
Chin J Plant Ecol ›› 2020, Vol. 44 ›› Issue (1): 22-32.DOI: 10.17521/cjpe.2019.0260 cstr: 32100.14.cjpe.2019.0260
Special Issue: 生物多样性
• Research Articles • Previous Articles Next Articles
WANG Yu-Bing1,2,SUN Yi-Han1,2,DING Wei3,ZHANG En-Tao1,2,LI Wen-Huai1,CHI Yong-Gang4,*( ),ZHENG Shu-Xia1,*()
),ZHENG Shu-Xia1,*()
Received:2019-10-01
Revised:2020-01-09
Online:2020-01-20
Published:2020-02-24
Contact:
CHI Yong-Gang,ZHENG Shu-Xia
Supported by:WANG Yu-Bing,SUN Yi-Han,DING Wei,ZHANG En-Tao,LI Wen-Huai,CHI Yong-Gang,ZHENG Shu-Xia. Effects and pathways of long-term nitrogen addition on plant diversity and primary productivity in a typical steppe[J]. Chin J Plant Ecol, 2020, 44(1): 22-32.
Add to citation manager EndNote|Ris|BibTeX
URL: https://www.plant-ecology.com/EN/10.17521/cjpe.2019.0260
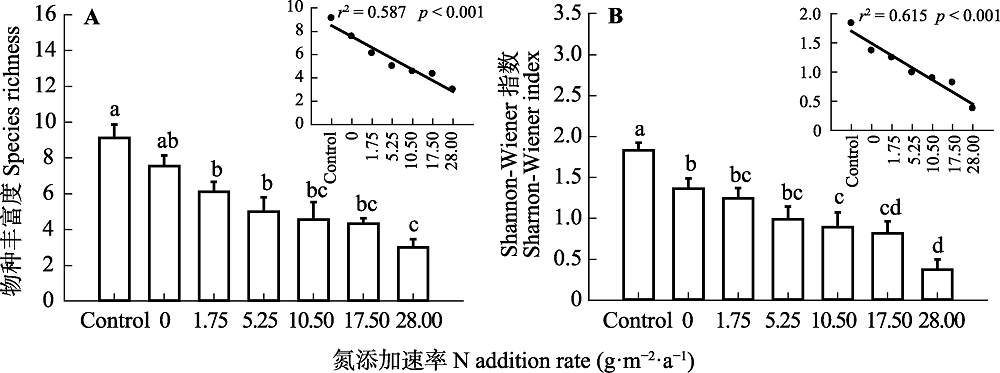
Fig. 1 Effects of N addition on species richness (A) and Shannon-Wiener index (B) in a typical steppe (mean + SE). Control, absolute control, no fertilizer added. Different lowercase letters indicate significant difference among treatments (p < 0.05).
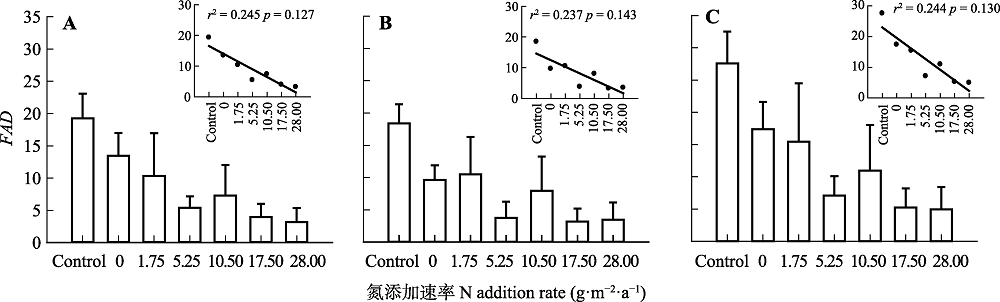
Fig. 2 Effect of N addition on functional attribute diversity (FAD) in a typical steppe (mean + SE). A, Plant trait functional diversity, calculated by plant height (PH), individual biomass (PB) and stem:leaf biomass ratio (SLR). B, Leaf trait functional diversity, calculated by specific leaf area (SLA), leaf dry mass content (LDMC), and leaf N content (LNC). C, Whole-plant trait functional diversity, calculated by six indices of plant and leaf traits. Control, absolute control, no fertilizer added. No significant difference among treatments (p > 0.05).
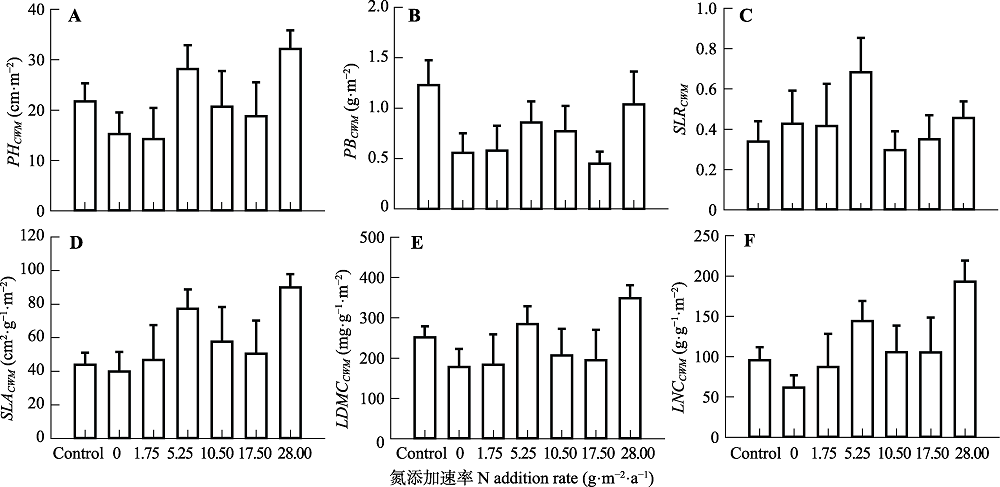
Fig. 3 Effects of N addition on community-weighted mean traits in a typical steppe (mean + SE). Community-weighted mean traits included community-weighted plant height (PHCWM), individual biomass (PBCWM), stem:leaf biomass ratio (SLRCWM), specific leaf area (SLACWM), leaf dry mass content (LDMCCWM), and leaf N content (LNCCWM). Control, absolute control, no fertilizer added. No significant difference among treatments (p > 0.05).
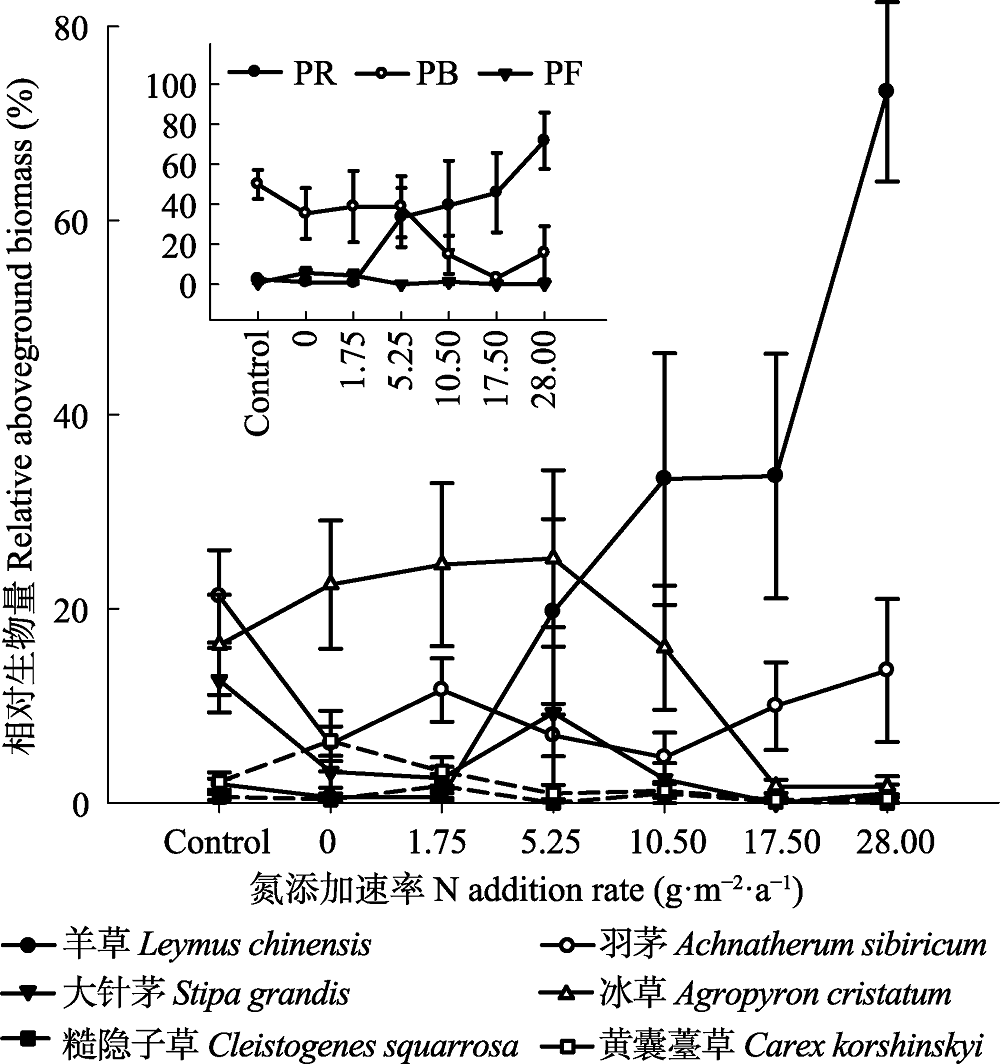
Fig. 4 Effects of N addition on the relative aboveground biomass (RAB) of six dominant species in a typical steppe (mean ± SE). PR, perennial rhizome grasses; PB, perennial bunch grasses; PF, perennial forbs. Control, absolute control, no fertilizer added.
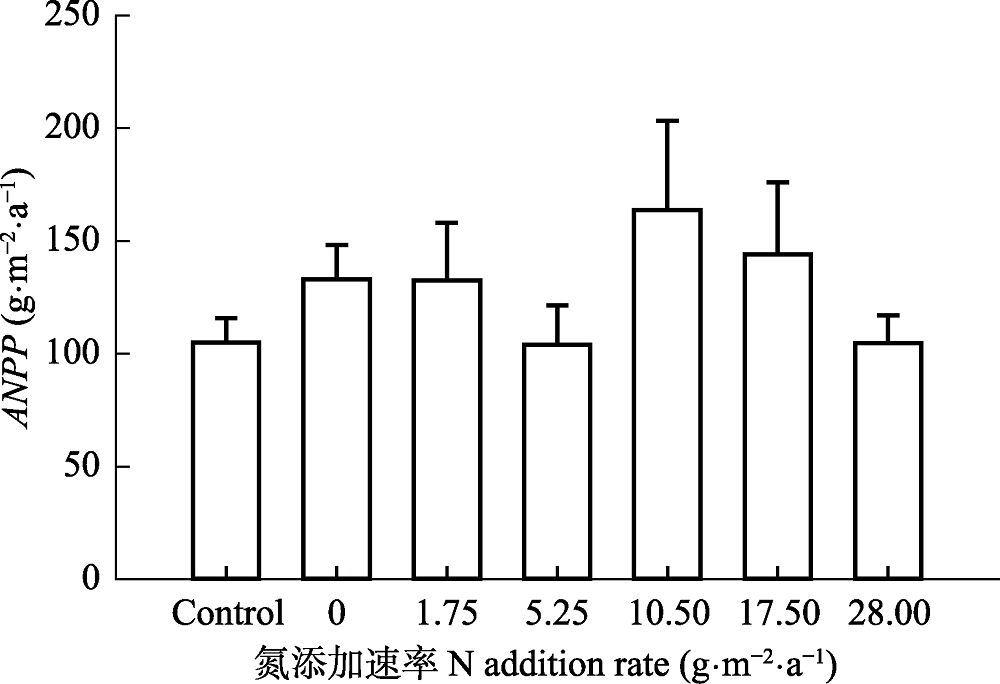
Fig. 5 Effect of N addition on aboveground net primary productivity (ANPP) in a typical steppe (mean + SE). Control, absolute control, no fertilizer added. No significant difference among treatments (p > 0.05).
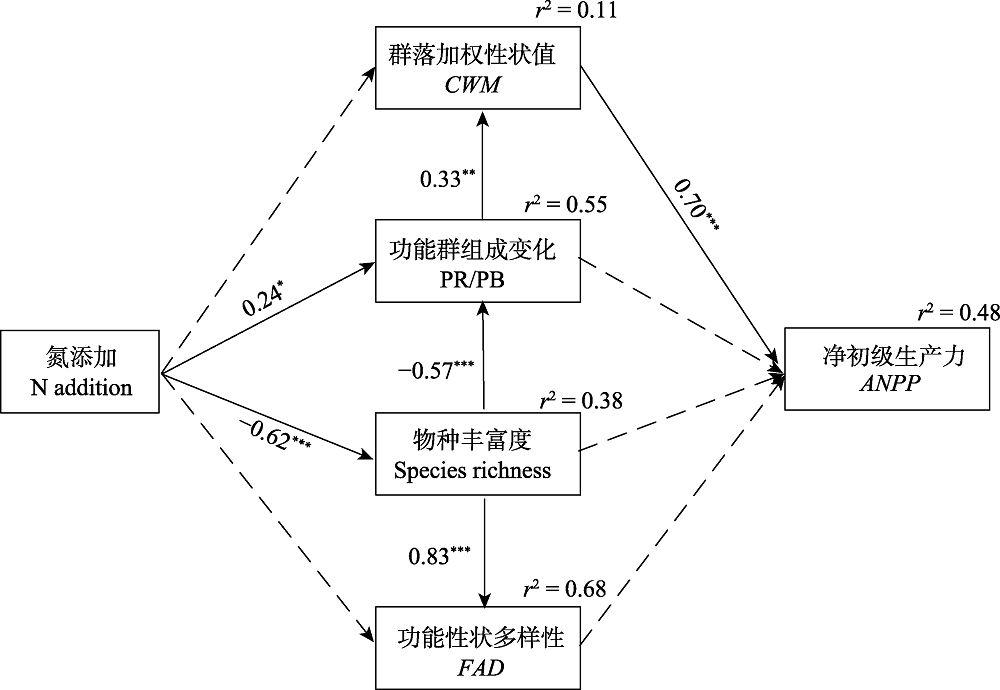
Fig. 6 Structural equation model (SEM) analyses of pathways of N addition on aboveground net primary productivity (ANPP). Results of SEM fitting: c2 = 3.611, p = 0.89, df = 8, n = 13, RMSEA < 0.001, AIC = 29.61, CFI = 1.000. ANPP, aboveground net primary productivity; CWM, community-weighted mean traits; FAD, functional attribute diversity; PR/PB, biomass ratio of perennial rhizome grass to perennial bunchgrass, indicating a shift in functional group composition. Solid and dashed arrows indicate significant (p < 0.05) and non-significant (p > 0.05) effects, respectively. Values associated with solid arrows represent standardized path coefficients (β); *, p < 0.05; **, p < 0.01; ***, p < 0.001. r2 values associated with response variables indicate the proportion of variation explained by relationships with other variables.
| [1] |
Bai YF, Wu JG, Clark CM, Naeem S, Pan QM, Huang JH, Zhang LX, Han XG (2010). Tradeoffs and thresholds in the effects of nitrogen addition on biodiversity and ecosystem functioning: Evidence from Inner Mongolia grasslands. Global Change Biology, 16, 358-372.
DOI URL |
| [2] | Bennett JA, Riibak K, Tamme R, Lewis RJ, Pärtel M (2016). The reciprocal relationship between competition and intraspecific trait variation. Journal of Ecology, 104, 1410-1420. |
| [3] | Bernhardt-Römermann M, Römermann C, Pillar VD, Kudernatsch T, Fischer A (2010). High functional diversity is related to high nitrogen availability in a deciduous forest—Evidence from a functional trait approach. Folia Geobotanica, 45, 111-124. |
| [4] |
Brym ZT, Lake JK, Allen D, Ostling A (2011). Plant functional traits suggest novel ecological strategy for an invasive shrub in an understorey woody plant community. Journal of Applied Ecology, 48, 1098-1106.
DOI URL |
| [5] | Cadotte MW (2017). Functional traits explain ecosystem function through opposing mechanisms. Ecology Letters, 20, 989-996. |
| [6] | Cardinale BJ, Wright JP, Cadotte MW, Carroll IT, Hector A, Srivastava DS, Loreau M, Weis JJ (2007). Impacts of plant diversity on biomass production increase through time because of species complementarity. Proceedings of the National Academy of Sciences of the United States of America, 104, 18123-18128. |
| [7] | Chen DM, Xing W, Lan ZC, Saleem M, Wu Y, Hu SJ, Bai YF (2019). Direct and indirect effects of nitrogen enrichment on soil organisms and carbon and nitrogen mineralization in a semi-arid grassland. Functional Ecology, 33, 175-187. |
| [8] | Clark CM, Tilman D (2008). Loss of plant species after chronic low-level nitrogen deposition to prairie grasslands. Nature, 451, 712-715. |
| [9] | de Bello F, Lepš J, Sebastià MT (2006). Variations in species and functional plant diversity along climatic and grazing gradients. Ecography, 29, 801-810. |
| [10] | Duprè C, Stevens CJ, Ranke T, Bleeker A, Peppler-Lisbach C, Gowing DJG, Dise NB, Dorland E, Bobbink R, Diekmann M (2010). Changes in species richness and composition in European acidic grasslands over the past 70 years: The contribution of cumulative atmospheric nitrogen deposition. Global Change Biology, 16, 344-357. |
| [11] | Finegan B, Peña-Claros M, de Oliveira A, Ascarrunz N, Bret-Harte MS, Carreño-Rocabado G, Casanoves F, Díaz S, Eguiguren Velepucha P, Fernandez F, Licona JC, Lorenzo L, Salgado Negret B, Vaz M, Poorter L (2015). Does functional trait diversity predict above-ground biomass and productivity of tropical forests? Testing three alternative hypotheses. Journal of Ecology, 103, 191-201. |
| [12] | Gagic V, Bartomeus I, Jonsson T, Taylor A, Winqvist C, Fischer C, Slade EM, Steffan-Dewenter I, Emmerson M, Potts SG, Tscharntke T, Weisser W, Bommarco R (2015). Functional identity and diversity of animals predict ecosystem functioning better than species-based indices. Proceedings of the Royal Society B, 282, 20142620. DOI: 10.1098/rspb.2014.2620. |
| [13] | Grime JP (1998). Benefits of plant diversity to ecosystems: Immediate, filter and founder effects. Journal of Ecology, 86, 902-910. |
| [14] | Gross N, Kunstler G, Liancourt P, de Bello F, Suding KN, Lavorel S (2009). Linking individual response to biotic interactions with community structure: A trait-based framework. Functional Ecology, 23, 1167-1178. |
| [15] | Hooper DU, Chapin III FS, Ewel JJ, Hector A, Inchausti P, Lavorel S, Lawton JH, Lodge DM, Loreau M, Naeem S, Schmid B, Setälä H, Symstad AJ, Vandermeer J, Wardle DA (2005). Effects of biodiversity on ecosystem functioning: A consensus of current knowledge. Ecological Monographs, 75, 3-35. |
| [16] | Hooper DU, Vitousek PM (1998). Effects of plant composition and diversity on nutrient cycling. Ecological Monographs, 68, 121-149. |
| [17] | Kardol P, Wardle DA (2010). How understanding aboveground-belowground linkages can assist restoration ecology. Trends in Ecology & Evolution, 25, 670-679. |
| [18] | Knops JM, Reinhart K (2000). Specific leaf area along a nitrogen fertilization gradient. The American Midland Naturalist, 144, 265-272. |
| [19] | Kraft NJB, Godoy O, Levine JM (2015). Plant functional traits and the multidimensional nature of species coexistence. Proceedings of the National Academy of Sciences of the United States of America, 112, 797-802. |
| [20] | Kunstler G, Falster D, Coomes DA, Hui F, Kooyman RM, Laughlin DC, Poorter L, Vanderwel M, Vieilledent G, Wright SJ, Aiba M, Baraloto C, Caspersen J, Cornelissen JHC, Gourlet-Fleury S, Hanewinkel M, Herault B, Kattge J, Kurokawa H, Onoda Y, Peñuelas J, Poorter H, Uriarte M, Richardson S, Ruiz-Benito P, Sun IF, Ståhl G, Swenson NG, Thompson J, Westerlund B, Wirth C, Zavala MA, Zeng HC, Zimmerman JK, Zimmermann NE, Westoby M (2016). Plant functional traits have globally consistent effects on competition. Nature, 529, 204-207. |
| [21] | Lan ZC, Bai YF (2012). Testing mechanisms of N-enrichment-induced species loss in a semiarid Inner Mongolia grassland: Critical thresholds and implications for long-term ecosystem responses. Philosophical Transactions of the Royal Society B-Biological Sciences, 367, 3125-3134. |
| [22] | Lan ZC, Jenerette GD, Zhan SX, Li WH, Zheng SX, Bai YF (2015). Testing the scaling effects and mechanisms of N-induced biodiversity loss: Evidence from a decade-long grassland experiment. Journal of Ecology, 103, 750-760. |
| [23] | Laughlin DC (2014). Applying trait-based models to achieve functional targets for theory-driven ecological restoration. Ecology Letters, 17, 771-784. |
| [24] | Lavorel S, Grigulis K, McIntyre S, Williams NSG, Garden D, Dorrough J, Berman S, Quétier F, Thébault A, Bonis A (2008). Assessing functional diversity in the field—Methodology matters! Functional Ecology, 22, 134-147. |
| [25] | Li W, Cheng JM, Yu KL, Epstein HE, Guo L, Jing GH, Zhao J, Du GZ (2015). Plant functional diversity can be independent of species diversity: Observations based on the impact of 4-yrs of nitrogen and phosphorus additions in an alpine meadow. PLOS ONE, 10, e0136040. DOI: 10.1371/journal.pone.0136040. |
| [26] | Li XG, Zhu ZH, Zhou XS, Yuan FR, Fan RJ, Xu ML (2011). Effects of clipping, fertilizing and watering on the relationship between species diversity, functional diversity and primary productivity in alpine meadow of China. Chinese Journal of Plant Ecology, 35, 1136-1147. |
| [ 李晓刚, 朱志红, 周晓松, 袁芙蓉, 樊瑞俭, 许曼丽 (2011). 刈割、施肥和浇水对高寒草甸物种多样性、功能多样性与初级生产力关系的影响. 植物生态学报, 35, 1136-1147.] | |
| [27] | Maire V, Gross N, Börger L, Proulx R, Wirth C, da Silveira Pontes L, Soussana JF, Louault F (2012). Habitat filtering and niche differentiation jointly explain species relative abundance within grassland communities along fertility and disturbance gradients. New Phytologist, 196, 497-509. |
| [28] | Mokany K, Ash J, Roxburgh S (2008). Functional identity is more important than diversity in influencing ecosystem processes in a temperate native grassland. Journal of Ecology, 96, 884-893. |
| [29] | Mouchet MA, Villéger S, Mason NWH, Mouillot D (2010). Functional diversity measures: An overview of their redundancy and their ability to discriminate community assembly rules. Functional Ecology, 24, 867-876. |
| [30] |
Naeem S, Wright JP (2003). Disentangling biodiversity effects on ecosystem functioning: Deriving solutions to a seemingly insurmountable problem. Ecology Letters, 6, 567-579.
DOI URL |
| [31] | Orwin KH, Buckland SM, Johnson D, Turner BL, Smart S, Oakley S, Bardgett RD (2010). Linkages of plant traits to soil properties and the functioning of temperate grassland. Journal of Ecology, 98, 1074-1083. |
| [32] | Pan QM, Bai YF, Wu JG, Han XG (2011). Hierarchical plant responses and diversity loss after nitrogen addition: Testing three functionally-based hypotheses in the Inner Mongolia grassland. PLOS ONE, 6, e20078. DOI: 10.1371/journal.pone.0020078. |
| [33] | Pardo LH, Fenn ME, Goodale CL, Geiser LH, Driscoll CT, Allen EB, Baron JS, Bobbink R, Bowman WD, Clark CM, Emmett B, Gilliam FS, Greaver TL, Hall SJ, Lilleskov EA, Liu LL, Lynch JA, Nadelhoffer KJ, Perakis SS, Robin-Abbott MJ, Stoddard JL, Weathers KC, Dennis RL (2011). Effects of nitrogen deposition and empirical nitrogen critical loads for ecoregions of the United States. Ecological Applications, 21, 3049-3082. |
| [34] | Payne RJ, Dise NB, Field CD, Dore AJ, Caporn SJ, Stevens CJ (2017). Nitrogen deposition and plant biodiversity: Past, present, and future. Frontiers in Ecology and the Environment, 15, 431-436. |
| [35] | Pérez-Ramos IM, Díaz-Delgado R, de la Riva EG, Villar R, Lloret F, Marañón T (2017). Climate variability and community stability in Mediterranean shrublands: The role of functional diversity and soil environment. Journal of Ecology, 105, 1335-1346. |
| [36] | Petchey OL, Gaston KJ (2002). Functional diversity (FD), species richness and community composition. Ecology Letters, 5, 402-411. |
| [37] |
Prado-Junior JA, Schiavini I, Vale VS, Arantes CS, van der Sande MT, Lohbeck M, Poorter L (2016). Conservative species drive biomass productivity in tropical dry forests. Journal of Ecology, 104, 817-827.
DOI URL |
| [38] | Spasojevic MJ, Suding KN (2012). Inferring community assembly mechanisms from functional diversity patterns: The importance of multiple assembly processes. Journal of Ecology, 100, 652-661. |
| [39] | Suding KN, Collins SL, Gough L, Clark C, Cleland EE, Gross KL, Milchunas DG, Pennings S (2005). Functional- and abundance-based mechanisms explain diversity loss due to N fertilization. Proceedings of the National Academy of Sciences of the United States of America, 102, 4387-4392. |
| [40] | Tylianakis JM, Didham RK, Bascompte J, Wardle DA (2008). Global change and species interactions in terrestrial ecosystems. Ecology Letters, 11, 1351-1363. |
| [41] | Valencia E, Maestre FT, Le Bagousse-Pinguet Y, Quero JL, Tamme R, Börger L, García-Gómez M, Gross N (2015). Functional diversity enhances the resistance of ecosystem multifunctionality to aridity in Mediterranean drylands. New Phytologist, 206, 660-671. |
| [42] |
Violle C, Navas ML, Vile D, Kazakou E, Fortunel C, Hummel I, Garnier E (2007). Let the concept of trait be functional! Oikos, 116, 882-892.
DOI URL |
| [43] | Walker B, Kinzig A, Langridge J (1999). Original articles: Plant attribute diversity, resilience, and ecosystem function: The nature and significance of dominant and minor species. Ecosystems, 2, 95-113. |
| [44] | Wan HW, Yang Y, Bai SQ, Xu YH, Bai YF (2008). Variations in leaf functional traits of six species along a nitrogen addition gradient in Leymus chinensis steppe in Inner Mongolia. Journal of Plant Ecology (Chinese version), 32, 611-621. |
| [ 万宏伟, 杨阳, 白世勤, 徐云虎, 白永飞 (2008). 羊草草原群落6种植物叶片功能特性对氮素添加的响应. 植物生态学报, 32, 611-621.] | |
| [45] | Xu ZW, Li MH, Zimmermann NE, Li SP, Li H, Ren HY, Sun H, Han XG, Jiang Y, Jiang L (2018). Plant functional diversity modulates global environmental change effects on grassland productivity. Journal of Ecology, 106, 1941-1951. |
| [46] | Yang HJ, Li Y, Wu MY, Zhang Z, Li LH, Wan SQ (2011). Plant community responses to nitrogen addition and increased precipitation: The importance of water availability and species traits. Global Change Biology, 17, 2936-2944. |
| [1] | Qing He, Xudong Yuan, Boshen Ren, Zhiyang Feng, Mengzhen Lu, Qiaoling Lin, Qinghu Jiang, Linsen Yang, Huiliang Yu, Hui Yao, Jingyuan Yang, Feng Liu, Mingxi Jiang. Effects of Erigeron annuus invasion on plant community structure and diversity in subalpine peat wetlands [J]. Chin J Plant Ecol, 2026, 50(预发表): 0-. |
| [2] | ZHANG An-Ning, XIAO Ya-Ning, ZHAO Xia, ZHANG Miao, CUI Han Wen, Chen Shu-Yan, AN Li-Zhe. Interactions between shrub encroachment and nitrogen addition on nematode community and functional traits on Qinghai–Tibetan Plateau [J]. Chin J Plant Ecol, 2026, 50(预发表): 0-. |
| [3] | XI Nian-Xun. The impact of multiple global change factors on traits of mycorrhizal plants [J]. , 2026, 50(预发表): 0-. |
| [4] | MA Jian-Hui, TONG Xin, ZHANG Si-Rong, MAO Zi-Kun, QIN Jun, MA Ke-Ping. Research advances and perspectives on physiological and ecological functions of mycorrhizal fungi [J]. Chin J Plant Ecol, 2026, 50(3): 498-514. |
| [5] | ZHENG Zi-Yi, CHEN Jiang-Hui, LIU Hui-Ying. Climate warming increases root exudation rates of dominant species in alpine meadow on the Qingzang Plateau [J]. Chin J Plant Ecol, 2025, 49(9): 1363-1373. |
| [6] | CUI Dong-Qing, TIAN Chen, SONG Hui-Min, LU Xiao-Ming, SA Qi-Ri, XU Guo-Qing, YANG Pei-Zhi, BAI Yong-Fei, TIAN Jian-Qing. Response mechanisms of rhizosphere bacterial community diversity and functional group composition of dominant plants in typical grasslands to long-term grazing [J]. Chin J Plant Ecol, 2025, 49(7): 1163-1176. |
| [7] | HUI Cheng-Yang, ZHANG Qiao-Yi, LIU Teng-Teng, LIU Wei-Yong, ZHOU Li-Na, JIN Xin-Jie, ZHANG Yong-Hua, LIU Jin-Liang. Main vegetation types and species composition of Daluo Mountain, Wenzhou, Zhejiang, China [J]. Chin J Plant Ecol, 2025, 49(6): 990-998. |
| [8] | MA Fu-Long, WANG Yu-Qing, HAO Yu, DUAN Ji-Chao, LIU Fei-Fei, XI Lin-Qiao, HAN Lu. Effects of altitude gradient on plant and soil microbial community structure and diversity in the middle part of the northern slope of the Kunlun Mountains, China [J]. Chin J Plant Ecol, 2025, 49(5): 732-747. |
| [9] | DU Ying-Jie, FAN Ai-Lian, WANG Xue, YAN Xiao-Jun, CHEN Ting-Ting, JIA Lin-Qiao, JIANG Qi, CHEN Guang-Shui. Coordination and differences in root-leaf functional traits between tree species and understory shrub species in a subtropical natural evergreen broadleaf forest [J]. Chin J Plant Ecol, 2025, 49(4): 585-595. |
| [10] | WU Yan-Ning, HAO Min-Hui, HE Huai-Jiang, ZHANG Chun-Yu, ZHAO Xiu-Hai. Relationships between functional diversity and aboveground carbon sink functions and their changes with forest succession in Changbai Mountains, China [J]. Chin J Plant Ecol, 2025, 49(2): 232-243. |
| [11] | BIAN Jia-Chen, WANG Rui, GAO Yang-Yang, LIANG Wen-Jun, JIN Hong, ZHANG Wen-Xuan, ZHANG Xiao-Rong, HAO Jie, $\boxed{\hbox{WANG Chang-Hui}}$, DONG Kuan-Hu, DIAO Hua-Jie. Effect of different levels of nitrogen addition on aboveground and belowground net primary productivity in Leymus secalinus grassland in Northern Shanxi, China [J]. Chin J Plant Ecol, 2025, 49(12): 2043-2053. |
| [12] | QIU Dan-Ni, PENG Qing-Qing, ZHANG Hui-Ling, WEN Hui-Hui, WU Fu-Zhong. Seasonal effects of typical canopy tree species on ant community dynamics in mid-subtropical evergreen broadleaf forests [J]. Chin J Plant Ecol, 2025, 49(11): 1805-1816. |
| [13] | GUO Huan-Min, SHEN Xiao-Xue, LI Rui-Li. Species co-occurrence and distribution probability in Futian Mangrove Nature Reserve of Shenzhen Bay [J]. Chin J Plant Ecol, 2025, 49(11): 1833-1843. |
| [14] | ZHANG Xiao-Ting, WANG Jun-Jie. Chlorophyll fluorescence characteristics of mangrove plants under salt and copper treatments and their relationship with leaf structure and biochemical components [J]. Chin J Plant Ecol, 2025, 49(11): 1944-1956. |
| [15] | TONG Jin-Lian, ZHANG Bo-Na, TANG Lu-Yao, YE Lin-Feng, LI Shu-Wen, XIE Jiang-Bo, LI Yan, WANG Zhong-Yuan. Regional differentiation of functional trait network of C4 plants Setaria viridis along precipitation gradient [J]. Chin J Plant Ecol, 2025, 49(11): 1817-1832. |
| Viewed | ||||||
|
Full text |
|
|||||
|
Abstract |
|
|||||
Copyright © 2026 Chinese Journal of Plant Ecology
Tel: 010-62836134, 62836138, E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
![]()