



草地是地球上分布范围最广的陆地生态系统类型, 占陆地生态系统面积的40%, 不仅是畜牧业生产的重要基地, 也是防止土地风蚀沙化、涵养水源的重要生态屏障(王明君等, 2010)。植物生产力和物种多样性是评价草地生态系统功能和结构的重要指标(王晶等, 2016)。生产力是草地最重要的生态系统功能, 很大程度上影响着其他生态系统功能(如凋落物分解、光截留率、土壤有机质含量), 其变化与群落物种组成和多样性有着密切联系(方精云等, 2018)。然而, 近几十年来由于化石燃料的燃烧和化学肥料的使用导致全球范围内的养分富集越来越严重(Galloway et al., 2008; Clark et al., 2017)。一方面养分富集可以缓解植物所受到的养分限制, 导致草地群落生产力的提高(Zhao et al., 2019; 张馨文等, 2021)。另一方面养分富集会打破不同物种间原有的竞争平衡, 引起草地群落多样性的丧失(Hautier et al., 2009; Isbell et al., 2013; Niu et al., 2018; Zhao et al., 2019)。目前, 养分富集背景下的草地生物量与多样性变化机制已成为草地生态学关注的热点问题。
氮是全球范围内限制草地植物生长的最重要元素, 适当的氮沉降可以补充土壤养分, 在一定程度上能够缓解植物生长所受到的氮限制, 对植物生长起到良好的促进作用(LeBauer & Treseder, 2008)。氮的富集通常会导致喜氮且可以无性繁殖的禾草类物种生物量增加, 进而增加群落整体的生物量(李禄军等, 2010; Xu et al., 2015; 王玉冰等, 2020), 该过程往往会通过光竞争来降低群落物种多样性, 因为禾草类物种高度较高, 在氮添加后占据群落上层空间, 造成群落下层的光资源减少与稀有种丧失(Wedin & Tilman, 1996; 杨倩等, 2018)。磷也是草地生态系统的重要限制养分, 尽管它的限制范围不及氮广泛(Vance et al., 2003; Soons et al., 2017; Gao et al., 2018); 但仍然有一些研究发现, 添加磷也可以改变地上生物量、物种组成和物种丰富度(张彦东等, 2004; Fay et al., 2015; 王玲等, 2019)。更为重要的是, Elser等(2007)发现, 氮和磷在陆地生态系统中具有类似的限制效应, 同时添加氮和磷组合比二者单独添加所产生的影响更为显著; 杨元武等(2017)也认为草原初级生产力同时受到氮和磷的限制。此外, 钾也是植物生长的限制因子。虽然Janssens等(1998)发现钾对植物生长的影响大于磷, 但大多数实验并没有发现钾的显著影响(贺星, 2014; 周小龙, 2016)。不同的草地类型其土壤性质与群落物种组成往往差异较大, 而阐明不同草地类型的养分限制状况可以为提高草地生产力提供重要信息, 具有重要的理论与实践意义。
群落物种多样性对养分添加的响应机理比较复杂, 大量研究表明养分添加导致植物群落物种多样性减少(Ren et al., 2010; 贺星, 2014; 周小龙, 2016; 杜忠毓等, 2020)。近年来, Harpole和Tilman (2007)、Harpole等(2016)提出的生态位维度-多样性假说受到越来越多的关注, 该假说认为群落中的物种通过土壤中的限制性资源维系其共存关系, 养分添加导致土壤中限制性资源减少进而导致土壤生态位维度减少, 最终使得多样性下降。同时也有研究表明, 养分添加后物种多样性增加(张杰琦等, 2010)或维持不变(丁睿等, 2017)。造成植物物种多样性不同程度变化的原因可能在于土壤发育环境、养分循环特征、物种初始组成和非生物因子(如降雨和气温等)差异所致(沈景林等, 1999)。
多样性与生产力之间的关系一直是生态学的研究热点(Tilman & Downing, 1994; Huberty et al., 1998; 王长庭等, 2005; 于丽等, 2015; Gao et al., 2018; 刘万第等, 2021), 但其结果却并不一致。全球尺度上的meta分析结果显示, 二者关系表现各异(Tang et al., 2017), 例如Waide等(1999)研究中发现大约200个生产力与多样性的关系中30%呈现单峰曲线关系, 26%呈现线性正相关关系, 12%呈现线性负相关关系, 32%显示二者关系不显著。针对多样性与生产力之间关系的一般模式以及产生这些模式的机制, 生态学家们进行了大量的实验研究提出诸多假说予以解释, 例如生态位互补假说和质量比假说。生态位互补假说认为生态系统功能由群落内物种的生态位分化程度所决定, 生态位分化程度高则能充分地利用各类资源有助于增强生态系统的功能(Tilman et al., 1997)。质量比假说则认为群落的地上生物量取决于群落中占优势的物种, 与群落的物种数无关(Aarssen, 1997; Tilman et al., 1997, 2001)。生态位互补假说可以很好地解释生物量与多样性之间的正相关关系(周小龙, 2016; 刘万第等, 2021), 而质量比假说则可以解释二者之间的负相关关系(Finegan et al., 2015; 周小龙, 2016)。目前在高寒草地中生产力变化受何种机制调控, 仍然是尚未解决的问题。
巴音布鲁克草原位于天山南麓, 是新疆最大的畜牧业基地, 也是新疆乃至中亚的生物多样性热点地区。同时又因其特殊的地理位置, 高寒的气候条件, 植物生长对环境的变化十分敏感, 也是生态脆弱区和气候变化的敏感区(Zhou et al., 2019)。基于上述原因, 本研究以天山高寒草地为研究对象, 通过多重养分添加实验, 分析不同元素添加处理组合条件下, 天山高寒草地群落物种多样性和地上生物量的变化差异, 以及二者关系的响应模式, 最终解析二者关系的潜在生态学机制。
1 材料和方法
1.1 研究区域概况
本研究的野外工作依托中国科学院巴音布鲁克草原生态系统研究站(42.88°-42.89° N, 83.70°- 83.72° E)开展。研究地点海拔约2 470 m, 年降水量265.7 mm, 年平均气温-4.8 ℃, 年蒸发量1 022.9- 1 247.5 mm, 年日照时间2 466-2 616 h, 全年积雪日达150-180天, 无绝对无霜期, 属于典型的高寒气候。植被类型为高寒草原, 优势物种为羊茅(Festuca ovina)、  草(Koeleria macrantha)、草地早熟禾(Poa pratensis)等。
草(Koeleria macrantha)、草地早熟禾(Poa pratensis)等。
1.2 实验设计
本研究基于全球营养网络(NutNet)巴音布鲁克站点的实验基础开展, 选择常年禁牧、生境均匀, 植被长势良好的典型高寒草原为研究对象, 通过使用尿素、磷酸二氢钙和硫酸钾, 换算为实际氮、磷和钾含量, 以10 g·m-2为养分添加单位, 进行不同养分类型的添加处理组合, 包括对照、氮(N)、磷(P)、钾(K)、N + P、N + K、P + K和N + P + K, 共计8个处理, 每个处理3次重复, 共计24个样方。实验布局采用随机区组设计, 每个处理的小区面积为25 m2 (5 m × 5 m), 各个样地之间用1 m的缓冲带隔开。2018年进行背景调查, 养分添加试验于2019年开始进行, 将预先称好的颗粒状肥料一次性均匀撒在样方内进行养分添加。养分添加时间为每年的5月中旬, 为了保证养分添加效果, 养分添加选择在阴雨天进行。
1.3 测定方法
2019年8月中旬和2020年9月中旬(因新冠疫情原因推迟)进行植被调查, 并采用收获法对地上生物量进行测定。为避免边缘效应, 实验均在每个样方的核心区(4 m × 4 m)内进行, 在每个样方核心区内设置2个0.1 m × 1 m小样方用于植物群落组成和地上生物量的测定, 先统计小样方内每个物种的个数, 有无性分株的禾本科个体只记为1株; 然后将小样方内所有植物分物种剪下装入信封, 在65 ℃下烘48 h至恒质量后将地上生物量按物种称量(精度为0.01 g), 对同一样方的2个0.1 m × 1 m小样方内各个物种的干物质质量累加计算地上生物量(g·m-2)。
1.4 植物多样性测定
采用物种丰富度、香农威纳指数(H')、辛普森指数(D)和均匀度指数(E)对群落的多样性进行评价, 本研究中物种丰富度指数用0.2 m2样方内出现的物种数表示, 即物种丰富度=出现在0.2 m2样方内的物种数。其余指数计算公式如下:
1.5 数据处理
物种的相对多度定义为每个物种的生物量除以样方中所有物种的生物量总和, 为了评估各个处理中物种相对多度(SRA)的变化, 计算了每个处理中物种相对多度的对数响应比(RSRA), 计算公式为RSRA = log(SRAf/SRAc), 其中SRAf与SRAc分别为养分添加和对照处理6个重复中的相对多度的均值, 若RSRA为正值则代表施肥以后该物种的相对多度是增加的; 若RSRA为负值则代表施肥以后该物种的相对多度是减小的(Niu et al., 2010; Zhou et al., 2016)。使用单因素方差分析和多重比较(HSD)对不同养分添加处理中的植物群落物种丰富度、地上生物量、物种多样性进行差异分析。使用一元线性回归对地上生物量和物种多样性之间的关系进行分析。所有数据利用SPSS 20.0软件进行统计分析和GraphPad Prism 8.0软件作图。
2 结果和分析
2.1 养分添加对群落内常见种相对多度的影响
分析养分添加对群落内10种常见种相对多度的影响, 结果显示: 包含氮添加的处理中(N、N + P、N + K、N + P + K) 4种禾草类(羊茅、冰草(Agropyron cristatum)、  草、草地早熟禾)的相对多度显著增加, 但是养分添加对天山赖草(Leymus tianschanicus)和细果薹草(Carex stenocarpa)的相对多度的影响没有一致规律。与禾草类相反, 包含氮添加的处理中(N、N + P、N + K、N + P + K) 2种豆科植物(小花棘豆(Oxytropis glabra)和高山黄耆(Astragalus alpinus)的相对多度降低。养分添加对2种杂草类(莓叶委陵菜(Potentilla fragarioides)和二裂委陵菜(P. bifurca)相对多度的影响没有一致规律(表1)。
草、草地早熟禾)的相对多度显著增加, 但是养分添加对天山赖草(Leymus tianschanicus)和细果薹草(Carex stenocarpa)的相对多度的影响没有一致规律。与禾草类相反, 包含氮添加的处理中(N、N + P、N + K、N + P + K) 2种豆科植物(小花棘豆(Oxytropis glabra)和高山黄耆(Astragalus alpinus)的相对多度降低。养分添加对2种杂草类(莓叶委陵菜(Potentilla fragarioides)和二裂委陵菜(P. bifurca)相对多度的影响没有一致规律(表1)。
表1 2020年氮(N)、磷(P)、钾(K)养分添加对天山高寒草地群落内常见物种相对多度的影响
Table 1
| 种名 Species | FG | RSRAN | RSRAP | RSRAK | RSRAN + P | RSRAN + K | RSRAP + K | RSRAN + P + K |
|---|---|---|---|---|---|---|---|---|
| 冰草 Agropyron cristatum | G | 0.45 | 0.37 | 0.57 | 0.88 | 1.28 | 0.31 | 0.78 |
| 草地早熟禾 Poa pratensis | G | 0.45 | 0.53 | 0.14 | 0.67 | 0.34 | 0.09 | 0.84 |
| G | -0.03 | -0.23 | 0.18 | 0.29 | 0.03 | 0.17 | 0.38 | |
| 天山赖草 Leymus tianschanicus | G | 0.48 | 0.06 | -1.04 | -1.49 | 0.21 | -0.20 | -0.13 |
| 细果薹草 Carex stenocarpa | G | -0.12 | 0.02 | -0.04 | 0.37 | -0.15 | -0.20 | 0.50 |
| 羊茅 Festuca ovina | G | 0.50 | 0.09 | 0.17 | 0.53 | 0.49 | 0.08 | 0.54 |
| 二裂委陵菜 Potentilla bifurca | F | 0.15 | 0.02 | -0.15 | 0.08 | -0.33 | -0.09 | 0.07 |
| 莓叶委陵菜 Potentilla fragarioides | F | 0.23 | -0.67 | -0.29 | -0.09 | 0.12 | -0.32 | -0.44 |
| 高山黄耆 Astragalus alpinus | L | -0.56 | 0.28 | 0.01 | -0.97 | -1.06 | 0.17 | -0.84 |
| 小花棘豆 Oxytropis glabra | L | -0.29 | -0.08 | 0.41 | -1.30 | 0.27 | -0.49 | -0.57 |
RSRA, 相对多度对数响应比。FG, 功能群。F, 杂类草; G, 禾草类; L, 豆科。
RSRA, log response of species relative abundance. FG, functional group. F, forb; G, grass; L, legume.
2.2 养分添加对天山高寒草地物种多样性的影响
用物种丰富度、香农威纳指数、辛普森指数和均匀度指数反映养分添加对群落植物多样性的影响。结果显示, 在所有的处理中物种丰富度几乎不发生变化(图1A、1B)。2019年均匀度指数几乎没有发生变化(图1G)。与对照相比, 2019年的香农威纳指数和辛普森指数只在N + P + K处理中差异显著(图1C、1E)。2020年香农威纳指数对P、N + P、N + K、P + K、N + P + K处理均响应显著(图1D), 2020年辛普森指数对N + P、N + K、N + P + K处理响应显著 (图1F), 2020年均匀度指数对P、N + P、N + K、P + K、N + P + K处理均响应显著(图1H)。此外, K添加对上述多样性指标几乎都不产生影响。
图1
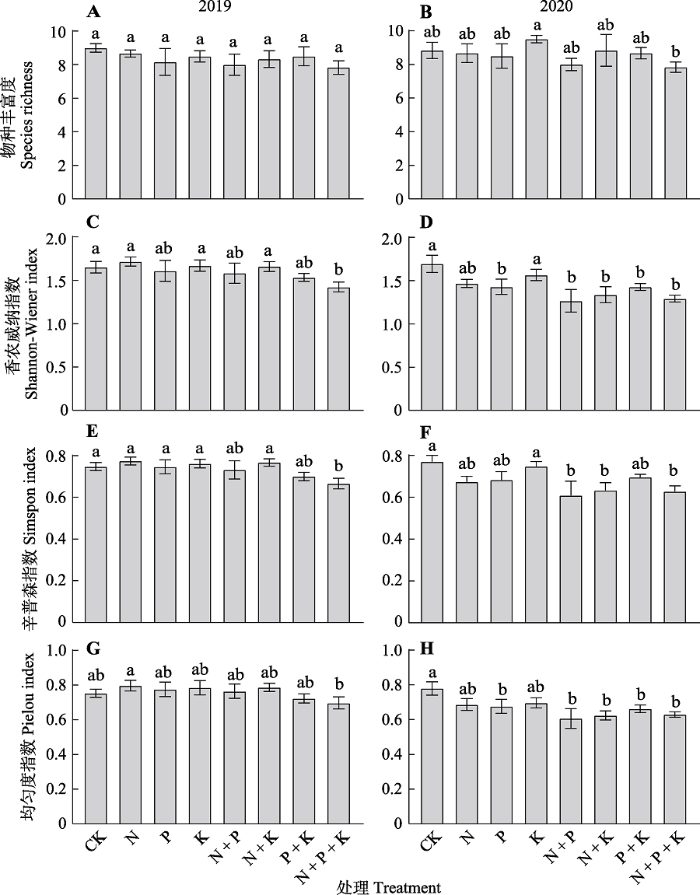
图1
养分添加对天山高寒草地群落物种多样性指数的影响(平均值±标准误, n = 6)。CK, 对照; N, 氮添加; P, 磷添加; K, 钾添加。不同的小写字母表示不同处理间差异显著(p < 0.05)。
Fig. 1
Effects of nutrient addition on plant species diversity index of alpine grassland community in Tianshan Mountains (mean ± SE, n = 6). CK, control; N, nitrogen addition; P, phosphorus addition; K, potassium addition. Different lowercase letters indicate significant differences among different treatments (p < 0.05).
2.3 养分添加对天山高寒草地地上生物量的影响
图2
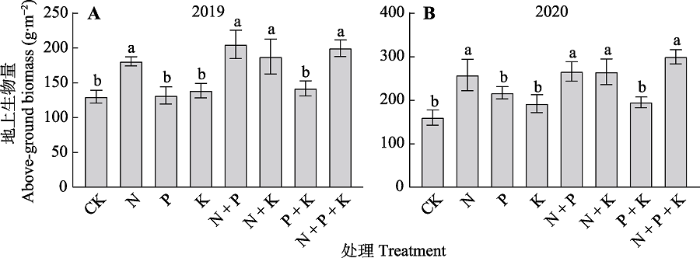
图2
养分添加对天山高寒草地地上生物量的影响(平均值±标准误, n = 6)。CK, 对照; N, 氮添加; P, 磷添加; K, 钾添加。不同的小写字母表示不同处理间差异显著(p < 0.05)。
Fig. 2
Effects of nutrient addition on above-ground biomass of alpine grassland in Tianshan Mountains (mean ± SE, n = 6). CK, control; N, nitrogen addition; P, phosphorus addition; K, potassium addition. Different lowercase letters indicate significant differences among different treatments (p < 0.05).
图3
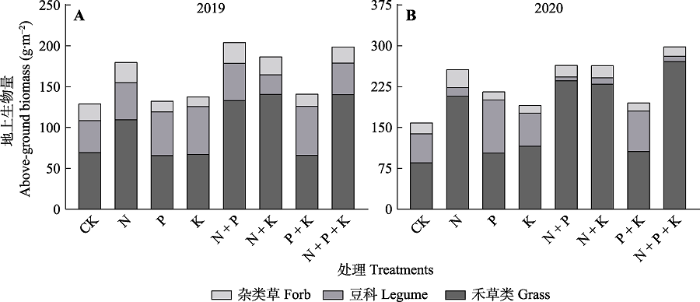
图3
养分添加对天山高寒草地植物功能群地上生物量的影响。CK, 对照; N, 氮添加; P, 磷添加; K, 钾添加。
Fig. 3
Effects of nutrient addition on aboveground biomass of plant functional groups in alpine grassland of Tianshan Mountains CK, control; N, nitrogen addition; P, phosphorus addition; K, potassium addition.
2.4 养分添加条件下地上生物量与物种多样性的关系
图4
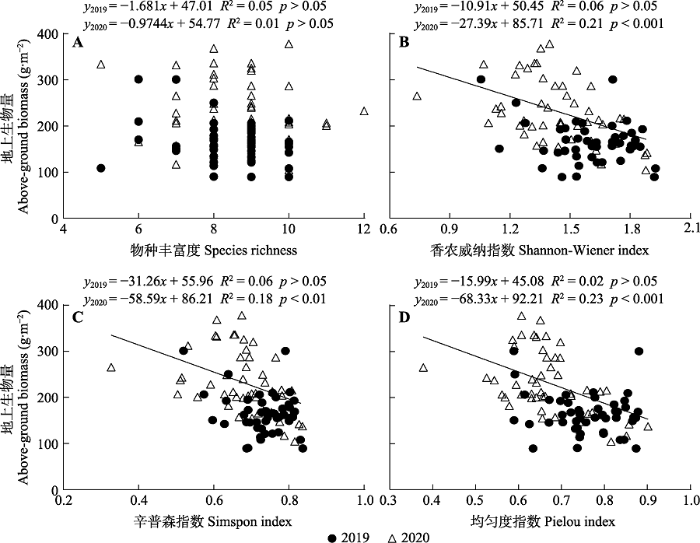
图4
养分添加下天山高寒草地地上生物量与物种多样性之间的关系。
Fig. 4
Relationship between aboveground biomass and species diversity in alpine grassland of Tianshan Mountains under nutrient addition.
3 讨论和结论
3.1 多重养分添加导致物种多样性下降
通过连续两年养分添加实验, 结果发现氮添加及其组合(除2020年N + P + K组合外)对物种丰富度几乎没有影响, 但是导致当地群落香农维纳指数与辛普森指数降低(图1), 究其原因在于养分添加后, 一些养分利用效率高的物种(如羊茅和  草)会快速生长且逐渐占据优势(Tian et al., 2016), 从而减少了其他物种(如莓叶委陵菜)所需的养分和空间, 进而造成稀有种数量减少甚至消失(表1; 图3)。同时, 在包含氮添加的处理中, 豆科物种因为固氮优势的丧失而引起相对多度减少(Salvagiotti et al., 2008; Nyfeler et al., 2011; 李强等, 2021)(表1)和生物量降低(图3), 最终导致当地群落均匀度下降(图1G、1H), 群落物种组成由禾草类(对照中53%)与豆科(对照中33%)共同占优势转变为禾草类占据绝对优势(N + P + K处理中91%)。大量研究表明物种对光资源的竞争也是导致群落组成和物种多样性变化的主要因素(Hautier et al., 2009; 杨倩等, 2018), 即养分添加使土壤中的可利用养分增加, 物种之间的竞争由地下部分对养分的竞争转变为地上部分对光的竞争, 由于那些个体高度较高的物种(如禾草类)通过增加自身光合速率以及对周围植物的遮阴, 导致底层的光限制加剧, 进而排除小个体的物种(李禄军等, 2010; 杜忠毓等, 2020)。值得注意的是在本研究中, 相比较单独养分添加而言, 多重养分添加更易导致物种多样性的丧失, 尤其在N + P + K添加处理条件下, 该结果与经典的罗斯坦德公园草地实验(Silvertown et al., 2006)一致, 说明养分添加导致生态位维度的下降是导致当地植物群落多样性丧失的重要原因, 支持生态位维度-多样性假说(图1)(Harpole & Tilman, 2007; Harpole et al., 2016), 该假说认为物种对土壤限制资源的生态位分化是驱动物种多样性共存的主要机制, 即物种共存通过限制资源的数目而维系物种间的共存, 当受到一或两个因素的限制时大量物种可以共存, 而增加多种限制性资源的供应可以降低生态位维度进而导致物种丧失。
草)会快速生长且逐渐占据优势(Tian et al., 2016), 从而减少了其他物种(如莓叶委陵菜)所需的养分和空间, 进而造成稀有种数量减少甚至消失(表1; 图3)。同时, 在包含氮添加的处理中, 豆科物种因为固氮优势的丧失而引起相对多度减少(Salvagiotti et al., 2008; Nyfeler et al., 2011; 李强等, 2021)(表1)和生物量降低(图3), 最终导致当地群落均匀度下降(图1G、1H), 群落物种组成由禾草类(对照中53%)与豆科(对照中33%)共同占优势转变为禾草类占据绝对优势(N + P + K处理中91%)。大量研究表明物种对光资源的竞争也是导致群落组成和物种多样性变化的主要因素(Hautier et al., 2009; 杨倩等, 2018), 即养分添加使土壤中的可利用养分增加, 物种之间的竞争由地下部分对养分的竞争转变为地上部分对光的竞争, 由于那些个体高度较高的物种(如禾草类)通过增加自身光合速率以及对周围植物的遮阴, 导致底层的光限制加剧, 进而排除小个体的物种(李禄军等, 2010; 杜忠毓等, 2020)。值得注意的是在本研究中, 相比较单独养分添加而言, 多重养分添加更易导致物种多样性的丧失, 尤其在N + P + K添加处理条件下, 该结果与经典的罗斯坦德公园草地实验(Silvertown et al., 2006)一致, 说明养分添加导致生态位维度的下降是导致当地植物群落多样性丧失的重要原因, 支持生态位维度-多样性假说(图1)(Harpole & Tilman, 2007; Harpole et al., 2016), 该假说认为物种对土壤限制资源的生态位分化是驱动物种多样性共存的主要机制, 即物种共存通过限制资源的数目而维系物种间的共存, 当受到一或两个因素的限制时大量物种可以共存, 而增加多种限制性资源的供应可以降低生态位维度进而导致物种丧失。
3.2 养分添加增加了群落及禾草类生物量
研究结果表明养分添加能显著增加高寒草地植物群落的地上生物量, 而这种增加主要是由于禾草类生物量增加所导致, 与之相反的是豆科和杂类草在总生物量中所占比例降低(图2, 图3), 这与前人的研究结果(邱波和罗燕江, 2004; Song et al., 2011; 杨晓霞等, 2014; 于丽等, 2015; Tian et al., 2016)一致, 其原因是禾草类植物具有较高的氮利用率(Song et al., 2011), 同时禾草类物种植株高度较高, 在群落中占据上层空间以获取更多的光资源, 在养分充足时可以更快生长, 尤其是氮、磷的添加缓解了土壤养分对禾草类植物生长的限制, 其地上部分迅速生长导致禾草类生物量增加(邱波和罗燕江, 2004; Tian et al., 2016; 张馨文等, 2021)。同以前的研究(Ren et al., 2010; 贺星, 2014; 周小龙, 2016)一致, 本研究表明2019和2020年氮添加处理都能使草地植物群落地上生物量提高, 而磷、钾的单独添加以及混合添加对地上生物量都无显著影响(图2), 该结果说明在本研究区域中, 氮为生物量的优先限制养分, 而非磷、钾。另一方面, 凡是包含氮元素的养分添加处理(N、N + P、N + K、N + P + K)都显著地提高生物量, 尤其是N + P + K混合养分添加处理条件下的地上生物量增加以及物种多样性减少都要比单独的养分添加处理更为剧烈, 这个结果表明在氮、磷和钾之间存在着交互作用, 解除氮限制以后磷和钾成为限制性养分, 这充分说明与养分的单独添加相比, 混合添加对草地植物群落地上生物量影响更为显著(周小龙, 2016; Gao et al., 2018)。在本研究中, 相比较第一年而言, 第二年养分添加的效果更加明显, 这可能是土壤中的养分逐渐累积的效果使然。
3.3 群落优势种而非物种数决定养分添加条件下的地上生物量
在本研究中, 2019年植物群落地上生物量与4种多样性指数无显著关系。养分添加两年之后, 植物群落地上生物量与植物丰富度依然无显著回归关系; 但是植物群落地上生物量与香农威纳指数、辛普森指数和均匀度指数呈显著的负线性回归关系(图3), 这与青藏高原高寒草甸、内蒙古针茅草原以及罗斯坦德公园草地实验的结果(Crawley et al., 2005; 马涛等, 2008; Silvertown et al., 2006)一致。当前研究对多样性和生产力关系解释各异(江小雷等, 2004; Xia & Wan, 2008; Gao et al., 2018; Liu et al., 2021)。本研究表明养分添加后, 养分利用效率高的禾本科植物生物量增加导致群落地上生物量增加, 与前人研究(李禄军等, 2010; Xu et al., 2015; 王玉冰等, 2020)一致, 即群落地上生物量主要取决于群落中占优势的物种(如N + P + K处理中占23%的羊茅和占53%的  草), 同时回归分析结果表明生物量与物种丰富度之间不存在显著回归关系, 本实验结果表明质量比假说是养分添加条件下天山高寒草地生物量变化的重要机制。
草), 同时回归分析结果表明生物量与物种丰富度之间不存在显著回归关系, 本实验结果表明质量比假说是养分添加条件下天山高寒草地生物量变化的重要机制。
综上所述, 本研究表明: 1)短期的养分添加降低了群落的香农威纳指数与辛普森指数, 但不会导致当地群落丰富度剧烈下降。2)氮是天山高寒草地地上生物量的主要限制性养分, 氮添加以后地上生物量显著增加; 在解除氮限制以后磷和钾成为限制性养分。3)养分添加后群落生物量增加主要由禾草类生物量增加导致, 当地群落的地上生物量主要由优势种所决定, 而非群落物种数决定。
致谢
感谢中国科学院巴音布鲁克草原生态系统研究站在野外工作中提供便利。
参考文献
High productivity in grassland ecosystems: effected by species diversity or productive species?
DOI:10.2307/3546531 URL [本文引用: 1]
Nitrogen-induced terrestrial eutrophication: cascading effects and impacts on ecosystem services
DOI:10.1002/ecs2.1877 [本文引用: 1]
Determinants of species richness in the Park Grass Experiment
DOI:10.1086/427270
PMID:15729649
[本文引用: 1]

The Park Grass Experiment at Rothamsted in southeast England was started in 1856, making it the longest-running experiment in plant ecology anywhere in the world. Experimental inputs include a range of fertilizers (nitrogen, phosphorus, potassium, and organic manures) applied annually, with lime applied occasionally, and these have led to an increase in biomass and, where nitrogen was applied in the form of ammonium sulfate, to substantial decreases in soil pH. The number of species per plot varies from three to 44 per 200 m(2), affording a unique opportunity to study the determinants of plant species richness and to estimate the effect sizes attributable to different factors. The response of species richness to biomass depends on the amount and type of nitrogen applied; richness declined monotonically with increasing biomass on plots receiving no nitrogen or receiving nitrogen in the form of sodium nitrate, but there was no relationship between species richness and biomass on plots acidified by ammonium sulfate application. The response to lime also depended on the type of nitrogen applied; there was no relationship between lime treatment and species richness, except in plots receiving nitrogen in the form of ammonium sulfate, where species richness increased sharply with increasing soil pH. The addition of phosphorus reduced species richness, and application of potassium along with phosphorus reduced species richness further, but the biggest negative effects were when nitrogen and phosphorus were applied together. The analysis demonstrates how multiple factors contribute to the observed diversity patterns and how environmental regulation of species pools can operate at the same spatial and temporal scale as biomass effects.
Analysis of current research situation on impact of nitrogen & phosphorus addition on species diversity and productivity of grassland ecological system
氮磷添加对草原生态系统物种多样性及生产力的影响研究现状分析
Effects of nutrient addition and precipitation manipulation on plant species diversity and biomass of a desert grassland
养分添加和降水变化对荒漠草原植物群落物种多样性和生物量的影响
Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems
DOI:10.1111/j.1461-0248.2007.01113.x URL [本文引用: 1]
The responses of ecosystems to global warming
生态系统对全球变暖的响应
Grassland productivity limited by multiple nutrients
DOI:10.1038/nplants.2015.80 URL [本文引用: 1]
Does functional trait diversity predict above-ground biomass and productivity of tropical forests? Testing three alternative hypotheses
DOI:10.1111/1365-2745.12346 URL [本文引用: 1]
Transformation of the nitrogen cycle: recent trends, questions, and potential solutions
DOI:10.1126/science.1136674
PMID:18487183
[本文引用: 1]
Humans continue to transform the global nitrogen cycle at a record pace, reflecting an increased combustion of fossil fuels, growing demand for nitrogen in agriculture and industry, and pervasive inefficiencies in its use. Much anthropogenic nitrogen is lost to air, water, and land to cause a cascade of environmental and human health problems. Simultaneously, food production in some parts of the world is nitrogen-deficient, highlighting inequities in the distribution of nitrogen-containing fertilizers. Optimizing the need for a key human resource while minimizing its negative consequences requires an integrated interdisciplinary approach and the development of strategies to decrease nitrogen-containing waste.
Nitrogen, not phosphorus, enrichment controls biomass production in alpine wetlands on the Tibetan Plateau, China
DOI:10.1016/j.ecoleng.2018.02.016 URL [本文引用: 4]
Addition of multiple limiting resources reduces grassland diversity
DOI:10.1038/nature19324 URL [本文引用: 2]
Grassland species loss resulting from reduced niche dimension
DOI:10.1038/nature05684 URL [本文引用: 2]
Competition for light causes plant biodiversity loss after eutrophication
DOI:10.1126/science.1169640
PMID:19407202
[本文引用: 2]
Human activities have increased the availability of nutrients in terrestrial and aquatic ecosystems. In grasslands, this eutrophication causes loss of plant species diversity, but the mechanism of this loss has been difficult to determine. Using experimental grassland plant communities, we found that addition of light to the grassland understory prevented the loss of biodiversity caused by eutrophication. There was no detectable role for competition for soil resources in diversity loss. Thus, competition for light is a major mechanism of plant diversity loss after eutrophication and explains the particular threat of eutrophication to plant diversity. Our conclusions have implications for grassland management and conservation policy and underscore the need to control nutrient enrichment if plant diversity is to be preserved.
Effects of Nutrient Addition on Community Biomass and Diversity Varied Among Different Grassland Ecosystems of Inner Mongolia
养分添加对内蒙古草原生物量和多样性的影响
Effects of nitrogen addition on successional dynamics and species diversity in Michigan old-fields
DOI:10.1046/j.1365-2745.1998.8650794.x URL [本文引用: 1]
Nutrient enrichment, biodiversity loss, and consequent declines in ecosystem productivity
Relationship between soil chemical factors and grassland diversity
DOI:10.1023/A:1004389614865 URL [本文引用: 1]
Effects of plant species diversity on productivity of ecosystem
植物群落物种多样性对生态系统生产力的影响
Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed
Our meta-analysis of 126 nitrogen addition experiments evaluated nitrogen (N) limitation of net primary production (NPP) in terrestrial ecosystems. We tested the hypothesis that N limitation is widespread among biomes and influenced by geography and climate. We used the response ratio (R approximately equal ANPP(N)/ANPP(ctrl)) of aboveground plant growth in fertilized to control plots and found that most ecosystems are nitrogen limited with an average 29% growth response to nitrogen (i.e., R = 1.29). The response ratio was significant within temperate forests (R = 1.19), tropical forests (R = 1.60), temperate grasslands (R = 1.53), tropical grasslands (R = 1.26), wetlands (R = 1.16), and tundra (R = 1.35), but not deserts. Eight tropical forest studies had been conducted on very young volcanic soils in Hawaii, and this subgroup was strongly N limited (R = 2.13), which resulted in a negative correlation between forest R and latitude. The degree of N limitation in the remainder of the tropical forest studies (R = 1.20) was comparable to that of temperate forests, and when the young Hawaiian subgroup was excluded, forest R did not vary with latitude. Grassland response increased with latitude, but was independent of temperature and precipitation. These results suggest that the global N and C cycles interact strongly and that geography can mediate ecosystem response to N within certain biome types.
Effects of fertilizations on species composition and diversity of grassland in Keerqin Sandy Lands
施肥对科尔沁沙质草地群落物种组成和多样性的影响
Mechanism of the trade-off between biological nitrogen fixation and phosphorus acquisition strategies of herbaceous legumes under nitrogen and phosphorus addition
DOI:10.17521/cjpe.2020.0241 URL [本文引用: 1]
土壤氮磷添加下豆科草本植物生物固氮与磷获取策略的权衡机制
Plant diversity in the process of succession of artificial vegetation types and environment in an arid desert region of China
我国干旱沙漠地区人工植被与环境演变过程中植物多样性的研究
以我国干旱沙漠区流沙治理的成功模式包兰铁路沙坡头地段人工植被防护体系作为研究对象,研究了区域人工植被与环境演变过程中植物多样性的变化特点。研究表明:该区人工植被经过40余年的演变其植物种的组成趋于动态平衡;植物多样性在时间尺度上的变化表现为随群落演替的进行而增大,42年前(1956年)建立的人工植被多样性指数达到D=0.706~0.822或H′=1.393~1.893,10年前(1987年)建立的人工植被其多样性仅为D=0.501~0.702或H′=0.819~1.074;β多样性的测度表明,沙坡头地区人工植被在其演变的历程中经历了2次物种周转速率相对较快的阶段,这一特点与植被演替密切相关,对干旱沙漠地区的生态恢复和人工绿洲建设与管理具有重要的参考价值。本文还进一步研究了土壤基质条件等变化对植物多样性的影响。
Community diversity, patterns of productivity, and factors influencing them in Stipa in Ningxia grassland
宁夏草原针茅属植物群落物种多样性和生产力格局及影响因素研究
Relationships of species diversity and productivity under the fertilization levels in alpine meadow of the Qinghai-Tibetan Plateau
不同施肥处理高寒草甸植物群落物种多样性与生产力的关系
The impact of nitrogen enrichment on grassland ecosystem stability depends on nitrogen addition level
DOI:10.1016/j.scitotenv.2017.09.318 URL [本文引用: 1]
Linking grazing response of species abundance to functional traits in the Tibetan alpine meadow
DOI:10.1007/s11104-009-0194-8 URL [本文引用: 1]
Grass-legume mixtures can yield more nitrogen than legume pure stands due to mutual stimulation of nitrogen uptake from symbiotic and non-symbiotic sources
DOI:10.1016/j.agee.2010.11.022 URL [本文引用: 1]
Effects of fertilizer gradients on productivity and species diversity in a degraded alpine meadow
不同施肥梯度对甘南退化高寒草甸生产力和物种多样性的影响
Effects of resource additions on species richness and ANPP in an alpine meadow community
DOI:10.1093/jpe/rtp034 URL [本文引用: 2]
Nitrogen uptake, fixation and response to fertilizer N in soybeans: a review
DOI:10.1016/j.fcr.2008.03.001 URL [本文引用: 1]
Experimental study of improvement measures on the depredated grassland in alpine areas
高寒地区退化草地改良试验研究
The Park Grass Experiment 1856-2006: its contribution to ecology
DOI:10.1111/j.1365-2745.2006.01145.x URL [本文引用: 2]
Nitrogen enrichment enhances the dominance of grasses over forbs in a temperate steppe ecosystem
DOI:10.5194/bg-8-2341-2011 URL [本文引用: 2]
Nitrogen effects on plant species richness in herbaceous communities are more widespread and stronger than those of phosphorus
DOI:10.1016/j.biocon.2016.12.006 URL [本文引用: 1]
The effect of nitrogen addition on community structure and productivity in grasslands: a meta-analysis
DOI:10.1016/j.ecoleng.2016.11.039 URL [本文引用: 1]
A novel soil manganese mechanism drives plant species loss with increased nitrogen deposition in a temperate steppe
DOI:10.1890/15-0917.1 URL [本文引用: 3]
Biodiversity and stability in grasslands
DOI:10.1038/367363a0 URL [本文引用: 1]
The influence of functional diversity and composition on ecosystem processes
DOI:10.1126/science.277.5330.1300 URL [本文引用: 2]
Diversity and productivity in a long-term grassland experiment
Plant diversity and niche complementarity had progressively stronger effects on ecosystem functioning during a 7-year experiment, with 16-species plots attaining 2.7 times greater biomass than monocultures. Diversity effects were neither transients nor explained solely by a few productive or unviable species. Rather, many higher-diversity plots outperformed the best monoculture. These results help resolve debate over biodiversity and ecosystem functioning, show effects at higher than expected diversity levels, and demonstrate, for these ecosystems, that even the best-chosen monocultures cannot achieve greater productivity or carbon stores than higher-diversity sites.
Phosphorus acquisition and use: critical adaptations by plants for securing a nonrenewable resource
DOI:10.1046/j.1469-8137.2003.00695.x URL [本文引用: 1]
The relationship between productivity and species richness
DOI:10.1146/annurev.ecolsys.30.1.257 URL [本文引用: 1]
Relationship between species diversity and productivity in four types of alpine meadow plant communities
高寒草甸不同草地群落物种多样性与生产力关系研究
Influence of nitrogen addition on the primary production in Nei Mongol degraded grassland
DOI:10.17521/cjpe.2016.0141 URL [本文引用: 1]
氮素添加对内蒙古退化草原生产力的短期影响
DOI:10.17521/cjpe.2016.0141
[本文引用: 1]
不合理的土地利用方式以及气候变化导致我国草原生态系统普遍退化, 主要表现在土壤养分降低、植被覆盖度减少、生产力下降。外源氮素添加是促进退化草原尽快恢复的一项重要措施, 尤其是对那些退化较为严重的草原。该研究选取内蒙古东乌珠穆沁旗不同退化程度(轻度、中度和重度)的草原群落, 于2014-2015年开展连续两年的氮素添加实验, 设置对照(不添加)、低水平(5.0 g N·m<sup>-2</sup>·a<sup>-1</sup>)、中水平(10.0 g N·m<sup>-2</sup>·a<sup>-1</sup>)和高水平(20.0 g N·m<sup>-2</sup>·a<sup>-1</sup>) 4种氮素添加处理, 探讨退化草原群落生产力在恢复过程中对不同水平氮素添加的响应。结果显示: (1)高、中水平氮素添加显著提高了轻度退化群落的地上生物量, 分别比对照增加了53.1%、51.6%, 氮素各水平添加对中度、重度群落地上生物量无显著影响; (2)高、中水平氮素添加显著提高了轻度退化群落中多年生根茎型禾草地上生物量, 分别比对照增加了45.1%、47.7%, 而多年生杂类草地上生物量分别比对照减少了37.4%、42.1%, 但中度和重度退化群落各功能群生物量的响应不显著; (3)三种水平氮素添加对轻、中、重度退化群落物种丰富度在试验期间均没有显著影响。研究结果表明氮素添加有助于提高轻度退化草原中多年生根茎型禾草的生物量, 进而提高群落的生物量, 但多年生杂类草会被逐渐替代, 导致生物量降低, 可见施氮对草原恢复的影响取决于草原退化 程度。
Effects of nitrogen and phosphorus addition on community diversity and biomass of alpine steppe
氮、磷添加对高寒草原群落多样性和生物量的影响
Effects of grazing intensity on the biodiversity and productivity of meadow steppe
放牧强度对草甸草原生产力和多样性的影响
Effects and pathways of long-term nitrogen addition on plant diversity and primary productivity in a typical steppe
DOI:10.17521/cjpe.2019.0260 URL [本文引用: 2]
长期氮添加对典型草原植物多样性与初级生产力的影响及途径
DOI:10.17521/cjpe.2019.0260
[本文引用: 2]
氮(N)沉降对陆地生态系统的结构和功能已产生了重要的影响, N也是中国北方草原植物生长和初级生产力的主要限制性元素。物种多样性和功能多样性是揭示生物多样性对生态系统功能维持机制的关键指标, 然而, 关于长期N添加下草原物种多样性与功能多样性的关系, 及其对初级生产力的影响途径及机制, 尚不十分清楚。为此, 该研究依托在内蒙古典型草原建立的长期N添加实验平台, 实验处理包括1个完全对照(不添加任何肥料)和6个N添加水平(0、1.75、5.25、10.50、17.50和28.00 g·m<sup>-2</sup>·a<sup>-1</sup>), 研究了长期N添加对典型草原物种多样性、功能多样性和初级生产力的影响大小及途径。结果表明: 1) N添加显著降低了典型草原的物种丰富度和Shannon-Wiener指数, 但对功能多样性(包括功能性状多样性指数和群落加权性状值)无显著的影响。2)结构方程模型分析表明, 功能多样性主要受物种丰富度的影响, 但是物种多样性减少并没有导致功能多样性降低, 其原因主要是功能群组成发生了改变, 即群落内多年生根茎禾草所占比例显著增加, 以致群落加权性状值变化不大。3) N通过影响物种丰富度和功能群组成, 间接影响群落加权性状值, 进而影响群落净初级生产力。其中, 群落加权性状值是最重要的影响因子, 可解释48%的初级生产力变化, 表明初级生产力主要是由群落内优势物种的生物量及功能性状所决定, 因此该研究的结果很好地支持了质量比假说。
Influence of nitrogen loading and species composition on the carbon balance of grasslands
In a 12-year experimental study of nitrogen (N) deposition on Minnesota grasslands, plots dominated by native warm-season grasses shifted to low-diversity mixtures dominated by cool-season grasses at all but the lowest N addition rates. This shift was associated with decreased biomass carbon (C):N ratios, increased N mineralization, increased soil nitrate, high N losses, and low C storage. In addition, plots originally dominated by nonnative cool-season grasses retained little added N and stored little C, even at low N input rates. Thus, grasslands with high N retention and C storage rates were the most vulnerable to species losses and major shifts in C and N cycling.
Global response patterns of terrestrial plant species to nitrogen addition
DOI:10.1111/j.1469-8137.2008.02488.x URL [本文引用: 1]
Influences of nitrogen, phosphorus and silicon addition on plant productivity and species richness in an alpine meadow
DOI:10.1093/aobpla/plv125 URL [本文引用: 2]
Plant diversity change in grassland communities along a grazing disturbance gradient in the northeast china transect
中国东北样带草地群落放牧干扰植物多样性的变化
放牧干扰是草地群落植物多样性变化的主要影响因素之一。中国东北样带9个草地群落放牧干扰植物多样性变化的研究结果表明:中牧或重牧阶段Shannon指数达最大值,形成中牧(重牧)>重牧(中牧)>轻牧>过牧的规律。群落物种丰富度、均匀度与多样性的相关分析表明,均匀度变化对多样性变化具有更大的贡献率,而丰富度呈下降趋势,即轻牧(中牧)>中牧(轻牧)>重牧>过牧。生活型功能群多样性也表现出明显的变化。中国东北样带草地群落植物多样性的分布格局是:草甸草原>典型草原>典型草原 >荒漠草原>碱化草原,并且群落物种丰富度对多样性有更大贡献率。
Effects of nitrogen addition on the plant diversity and biomass of degraded grasslands of Nei Mongol, China
DOI:10.17521/cjpe.2017.0135 URL [本文引用: 2]
氮添加对内蒙古退化草地植物群落多样性和生物量的影响
DOI:10.17521/cjpe.2017.0135
[本文引用: 2]
人为干扰及气候变化导致内蒙古草地发生了大面积退化, 氮添加是促进退化草地生产力恢复的一项重要措施。该文基于2011年建立的氮肥添加实验平台, 以3个不同退化程度(中度退化、重度退化、极度退化)草地群落为研究对象, 设置对照、10、20、30、40和50 g·m <sup>-2</sup>·a <sup>-1</sup> 6种氮添加处理, 分析氮添加对退化草地恢复过程中群落多样性和生物量的影响。结果表明: (1)氮添加降低了中度、重度退化草地恢复进程中物种丰富度和多样性, 对极度退化草地恢复进程中物种丰富度和多样性无明显影响。(2)氮添加促进了3个不同退化程度草地恢复进程中群落地上生物量的增加。(3)氮添加显著增加了群落中禾草的地上生物量及其在群落地上生物量中所占的比例, 降低了杂类草在群落地上生物量中的比例, 但对杂类草地上生物量无显著影响。研究表明在利用施肥措施治理退化草地的过程中, 需要充分考虑草地退化程度以及由氮添加引起的群落多样性和生产力的改变对草地生态系统功能的影响。
Responses of plant community biomass to nitrogen and phosphorus additions in an alpine meadow on the Qinghai-Xizang Plateau
DOI:10.3724/SP.J.1258.2014.00014 URL [本文引用: 1]
青藏高原高寒草甸植物群落生物量对氮、磷添加的响应
DOI:10.3724/SP.J.1258.2014.00014
[本文引用: 1]
青藏高原正经历着明显的温暖化过程, 由此引起的土壤温度的升高促进了土壤中微生物的活性, 同时青藏高原东缘地区大气氮沉降十分明显, 并呈逐年增加的趋势, 这些环境变化均促使土壤中可利用营养元素增加, 因此深入了解青藏高原高寒草甸植物生物量对可利用营养元素增加的响应, 是准确预测未来全球变化背景下青藏高原高寒草甸碳循环过程的重要基础。该研究基于在青藏高原高寒草甸连续4年(2009-2012年)氮、磷添加后对不同功能群植物地上生物量、群落地上和地下生物量的测定, 探讨高寒草甸生态系统碳输入对氮、磷添加的响应。结果表明: (1)氮、磷添加均极显著增加了禾草的地上绝对生物量及其在群落总生物量中所占的比例, 同时均显著降低了杂类草在群落总生物量中的比例, 此外磷添加极显著降低了莎草地上绝对生物量及其在群落总生物量中所占的比例。(2)氮、磷添加均显著促进了青藏高原高寒草甸的地上生物量增加, 分别增加了24%和52%。(3)氮添加对高寒草甸地下生物量无显著影响, 而磷添加后地下生物量有增加的趋势。(4)氮添加对高寒草甸植物总生物量无显著影响, 而磷添加后植物总生物量显著增加。研究表明, 氮、磷添加可缓解青藏高原高寒草甸植物生长的营养限制, 促进植物地上部分的生长, 然而高寒草甸植物的生长极有可能更受土壤中可利用磷含量的限制。
Initial response of species diversity and productivity to nutrients addition on alpine meadow
高寒草甸物种多样性和生产力对养分添加的初期响应
Effects of nutrient addition on plant diversity and productivity in a Stipa baicalensis grassland in Inner Mongolia, China
养分添加对内蒙古贝加尔针茅草原植物多样性与生产力的影响
Effects of nitrogen addition on species richness and relationship between species richness and aboveground productivity of alpine meadow of the Qinghai-Tibetan Plateau, China
氮素添加对青藏高原高寒草甸植物群落物种丰富度及其与地上生产力关系的影响
DOI:10.3773/j.issn.1005-264x.2010.10.001
[本文引用: 1]
植物种群对有限资源的竞争是决定植物群落物种组成、多样性和生产力等群落结构和功能的主要因素。该文以青藏高原高寒草甸为研究对象, 研究了短期内不同水平的氮素添加对高寒草甸植物群落的影响。结果表明: 1)氮素添加提高了土壤中NO<sub>3</sub><sup>–</sup>-N等可利用资源的含量, 增加了植物群落植被的盖度, 减小了植被的透光率, 随着施氮量的增加, 群落中物种丰富度显著降低(p p p 3</sub><sup>–</sup>-N含量呈线性正相关(p 3</sub><sup>–</sup>-N等可利用资源的含量而对植物群落物种组成和地上生产力产生影响。
Effects of short-term nitrogen addition on plant community composition and stability of desert steppe
短期氮添加对荒漠草原植物群落组成及稳定性的影响
Fertilization effects of N, P on a grass community at the dry valley of Jinsha River
金沙江干旱河谷退化草地群落对氮磷施肥的反应
Community composition, structure and productivity in response to nitrogen and phosphorus additions in a temperate meadow
DOI:10.1016/j.scitotenv.2018.11.155 URL [本文引用: 2]
The Effect of Fertilization on Community Assembly and Production in Alpine Meadow Community
高寒草甸植物群落结构组建和生产力对施肥的响应机制
The competition- dispersal trade-off exists in forbs but not in graminoids: a case study from multispecies alpine grassland communities
DOI:10.1002/ece3.4856 URL [本文引用: 1]
The effects of fertilization on the trait-abundance relationships in a Tibetan alpine meadow community

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}