 2]
nlme: Linear and nonlinear mixed effects models
Global patterns of plant leaf N and P in relation to temperature and latitude
DOI:10.1073/pnas.0403588101 URL [本文引用: 3]
Reinforcing loose foundation stones in trait-based plant ecology
DOI:10.1007/s00442-016-3549-x
PMID:26796410
[本文引用: 1]
The promise of "trait-based" plant ecology is one of generalized prediction across organizational and spatial scales, independent of taxonomy. This promise is a major reason for the increased popularity of this approach. Here, we argue that some important foundational assumptions of trait-based ecology have not received sufficient empirical evaluation. We identify three such assumptions and, where possible, suggest methods of improvement: (i) traits are functional to the degree that they determine individual fitness, (ii) intraspecific variation in functional traits can be largely ignored, and (iii) functional traits show general predictive relationships to measurable environmental gradients.
Incorporating intraspecific variation in tests of trait-based community assembly
DOI:10.1007/s00442-012-2351-7
PMID:22580640
[本文引用: 1]
Environmental filtering and niche differentiation are processes proposed to drive community assembly, generating nonrandom patterns in community trait distributions. Despite the substantial intraspecific trait variation present in plant communities, most previous studies of trait-based community assembly have used species mean trait values and therefore not accounted for intraspecific variation. Using a null model approach, I tested for environmental filtering and niche differentiation acting on three key functional traits--vegetative height, specific leaf area (SLA), and leaf dry matter content (LDMC)-in old-field plant communities. I also examined how accounting for intraspecific variation at the among-plot and individual levels affected the detection of nonrandom assembly patterns. Tests using fixed species mean trait values provided evidence of environmental filtering acting on height and SLA and niche differentiation acting on SLA. Including plot-level intraspecific variation increased the strength of these patterns, indicating an important role of intraspecific variation in community assembly. Tests using individual trait data indicated strong environmental filtering acting on all traits, but provided no evidence of niche differentiation, although these signals may have been obscured by the effects of dispersal limitation and spatial aggregation of conspecific individuals. There was also strong evidence of nonrandom assembly of individuals within single species, with the strength of environmental filtering varying among species. This study demonstrates that, while analyses using fixed species mean trait values can provide insights into community assembly processes, accounting for intraspecific variation provides a more complete view of communities and the processes driving their assembly.
A global meta- analysis of the relative extent of intraspecific trait variation in plant communities
DOI:10.1111/ele.12508
PMID:26415616
[本文引用: 3]
Recent studies have shown that accounting for intraspecific trait variation (ITV) may better address major questions in community ecology. However, a general picture of the relative extent of ITV compared to interspecific trait variation in plant communities is still missing. Here, we conducted a meta-analysis of the relative extent of ITV within and among plant communities worldwide, using a data set encompassing 629 communities (plots) and 36 functional traits. Overall, ITV accounted for 25% of the total trait variation within communities and 32% of the total trait variation among communities on average. The relative extent of ITV tended to be greater for whole-plant (e.g. plant height) vs. organ-level traits and for leaf chemical (e.g. leaf N and P concentration) vs. leaf morphological (e.g. leaf area and thickness) traits. The relative amount of ITV decreased with increasing species richness and spatial extent, but did not vary with plant growth form or climate. These results highlight global patterns in the relative importance of ITV in plant communities, providing practical guidelines for when researchers should include ITV in trait-based community and ecosystem studies. © 2015 John Wiley & Sons Ltd/CNRS.
Interspecific and intraspecific variation in functional traits of subtropical evergreen and deciduous broad-leaved mixed forests
DOI:10.17520/biods.2015200 URL [本文引用: 2]
亚热带常绿落叶阔叶混交林植物功能性状的种间和种内变异
DOI:10.17520/biods.2015200
[本文引用: 2]
不同物种间的功能性状差异是自然生态系统中物种共存的基础, 而物种内个体间的性状变异对物种的共存和分布同样具有重要作用。本文以湖北星斗山自然保护区亚热带常绿落叶阔叶混交林内28种主要树种(通过物种多度排序获得, 其中常绿和落叶树种各14种)为研究对象, 探讨不同叶习性树种的4种功能性状(比叶面积、叶干物质含量、叶面积和比茎密度)在种间和种内的差异程度。结果表明: (1)常绿和落叶树种在4种功能性状上均存在显著差异, 常绿树种的比叶面积和叶面积显著低于落叶树种, 但叶干物质含量和比茎密度则显著高于落叶树种; (2)比叶面积的变化主要来源于叶习性(57.49%), 叶面积变化主要来源于种间(66.80%)和种内变异(27.52%), 叶干物质含量的变化主要来源于种间(38.12%)和种内(33.88%)变异, 但比茎密度的变化主要来源于种内变异(51.50%), 其次为种间变异(32.52%); (3)常绿和落叶树种种间水平的性状相关性可能掩盖各功能性状之间的相关性。种内变异能够显著影响群落间的植物功能性状差异, 但不同功能性状的种内变异程度存在差异。
Let the concept of trait be functional!
DOI:10.1111/j.0030-1299.2007.15559.x URL [本文引用: 1]
The return of the variance: intraspecific variability in community ecology
DOI:10.1016/j.tree.2011.11.014 URL [本文引用: 2]
Pattern and process in the plant community
DOI:10.2307/2256497 URL [本文引用: 1]
A structured and dynamic framework to advance traits-based theory and prediction in ecology
DOI:10.1111/j.1461-0248.2010.01444.x URL [本文引用: 1]
Analysis of leaf functional traits and functional diversity of woody plants in evergreen and deciduous broad-leaved mixed forest of Xingdoushan
星斗山常绿落叶阔叶混交林木本植物叶功能性状及其多样性特征分析
Spatial distribution pattern of soil seed bank in canopy gaps of various sizes in temperate secondary forests, Northeast China
DOI:10.1007/s11104-009-0172-1 URL [本文引用: 1]
Soil health and agriculture green development: opportunities and challenges
土壤健康与农业绿色发展: 机遇与对策
Source of variation of plant functional traits in the Yanhe River watershed: the influence of environment and phylogenetic background
DOI:10.5846/stxb201212131800 URL [本文引用: 1]
延河流域植物功能性状变异来源分析
Responses of growth, leaf traits and biomass allocation of Phoebe sheareri seedlings to different habitats in forest gap
紫楠幼苗生长、叶性状和生物量分配对林窗不同生境的响应
Effects of gaps on regeneration of woody plants: a meta-analysis
DOI:10.1007/s11676-014-0489-3 URL [本文引用: 2]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}