The sedimentary carbon-sulfur-iron interplay—A lesson from East Anglian salt marsh sediments
1
2019
... (3)盐沼湿地土壤碳库的稳定性及其横向碳流动.盐沼湿地蓝碳碳汇功能主要体现在垂直方向上植物的碳固定和沉积物埋藏, 以及水平方向上土壤有机碳和无机碳在潮汐作用下与近海水体的交换.周期性潮汐水动力过程及由其引起的干湿交替对盐沼湿地横向方向上碳交换的影响还相对薄弱(韩广轩, 2017).同时, 当前关于盐沼湿地土壤有机碳分解转化和碳库组分化学结构转化与土壤微生物的耦合关系尚不明确, 需要深入认识土壤碳库的微生物相关过程(Antler et al., 2019).同时, 当前关于盐沼湿地土壤有机碳分解转化和碳库组分化学结构转化与土壤微生物的耦合关系尚不明确, 需要深入认识土壤碳库的微生物相关过程.目前大部分研究主要关注单一的碳代谢过程, 综合考虑多群落多生态位的碳循环耦联以及碳与其他元素循环的协同作用对盐沼湿地碳储存的影响是目前研究的重点和难点.另外, 多群落环境生物地球化学过程复杂, 目前技术手段揭示的生态过程与生态位真实内在微环境下微生物的功能还存在一定差距甚至出现悖论, 因此亟需更加准确可靠的表征手段, 比如多通道实时检测设备, 以全面正确认识盐沼湿地土壤微生物参与的碳代谢过程. ...
中国滨海盐沼湿地碳收支与碳循环过程研究进展
1
2013
... 全球滨海湿地面积约为20.3 × 104 km2, 我国滨海湿地面积约占全球滨海湿地总面积的1/4, 其中滨海盐沼湿地是我国最主要的滨海湿地类型(曹磊等, 2013).据估算, 我国蓝碳生态系统的碳年埋藏量为0.349-0.835 Tg, 其中盐沼湿地约占80%, 远高于红树林和海草床, 是我国蓝碳碳汇的主要贡献者(周晨昊等, 2016).模型模拟结果表明, 未来气候变暖和海平面上升可能使得盐沼湿地能够更迅速捕获和埋藏大气中的碳(Kirwan & Mudd, 2012).另外, 加强全球滨海湿地的保护和修复, 每年能够吸收、储存和减少0.32-0.89 Tg CO2排放(Hoegh-Guldberg et al., 2019).2010年, 保护国际基金会、世界自然保护联盟、联合国教科文组织政府间海洋学委员会共同发起“蓝碳倡议” (The Blue Carbon Initiative), 旨在支撑全球蓝碳的科学研究、项目实施和政策制定等, 通过恢复和保护受损蓝碳生态系统工作而产生碳效益, 减缓气候变化. ...
中国滨海盐沼湿地碳收支与碳循环过程研究进展
1
2013
... 全球滨海湿地面积约为20.3 × 104 km2, 我国滨海湿地面积约占全球滨海湿地总面积的1/4, 其中滨海盐沼湿地是我国最主要的滨海湿地类型(曹磊等, 2013).据估算, 我国蓝碳生态系统的碳年埋藏量为0.349-0.835 Tg, 其中盐沼湿地约占80%, 远高于红树林和海草床, 是我国蓝碳碳汇的主要贡献者(周晨昊等, 2016).模型模拟结果表明, 未来气候变暖和海平面上升可能使得盐沼湿地能够更迅速捕获和埋藏大气中的碳(Kirwan & Mudd, 2012).另外, 加强全球滨海湿地的保护和修复, 每年能够吸收、储存和减少0.32-0.89 Tg CO2排放(Hoegh-Guldberg et al., 2019).2010年, 保护国际基金会、世界自然保护联盟、联合国教科文组织政府间海洋学委员会共同发起“蓝碳倡议” (The Blue Carbon Initiative), 旨在支撑全球蓝碳的科学研究、项目实施和政策制定等, 通过恢复和保护受损蓝碳生态系统工作而产生碳效益, 减缓气候变化. ...
Dynamics of carbon sequestration in a coastal wetland using radiocarbon measurements
1
2004
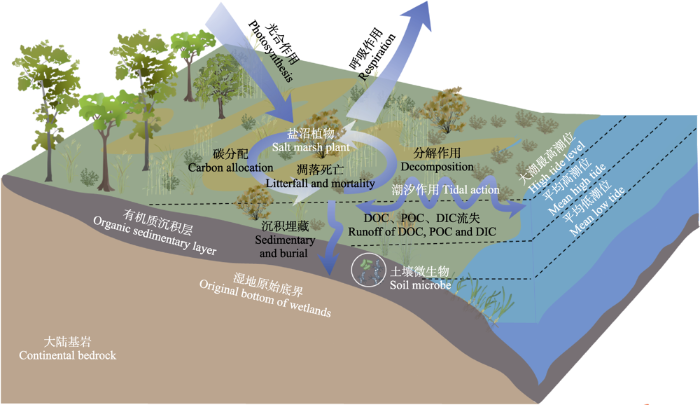
... 盐沼湿地的蓝碳功能是光合碳吸收、碳沉积埋藏、碳输出等多过程相互作用和平衡的结果(图1), 其关键过程主要包括: (1)植被光合固碳.盐沼湿地通常具有高的净初级生产力, 尤其在河口区, 植被净初级生产力可达1 745 g C·m-2·a-1 (Sousa et al., 2010).(2)光合碳分配.植物通过调节碳在不同部位的分配模式来满足自身生长与繁殖的需求或者应对周边环境变化(Wang et al., 2019a).光合固定的碳向根系和土壤碳库的分配和转移规律, 直接影响着土壤碳库的走向(Xu et al., 2020).盐沼植被根冠比可达1.4-50, 有大量的初级生产力所固定的碳被储存在地下生物量中, 通过根系周转进入土壤碳库(王秀君等, 2016).(3)碳沉积埋藏.由于滨海盐沼湿地不断向下沉积, 土壤碳库很难达到饱和, 储存的碳可以在土壤中保存数千年(Radabaugh et al., 2018).因此, 盐沼湿地的碳封存效率远高于森林或其他陆地生态系统(Wang et al., 2021a).(4)土壤碳流失.潮汐作用导致盐沼湿地间歇性厌氧好氧交替, 氧化还原过程频繁发生, 陆地和海洋微生物交互作用; 同时, 周期性潮汐携带大量的SO42-阻碍CH4产生, 从而降低盐沼湿地碳排放量(Choi & Wang, 2004).另外, 潮汐作用下盐沼湿地可溶性有机碳(DOC)、可溶性无机碳(DIC)、颗粒有机碳(POC)交换及其输出也是其碳流失中不容忽视的组成部分(韩广轩, 2017). ...
Dual effect of precipitation redistribution on net ecosystem CO2 exchange of a coastal wetland in the Yellow River Delta
1
2018
... (1)周期性潮汐作用下, 盐沼湿地植被海陆梯度分布格局对碳吸收能力和碳分配的影响.盐沼植物群落呈明显的带状分布, 主要特点为耐盐耐淹的先锋植物分布在高程较低处, 而偏中生性植物分布在高程较高处.以黄河三角洲滨海湿地为例, 芦苇湿地植被净初级生产力为650-1 590 g C·m-2·a-1, 芦苇植被净生态系统CO2交换量(NEE)范围在-164- -269 g C·m-2·a-1, 盐地碱蓬(Suaeda salsa)湿地NEE则为-8- -85 g C·m-2·a-1 (Han et al., 2015; Chu et al., 2018).除本地植物以外, 我国盐沼湿地普遍受到外来植物互花米草入侵的威胁, 从而进一步影响湿地植被演替及其碳吸收能力. ...
An arctic paradox: response of fluvial Hg inputs and bioavailability to global climate change in an extreme coastal environment
1
2020
... 气候变暖导致全球海平面自20世纪初以来持续上升, 盐沼湿地等蓝碳生态系统受到严重威胁.在全球气候变化的背景下, 全球河口海岸带区域极端气候事件发生频率明显增加(Han et al., 2018; de Lacerda et al., 2020; Newton et al., 2020).气候变化会造成热量、土壤水分与盐度等环境要素的改变, 改变原有盐沼湿地生态系统的组成与结构, 影响盐沼湿地群落的演替方向与进程, 直接改变盐沼湿地的碳循环关键过程(Spencer et al., 2016).有研究表明, 气候变暖条件下盐沼湿地净碳积累取决于矿化沉积物沉积速率与有机质积累速率之间的权衡, 模型预测在21世纪上半叶全球气候变化将导致盐沼湿地碳埋藏速率增加(Kirwan & Mudd, 2012).然而也有研究指出, 气候变暖引起的海平面上升会导致大量的盐沼湿地被海水淹没, 使得气候变化对湿地固碳的促进作用最终消失(DeLaune & White, 2012; Rogers et al., 2019).由此可见, 气候变化对盐沼湿地固碳功能存在着复杂、长期且动态的影响.因此, 探索滨海盐沼湿地固碳功能对气候变化的响应时, 应充分考虑气候变化及其导致后果的多重影响, 并需要开展长期的模拟预测. ...
Will coastal wetlands continue to sequester carbon in response to an increase in global sea level?: a case study of the rapidly subsiding Mississippi river deltaic plain
2
2012
... 气候变暖导致全球海平面自20世纪初以来持续上升, 盐沼湿地等蓝碳生态系统受到严重威胁.在全球气候变化的背景下, 全球河口海岸带区域极端气候事件发生频率明显增加(Han et al., 2018; de Lacerda et al., 2020; Newton et al., 2020).气候变化会造成热量、土壤水分与盐度等环境要素的改变, 改变原有盐沼湿地生态系统的组成与结构, 影响盐沼湿地群落的演替方向与进程, 直接改变盐沼湿地的碳循环关键过程(Spencer et al., 2016).有研究表明, 气候变暖条件下盐沼湿地净碳积累取决于矿化沉积物沉积速率与有机质积累速率之间的权衡, 模型预测在21世纪上半叶全球气候变化将导致盐沼湿地碳埋藏速率增加(Kirwan & Mudd, 2012).然而也有研究指出, 气候变暖引起的海平面上升会导致大量的盐沼湿地被海水淹没, 使得气候变化对湿地固碳的促进作用最终消失(DeLaune & White, 2012; Rogers et al., 2019).由此可见, 气候变化对盐沼湿地固碳功能存在着复杂、长期且动态的影响.因此, 探索滨海盐沼湿地固碳功能对气候变化的响应时, 应充分考虑气候变化及其导致后果的多重影响, 并需要开展长期的模拟预测. ...
... (4)气候变化和海平面上升背景下盐沼湿地蓝碳模拟与增汇潜力评估.气候变化和海平面上升改变了盐沼湿地的生物地球化学过程、水文条件以及植被生长和演替, 导致了盐沼湿地蓝碳潜力存在极大的不确定性.气候变化和海平面上升增加滨海湿地碳汇功能(Langley et al., 2009; Wang et al., 2019b, 2021a), 也会通过海岸侵蚀导致更多的碳流失(DeLaune & White, 2012), 但现有模型大多基于气候驱动因素、沉积和碳累积速率等, 缺乏针对生态系统和植被演替过程的考虑.另外, 增温、富营养化和海平面上升对滨海湿地碳储存的影响机制不同(Spivak et al., 2019), 这些因素的改变在极大程度上影响了滨海盐沼湿地的固碳能力, 但目前很难精确地预测这种未知影响.因此, 量化这些因素如何影响盐沼湿地有机碳固存与分解对于构建更具预测性的蓝碳模型至关重要.同时急需构建不同尺度气候变化、海平面上升、生态系统演替耦合驱动的盐沼湿地碳循环关键过程模拟预测新范式, 增强盐沼湿地蓝碳增汇潜力评估的可靠性. ...
Environmental effects on carbon isotope discrimination from assimilation to respiration in a coniferous and broad-leaved mixed forest of northeast China
1
2020
... 植物光合碳是植物-土壤系统碳循环的起点, 解析植物-土壤系统中光合碳的分配过程及机制是估算盐沼湿地固碳功能的先决条件.植物主要通过光合作用将CO2转化为多糖等光合产物, 由叶片通过枝、干向下运输, 分配到植物体各个组织器官.在生态系统碳循环过程中, 植物会通过调节碳在不同部位的分配来满足自身生长与繁殖的需求或应对周边环境变化, 从而间接影响生物地球化学循环(Wang et al., 2019a).气候条件、土壤理化性质、微生物活性和植物生长发育等均会影响植物对光合碳分配的模式, 改变植物向地上、地下部分的碳输入(Diao et al., 2020).此外, 光合碳固定是碳分配的主要驱动力, 环境胁迫还可以通过影响光合作用影响碳吸收, 从而间接影响光同化物的分配(Pausch & Kuzyakov, 2018).目前, 碳同位素技术是研究植物光合碳分配、周转和固定的重要手段.光合碳在植物体各组织器官内的迁移和转化十分迅速, 碳同化后的第1天从叶片到根系的分配就达到峰值(Pausch & Kuzyakov, 2018).同时, 新合成的光合产物可以通过根系分泌物的形式刺激土壤有机质(老碳)的分解, 即土壤碳输入的激发效应(Street et al., 2020).另外, 光合碳分配在物种间也存在差异.Meta分析发现, 与禾本科植物相比, 莎草科植物可以通过根际沉积将更多的碳转移至根和土壤中, 增加地下碳固存(Mou et al., 2018); 采用稳定碳同位素方法, 发现互花米草(Spartina alterniflora)可以将33%和7%的光合同化碳分配到茎和根, 显著高于芦苇(Phragmites australis)的分配比例(Zhang et al., 2017). ...
Greenhouse gas fluxes from coastal wetlands at the intersection of urban pollution and saltwater intrusion: a soil core experiment
2
2019
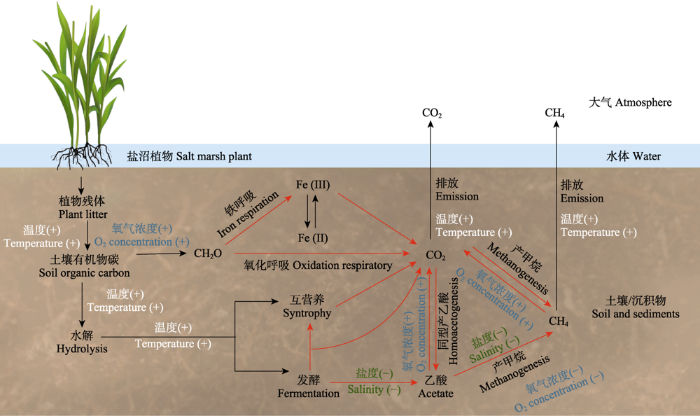
... 目前, 国内外学者已经使用多种技术手段, 如13C或14C标记, 对农田、草地和森林等生态系统的土壤碳矿化过程、行为及参与关键单元开展了研究.但是盐沼湿地有关研究相对滞后, 主要集中在原位或者微宇宙培养下, 生物地球化学参数、基础微生物等指标的测定分析(Doroski et al., 2019).近期研究发现盐沼湿地土壤碳稳定性与土壤碳官能团种类息息相关(Luk et al., 2021).如Roth等(2019)研究指出可溶性有机物质在土壤中持久性保存归因于其在沉积过程中的分子变化.沉积过程中易被利用的烷氧碳比例逐渐降低, 而稳定性较高的烷基碳比例逐渐升高.盐沼湿地处于长期淹水或间歇性淹水, 氧化还原过程频繁发生(图2), 微生物-矿物-有机碳的相互作用对盐沼湿地碳的保存和流失起重要的调控作用.除盐沼土壤内部活跃的生物地球化学过程, 在长期外源物输入条件下, 微生物群落调节着土壤有机碳的种类和含量(Georgiou et al., 2017).外源有机质的多样性将会影响微生物胞外酶的分泌, 进而影响土壤有机质的形成和分解过程(Lehmann et al., 2020).另外, 海水入侵对盐沼湿地微生物参与的土壤碳矿化过程和CO2/CH4排放具有一定的抑制作用(Doroski et al., 2019).但是, 目前关于盐沼湿地土壤有机碳分子结构转换与土壤微生物的耦合关系尚不明确.深入认识微生物相关过程是理解土壤碳库稳定性的基础, 也是认知蓝碳潜力的关键. ...
... ., 2019).但是, 目前关于盐沼湿地土壤有机碳分子结构转换与土壤微生物的耦合关系尚不明确.深入认识微生物相关过程是理解土壤碳库稳定性的基础, 也是认知蓝碳潜力的关键. ...
The role of coastal plant communities for climate change mitigation and adaptation
1
2013
... 2009年, 联合国环境规划署、粮农组织和教科文组织政府间海洋学委员会联合发布《蓝碳: 健康海洋对碳的固定作用》(《Blue Carbon: the Role of Healthy Oceans in Binding Carbon》), 确认了海洋在减缓全球气候变化和碳循环过程中至关重要的作用, 首次提出了“蓝碳”的概念, 并明确指出盐沼湿地、红树林和海草床三类海岸带蓝碳是相对易于管理的(Nellemann et al., 2009).2019年, 联合国政府间气候变化专门委员会(IPCC)发布《气候变化中的海洋与冰冻圈特别报告》(《Special Report on the Ocean and Cryosphere in a Changing Climate》), 明确指出是否“易于管理”是界定蓝碳的必要条件.滨海盐沼湿地(coastal salt marshes)是指海岸沿线受海洋潮汐周期性或间歇性影响, 有盐生植物覆盖的咸水或淡咸水淤泥质滩涂, 是地球上高生产力植被类型之一.盐沼湿地具有很高的固碳能力, 同时土壤能够捕获和储存大量的碳, 其土壤碳埋藏速率为(218 ± 24) g·m-2·a-1, 比森林生态系统高40倍左右(Mcleod et al., 2011; Macreadie et al., 2019).据估计, 盐沼湿地上层(约1 m)土壤含有约917 t CO2 e·hm-2 (每hm2 CO2当量), 年固碳量约为(8.0 ± 8.5) t CO2 e·hm-2 (Sapkota & White, 2021).因此, 滨海盐沼湿地与红树林、海草床等海岸带生态系统被称为蓝碳生态系统.虽然海岸带蓝碳生态系统只占全球海洋面积的0.2%, 但贡献了海洋沉积物碳总储量的50% (Duarte et al., 2013). ...
Ecosystem engineering by large grazers enhances carbon stocks in a tidal salt marsh
1
2015
... 在全球范围内, 已开展了气候变暖、海平面上升、泥沙沉积等因素对盐沼湿地及其碳循环过程影响的模拟预测研究(Spencer et al., 2016; Schuerch et al., 2018; Ewers Lewis et al., 2020).尽管这些研究横跨了多个时空尺度, 并解释了盐沼湿地及其碳循环是如何受未来全球变化的影响, 但大量国内外学者在开展盐沼湿地固碳动态模拟时均忽略了由于自身演替而导致的湿地植被群落景观格局的变化, 及其带来的景观-区域尺度盐沼湿地固碳功能的改变(Elschot et al., 2015; Gu et al., 2018).这种影响在快速演替的年轻河口盐沼湿地中表现得更为明显.因此, 想要科学地认识盐沼湿地对气候变化影响的响应, 准确地评估滨海盐沼湿地的固碳功能在未来减缓气候变化中所发挥的作用, 除了需要深入地了解全球变化对盐沼植被碳吸收、碳分配、碳埋藏等过程的影响外, 还需要全面地把握全球变化带来的环境改变而引起的盐沼植被群落空间分布及其演替动态对整个生态系统碳循环过程的影响.在此基础之上, 耦合植被群落演替与碳循环两类模型是实现生态系统-景观-区域水平的碳循环过程的模拟与碳汇预测的有效途径.因此, 亟待构建区域尺度气候变化、海平面上升、生态系统演替耦合驱动的盐沼湿地碳循环关键过程模拟预测新范式, 增强盐沼湿地蓝碳增汇潜力评估的可靠性. ...
Drivers and modelling of blue carbon stock variability in sediments of southeastern Australia
1
2020
... 在全球范围内, 已开展了气候变暖、海平面上升、泥沙沉积等因素对盐沼湿地及其碳循环过程影响的模拟预测研究(Spencer et al., 2016; Schuerch et al., 2018; Ewers Lewis et al., 2020).尽管这些研究横跨了多个时空尺度, 并解释了盐沼湿地及其碳循环是如何受未来全球变化的影响, 但大量国内外学者在开展盐沼湿地固碳动态模拟时均忽略了由于自身演替而导致的湿地植被群落景观格局的变化, 及其带来的景观-区域尺度盐沼湿地固碳功能的改变(Elschot et al., 2015; Gu et al., 2018).这种影响在快速演替的年轻河口盐沼湿地中表现得更为明显.因此, 想要科学地认识盐沼湿地对气候变化影响的响应, 准确地评估滨海盐沼湿地的固碳功能在未来减缓气候变化中所发挥的作用, 除了需要深入地了解全球变化对盐沼植被碳吸收、碳分配、碳埋藏等过程的影响外, 还需要全面地把握全球变化带来的环境改变而引起的盐沼植被群落空间分布及其演替动态对整个生态系统碳循环过程的影响.在此基础之上, 耦合植被群落演替与碳循环两类模型是实现生态系统-景观-区域水平的碳循环过程的模拟与碳汇预测的有效途径.因此, 亟待构建区域尺度气候变化、海平面上升、生态系统演替耦合驱动的盐沼湿地碳循环关键过程模拟预测新范式, 增强盐沼湿地蓝碳增汇潜力评估的可靠性. ...
生物标志物及其在生态系统研究中的应用
3
2020
... 解析滨海湿地埋藏碳的来源所使用的方法主要是碳氮比值法(C:N)、稳定同位素组成法和生物标记物法(Xia et al., 2021a).综合利用碳、氮稳定性同位素丰度(δ)可以解析湿地沉积物有机质的来源(Xia et al., 2021b).通过对沉积物C:N的测定可以区分其碳的来源是水生植被还是陆生植被, 水生植物C:N一般在4-10之间, 而陆生维管植物和挺水植物C:N一般大于20.同时, 沉积物有机质中δ13C和δ15N与有机质来源紧密相关, 通过对滨海湿地地上植被和周围水体主要植物体δ13C的测定可以区分其沉积物中内源有机碳的来源(黄梅等, 2018).除了δ13C, δ15N也被经常用于解析有机质的来源.典型陆地植物的δ15N变化范围为0-11‰, 海草δ15N变化范围为6‰-12‰, 而海洋中藻类的δ15N变化范围为0-4‰ (Samper- Villarreal et al., 2016).另外, 生物标志物是环境和地质体中记载着原始生物母质分子结构信息的有机化合物, 被广泛用于研究埋藏碳的来源和转化特征(冯晓娟等, 2020).例如, 正构烷烃广泛分布于低等细菌藻类与高等植物等不同类型的生物体的蜡质脂类中, 来源于不同生物体的正构烷烃, 其碳数组成与分布特征不同(冯晓娟等, 2020), 可以用来探究沉积物有机质内源菌藻、水生植物、草本植物与木本植物的相对贡献.氨基糖在土壤中更稳定而且保存更长时间(Joergensen, 2018), 是土壤中微生物残体碳的主要标记物, 被广泛应用于研究微生物残体对土壤有机碳和各种微环境流通量的贡献(Joergensen, 2018).综合运用C:N法、稳定同位素组成法和生物标记物法可以有效地解析湿地沉积物中有机碳的来源及其转化过程, 是当前湿地碳循环研究的热点方向(冯晓娟等, 2020; Xia et al., 2021b; Yuan et al., 2021). ...
... ).例如, 正构烷烃广泛分布于低等细菌藻类与高等植物等不同类型的生物体的蜡质脂类中, 来源于不同生物体的正构烷烃, 其碳数组成与分布特征不同(冯晓娟等, 2020), 可以用来探究沉积物有机质内源菌藻、水生植物、草本植物与木本植物的相对贡献.氨基糖在土壤中更稳定而且保存更长时间(Joergensen, 2018), 是土壤中微生物残体碳的主要标记物, 被广泛应用于研究微生物残体对土壤有机碳和各种微环境流通量的贡献(Joergensen, 2018).综合运用C:N法、稳定同位素组成法和生物标记物法可以有效地解析湿地沉积物中有机碳的来源及其转化过程, 是当前湿地碳循环研究的热点方向(冯晓娟等, 2020; Xia et al., 2021b; Yuan et al., 2021). ...
... ).综合运用C:N法、稳定同位素组成法和生物标记物法可以有效地解析湿地沉积物中有机碳的来源及其转化过程, 是当前湿地碳循环研究的热点方向(冯晓娟等, 2020; Xia et al., 2021b; Yuan et al., 2021). ...
生物标志物及其在生态系统研究中的应用
3
2020
... 解析滨海湿地埋藏碳的来源所使用的方法主要是碳氮比值法(C:N)、稳定同位素组成法和生物标记物法(Xia et al., 2021a).综合利用碳、氮稳定性同位素丰度(δ)可以解析湿地沉积物有机质的来源(Xia et al., 2021b).通过对沉积物C:N的测定可以区分其碳的来源是水生植被还是陆生植被, 水生植物C:N一般在4-10之间, 而陆生维管植物和挺水植物C:N一般大于20.同时, 沉积物有机质中δ13C和δ15N与有机质来源紧密相关, 通过对滨海湿地地上植被和周围水体主要植物体δ13C的测定可以区分其沉积物中内源有机碳的来源(黄梅等, 2018).除了δ13C, δ15N也被经常用于解析有机质的来源.典型陆地植物的δ15N变化范围为0-11‰, 海草δ15N变化范围为6‰-12‰, 而海洋中藻类的δ15N变化范围为0-4‰ (Samper- Villarreal et al., 2016).另外, 生物标志物是环境和地质体中记载着原始生物母质分子结构信息的有机化合物, 被广泛用于研究埋藏碳的来源和转化特征(冯晓娟等, 2020).例如, 正构烷烃广泛分布于低等细菌藻类与高等植物等不同类型的生物体的蜡质脂类中, 来源于不同生物体的正构烷烃, 其碳数组成与分布特征不同(冯晓娟等, 2020), 可以用来探究沉积物有机质内源菌藻、水生植物、草本植物与木本植物的相对贡献.氨基糖在土壤中更稳定而且保存更长时间(Joergensen, 2018), 是土壤中微生物残体碳的主要标记物, 被广泛应用于研究微生物残体对土壤有机碳和各种微环境流通量的贡献(Joergensen, 2018).综合运用C:N法、稳定同位素组成法和生物标记物法可以有效地解析湿地沉积物中有机碳的来源及其转化过程, 是当前湿地碳循环研究的热点方向(冯晓娟等, 2020; Xia et al., 2021b; Yuan et al., 2021). ...
... ).例如, 正构烷烃广泛分布于低等细菌藻类与高等植物等不同类型的生物体的蜡质脂类中, 来源于不同生物体的正构烷烃, 其碳数组成与分布特征不同(冯晓娟等, 2020), 可以用来探究沉积物有机质内源菌藻、水生植物、草本植物与木本植物的相对贡献.氨基糖在土壤中更稳定而且保存更长时间(Joergensen, 2018), 是土壤中微生物残体碳的主要标记物, 被广泛应用于研究微生物残体对土壤有机碳和各种微环境流通量的贡献(Joergensen, 2018).综合运用C:N法、稳定同位素组成法和生物标记物法可以有效地解析湿地沉积物中有机碳的来源及其转化过程, 是当前湿地碳循环研究的热点方向(冯晓娟等, 2020; Xia et al., 2021b; Yuan et al., 2021). ...
... ).综合运用C:N法、稳定同位素组成法和生物标记物法可以有效地解析湿地沉积物中有机碳的来源及其转化过程, 是当前湿地碳循环研究的热点方向(冯晓娟等, 2020; Xia et al., 2021b; Yuan et al., 2021). ...
Microbial community-level regulation explains soil carbon responses to long-term litter manipulations
1
2017
... 目前, 国内外学者已经使用多种技术手段, 如13C或14C标记, 对农田、草地和森林等生态系统的土壤碳矿化过程、行为及参与关键单元开展了研究.但是盐沼湿地有关研究相对滞后, 主要集中在原位或者微宇宙培养下, 生物地球化学参数、基础微生物等指标的测定分析(Doroski et al., 2019).近期研究发现盐沼湿地土壤碳稳定性与土壤碳官能团种类息息相关(Luk et al., 2021).如Roth等(2019)研究指出可溶性有机物质在土壤中持久性保存归因于其在沉积过程中的分子变化.沉积过程中易被利用的烷氧碳比例逐渐降低, 而稳定性较高的烷基碳比例逐渐升高.盐沼湿地处于长期淹水或间歇性淹水, 氧化还原过程频繁发生(图2), 微生物-矿物-有机碳的相互作用对盐沼湿地碳的保存和流失起重要的调控作用.除盐沼土壤内部活跃的生物地球化学过程, 在长期外源物输入条件下, 微生物群落调节着土壤有机碳的种类和含量(Georgiou et al., 2017).外源有机质的多样性将会影响微生物胞外酶的分泌, 进而影响土壤有机质的形成和分解过程(Lehmann et al., 2020).另外, 海水入侵对盐沼湿地微生物参与的土壤碳矿化过程和CO2/CH4排放具有一定的抑制作用(Doroski et al., 2019).但是, 目前关于盐沼湿地土壤有机碳分子结构转换与土壤微生物的耦合关系尚不明确.深入认识微生物相关过程是理解土壤碳库稳定性的基础, 也是认知蓝碳潜力的关键. ...
Losses of salt marsh in China: trends, threats and management
1
2018
... 在全球范围内, 已开展了气候变暖、海平面上升、泥沙沉积等因素对盐沼湿地及其碳循环过程影响的模拟预测研究(Spencer et al., 2016; Schuerch et al., 2018; Ewers Lewis et al., 2020).尽管这些研究横跨了多个时空尺度, 并解释了盐沼湿地及其碳循环是如何受未来全球变化的影响, 但大量国内外学者在开展盐沼湿地固碳动态模拟时均忽略了由于自身演替而导致的湿地植被群落景观格局的变化, 及其带来的景观-区域尺度盐沼湿地固碳功能的改变(Elschot et al., 2015; Gu et al., 2018).这种影响在快速演替的年轻河口盐沼湿地中表现得更为明显.因此, 想要科学地认识盐沼湿地对气候变化影响的响应, 准确地评估滨海盐沼湿地的固碳功能在未来减缓气候变化中所发挥的作用, 除了需要深入地了解全球变化对盐沼植被碳吸收、碳分配、碳埋藏等过程的影响外, 还需要全面地把握全球变化带来的环境改变而引起的盐沼植被群落空间分布及其演替动态对整个生态系统碳循环过程的影响.在此基础之上, 耦合植被群落演替与碳循环两类模型是实现生态系统-景观-区域水平的碳循环过程的模拟与碳汇预测的有效途径.因此, 亟待构建区域尺度气候变化、海平面上升、生态系统演替耦合驱动的盐沼湿地碳循环关键过程模拟预测新范式, 增强盐沼湿地蓝碳增汇潜力评估的可靠性. ...
潮汐作用和干湿交替对盐沼湿地碳交换的影响机制研究进展
3
2017
... 由于盐沼湿地处于陆海过渡带, 周期性潮汐作用及其所控制的植被分布与演替过程是控制和维持盐沼湿地碳循环过程和碳汇功能的特异性机制(韩广轩, 2017).受陆海动力交替作用, 盐沼湿地植被时空动态演替复杂而迅速.同时, 盐沼植被类型由海向陆具有明显的梯度分布.陆域向海扩张促使植被向海方向不断演替更迭的同时, 原来靠近海域的地方渐渐深入内地, 海洋环境特征逐渐减弱, 盐生植被向着淡水和旱生植被演替.因而, 盐沼湿地植被的演替不仅表现在空间地域上的植被类型的更替, 也表现在时间序列上的演进(韩广轩等, 2018).同时, 盐沼湿地具有横向扩张和快速淤积的发育特征, 潮汐运动通过沉积物的供给直接影响盐沼湿地土壤碳封存能力, 从而影响盐沼湿地碳汇形成机制. ...
... 盐沼湿地的蓝碳功能是光合碳吸收、碳沉积埋藏、碳输出等多过程相互作用和平衡的结果(图1), 其关键过程主要包括: (1)植被光合固碳.盐沼湿地通常具有高的净初级生产力, 尤其在河口区, 植被净初级生产力可达1 745 g C·m-2·a-1 (Sousa et al., 2010).(2)光合碳分配.植物通过调节碳在不同部位的分配模式来满足自身生长与繁殖的需求或者应对周边环境变化(Wang et al., 2019a).光合固定的碳向根系和土壤碳库的分配和转移规律, 直接影响着土壤碳库的走向(Xu et al., 2020).盐沼植被根冠比可达1.4-50, 有大量的初级生产力所固定的碳被储存在地下生物量中, 通过根系周转进入土壤碳库(王秀君等, 2016).(3)碳沉积埋藏.由于滨海盐沼湿地不断向下沉积, 土壤碳库很难达到饱和, 储存的碳可以在土壤中保存数千年(Radabaugh et al., 2018).因此, 盐沼湿地的碳封存效率远高于森林或其他陆地生态系统(Wang et al., 2021a).(4)土壤碳流失.潮汐作用导致盐沼湿地间歇性厌氧好氧交替, 氧化还原过程频繁发生, 陆地和海洋微生物交互作用; 同时, 周期性潮汐携带大量的SO42-阻碍CH4产生, 从而降低盐沼湿地碳排放量(Choi & Wang, 2004).另外, 潮汐作用下盐沼湿地可溶性有机碳(DOC)、可溶性无机碳(DIC)、颗粒有机碳(POC)交换及其输出也是其碳流失中不容忽视的组成部分(韩广轩, 2017). ...
... (3)盐沼湿地土壤碳库的稳定性及其横向碳流动.盐沼湿地蓝碳碳汇功能主要体现在垂直方向上植物的碳固定和沉积物埋藏, 以及水平方向上土壤有机碳和无机碳在潮汐作用下与近海水体的交换.周期性潮汐水动力过程及由其引起的干湿交替对盐沼湿地横向方向上碳交换的影响还相对薄弱(韩广轩, 2017).同时, 当前关于盐沼湿地土壤有机碳分解转化和碳库组分化学结构转化与土壤微生物的耦合关系尚不明确, 需要深入认识土壤碳库的微生物相关过程(Antler et al., 2019).同时, 当前关于盐沼湿地土壤有机碳分解转化和碳库组分化学结构转化与土壤微生物的耦合关系尚不明确, 需要深入认识土壤碳库的微生物相关过程.目前大部分研究主要关注单一的碳代谢过程, 综合考虑多群落多生态位的碳循环耦联以及碳与其他元素循环的协同作用对盐沼湿地碳储存的影响是目前研究的重点和难点.另外, 多群落环境生物地球化学过程复杂, 目前技术手段揭示的生态过程与生态位真实内在微环境下微生物的功能还存在一定差距甚至出现悖论, 因此亟需更加准确可靠的表征手段, 比如多通道实时检测设备, 以全面正确认识盐沼湿地土壤微生物参与的碳代谢过程. ...
潮汐作用和干湿交替对盐沼湿地碳交换的影响机制研究进展
3
2017
... 由于盐沼湿地处于陆海过渡带, 周期性潮汐作用及其所控制的植被分布与演替过程是控制和维持盐沼湿地碳循环过程和碳汇功能的特异性机制(韩广轩, 2017).受陆海动力交替作用, 盐沼湿地植被时空动态演替复杂而迅速.同时, 盐沼植被类型由海向陆具有明显的梯度分布.陆域向海扩张促使植被向海方向不断演替更迭的同时, 原来靠近海域的地方渐渐深入内地, 海洋环境特征逐渐减弱, 盐生植被向着淡水和旱生植被演替.因而, 盐沼湿地植被的演替不仅表现在空间地域上的植被类型的更替, 也表现在时间序列上的演进(韩广轩等, 2018).同时, 盐沼湿地具有横向扩张和快速淤积的发育特征, 潮汐运动通过沉积物的供给直接影响盐沼湿地土壤碳封存能力, 从而影响盐沼湿地碳汇形成机制. ...
... 盐沼湿地的蓝碳功能是光合碳吸收、碳沉积埋藏、碳输出等多过程相互作用和平衡的结果(图1), 其关键过程主要包括: (1)植被光合固碳.盐沼湿地通常具有高的净初级生产力, 尤其在河口区, 植被净初级生产力可达1 745 g C·m-2·a-1 (Sousa et al., 2010).(2)光合碳分配.植物通过调节碳在不同部位的分配模式来满足自身生长与繁殖的需求或者应对周边环境变化(Wang et al., 2019a).光合固定的碳向根系和土壤碳库的分配和转移规律, 直接影响着土壤碳库的走向(Xu et al., 2020).盐沼植被根冠比可达1.4-50, 有大量的初级生产力所固定的碳被储存在地下生物量中, 通过根系周转进入土壤碳库(王秀君等, 2016).(3)碳沉积埋藏.由于滨海盐沼湿地不断向下沉积, 土壤碳库很难达到饱和, 储存的碳可以在土壤中保存数千年(Radabaugh et al., 2018).因此, 盐沼湿地的碳封存效率远高于森林或其他陆地生态系统(Wang et al., 2021a).(4)土壤碳流失.潮汐作用导致盐沼湿地间歇性厌氧好氧交替, 氧化还原过程频繁发生, 陆地和海洋微生物交互作用; 同时, 周期性潮汐携带大量的SO42-阻碍CH4产生, 从而降低盐沼湿地碳排放量(Choi & Wang, 2004).另外, 潮汐作用下盐沼湿地可溶性有机碳(DOC)、可溶性无机碳(DIC)、颗粒有机碳(POC)交换及其输出也是其碳流失中不容忽视的组成部分(韩广轩, 2017). ...
... (3)盐沼湿地土壤碳库的稳定性及其横向碳流动.盐沼湿地蓝碳碳汇功能主要体现在垂直方向上植物的碳固定和沉积物埋藏, 以及水平方向上土壤有机碳和无机碳在潮汐作用下与近海水体的交换.周期性潮汐水动力过程及由其引起的干湿交替对盐沼湿地横向方向上碳交换的影响还相对薄弱(韩广轩, 2017).同时, 当前关于盐沼湿地土壤有机碳分解转化和碳库组分化学结构转化与土壤微生物的耦合关系尚不明确, 需要深入认识土壤碳库的微生物相关过程(Antler et al., 2019).同时, 当前关于盐沼湿地土壤有机碳分解转化和碳库组分化学结构转化与土壤微生物的耦合关系尚不明确, 需要深入认识土壤碳库的微生物相关过程.目前大部分研究主要关注单一的碳代谢过程, 综合考虑多群落多生态位的碳循环耦联以及碳与其他元素循环的协同作用对盐沼湿地碳储存的影响是目前研究的重点和难点.另外, 多群落环境生物地球化学过程复杂, 目前技术手段揭示的生态过程与生态位真实内在微环境下微生物的功能还存在一定差距甚至出现悖论, 因此亟需更加准确可靠的表征手段, 比如多通道实时检测设备, 以全面正确认识盐沼湿地土壤微生物参与的碳代谢过程. ...
Effects of episodic flooding on the net ecosystem CO2 exchange of a supratidal wetland in the Yellow River Delta
1
2015
... (1)周期性潮汐作用下, 盐沼湿地植被海陆梯度分布格局对碳吸收能力和碳分配的影响.盐沼植物群落呈明显的带状分布, 主要特点为耐盐耐淹的先锋植物分布在高程较低处, 而偏中生性植物分布在高程较高处.以黄河三角洲滨海湿地为例, 芦苇湿地植被净初级生产力为650-1 590 g C·m-2·a-1, 芦苇植被净生态系统CO2交换量(NEE)范围在-164- -269 g C·m-2·a-1, 盐地碱蓬(Suaeda salsa)湿地NEE则为-8- -85 g C·m-2·a-1 (Han et al., 2015; Chu et al., 2018).除本地植物以外, 我国盐沼湿地普遍受到外来植物互花米草入侵的威胁, 从而进一步影响湿地植被演替及其碳吸收能力. ...
Precipitation events reduce soil respiration in a coastal wetland based on four-year continuous field measurements
2
2018
... 植物光合碳分配也是植物采用的一种应对环境胁迫的适应性策略(Stefan et al., 2018; Wang et al., 2021b).例如随着植物可用养分在干旱胁迫环境下变得越来越有限, 植物可能向根部分配更多的光合碳用来吸收水分, 从而引发土壤有机质分解并增加植物养分的可利用性(Williams & de Vries, 2020).同时干旱还可以通过调节根际分泌物的质量和数量等加快或减缓土壤碳循环(Preece et al., 2018).相应地, 滨海湿地生态系统水文过程主要受周期性潮汐作用以及地表淡水与地下咸水之间的相互作用, 导致湿地土壤盐碱化或地表淹水(Han et al., 2018), 也可能改变植被光合碳分配策略.目前, 国内外学者已经使用13C或14C标记对农田、草地和森林等旱地生态系统的光合碳及其后续转化进行了定量分析(Pausch & Kuzyakov, 2018; Wang et al., 2019a).由于盐沼湿地长期或周期性淹水和厌氧, 其植物光合碳分配途径有别于其他生态系统, 因此对旱地生态系统的归纳不一定能够阐明湿地生态系统碳循环的关键过程(Liu et al., 2019).因此, 在全球气候变化背景下, 迫切需要了解盐沼湿地植物-土壤中光合碳分配过程及比例, 这对于理解和精确评估盐沼湿地碳汇功能及构建模型具有重要意义. ...
... 气候变暖导致全球海平面自20世纪初以来持续上升, 盐沼湿地等蓝碳生态系统受到严重威胁.在全球气候变化的背景下, 全球河口海岸带区域极端气候事件发生频率明显增加(Han et al., 2018; de Lacerda et al., 2020; Newton et al., 2020).气候变化会造成热量、土壤水分与盐度等环境要素的改变, 改变原有盐沼湿地生态系统的组成与结构, 影响盐沼湿地群落的演替方向与进程, 直接改变盐沼湿地的碳循环关键过程(Spencer et al., 2016).有研究表明, 气候变暖条件下盐沼湿地净碳积累取决于矿化沉积物沉积速率与有机质积累速率之间的权衡, 模型预测在21世纪上半叶全球气候变化将导致盐沼湿地碳埋藏速率增加(Kirwan & Mudd, 2012).然而也有研究指出, 气候变暖引起的海平面上升会导致大量的盐沼湿地被海水淹没, 使得气候变化对湿地固碳的促进作用最终消失(DeLaune & White, 2012; Rogers et al., 2019).由此可见, 气候变化对盐沼湿地固碳功能存在着复杂、长期且动态的影响.因此, 探索滨海盐沼湿地固碳功能对气候变化的响应时, 应充分考虑气候变化及其导致后果的多重影响, 并需要开展长期的模拟预测. ...
1
2018
... 由于盐沼湿地处于陆海过渡带, 周期性潮汐作用及其所控制的植被分布与演替过程是控制和维持盐沼湿地碳循环过程和碳汇功能的特异性机制(韩广轩, 2017).受陆海动力交替作用, 盐沼湿地植被时空动态演替复杂而迅速.同时, 盐沼植被类型由海向陆具有明显的梯度分布.陆域向海扩张促使植被向海方向不断演替更迭的同时, 原来靠近海域的地方渐渐深入内地, 海洋环境特征逐渐减弱, 盐生植被向着淡水和旱生植被演替.因而, 盐沼湿地植被的演替不仅表现在空间地域上的植被类型的更替, 也表现在时间序列上的演进(韩广轩等, 2018).同时, 盐沼湿地具有横向扩张和快速淤积的发育特征, 潮汐运动通过沉积物的供给直接影响盐沼湿地土壤碳封存能力, 从而影响盐沼湿地碳汇形成机制. ...
1
2018
... 由于盐沼湿地处于陆海过渡带, 周期性潮汐作用及其所控制的植被分布与演替过程是控制和维持盐沼湿地碳循环过程和碳汇功能的特异性机制(韩广轩, 2017).受陆海动力交替作用, 盐沼湿地植被时空动态演替复杂而迅速.同时, 盐沼植被类型由海向陆具有明显的梯度分布.陆域向海扩张促使植被向海方向不断演替更迭的同时, 原来靠近海域的地方渐渐深入内地, 海洋环境特征逐渐减弱, 盐生植被向着淡水和旱生植被演替.因而, 盐沼湿地植被的演替不仅表现在空间地域上的植被类型的更替, 也表现在时间序列上的演进(韩广轩等, 2018).同时, 盐沼湿地具有横向扩张和快速淤积的发育特征, 潮汐运动通过沉积物的供给直接影响盐沼湿地土壤碳封存能力, 从而影响盐沼湿地碳汇形成机制. ...
The Ocean as a Solution to Climate Change: Five Opportunities for Action
1
2019
... 全球滨海湿地面积约为20.3 × 104 km2, 我国滨海湿地面积约占全球滨海湿地总面积的1/4, 其中滨海盐沼湿地是我国最主要的滨海湿地类型(曹磊等, 2013).据估算, 我国蓝碳生态系统的碳年埋藏量为0.349-0.835 Tg, 其中盐沼湿地约占80%, 远高于红树林和海草床, 是我国蓝碳碳汇的主要贡献者(周晨昊等, 2016).模型模拟结果表明, 未来气候变暖和海平面上升可能使得盐沼湿地能够更迅速捕获和埋藏大气中的碳(Kirwan & Mudd, 2012).另外, 加强全球滨海湿地的保护和修复, 每年能够吸收、储存和减少0.32-0.89 Tg CO2排放(Hoegh-Guldberg et al., 2019).2010年, 保护国际基金会、世界自然保护联盟、联合国教科文组织政府间海洋学委员会共同发起“蓝碳倡议” (The Blue Carbon Initiative), 旨在支撑全球蓝碳的科学研究、项目实施和政策制定等, 通过恢复和保护受损蓝碳生态系统工作而产生碳效益, 减缓气候变化. ...
米草引种对潮滩沉积物有机质的贡献及碳埋藏的影响
1
2018
... 解析滨海湿地埋藏碳的来源所使用的方法主要是碳氮比值法(C:N)、稳定同位素组成法和生物标记物法(Xia et al., 2021a).综合利用碳、氮稳定性同位素丰度(δ)可以解析湿地沉积物有机质的来源(Xia et al., 2021b).通过对沉积物C:N的测定可以区分其碳的来源是水生植被还是陆生植被, 水生植物C:N一般在4-10之间, 而陆生维管植物和挺水植物C:N一般大于20.同时, 沉积物有机质中δ13C和δ15N与有机质来源紧密相关, 通过对滨海湿地地上植被和周围水体主要植物体δ13C的测定可以区分其沉积物中内源有机碳的来源(黄梅等, 2018).除了δ13C, δ15N也被经常用于解析有机质的来源.典型陆地植物的δ15N变化范围为0-11‰, 海草δ15N变化范围为6‰-12‰, 而海洋中藻类的δ15N变化范围为0-4‰ (Samper- Villarreal et al., 2016).另外, 生物标志物是环境和地质体中记载着原始生物母质分子结构信息的有机化合物, 被广泛用于研究埋藏碳的来源和转化特征(冯晓娟等, 2020).例如, 正构烷烃广泛分布于低等细菌藻类与高等植物等不同类型的生物体的蜡质脂类中, 来源于不同生物体的正构烷烃, 其碳数组成与分布特征不同(冯晓娟等, 2020), 可以用来探究沉积物有机质内源菌藻、水生植物、草本植物与木本植物的相对贡献.氨基糖在土壤中更稳定而且保存更长时间(Joergensen, 2018), 是土壤中微生物残体碳的主要标记物, 被广泛应用于研究微生物残体对土壤有机碳和各种微环境流通量的贡献(Joergensen, 2018).综合运用C:N法、稳定同位素组成法和生物标记物法可以有效地解析湿地沉积物中有机碳的来源及其转化过程, 是当前湿地碳循环研究的热点方向(冯晓娟等, 2020; Xia et al., 2021b; Yuan et al., 2021). ...
米草引种对潮滩沉积物有机质的贡献及碳埋藏的影响
1
2018
... 解析滨海湿地埋藏碳的来源所使用的方法主要是碳氮比值法(C:N)、稳定同位素组成法和生物标记物法(Xia et al., 2021a).综合利用碳、氮稳定性同位素丰度(δ)可以解析湿地沉积物有机质的来源(Xia et al., 2021b).通过对沉积物C:N的测定可以区分其碳的来源是水生植被还是陆生植被, 水生植物C:N一般在4-10之间, 而陆生维管植物和挺水植物C:N一般大于20.同时, 沉积物有机质中δ13C和δ15N与有机质来源紧密相关, 通过对滨海湿地地上植被和周围水体主要植物体δ13C的测定可以区分其沉积物中内源有机碳的来源(黄梅等, 2018).除了δ13C, δ15N也被经常用于解析有机质的来源.典型陆地植物的δ15N变化范围为0-11‰, 海草δ15N变化范围为6‰-12‰, 而海洋中藻类的δ15N变化范围为0-4‰ (Samper- Villarreal et al., 2016).另外, 生物标志物是环境和地质体中记载着原始生物母质分子结构信息的有机化合物, 被广泛用于研究埋藏碳的来源和转化特征(冯晓娟等, 2020).例如, 正构烷烃广泛分布于低等细菌藻类与高等植物等不同类型的生物体的蜡质脂类中, 来源于不同生物体的正构烷烃, 其碳数组成与分布特征不同(冯晓娟等, 2020), 可以用来探究沉积物有机质内源菌藻、水生植物、草本植物与木本植物的相对贡献.氨基糖在土壤中更稳定而且保存更长时间(Joergensen, 2018), 是土壤中微生物残体碳的主要标记物, 被广泛应用于研究微生物残体对土壤有机碳和各种微环境流通量的贡献(Joergensen, 2018).综合运用C:N法、稳定同位素组成法和生物标记物法可以有效地解析湿地沉积物中有机碳的来源及其转化过程, 是当前湿地碳循环研究的热点方向(冯晓娟等, 2020; Xia et al., 2021b; Yuan et al., 2021). ...
Amino sugars as specific indices for fungal and bacterial residues in soil
2
2018
... 解析滨海湿地埋藏碳的来源所使用的方法主要是碳氮比值法(C:N)、稳定同位素组成法和生物标记物法(Xia et al., 2021a).综合利用碳、氮稳定性同位素丰度(δ)可以解析湿地沉积物有机质的来源(Xia et al., 2021b).通过对沉积物C:N的测定可以区分其碳的来源是水生植被还是陆生植被, 水生植物C:N一般在4-10之间, 而陆生维管植物和挺水植物C:N一般大于20.同时, 沉积物有机质中δ13C和δ15N与有机质来源紧密相关, 通过对滨海湿地地上植被和周围水体主要植物体δ13C的测定可以区分其沉积物中内源有机碳的来源(黄梅等, 2018).除了δ13C, δ15N也被经常用于解析有机质的来源.典型陆地植物的δ15N变化范围为0-11‰, 海草δ15N变化范围为6‰-12‰, 而海洋中藻类的δ15N变化范围为0-4‰ (Samper- Villarreal et al., 2016).另外, 生物标志物是环境和地质体中记载着原始生物母质分子结构信息的有机化合物, 被广泛用于研究埋藏碳的来源和转化特征(冯晓娟等, 2020).例如, 正构烷烃广泛分布于低等细菌藻类与高等植物等不同类型的生物体的蜡质脂类中, 来源于不同生物体的正构烷烃, 其碳数组成与分布特征不同(冯晓娟等, 2020), 可以用来探究沉积物有机质内源菌藻、水生植物、草本植物与木本植物的相对贡献.氨基糖在土壤中更稳定而且保存更长时间(Joergensen, 2018), 是土壤中微生物残体碳的主要标记物, 被广泛应用于研究微生物残体对土壤有机碳和各种微环境流通量的贡献(Joergensen, 2018).综合运用C:N法、稳定同位素组成法和生物标记物法可以有效地解析湿地沉积物中有机碳的来源及其转化过程, 是当前湿地碳循环研究的热点方向(冯晓娟等, 2020; Xia et al., 2021b; Yuan et al., 2021). ...
... ), 是土壤中微生物残体碳的主要标记物, 被广泛应用于研究微生物残体对土壤有机碳和各种微环境流通量的贡献(Joergensen, 2018).综合运用C:N法、稳定同位素组成法和生物标记物法可以有效地解析湿地沉积物中有机碳的来源及其转化过程, 是当前湿地碳循环研究的热点方向(冯晓娟等, 2020; Xia et al., 2021b; Yuan et al., 2021). ...
Total ecosystem carbon stocks at the marine-terrestrial interface: blue carbon of the Pacific Northwest Coast, United States
1
2020
... 盐沼湿地的碳汇通常以有机碳在土壤或沉积物中的积累为主要特征, 明确土壤碳的存在形式以及与微生物的协同关系成为理解土壤碳库稳定性的基础.盐沼湿地是巨大的土壤碳库, 较高的碳固持能力赋予了盐沼湿地显著的蓝碳碳汇功能, 受海洋潮汐、地表径流及土壤生物生命活动的影响, 土壤碳库的微小变化会极大地调节生态系统碳收支平衡, 成为制约盐沼湿地碳汇潜力的重要因素(Sanderman et al., 2017; Kauffman et al., 2020).同时, 在海洋潮汐和地表径流等水文作用下, 土壤有机碳以溶解性形式进入邻近水域, 是盐沼湿地重要的碳流失途径.除水文作用外, 土壤碳在微生物参与下发生的矿化分解也是关键的碳丢失途径, 包括盐沼植物的凋落分解、土壤有机碳水解、发酵、产酸、产CO2和CH4等(图2).因此, 盐沼湿地土壤碳库稳定性在形成全球蓝碳碳库过程中扮演着至关重要的角色.寻求蓝碳“开源”途径的同时, 深入认识并合理控制“碳节流”也是提高盐沼湿地碳储存能力的重要途径. ...
A national approach to greenhouse gas abatement through blue carbon management
1
2020
... (5)盐沼湿地蓝碳监测、核算及其增汇技术.当前盐沼湿地蓝碳监测技术、评价标准及核算体系等国内外还没有统一的规范和标准, 碳储量核算存在较大的争议, 亟待建立和完善盐沼湿地蓝碳监测与核算技术标准, 编制国际认可的蓝碳核算技术体系; 同时, 建立盐沼湿地碳汇观测体系, 强化固碳过程精细化观测体系, 建立融合现场观测-遥感反演-模型模拟等研究手段, 提高对蓝碳增汇机制的科学认识和对未来碳汇强度的预测能力.与近海环境综合整治、退养还滩还湿、退化滨海湿地生态恢复相结合, 研发盐沼湿地固碳增汇关键技术体系和综合管理方法, 并进行规模化应用.例如, 通过控制陆源污染排放和水体富营养化, 恢复湿地潮汐等水文过程, 增加蓝碳累积(Macreadie et al., 2017; Kelleway et al., 2020).通过重建高生物量、高碳汇型水生生物群落, 改善湿地土壤及水体环境等措施来建立海岸带退化湿地的固碳增汇技术体系(李捷等, 2019).在中国滨海湿地生态系统中, 通过实施“南红北柳”项目, 恢复和种植红树林、柽柳(Tamarix chinensis)、芦苇和盐地碱蓬等植物, 扩大滨海湿地面积, 增强蓝色碳汇潜力. ...
Response of salt-marsh carbon accumulation to climate change
2
2012
... 全球滨海湿地面积约为20.3 × 104 km2, 我国滨海湿地面积约占全球滨海湿地总面积的1/4, 其中滨海盐沼湿地是我国最主要的滨海湿地类型(曹磊等, 2013).据估算, 我国蓝碳生态系统的碳年埋藏量为0.349-0.835 Tg, 其中盐沼湿地约占80%, 远高于红树林和海草床, 是我国蓝碳碳汇的主要贡献者(周晨昊等, 2016).模型模拟结果表明, 未来气候变暖和海平面上升可能使得盐沼湿地能够更迅速捕获和埋藏大气中的碳(Kirwan & Mudd, 2012).另外, 加强全球滨海湿地的保护和修复, 每年能够吸收、储存和减少0.32-0.89 Tg CO2排放(Hoegh-Guldberg et al., 2019).2010年, 保护国际基金会、世界自然保护联盟、联合国教科文组织政府间海洋学委员会共同发起“蓝碳倡议” (The Blue Carbon Initiative), 旨在支撑全球蓝碳的科学研究、项目实施和政策制定等, 通过恢复和保护受损蓝碳生态系统工作而产生碳效益, 减缓气候变化. ...
... 气候变暖导致全球海平面自20世纪初以来持续上升, 盐沼湿地等蓝碳生态系统受到严重威胁.在全球气候变化的背景下, 全球河口海岸带区域极端气候事件发生频率明显增加(Han et al., 2018; de Lacerda et al., 2020; Newton et al., 2020).气候变化会造成热量、土壤水分与盐度等环境要素的改变, 改变原有盐沼湿地生态系统的组成与结构, 影响盐沼湿地群落的演替方向与进程, 直接改变盐沼湿地的碳循环关键过程(Spencer et al., 2016).有研究表明, 气候变暖条件下盐沼湿地净碳积累取决于矿化沉积物沉积速率与有机质积累速率之间的权衡, 模型预测在21世纪上半叶全球气候变化将导致盐沼湿地碳埋藏速率增加(Kirwan & Mudd, 2012).然而也有研究指出, 气候变暖引起的海平面上升会导致大量的盐沼湿地被海水淹没, 使得气候变化对湿地固碳的促进作用最终消失(DeLaune & White, 2012; Rogers et al., 2019).由此可见, 气候变化对盐沼湿地固碳功能存在着复杂、长期且动态的影响.因此, 探索滨海盐沼湿地固碳功能对气候变化的响应时, 应充分考虑气候变化及其导致后果的多重影响, 并需要开展长期的模拟预测. ...
Elevated CO2 stimulates marsh elevation gain, counterbalancing sea-level rise
1
2009
... (4)气候变化和海平面上升背景下盐沼湿地蓝碳模拟与增汇潜力评估.气候变化和海平面上升改变了盐沼湿地的生物地球化学过程、水文条件以及植被生长和演替, 导致了盐沼湿地蓝碳潜力存在极大的不确定性.气候变化和海平面上升增加滨海湿地碳汇功能(Langley et al., 2009; Wang et al., 2019b, 2021a), 也会通过海岸侵蚀导致更多的碳流失(DeLaune & White, 2012), 但现有模型大多基于气候驱动因素、沉积和碳累积速率等, 缺乏针对生态系统和植被演替过程的考虑.另外, 增温、富营养化和海平面上升对滨海湿地碳储存的影响机制不同(Spivak et al., 2019), 这些因素的改变在极大程度上影响了滨海盐沼湿地的固碳能力, 但目前很难精确地预测这种未知影响.因此, 量化这些因素如何影响盐沼湿地有机碳固存与分解对于构建更具预测性的蓝碳模型至关重要.同时急需构建不同尺度气候变化、海平面上升、生态系统演替耦合驱动的盐沼湿地碳循环关键过程模拟预测新范式, 增强盐沼湿地蓝碳增汇潜力评估的可靠性. ...
Persistence of soil organic carbon caused by functional complexity
1
2020
... 目前, 国内外学者已经使用多种技术手段, 如13C或14C标记, 对农田、草地和森林等生态系统的土壤碳矿化过程、行为及参与关键单元开展了研究.但是盐沼湿地有关研究相对滞后, 主要集中在原位或者微宇宙培养下, 生物地球化学参数、基础微生物等指标的测定分析(Doroski et al., 2019).近期研究发现盐沼湿地土壤碳稳定性与土壤碳官能团种类息息相关(Luk et al., 2021).如Roth等(2019)研究指出可溶性有机物质在土壤中持久性保存归因于其在沉积过程中的分子变化.沉积过程中易被利用的烷氧碳比例逐渐降低, 而稳定性较高的烷基碳比例逐渐升高.盐沼湿地处于长期淹水或间歇性淹水, 氧化还原过程频繁发生(图2), 微生物-矿物-有机碳的相互作用对盐沼湿地碳的保存和流失起重要的调控作用.除盐沼土壤内部活跃的生物地球化学过程, 在长期外源物输入条件下, 微生物群落调节着土壤有机碳的种类和含量(Georgiou et al., 2017).外源有机质的多样性将会影响微生物胞外酶的分泌, 进而影响土壤有机质的形成和分解过程(Lehmann et al., 2020).另外, 海水入侵对盐沼湿地微生物参与的土壤碳矿化过程和CO2/CH4排放具有一定的抑制作用(Doroski et al., 2019).但是, 目前关于盐沼湿地土壤有机碳分子结构转换与土壤微生物的耦合关系尚不明确.深入认识微生物相关过程是理解土壤碳库稳定性的基础, 也是认知蓝碳潜力的关键. ...
中国海岸带蓝碳现状分析
1
2019
... (5)盐沼湿地蓝碳监测、核算及其增汇技术.当前盐沼湿地蓝碳监测技术、评价标准及核算体系等国内外还没有统一的规范和标准, 碳储量核算存在较大的争议, 亟待建立和完善盐沼湿地蓝碳监测与核算技术标准, 编制国际认可的蓝碳核算技术体系; 同时, 建立盐沼湿地碳汇观测体系, 强化固碳过程精细化观测体系, 建立融合现场观测-遥感反演-模型模拟等研究手段, 提高对蓝碳增汇机制的科学认识和对未来碳汇强度的预测能力.与近海环境综合整治、退养还滩还湿、退化滨海湿地生态恢复相结合, 研发盐沼湿地固碳增汇关键技术体系和综合管理方法, 并进行规模化应用.例如, 通过控制陆源污染排放和水体富营养化, 恢复湿地潮汐等水文过程, 增加蓝碳累积(Macreadie et al., 2017; Kelleway et al., 2020).通过重建高生物量、高碳汇型水生生物群落, 改善湿地土壤及水体环境等措施来建立海岸带退化湿地的固碳增汇技术体系(李捷等, 2019).在中国滨海湿地生态系统中, 通过实施“南红北柳”项目, 恢复和种植红树林、柽柳(Tamarix chinensis)、芦苇和盐地碱蓬等植物, 扩大滨海湿地面积, 增强蓝色碳汇潜力. ...
中国海岸带蓝碳现状分析
1
2019
... (5)盐沼湿地蓝碳监测、核算及其增汇技术.当前盐沼湿地蓝碳监测技术、评价标准及核算体系等国内外还没有统一的规范和标准, 碳储量核算存在较大的争议, 亟待建立和完善盐沼湿地蓝碳监测与核算技术标准, 编制国际认可的蓝碳核算技术体系; 同时, 建立盐沼湿地碳汇观测体系, 强化固碳过程精细化观测体系, 建立融合现场观测-遥感反演-模型模拟等研究手段, 提高对蓝碳增汇机制的科学认识和对未来碳汇强度的预测能力.与近海环境综合整治、退养还滩还湿、退化滨海湿地生态恢复相结合, 研发盐沼湿地固碳增汇关键技术体系和综合管理方法, 并进行规模化应用.例如, 通过控制陆源污染排放和水体富营养化, 恢复湿地潮汐等水文过程, 增加蓝碳累积(Macreadie et al., 2017; Kelleway et al., 2020).通过重建高生物量、高碳汇型水生生物群落, 改善湿地土壤及水体环境等措施来建立海岸带退化湿地的固碳增汇技术体系(李捷等, 2019).在中国滨海湿地生态系统中, 通过实施“南红北柳”项目, 恢复和种植红树林、柽柳(Tamarix chinensis)、芦苇和盐地碱蓬等植物, 扩大滨海湿地面积, 增强蓝色碳汇潜力. ...
Carbon input and allocation by rice into paddy soils: a review
1
2019
... 植物光合碳分配也是植物采用的一种应对环境胁迫的适应性策略(Stefan et al., 2018; Wang et al., 2021b).例如随着植物可用养分在干旱胁迫环境下变得越来越有限, 植物可能向根部分配更多的光合碳用来吸收水分, 从而引发土壤有机质分解并增加植物养分的可利用性(Williams & de Vries, 2020).同时干旱还可以通过调节根际分泌物的质量和数量等加快或减缓土壤碳循环(Preece et al., 2018).相应地, 滨海湿地生态系统水文过程主要受周期性潮汐作用以及地表淡水与地下咸水之间的相互作用, 导致湿地土壤盐碱化或地表淹水(Han et al., 2018), 也可能改变植被光合碳分配策略.目前, 国内外学者已经使用13C或14C标记对农田、草地和森林等旱地生态系统的光合碳及其后续转化进行了定量分析(Pausch & Kuzyakov, 2018; Wang et al., 2019a).由于盐沼湿地长期或周期性淹水和厌氧, 其植物光合碳分配途径有别于其他生态系统, 因此对旱地生态系统的归纳不一定能够阐明湿地生态系统碳循环的关键过程(Liu et al., 2019).因此, 在全球气候变化背景下, 迫切需要了解盐沼湿地植物-土壤中光合碳分配过程及比例, 这对于理解和精确评估盐沼湿地碳汇功能及构建模型具有重要意义. ...
Soil organic carbon development and turnover in natural and disturbed salt marsh environments
1
2021
... 目前, 国内外学者已经使用多种技术手段, 如13C或14C标记, 对农田、草地和森林等生态系统的土壤碳矿化过程、行为及参与关键单元开展了研究.但是盐沼湿地有关研究相对滞后, 主要集中在原位或者微宇宙培养下, 生物地球化学参数、基础微生物等指标的测定分析(Doroski et al., 2019).近期研究发现盐沼湿地土壤碳稳定性与土壤碳官能团种类息息相关(Luk et al., 2021).如Roth等(2019)研究指出可溶性有机物质在土壤中持久性保存归因于其在沉积过程中的分子变化.沉积过程中易被利用的烷氧碳比例逐渐降低, 而稳定性较高的烷基碳比例逐渐升高.盐沼湿地处于长期淹水或间歇性淹水, 氧化还原过程频繁发生(图2), 微生物-矿物-有机碳的相互作用对盐沼湿地碳的保存和流失起重要的调控作用.除盐沼土壤内部活跃的生物地球化学过程, 在长期外源物输入条件下, 微生物群落调节着土壤有机碳的种类和含量(Georgiou et al., 2017).外源有机质的多样性将会影响微生物胞外酶的分泌, 进而影响土壤有机质的形成和分解过程(Lehmann et al., 2020).另外, 海水入侵对盐沼湿地微生物参与的土壤碳矿化过程和CO2/CH4排放具有一定的抑制作用(Doroski et al., 2019).但是, 目前关于盐沼湿地土壤有机碳分子结构转换与土壤微生物的耦合关系尚不明确.深入认识微生物相关过程是理解土壤碳库稳定性的基础, 也是认知蓝碳潜力的关键. ...
The future of Blue Carbon science
1
2019
... 2009年, 联合国环境规划署、粮农组织和教科文组织政府间海洋学委员会联合发布《蓝碳: 健康海洋对碳的固定作用》(《Blue Carbon: the Role of Healthy Oceans in Binding Carbon》), 确认了海洋在减缓全球气候变化和碳循环过程中至关重要的作用, 首次提出了“蓝碳”的概念, 并明确指出盐沼湿地、红树林和海草床三类海岸带蓝碳是相对易于管理的(Nellemann et al., 2009).2019年, 联合国政府间气候变化专门委员会(IPCC)发布《气候变化中的海洋与冰冻圈特别报告》(《Special Report on the Ocean and Cryosphere in a Changing Climate》), 明确指出是否“易于管理”是界定蓝碳的必要条件.滨海盐沼湿地(coastal salt marshes)是指海岸沿线受海洋潮汐周期性或间歇性影响, 有盐生植物覆盖的咸水或淡咸水淤泥质滩涂, 是地球上高生产力植被类型之一.盐沼湿地具有很高的固碳能力, 同时土壤能够捕获和储存大量的碳, 其土壤碳埋藏速率为(218 ± 24) g·m-2·a-1, 比森林生态系统高40倍左右(Mcleod et al., 2011; Macreadie et al., 2019).据估计, 盐沼湿地上层(约1 m)土壤含有约917 t CO2 e·hm-2 (每hm2 CO2当量), 年固碳量约为(8.0 ± 8.5) t CO2 e·hm-2 (Sapkota & White, 2021).因此, 滨海盐沼湿地与红树林、海草床等海岸带生态系统被称为蓝碳生态系统.虽然海岸带蓝碳生态系统只占全球海洋面积的0.2%, 但贡献了海洋沉积物碳总储量的50% (Duarte et al., 2013). ...
Can we manage coastal ecosystems to sequester more blue carbon
1
2017
... (5)盐沼湿地蓝碳监测、核算及其增汇技术.当前盐沼湿地蓝碳监测技术、评价标准及核算体系等国内外还没有统一的规范和标准, 碳储量核算存在较大的争议, 亟待建立和完善盐沼湿地蓝碳监测与核算技术标准, 编制国际认可的蓝碳核算技术体系; 同时, 建立盐沼湿地碳汇观测体系, 强化固碳过程精细化观测体系, 建立融合现场观测-遥感反演-模型模拟等研究手段, 提高对蓝碳增汇机制的科学认识和对未来碳汇强度的预测能力.与近海环境综合整治、退养还滩还湿、退化滨海湿地生态恢复相结合, 研发盐沼湿地固碳增汇关键技术体系和综合管理方法, 并进行规模化应用.例如, 通过控制陆源污染排放和水体富营养化, 恢复湿地潮汐等水文过程, 增加蓝碳累积(Macreadie et al., 2017; Kelleway et al., 2020).通过重建高生物量、高碳汇型水生生物群落, 改善湿地土壤及水体环境等措施来建立海岸带退化湿地的固碳增汇技术体系(李捷等, 2019).在中国滨海湿地生态系统中, 通过实施“南红北柳”项目, 恢复和种植红树林、柽柳(Tamarix chinensis)、芦苇和盐地碱蓬等植物, 扩大滨海湿地面积, 增强蓝色碳汇潜力. ...
A blueprint for blue carbon: toward an improved understanding of the role of vegetated coastal habitats in sequestering CO2
2
2011
... 2009年, 联合国环境规划署、粮农组织和教科文组织政府间海洋学委员会联合发布《蓝碳: 健康海洋对碳的固定作用》(《Blue Carbon: the Role of Healthy Oceans in Binding Carbon》), 确认了海洋在减缓全球气候变化和碳循环过程中至关重要的作用, 首次提出了“蓝碳”的概念, 并明确指出盐沼湿地、红树林和海草床三类海岸带蓝碳是相对易于管理的(Nellemann et al., 2009).2019年, 联合国政府间气候变化专门委员会(IPCC)发布《气候变化中的海洋与冰冻圈特别报告》(《Special Report on the Ocean and Cryosphere in a Changing Climate》), 明确指出是否“易于管理”是界定蓝碳的必要条件.滨海盐沼湿地(coastal salt marshes)是指海岸沿线受海洋潮汐周期性或间歇性影响, 有盐生植物覆盖的咸水或淡咸水淤泥质滩涂, 是地球上高生产力植被类型之一.盐沼湿地具有很高的固碳能力, 同时土壤能够捕获和储存大量的碳, 其土壤碳埋藏速率为(218 ± 24) g·m-2·a-1, 比森林生态系统高40倍左右(Mcleod et al., 2011; Macreadie et al., 2019).据估计, 盐沼湿地上层(约1 m)土壤含有约917 t CO2 e·hm-2 (每hm2 CO2当量), 年固碳量约为(8.0 ± 8.5) t CO2 e·hm-2 (Sapkota & White, 2021).因此, 滨海盐沼湿地与红树林、海草床等海岸带生态系统被称为蓝碳生态系统.虽然海岸带蓝碳生态系统只占全球海洋面积的0.2%, 但贡献了海洋沉积物碳总储量的50% (Duarte et al., 2013). ...
... (2)盐沼湿地土壤有机碳沉积和埋藏速率及其对全球变化的响应.盐沼湿地具有很高的碳捕获能力和长期持续固碳能力, 特别是在河流携带大量泥沙和营养元素的河口区, 其储碳机理主要是在沉积物厌氧环境对有机质分解的抑制作用下, 大量植物残体能够被长期地保存(Mcleod et al., 2011; 王秀君等, 2016).盐沼湿地和红树林的碳埋藏速率约为53.65 Tg·a-1, 换算成CO2当量为196.71 Tg·a-1 (Wang et al., 2021a), 这一数据相当于人类活动每年排放量的0.6% (王法明等, 2021).在未来气候变化和海平面上升情境下, 盐沼湿地蓝碳的沉积和埋藏潜力如何? ...
Tibetan sedges sequester more carbon belowground than grasses: a 13C labeling study
1
2018
... 植物光合碳是植物-土壤系统碳循环的起点, 解析植物-土壤系统中光合碳的分配过程及机制是估算盐沼湿地固碳功能的先决条件.植物主要通过光合作用将CO2转化为多糖等光合产物, 由叶片通过枝、干向下运输, 分配到植物体各个组织器官.在生态系统碳循环过程中, 植物会通过调节碳在不同部位的分配来满足自身生长与繁殖的需求或应对周边环境变化, 从而间接影响生物地球化学循环(Wang et al., 2019a).气候条件、土壤理化性质、微生物活性和植物生长发育等均会影响植物对光合碳分配的模式, 改变植物向地上、地下部分的碳输入(Diao et al., 2020).此外, 光合碳固定是碳分配的主要驱动力, 环境胁迫还可以通过影响光合作用影响碳吸收, 从而间接影响光同化物的分配(Pausch & Kuzyakov, 2018).目前, 碳同位素技术是研究植物光合碳分配、周转和固定的重要手段.光合碳在植物体各组织器官内的迁移和转化十分迅速, 碳同化后的第1天从叶片到根系的分配就达到峰值(Pausch & Kuzyakov, 2018).同时, 新合成的光合产物可以通过根系分泌物的形式刺激土壤有机质(老碳)的分解, 即土壤碳输入的激发效应(Street et al., 2020).另外, 光合碳分配在物种间也存在差异.Meta分析发现, 与禾本科植物相比, 莎草科植物可以通过根际沉积将更多的碳转移至根和土壤中, 增加地下碳固存(Mou et al., 2018); 采用稳定碳同位素方法, 发现互花米草(Spartina alterniflora)可以将33%和7%的光合同化碳分配到茎和根, 显著高于芦苇(Phragmites australis)的分配比例(Zhang et al., 2017). ...
Blue Carbon—The Role of Healthy Oceans in Binding Carbon
1
2009
... 2009年, 联合国环境规划署、粮农组织和教科文组织政府间海洋学委员会联合发布《蓝碳: 健康海洋对碳的固定作用》(《Blue Carbon: the Role of Healthy Oceans in Binding Carbon》), 确认了海洋在减缓全球气候变化和碳循环过程中至关重要的作用, 首次提出了“蓝碳”的概念, 并明确指出盐沼湿地、红树林和海草床三类海岸带蓝碳是相对易于管理的(Nellemann et al., 2009).2019年, 联合国政府间气候变化专门委员会(IPCC)发布《气候变化中的海洋与冰冻圈特别报告》(《Special Report on the Ocean and Cryosphere in a Changing Climate》), 明确指出是否“易于管理”是界定蓝碳的必要条件.滨海盐沼湿地(coastal salt marshes)是指海岸沿线受海洋潮汐周期性或间歇性影响, 有盐生植物覆盖的咸水或淡咸水淤泥质滩涂, 是地球上高生产力植被类型之一.盐沼湿地具有很高的固碳能力, 同时土壤能够捕获和储存大量的碳, 其土壤碳埋藏速率为(218 ± 24) g·m-2·a-1, 比森林生态系统高40倍左右(Mcleod et al., 2011; Macreadie et al., 2019).据估计, 盐沼湿地上层(约1 m)土壤含有约917 t CO2 e·hm-2 (每hm2 CO2当量), 年固碳量约为(8.0 ± 8.5) t CO2 e·hm-2 (Sapkota & White, 2021).因此, 滨海盐沼湿地与红树林、海草床等海岸带生态系统被称为蓝碳生态系统.虽然海岸带蓝碳生态系统只占全球海洋面积的0.2%, 但贡献了海洋沉积物碳总储量的50% (Duarte et al., 2013). ...
Anthropogenic, direct pressures on coastal wetlands
1
2020
... 气候变暖导致全球海平面自20世纪初以来持续上升, 盐沼湿地等蓝碳生态系统受到严重威胁.在全球气候变化的背景下, 全球河口海岸带区域极端气候事件发生频率明显增加(Han et al., 2018; de Lacerda et al., 2020; Newton et al., 2020).气候变化会造成热量、土壤水分与盐度等环境要素的改变, 改变原有盐沼湿地生态系统的组成与结构, 影响盐沼湿地群落的演替方向与进程, 直接改变盐沼湿地的碳循环关键过程(Spencer et al., 2016).有研究表明, 气候变暖条件下盐沼湿地净碳积累取决于矿化沉积物沉积速率与有机质积累速率之间的权衡, 模型预测在21世纪上半叶全球气候变化将导致盐沼湿地碳埋藏速率增加(Kirwan & Mudd, 2012).然而也有研究指出, 气候变暖引起的海平面上升会导致大量的盐沼湿地被海水淹没, 使得气候变化对湿地固碳的促进作用最终消失(DeLaune & White, 2012; Rogers et al., 2019).由此可见, 气候变化对盐沼湿地固碳功能存在着复杂、长期且动态的影响.因此, 探索滨海盐沼湿地固碳功能对气候变化的响应时, 应充分考虑气候变化及其导致后果的多重影响, 并需要开展长期的模拟预测. ...
Carbon input by roots into the soil: quantification of rhizodeposition from root to ecosystem scale
3
2018
... 植物光合碳是植物-土壤系统碳循环的起点, 解析植物-土壤系统中光合碳的分配过程及机制是估算盐沼湿地固碳功能的先决条件.植物主要通过光合作用将CO2转化为多糖等光合产物, 由叶片通过枝、干向下运输, 分配到植物体各个组织器官.在生态系统碳循环过程中, 植物会通过调节碳在不同部位的分配来满足自身生长与繁殖的需求或应对周边环境变化, 从而间接影响生物地球化学循环(Wang et al., 2019a).气候条件、土壤理化性质、微生物活性和植物生长发育等均会影响植物对光合碳分配的模式, 改变植物向地上、地下部分的碳输入(Diao et al., 2020).此外, 光合碳固定是碳分配的主要驱动力, 环境胁迫还可以通过影响光合作用影响碳吸收, 从而间接影响光同化物的分配(Pausch & Kuzyakov, 2018).目前, 碳同位素技术是研究植物光合碳分配、周转和固定的重要手段.光合碳在植物体各组织器官内的迁移和转化十分迅速, 碳同化后的第1天从叶片到根系的分配就达到峰值(Pausch & Kuzyakov, 2018).同时, 新合成的光合产物可以通过根系分泌物的形式刺激土壤有机质(老碳)的分解, 即土壤碳输入的激发效应(Street et al., 2020).另外, 光合碳分配在物种间也存在差异.Meta分析发现, 与禾本科植物相比, 莎草科植物可以通过根际沉积将更多的碳转移至根和土壤中, 增加地下碳固存(Mou et al., 2018); 采用稳定碳同位素方法, 发现互花米草(Spartina alterniflora)可以将33%和7%的光合同化碳分配到茎和根, 显著高于芦苇(Phragmites australis)的分配比例(Zhang et al., 2017). ...
... ).目前, 碳同位素技术是研究植物光合碳分配、周转和固定的重要手段.光合碳在植物体各组织器官内的迁移和转化十分迅速, 碳同化后的第1天从叶片到根系的分配就达到峰值(Pausch & Kuzyakov, 2018).同时, 新合成的光合产物可以通过根系分泌物的形式刺激土壤有机质(老碳)的分解, 即土壤碳输入的激发效应(Street et al., 2020).另外, 光合碳分配在物种间也存在差异.Meta分析发现, 与禾本科植物相比, 莎草科植物可以通过根际沉积将更多的碳转移至根和土壤中, 增加地下碳固存(Mou et al., 2018); 采用稳定碳同位素方法, 发现互花米草(Spartina alterniflora)可以将33%和7%的光合同化碳分配到茎和根, 显著高于芦苇(Phragmites australis)的分配比例(Zhang et al., 2017). ...
... 植物光合碳分配也是植物采用的一种应对环境胁迫的适应性策略(Stefan et al., 2018; Wang et al., 2021b).例如随着植物可用养分在干旱胁迫环境下变得越来越有限, 植物可能向根部分配更多的光合碳用来吸收水分, 从而引发土壤有机质分解并增加植物养分的可利用性(Williams & de Vries, 2020).同时干旱还可以通过调节根际分泌物的质量和数量等加快或减缓土壤碳循环(Preece et al., 2018).相应地, 滨海湿地生态系统水文过程主要受周期性潮汐作用以及地表淡水与地下咸水之间的相互作用, 导致湿地土壤盐碱化或地表淹水(Han et al., 2018), 也可能改变植被光合碳分配策略.目前, 国内外学者已经使用13C或14C标记对农田、草地和森林等旱地生态系统的光合碳及其后续转化进行了定量分析(Pausch & Kuzyakov, 2018; Wang et al., 2019a).由于盐沼湿地长期或周期性淹水和厌氧, 其植物光合碳分配途径有别于其他生态系统, 因此对旱地生态系统的归纳不一定能够阐明湿地生态系统碳循环的关键过程(Liu et al., 2019).因此, 在全球气候变化背景下, 迫切需要了解盐沼湿地植物-土壤中光合碳分配过程及比例, 这对于理解和精确评估盐沼湿地碳汇功能及构建模型具有重要意义. ...
Controls on sediment accretion and blue carbon burial in tidal saline wetlands: insights from the oregon coast, USA
1
2020
... 盐沼湿地碳埋藏速率通常通过沉积速率与沉积物碳密度计算而来.沉积物中放射性核素的含量被广泛应用于河口、海岸及陆架区域沉积物年龄的确定, 沉积过程的揭示和物质来源判断等.常用的天然核素为210Pb, 其半衰期为22.3 a, 定年上限为150 a.除天然核素外, 人类核试验等活动还产生了137Cs等人工核素, 137Cs半衰期为30.17 a, 在沉积物垂直分布中通常会出现峰值, 对应特定的核爆年份, 具有重要的纪年意义(Xia et al., 2016).沉积速率研究中一般将137Cs与210Pb纪年结果相互印证(Peck et al., 2020).例如, Ye等(2018)利用210Pb纪年方法研究了黄河三角洲不同植被类型湿地的沉积速率及碳埋藏速率, 发现黄河三角洲盐沼湿地的沉积速率在3.87-9.68 cm·a-1, 而碳埋藏速率则在586-1 784 g·m-2·a-1, 远高于全球盐沼湿地固碳速率的平均值(168 g·m-2·a-1), 显示出黄河三角洲滨海湿地极高的碳埋藏能力. ...
Thirsty tree roots exude more carbon
1
2018
... 植物光合碳分配也是植物采用的一种应对环境胁迫的适应性策略(Stefan et al., 2018; Wang et al., 2021b).例如随着植物可用养分在干旱胁迫环境下变得越来越有限, 植物可能向根部分配更多的光合碳用来吸收水分, 从而引发土壤有机质分解并增加植物养分的可利用性(Williams & de Vries, 2020).同时干旱还可以通过调节根际分泌物的质量和数量等加快或减缓土壤碳循环(Preece et al., 2018).相应地, 滨海湿地生态系统水文过程主要受周期性潮汐作用以及地表淡水与地下咸水之间的相互作用, 导致湿地土壤盐碱化或地表淹水(Han et al., 2018), 也可能改变植被光合碳分配策略.目前, 国内外学者已经使用13C或14C标记对农田、草地和森林等旱地生态系统的光合碳及其后续转化进行了定量分析(Pausch & Kuzyakov, 2018; Wang et al., 2019a).由于盐沼湿地长期或周期性淹水和厌氧, 其植物光合碳分配途径有别于其他生态系统, 因此对旱地生态系统的归纳不一定能够阐明湿地生态系统碳循环的关键过程(Liu et al., 2019).因此, 在全球气候变化背景下, 迫切需要了解盐沼湿地植物-土壤中光合碳分配过程及比例, 这对于理解和精确评估盐沼湿地碳汇功能及构建模型具有重要意义. ...
Coastal blue carbon assessment of mangroves, salt marshes, and salt barrens in Tampa Bay, Florida, USA
1
2018
... 盐沼湿地的蓝碳功能是光合碳吸收、碳沉积埋藏、碳输出等多过程相互作用和平衡的结果(图1), 其关键过程主要包括: (1)植被光合固碳.盐沼湿地通常具有高的净初级生产力, 尤其在河口区, 植被净初级生产力可达1 745 g C·m-2·a-1 (Sousa et al., 2010).(2)光合碳分配.植物通过调节碳在不同部位的分配模式来满足自身生长与繁殖的需求或者应对周边环境变化(Wang et al., 2019a).光合固定的碳向根系和土壤碳库的分配和转移规律, 直接影响着土壤碳库的走向(Xu et al., 2020).盐沼植被根冠比可达1.4-50, 有大量的初级生产力所固定的碳被储存在地下生物量中, 通过根系周转进入土壤碳库(王秀君等, 2016).(3)碳沉积埋藏.由于滨海盐沼湿地不断向下沉积, 土壤碳库很难达到饱和, 储存的碳可以在土壤中保存数千年(Radabaugh et al., 2018).因此, 盐沼湿地的碳封存效率远高于森林或其他陆地生态系统(Wang et al., 2021a).(4)土壤碳流失.潮汐作用导致盐沼湿地间歇性厌氧好氧交替, 氧化还原过程频繁发生, 陆地和海洋微生物交互作用; 同时, 周期性潮汐携带大量的SO42-阻碍CH4产生, 从而降低盐沼湿地碳排放量(Choi & Wang, 2004).另外, 潮汐作用下盐沼湿地可溶性有机碳(DOC)、可溶性无机碳(DIC)、颗粒有机碳(POC)交换及其输出也是其碳流失中不容忽视的组成部分(韩广轩, 2017). ...
Wetland carbon storage controlled by millennial-scale variation in relative sea-level rise
1
2019
... 气候变暖导致全球海平面自20世纪初以来持续上升, 盐沼湿地等蓝碳生态系统受到严重威胁.在全球气候变化的背景下, 全球河口海岸带区域极端气候事件发生频率明显增加(Han et al., 2018; de Lacerda et al., 2020; Newton et al., 2020).气候变化会造成热量、土壤水分与盐度等环境要素的改变, 改变原有盐沼湿地生态系统的组成与结构, 影响盐沼湿地群落的演替方向与进程, 直接改变盐沼湿地的碳循环关键过程(Spencer et al., 2016).有研究表明, 气候变暖条件下盐沼湿地净碳积累取决于矿化沉积物沉积速率与有机质积累速率之间的权衡, 模型预测在21世纪上半叶全球气候变化将导致盐沼湿地碳埋藏速率增加(Kirwan & Mudd, 2012).然而也有研究指出, 气候变暖引起的海平面上升会导致大量的盐沼湿地被海水淹没, 使得气候变化对湿地固碳的促进作用最终消失(DeLaune & White, 2012; Rogers et al., 2019).由此可见, 气候变化对盐沼湿地固碳功能存在着复杂、长期且动态的影响.因此, 探索滨海盐沼湿地固碳功能对气候变化的响应时, 应充分考虑气候变化及其导致后果的多重影响, 并需要开展长期的模拟预测. ...
Persistence of dissolved organic matter explained by molecular changes during its passage through soil
1
2019
... 目前, 国内外学者已经使用多种技术手段, 如13C或14C标记, 对农田、草地和森林等生态系统的土壤碳矿化过程、行为及参与关键单元开展了研究.但是盐沼湿地有关研究相对滞后, 主要集中在原位或者微宇宙培养下, 生物地球化学参数、基础微生物等指标的测定分析(Doroski et al., 2019).近期研究发现盐沼湿地土壤碳稳定性与土壤碳官能团种类息息相关(Luk et al., 2021).如Roth等(2019)研究指出可溶性有机物质在土壤中持久性保存归因于其在沉积过程中的分子变化.沉积过程中易被利用的烷氧碳比例逐渐降低, 而稳定性较高的烷基碳比例逐渐升高.盐沼湿地处于长期淹水或间歇性淹水, 氧化还原过程频繁发生(图2), 微生物-矿物-有机碳的相互作用对盐沼湿地碳的保存和流失起重要的调控作用.除盐沼土壤内部活跃的生物地球化学过程, 在长期外源物输入条件下, 微生物群落调节着土壤有机碳的种类和含量(Georgiou et al., 2017).外源有机质的多样性将会影响微生物胞外酶的分泌, 进而影响土壤有机质的形成和分解过程(Lehmann et al., 2020).另外, 海水入侵对盐沼湿地微生物参与的土壤碳矿化过程和CO2/CH4排放具有一定的抑制作用(Doroski et al., 2019).但是, 目前关于盐沼湿地土壤有机碳分子结构转换与土壤微生物的耦合关系尚不明确.深入认识微生物相关过程是理解土壤碳库稳定性的基础, 也是认知蓝碳潜力的关键. ...
Organic carbon in seagrass sediments is influenced by seagrass canopy complexity, turbidity, wave height, and water depth
1
2016
... 解析滨海湿地埋藏碳的来源所使用的方法主要是碳氮比值法(C:N)、稳定同位素组成法和生物标记物法(Xia et al., 2021a).综合利用碳、氮稳定性同位素丰度(δ)可以解析湿地沉积物有机质的来源(Xia et al., 2021b).通过对沉积物C:N的测定可以区分其碳的来源是水生植被还是陆生植被, 水生植物C:N一般在4-10之间, 而陆生维管植物和挺水植物C:N一般大于20.同时, 沉积物有机质中δ13C和δ15N与有机质来源紧密相关, 通过对滨海湿地地上植被和周围水体主要植物体δ13C的测定可以区分其沉积物中内源有机碳的来源(黄梅等, 2018).除了δ13C, δ15N也被经常用于解析有机质的来源.典型陆地植物的δ15N变化范围为0-11‰, 海草δ15N变化范围为6‰-12‰, 而海洋中藻类的δ15N变化范围为0-4‰ (Samper- Villarreal et al., 2016).另外, 生物标志物是环境和地质体中记载着原始生物母质分子结构信息的有机化合物, 被广泛用于研究埋藏碳的来源和转化特征(冯晓娟等, 2020).例如, 正构烷烃广泛分布于低等细菌藻类与高等植物等不同类型的生物体的蜡质脂类中, 来源于不同生物体的正构烷烃, 其碳数组成与分布特征不同(冯晓娟等, 2020), 可以用来探究沉积物有机质内源菌藻、水生植物、草本植物与木本植物的相对贡献.氨基糖在土壤中更稳定而且保存更长时间(Joergensen, 2018), 是土壤中微生物残体碳的主要标记物, 被广泛应用于研究微生物残体对土壤有机碳和各种微环境流通量的贡献(Joergensen, 2018).综合运用C:N法、稳定同位素组成法和生物标记物法可以有效地解析湿地沉积物中有机碳的来源及其转化过程, 是当前湿地碳循环研究的热点方向(冯晓娟等, 2020; Xia et al., 2021b; Yuan et al., 2021). ...
Soil carbon debt of 12,000 years of human land use
1
2017
... 盐沼湿地的碳汇通常以有机碳在土壤或沉积物中的积累为主要特征, 明确土壤碳的存在形式以及与微生物的协同关系成为理解土壤碳库稳定性的基础.盐沼湿地是巨大的土壤碳库, 较高的碳固持能力赋予了盐沼湿地显著的蓝碳碳汇功能, 受海洋潮汐、地表径流及土壤生物生命活动的影响, 土壤碳库的微小变化会极大地调节生态系统碳收支平衡, 成为制约盐沼湿地碳汇潜力的重要因素(Sanderman et al., 2017; Kauffman et al., 2020).同时, 在海洋潮汐和地表径流等水文作用下, 土壤有机碳以溶解性形式进入邻近水域, 是盐沼湿地重要的碳流失途径.除水文作用外, 土壤碳在微生物参与下发生的矿化分解也是关键的碳丢失途径, 包括盐沼植物的凋落分解、土壤有机碳水解、发酵、产酸、产CO2和CH4等(图2).因此, 盐沼湿地土壤碳库稳定性在形成全球蓝碳碳库过程中扮演着至关重要的角色.寻求蓝碳“开源”途径的同时, 深入认识并合理控制“碳节流”也是提高盐沼湿地碳储存能力的重要途径. ...
Long-term fate of rapidly eroding carbon stock soil profiles in coastal wetlands
1
2021
... 2009年, 联合国环境规划署、粮农组织和教科文组织政府间海洋学委员会联合发布《蓝碳: 健康海洋对碳的固定作用》(《Blue Carbon: the Role of Healthy Oceans in Binding Carbon》), 确认了海洋在减缓全球气候变化和碳循环过程中至关重要的作用, 首次提出了“蓝碳”的概念, 并明确指出盐沼湿地、红树林和海草床三类海岸带蓝碳是相对易于管理的(Nellemann et al., 2009).2019年, 联合国政府间气候变化专门委员会(IPCC)发布《气候变化中的海洋与冰冻圈特别报告》(《Special Report on the Ocean and Cryosphere in a Changing Climate》), 明确指出是否“易于管理”是界定蓝碳的必要条件.滨海盐沼湿地(coastal salt marshes)是指海岸沿线受海洋潮汐周期性或间歇性影响, 有盐生植物覆盖的咸水或淡咸水淤泥质滩涂, 是地球上高生产力植被类型之一.盐沼湿地具有很高的固碳能力, 同时土壤能够捕获和储存大量的碳, 其土壤碳埋藏速率为(218 ± 24) g·m-2·a-1, 比森林生态系统高40倍左右(Mcleod et al., 2011; Macreadie et al., 2019).据估计, 盐沼湿地上层(约1 m)土壤含有约917 t CO2 e·hm-2 (每hm2 CO2当量), 年固碳量约为(8.0 ± 8.5) t CO2 e·hm-2 (Sapkota & White, 2021).因此, 滨海盐沼湿地与红树林、海草床等海岸带生态系统被称为蓝碳生态系统.虽然海岸带蓝碳生态系统只占全球海洋面积的0.2%, 但贡献了海洋沉积物碳总储量的50% (Duarte et al., 2013). ...
Future response of global coastal wetlands to sea-level rise
1
2018
... 在全球范围内, 已开展了气候变暖、海平面上升、泥沙沉积等因素对盐沼湿地及其碳循环过程影响的模拟预测研究(Spencer et al., 2016; Schuerch et al., 2018; Ewers Lewis et al., 2020).尽管这些研究横跨了多个时空尺度, 并解释了盐沼湿地及其碳循环是如何受未来全球变化的影响, 但大量国内外学者在开展盐沼湿地固碳动态模拟时均忽略了由于自身演替而导致的湿地植被群落景观格局的变化, 及其带来的景观-区域尺度盐沼湿地固碳功能的改变(Elschot et al., 2015; Gu et al., 2018).这种影响在快速演替的年轻河口盐沼湿地中表现得更为明显.因此, 想要科学地认识盐沼湿地对气候变化影响的响应, 准确地评估滨海盐沼湿地的固碳功能在未来减缓气候变化中所发挥的作用, 除了需要深入地了解全球变化对盐沼植被碳吸收、碳分配、碳埋藏等过程的影响外, 还需要全面地把握全球变化带来的环境改变而引起的盐沼植被群落空间分布及其演替动态对整个生态系统碳循环过程的影响.在此基础之上, 耦合植被群落演替与碳循环两类模型是实现生态系统-景观-区域水平的碳循环过程的模拟与碳汇预测的有效途径.因此, 亟待构建区域尺度气候变化、海平面上升、生态系统演替耦合驱动的盐沼湿地碳循环关键过程模拟预测新范式, 增强盐沼湿地蓝碳增汇潜力评估的可靠性. ...
Productivity and nutrient cycling in salt marshes: contribution to ecosystem health
1
2010
... 盐沼湿地的蓝碳功能是光合碳吸收、碳沉积埋藏、碳输出等多过程相互作用和平衡的结果(图1), 其关键过程主要包括: (1)植被光合固碳.盐沼湿地通常具有高的净初级生产力, 尤其在河口区, 植被净初级生产力可达1 745 g C·m-2·a-1 (Sousa et al., 2010).(2)光合碳分配.植物通过调节碳在不同部位的分配模式来满足自身生长与繁殖的需求或者应对周边环境变化(Wang et al., 2019a).光合固定的碳向根系和土壤碳库的分配和转移规律, 直接影响着土壤碳库的走向(Xu et al., 2020).盐沼植被根冠比可达1.4-50, 有大量的初级生产力所固定的碳被储存在地下生物量中, 通过根系周转进入土壤碳库(王秀君等, 2016).(3)碳沉积埋藏.由于滨海盐沼湿地不断向下沉积, 土壤碳库很难达到饱和, 储存的碳可以在土壤中保存数千年(Radabaugh et al., 2018).因此, 盐沼湿地的碳封存效率远高于森林或其他陆地生态系统(Wang et al., 2021a).(4)土壤碳流失.潮汐作用导致盐沼湿地间歇性厌氧好氧交替, 氧化还原过程频繁发生, 陆地和海洋微生物交互作用; 同时, 周期性潮汐携带大量的SO42-阻碍CH4产生, 从而降低盐沼湿地碳排放量(Choi & Wang, 2004).另外, 潮汐作用下盐沼湿地可溶性有机碳(DOC)、可溶性无机碳(DIC)、颗粒有机碳(POC)交换及其输出也是其碳流失中不容忽视的组成部分(韩广轩, 2017). ...
Global coastal wetland change under sea-level rise and related stresses: the DIVA Wetland Change Model
2
2016
... 气候变暖导致全球海平面自20世纪初以来持续上升, 盐沼湿地等蓝碳生态系统受到严重威胁.在全球气候变化的背景下, 全球河口海岸带区域极端气候事件发生频率明显增加(Han et al., 2018; de Lacerda et al., 2020; Newton et al., 2020).气候变化会造成热量、土壤水分与盐度等环境要素的改变, 改变原有盐沼湿地生态系统的组成与结构, 影响盐沼湿地群落的演替方向与进程, 直接改变盐沼湿地的碳循环关键过程(Spencer et al., 2016).有研究表明, 气候变暖条件下盐沼湿地净碳积累取决于矿化沉积物沉积速率与有机质积累速率之间的权衡, 模型预测在21世纪上半叶全球气候变化将导致盐沼湿地碳埋藏速率增加(Kirwan & Mudd, 2012).然而也有研究指出, 气候变暖引起的海平面上升会导致大量的盐沼湿地被海水淹没, 使得气候变化对湿地固碳的促进作用最终消失(DeLaune & White, 2012; Rogers et al., 2019).由此可见, 气候变化对盐沼湿地固碳功能存在着复杂、长期且动态的影响.因此, 探索滨海盐沼湿地固碳功能对气候变化的响应时, 应充分考虑气候变化及其导致后果的多重影响, 并需要开展长期的模拟预测. ...
... 在全球范围内, 已开展了气候变暖、海平面上升、泥沙沉积等因素对盐沼湿地及其碳循环过程影响的模拟预测研究(Spencer et al., 2016; Schuerch et al., 2018; Ewers Lewis et al., 2020).尽管这些研究横跨了多个时空尺度, 并解释了盐沼湿地及其碳循环是如何受未来全球变化的影响, 但大量国内外学者在开展盐沼湿地固碳动态模拟时均忽略了由于自身演替而导致的湿地植被群落景观格局的变化, 及其带来的景观-区域尺度盐沼湿地固碳功能的改变(Elschot et al., 2015; Gu et al., 2018).这种影响在快速演替的年轻河口盐沼湿地中表现得更为明显.因此, 想要科学地认识盐沼湿地对气候变化影响的响应, 准确地评估滨海盐沼湿地的固碳功能在未来减缓气候变化中所发挥的作用, 除了需要深入地了解全球变化对盐沼植被碳吸收、碳分配、碳埋藏等过程的影响外, 还需要全面地把握全球变化带来的环境改变而引起的盐沼植被群落空间分布及其演替动态对整个生态系统碳循环过程的影响.在此基础之上, 耦合植被群落演替与碳循环两类模型是实现生态系统-景观-区域水平的碳循环过程的模拟与碳汇预测的有效途径.因此, 亟待构建区域尺度气候变化、海平面上升、生态系统演替耦合驱动的盐沼湿地碳循环关键过程模拟预测新范式, 增强盐沼湿地蓝碳增汇潜力评估的可靠性. ...
Global-change controls on soil-carbon accumulation and loss in coastal vegetated ecosystems
1
2019
... (4)气候变化和海平面上升背景下盐沼湿地蓝碳模拟与增汇潜力评估.气候变化和海平面上升改变了盐沼湿地的生物地球化学过程、水文条件以及植被生长和演替, 导致了盐沼湿地蓝碳潜力存在极大的不确定性.气候变化和海平面上升增加滨海湿地碳汇功能(Langley et al., 2009; Wang et al., 2019b, 2021a), 也会通过海岸侵蚀导致更多的碳流失(DeLaune & White, 2012), 但现有模型大多基于气候驱动因素、沉积和碳累积速率等, 缺乏针对生态系统和植被演替过程的考虑.另外, 增温、富营养化和海平面上升对滨海湿地碳储存的影响机制不同(Spivak et al., 2019), 这些因素的改变在极大程度上影响了滨海盐沼湿地的固碳能力, 但目前很难精确地预测这种未知影响.因此, 量化这些因素如何影响盐沼湿地有机碳固存与分解对于构建更具预测性的蓝碳模型至关重要.同时急需构建不同尺度气候变化、海平面上升、生态系统演替耦合驱动的盐沼湿地碳循环关键过程模拟预测新范式, 增强盐沼湿地蓝碳增汇潜力评估的可靠性. ...
Land use in mountain grasslands alters drought response and recovery of carbon allocation and plant-microbial interactions
1
2018
... 植物光合碳分配也是植物采用的一种应对环境胁迫的适应性策略(Stefan et al., 2018; Wang et al., 2021b).例如随着植物可用养分在干旱胁迫环境下变得越来越有限, 植物可能向根部分配更多的光合碳用来吸收水分, 从而引发土壤有机质分解并增加植物养分的可利用性(Williams & de Vries, 2020).同时干旱还可以通过调节根际分泌物的质量和数量等加快或减缓土壤碳循环(Preece et al., 2018).相应地, 滨海湿地生态系统水文过程主要受周期性潮汐作用以及地表淡水与地下咸水之间的相互作用, 导致湿地土壤盐碱化或地表淹水(Han et al., 2018), 也可能改变植被光合碳分配策略.目前, 国内外学者已经使用13C或14C标记对农田、草地和森林等旱地生态系统的光合碳及其后续转化进行了定量分析(Pausch & Kuzyakov, 2018; Wang et al., 2019a).由于盐沼湿地长期或周期性淹水和厌氧, 其植物光合碳分配途径有别于其他生态系统, 因此对旱地生态系统的归纳不一定能够阐明湿地生态系统碳循环的关键过程(Liu et al., 2019).因此, 在全球气候变化背景下, 迫切需要了解盐沼湿地植物-土壤中光合碳分配过程及比例, 这对于理解和精确评估盐沼湿地碳汇功能及构建模型具有重要意义. ...
Plant carbon allocation drives turnover of old soil organic matter in permafrost tundra soils
1
2020
... 植物光合碳是植物-土壤系统碳循环的起点, 解析植物-土壤系统中光合碳的分配过程及机制是估算盐沼湿地固碳功能的先决条件.植物主要通过光合作用将CO2转化为多糖等光合产物, 由叶片通过枝、干向下运输, 分配到植物体各个组织器官.在生态系统碳循环过程中, 植物会通过调节碳在不同部位的分配来满足自身生长与繁殖的需求或应对周边环境变化, 从而间接影响生物地球化学循环(Wang et al., 2019a).气候条件、土壤理化性质、微生物活性和植物生长发育等均会影响植物对光合碳分配的模式, 改变植物向地上、地下部分的碳输入(Diao et al., 2020).此外, 光合碳固定是碳分配的主要驱动力, 环境胁迫还可以通过影响光合作用影响碳吸收, 从而间接影响光同化物的分配(Pausch & Kuzyakov, 2018).目前, 碳同位素技术是研究植物光合碳分配、周转和固定的重要手段.光合碳在植物体各组织器官内的迁移和转化十分迅速, 碳同化后的第1天从叶片到根系的分配就达到峰值(Pausch & Kuzyakov, 2018).同时, 新合成的光合产物可以通过根系分泌物的形式刺激土壤有机质(老碳)的分解, 即土壤碳输入的激发效应(Street et al., 2020).另外, 光合碳分配在物种间也存在差异.Meta分析发现, 与禾本科植物相比, 莎草科植物可以通过根际沉积将更多的碳转移至根和土壤中, 增加地下碳固存(Mou et al., 2018); 采用稳定碳同位素方法, 发现互花米草(Spartina alterniflora)可以将33%和7%的光合同化碳分配到茎和根, 显著高于芦苇(Phragmites australis)的分配比例(Zhang et al., 2017). ...
Nitrogen addition alters photosynthetic carbon fixation, allocation of photoassimilates, and carbon partitioning of Leymus chinensis in a temperate grassland of Inner Mongolia
3
2019a
... 盐沼湿地的蓝碳功能是光合碳吸收、碳沉积埋藏、碳输出等多过程相互作用和平衡的结果(图1), 其关键过程主要包括: (1)植被光合固碳.盐沼湿地通常具有高的净初级生产力, 尤其在河口区, 植被净初级生产力可达1 745 g C·m-2·a-1 (Sousa et al., 2010).(2)光合碳分配.植物通过调节碳在不同部位的分配模式来满足自身生长与繁殖的需求或者应对周边环境变化(Wang et al., 2019a).光合固定的碳向根系和土壤碳库的分配和转移规律, 直接影响着土壤碳库的走向(Xu et al., 2020).盐沼植被根冠比可达1.4-50, 有大量的初级生产力所固定的碳被储存在地下生物量中, 通过根系周转进入土壤碳库(王秀君等, 2016).(3)碳沉积埋藏.由于滨海盐沼湿地不断向下沉积, 土壤碳库很难达到饱和, 储存的碳可以在土壤中保存数千年(Radabaugh et al., 2018).因此, 盐沼湿地的碳封存效率远高于森林或其他陆地生态系统(Wang et al., 2021a).(4)土壤碳流失.潮汐作用导致盐沼湿地间歇性厌氧好氧交替, 氧化还原过程频繁发生, 陆地和海洋微生物交互作用; 同时, 周期性潮汐携带大量的SO42-阻碍CH4产生, 从而降低盐沼湿地碳排放量(Choi & Wang, 2004).另外, 潮汐作用下盐沼湿地可溶性有机碳(DOC)、可溶性无机碳(DIC)、颗粒有机碳(POC)交换及其输出也是其碳流失中不容忽视的组成部分(韩广轩, 2017). ...
... 植物光合碳是植物-土壤系统碳循环的起点, 解析植物-土壤系统中光合碳的分配过程及机制是估算盐沼湿地固碳功能的先决条件.植物主要通过光合作用将CO2转化为多糖等光合产物, 由叶片通过枝、干向下运输, 分配到植物体各个组织器官.在生态系统碳循环过程中, 植物会通过调节碳在不同部位的分配来满足自身生长与繁殖的需求或应对周边环境变化, 从而间接影响生物地球化学循环(Wang et al., 2019a).气候条件、土壤理化性质、微生物活性和植物生长发育等均会影响植物对光合碳分配的模式, 改变植物向地上、地下部分的碳输入(Diao et al., 2020).此外, 光合碳固定是碳分配的主要驱动力, 环境胁迫还可以通过影响光合作用影响碳吸收, 从而间接影响光同化物的分配(Pausch & Kuzyakov, 2018).目前, 碳同位素技术是研究植物光合碳分配、周转和固定的重要手段.光合碳在植物体各组织器官内的迁移和转化十分迅速, 碳同化后的第1天从叶片到根系的分配就达到峰值(Pausch & Kuzyakov, 2018).同时, 新合成的光合产物可以通过根系分泌物的形式刺激土壤有机质(老碳)的分解, 即土壤碳输入的激发效应(Street et al., 2020).另外, 光合碳分配在物种间也存在差异.Meta分析发现, 与禾本科植物相比, 莎草科植物可以通过根际沉积将更多的碳转移至根和土壤中, 增加地下碳固存(Mou et al., 2018); 采用稳定碳同位素方法, 发现互花米草(Spartina alterniflora)可以将33%和7%的光合同化碳分配到茎和根, 显著高于芦苇(Phragmites australis)的分配比例(Zhang et al., 2017). ...
... 植物光合碳分配也是植物采用的一种应对环境胁迫的适应性策略(Stefan et al., 2018; Wang et al., 2021b).例如随着植物可用养分在干旱胁迫环境下变得越来越有限, 植物可能向根部分配更多的光合碳用来吸收水分, 从而引发土壤有机质分解并增加植物养分的可利用性(Williams & de Vries, 2020).同时干旱还可以通过调节根际分泌物的质量和数量等加快或减缓土壤碳循环(Preece et al., 2018).相应地, 滨海湿地生态系统水文过程主要受周期性潮汐作用以及地表淡水与地下咸水之间的相互作用, 导致湿地土壤盐碱化或地表淹水(Han et al., 2018), 也可能改变植被光合碳分配策略.目前, 国内外学者已经使用13C或14C标记对农田、草地和森林等旱地生态系统的光合碳及其后续转化进行了定量分析(Pausch & Kuzyakov, 2018; Wang et al., 2019a).由于盐沼湿地长期或周期性淹水和厌氧, 其植物光合碳分配途径有别于其他生态系统, 因此对旱地生态系统的归纳不一定能够阐明湿地生态系统碳循环的关键过程(Liu et al., 2019).因此, 在全球气候变化背景下, 迫切需要了解盐沼湿地植物-土壤中光合碳分配过程及比例, 这对于理解和精确评估盐沼湿地碳汇功能及构建模型具有重要意义. ...
Tidal wetland resilience to sea level rise increases their carbon sequestration capacity in United States
1
2019b
... (4)气候变化和海平面上升背景下盐沼湿地蓝碳模拟与增汇潜力评估.气候变化和海平面上升改变了盐沼湿地的生物地球化学过程、水文条件以及植被生长和演替, 导致了盐沼湿地蓝碳潜力存在极大的不确定性.气候变化和海平面上升增加滨海湿地碳汇功能(Langley et al., 2009; Wang et al., 2019b, 2021a), 也会通过海岸侵蚀导致更多的碳流失(DeLaune & White, 2012), 但现有模型大多基于气候驱动因素、沉积和碳累积速率等, 缺乏针对生态系统和植被演替过程的考虑.另外, 增温、富营养化和海平面上升对滨海湿地碳储存的影响机制不同(Spivak et al., 2019), 这些因素的改变在极大程度上影响了滨海盐沼湿地的固碳能力, 但目前很难精确地预测这种未知影响.因此, 量化这些因素如何影响盐沼湿地有机碳固存与分解对于构建更具预测性的蓝碳模型至关重要.同时急需构建不同尺度气候变化、海平面上升、生态系统演替耦合驱动的盐沼湿地碳循环关键过程模拟预测新范式, 增强盐沼湿地蓝碳增汇潜力评估的可靠性. ...
Global blue carbon accumulation in tidal wetlands increases with climate change
4
2021a
... 盐沼湿地的蓝碳功能是光合碳吸收、碳沉积埋藏、碳输出等多过程相互作用和平衡的结果(图1), 其关键过程主要包括: (1)植被光合固碳.盐沼湿地通常具有高的净初级生产力, 尤其在河口区, 植被净初级生产力可达1 745 g C·m-2·a-1 (Sousa et al., 2010).(2)光合碳分配.植物通过调节碳在不同部位的分配模式来满足自身生长与繁殖的需求或者应对周边环境变化(Wang et al., 2019a).光合固定的碳向根系和土壤碳库的分配和转移规律, 直接影响着土壤碳库的走向(Xu et al., 2020).盐沼植被根冠比可达1.4-50, 有大量的初级生产力所固定的碳被储存在地下生物量中, 通过根系周转进入土壤碳库(王秀君等, 2016).(3)碳沉积埋藏.由于滨海盐沼湿地不断向下沉积, 土壤碳库很难达到饱和, 储存的碳可以在土壤中保存数千年(Radabaugh et al., 2018).因此, 盐沼湿地的碳封存效率远高于森林或其他陆地生态系统(Wang et al., 2021a).(4)土壤碳流失.潮汐作用导致盐沼湿地间歇性厌氧好氧交替, 氧化还原过程频繁发生, 陆地和海洋微生物交互作用; 同时, 周期性潮汐携带大量的SO42-阻碍CH4产生, 从而降低盐沼湿地碳排放量(Choi & Wang, 2004).另外, 潮汐作用下盐沼湿地可溶性有机碳(DOC)、可溶性无机碳(DIC)、颗粒有机碳(POC)交换及其输出也是其碳流失中不容忽视的组成部分(韩广轩, 2017). ...
... 除了植被光合固碳外, 滨海盐沼湿地还具有较高的碳沉积埋藏速率, 成为盐沼湿地蓝碳功能的重要组成部分, 主要包括内源有机碳和外源有机碳.解析沉积物中碳的埋藏特征及来源过程是认知盐沼湿地固碳功能的基础和重点, 也是当前滨海湿地碳循环研究的热点和难点.沉积到滨海湿地的有机质有一部分经微生物降解和矿化作用生成CO2和CH4, 而另一部分将被埋藏在沉积物中, 成为滨海湿地固碳功能的重要组成部分(Xia et al., 2021a).滨海湿地的碳埋藏主要有两个来源, 首先是内源有机碳, 主要指湿地植被凋落物的累积; 其次是外源有机碳, 主要是潮汐淹没过程中海水中悬浮颗粒物有机碳的沉积, 包括海水中各种藻类和其他生物死亡残体以及从河流上游运输而来的各类颗粒有机质(Xiong et al., 2018).滨海湿地的碳埋藏速率因不同植被类型而变化较大, 同时受海平面上升速率的影响.最新的数据估算了全球尺度上盐沼湿地和红树林的碳埋藏速率约为53.65 Tg·a-1, 换算成CO2当量为196.71 Tg·a-1 (Wang et al., 2021a), 相当于人类活动每年排放量的0.6%.如果从单位面积碳埋藏速率估算, 滨海湿地蓝碳生态系统的碳埋藏速率是陆地生态系统固碳速率的15倍, 是海洋生态系统固碳速率的50倍左右, 显示出极强的碳汇功能.但是在河口地区, 外源有机碳会贡献更多的埋藏碳.在我国黄河口以及长江口的滨海湿地的沉积速率远超其他非河口地区(Ye et al., 2018), 其主要原因是河水的搬移作用携带了大量的上游颗粒物质在河口地区沉积下来, 因此, 这些河水中的颗粒物贡献了河口地区滨海湿地土壤有机碳的主要部分. ...
... (2)盐沼湿地土壤有机碳沉积和埋藏速率及其对全球变化的响应.盐沼湿地具有很高的碳捕获能力和长期持续固碳能力, 特别是在河流携带大量泥沙和营养元素的河口区, 其储碳机理主要是在沉积物厌氧环境对有机质分解的抑制作用下, 大量植物残体能够被长期地保存(Mcleod et al., 2011; 王秀君等, 2016).盐沼湿地和红树林的碳埋藏速率约为53.65 Tg·a-1, 换算成CO2当量为196.71 Tg·a-1 (Wang et al., 2021a), 这一数据相当于人类活动每年排放量的0.6% (王法明等, 2021).在未来气候变化和海平面上升情境下, 盐沼湿地蓝碳的沉积和埋藏潜力如何? ...
... (4)气候变化和海平面上升背景下盐沼湿地蓝碳模拟与增汇潜力评估.气候变化和海平面上升改变了盐沼湿地的生物地球化学过程、水文条件以及植被生长和演替, 导致了盐沼湿地蓝碳潜力存在极大的不确定性.气候变化和海平面上升增加滨海湿地碳汇功能(Langley et al., 2009; Wang et al., 2019b, 2021a), 也会通过海岸侵蚀导致更多的碳流失(DeLaune & White, 2012), 但现有模型大多基于气候驱动因素、沉积和碳累积速率等, 缺乏针对生态系统和植被演替过程的考虑.另外, 增温、富营养化和海平面上升对滨海湿地碳储存的影响机制不同(Spivak et al., 2019), 这些因素的改变在极大程度上影响了滨海盐沼湿地的固碳能力, 但目前很难精确地预测这种未知影响.因此, 量化这些因素如何影响盐沼湿地有机碳固存与分解对于构建更具预测性的蓝碳模型至关重要.同时急需构建不同尺度气候变化、海平面上升、生态系统演替耦合驱动的盐沼湿地碳循环关键过程模拟预测新范式, 增强盐沼湿地蓝碳增汇潜力评估的可靠性. ...
中国滨海湿地的蓝色碳汇功能及碳中和对策
1
2021
... (2)盐沼湿地土壤有机碳沉积和埋藏速率及其对全球变化的响应.盐沼湿地具有很高的碳捕获能力和长期持续固碳能力, 特别是在河流携带大量泥沙和营养元素的河口区, 其储碳机理主要是在沉积物厌氧环境对有机质分解的抑制作用下, 大量植物残体能够被长期地保存(Mcleod et al., 2011; 王秀君等, 2016).盐沼湿地和红树林的碳埋藏速率约为53.65 Tg·a-1, 换算成CO2当量为196.71 Tg·a-1 (Wang et al., 2021a), 这一数据相当于人类活动每年排放量的0.6% (王法明等, 2021).在未来气候变化和海平面上升情境下, 盐沼湿地蓝碳的沉积和埋藏潜力如何? ...
中国滨海湿地的蓝色碳汇功能及碳中和对策
1
2021
... (2)盐沼湿地土壤有机碳沉积和埋藏速率及其对全球变化的响应.盐沼湿地具有很高的碳捕获能力和长期持续固碳能力, 特别是在河流携带大量泥沙和营养元素的河口区, 其储碳机理主要是在沉积物厌氧环境对有机质分解的抑制作用下, 大量植物残体能够被长期地保存(Mcleod et al., 2011; 王秀君等, 2016).盐沼湿地和红树林的碳埋藏速率约为53.65 Tg·a-1, 换算成CO2当量为196.71 Tg·a-1 (Wang et al., 2021a), 这一数据相当于人类活动每年排放量的0.6% (王法明等, 2021).在未来气候变化和海平面上升情境下, 盐沼湿地蓝碳的沉积和埋藏潜力如何? ...
A novel 13C pulse-labelling method to quantify the contribution of rhizodeposits to soil respiration in a grassland exposed to drought and nitrogen addition
1
2021b
... 植物光合碳分配也是植物采用的一种应对环境胁迫的适应性策略(Stefan et al., 2018; Wang et al., 2021b).例如随着植物可用养分在干旱胁迫环境下变得越来越有限, 植物可能向根部分配更多的光合碳用来吸收水分, 从而引发土壤有机质分解并增加植物养分的可利用性(Williams & de Vries, 2020).同时干旱还可以通过调节根际分泌物的质量和数量等加快或减缓土壤碳循环(Preece et al., 2018).相应地, 滨海湿地生态系统水文过程主要受周期性潮汐作用以及地表淡水与地下咸水之间的相互作用, 导致湿地土壤盐碱化或地表淹水(Han et al., 2018), 也可能改变植被光合碳分配策略.目前, 国内外学者已经使用13C或14C标记对农田、草地和森林等旱地生态系统的光合碳及其后续转化进行了定量分析(Pausch & Kuzyakov, 2018; Wang et al., 2019a).由于盐沼湿地长期或周期性淹水和厌氧, 其植物光合碳分配途径有别于其他生态系统, 因此对旱地生态系统的归纳不一定能够阐明湿地生态系统碳循环的关键过程(Liu et al., 2019).因此, 在全球气候变化背景下, 迫切需要了解盐沼湿地植物-土壤中光合碳分配过程及比例, 这对于理解和精确评估盐沼湿地碳汇功能及构建模型具有重要意义. ...
中国海岸带及近海碳循环与蓝碳潜力
3
2016
... 盐沼湿地的蓝碳功能是光合碳吸收、碳沉积埋藏、碳输出等多过程相互作用和平衡的结果(图1), 其关键过程主要包括: (1)植被光合固碳.盐沼湿地通常具有高的净初级生产力, 尤其在河口区, 植被净初级生产力可达1 745 g C·m-2·a-1 (Sousa et al., 2010).(2)光合碳分配.植物通过调节碳在不同部位的分配模式来满足自身生长与繁殖的需求或者应对周边环境变化(Wang et al., 2019a).光合固定的碳向根系和土壤碳库的分配和转移规律, 直接影响着土壤碳库的走向(Xu et al., 2020).盐沼植被根冠比可达1.4-50, 有大量的初级生产力所固定的碳被储存在地下生物量中, 通过根系周转进入土壤碳库(王秀君等, 2016).(3)碳沉积埋藏.由于滨海盐沼湿地不断向下沉积, 土壤碳库很难达到饱和, 储存的碳可以在土壤中保存数千年(Radabaugh et al., 2018).因此, 盐沼湿地的碳封存效率远高于森林或其他陆地生态系统(Wang et al., 2021a).(4)土壤碳流失.潮汐作用导致盐沼湿地间歇性厌氧好氧交替, 氧化还原过程频繁发生, 陆地和海洋微生物交互作用; 同时, 周期性潮汐携带大量的SO42-阻碍CH4产生, 从而降低盐沼湿地碳排放量(Choi & Wang, 2004).另外, 潮汐作用下盐沼湿地可溶性有机碳(DOC)、可溶性无机碳(DIC)、颗粒有机碳(POC)交换及其输出也是其碳流失中不容忽视的组成部分(韩广轩, 2017). ...
... 综上所述, 盐沼等滨海湿地蓝碳潜力巨大, 但是其固碳的关键机理尚不清楚, 碳汇通量、封存量、增汇潜力的数据还很少, 目前尚无一个全球公认的机制来正确认知蓝碳的重要性(王秀君等, 2016; 周晨昊等, 2016).同时, 以往的研究多关注单一过程对盐沼湿地固碳功能的影响, 较少考虑多过程的系统影响及其相对贡献, 缺乏综合性机理探讨.另外, 气候变化和海平面上升背景下盐沼湿地蓝碳的增汇潜力如何, 目前尚无定论, 这在很大程度上制约着对盐沼湿地碳汇功能的整体理解和准确评估. ...
... (2)盐沼湿地土壤有机碳沉积和埋藏速率及其对全球变化的响应.盐沼湿地具有很高的碳捕获能力和长期持续固碳能力, 特别是在河流携带大量泥沙和营养元素的河口区, 其储碳机理主要是在沉积物厌氧环境对有机质分解的抑制作用下, 大量植物残体能够被长期地保存(Mcleod et al., 2011; 王秀君等, 2016).盐沼湿地和红树林的碳埋藏速率约为53.65 Tg·a-1, 换算成CO2当量为196.71 Tg·a-1 (Wang et al., 2021a), 这一数据相当于人类活动每年排放量的0.6% (王法明等, 2021).在未来气候变化和海平面上升情境下, 盐沼湿地蓝碳的沉积和埋藏潜力如何? ...
中国海岸带及近海碳循环与蓝碳潜力
3
2016
... 盐沼湿地的蓝碳功能是光合碳吸收、碳沉积埋藏、碳输出等多过程相互作用和平衡的结果(图1), 其关键过程主要包括: (1)植被光合固碳.盐沼湿地通常具有高的净初级生产力, 尤其在河口区, 植被净初级生产力可达1 745 g C·m-2·a-1 (Sousa et al., 2010).(2)光合碳分配.植物通过调节碳在不同部位的分配模式来满足自身生长与繁殖的需求或者应对周边环境变化(Wang et al., 2019a).光合固定的碳向根系和土壤碳库的分配和转移规律, 直接影响着土壤碳库的走向(Xu et al., 2020).盐沼植被根冠比可达1.4-50, 有大量的初级生产力所固定的碳被储存在地下生物量中, 通过根系周转进入土壤碳库(王秀君等, 2016).(3)碳沉积埋藏.由于滨海盐沼湿地不断向下沉积, 土壤碳库很难达到饱和, 储存的碳可以在土壤中保存数千年(Radabaugh et al., 2018).因此, 盐沼湿地的碳封存效率远高于森林或其他陆地生态系统(Wang et al., 2021a).(4)土壤碳流失.潮汐作用导致盐沼湿地间歇性厌氧好氧交替, 氧化还原过程频繁发生, 陆地和海洋微生物交互作用; 同时, 周期性潮汐携带大量的SO42-阻碍CH4产生, 从而降低盐沼湿地碳排放量(Choi & Wang, 2004).另外, 潮汐作用下盐沼湿地可溶性有机碳(DOC)、可溶性无机碳(DIC)、颗粒有机碳(POC)交换及其输出也是其碳流失中不容忽视的组成部分(韩广轩, 2017). ...
... 综上所述, 盐沼等滨海湿地蓝碳潜力巨大, 但是其固碳的关键机理尚不清楚, 碳汇通量、封存量、增汇潜力的数据还很少, 目前尚无一个全球公认的机制来正确认知蓝碳的重要性(王秀君等, 2016; 周晨昊等, 2016).同时, 以往的研究多关注单一过程对盐沼湿地固碳功能的影响, 较少考虑多过程的系统影响及其相对贡献, 缺乏综合性机理探讨.另外, 气候变化和海平面上升背景下盐沼湿地蓝碳的增汇潜力如何, 目前尚无定论, 这在很大程度上制约着对盐沼湿地碳汇功能的整体理解和准确评估. ...
... (2)盐沼湿地土壤有机碳沉积和埋藏速率及其对全球变化的响应.盐沼湿地具有很高的碳捕获能力和长期持续固碳能力, 特别是在河流携带大量泥沙和营养元素的河口区, 其储碳机理主要是在沉积物厌氧环境对有机质分解的抑制作用下, 大量植物残体能够被长期地保存(Mcleod et al., 2011; 王秀君等, 2016).盐沼湿地和红树林的碳埋藏速率约为53.65 Tg·a-1, 换算成CO2当量为196.71 Tg·a-1 (Wang et al., 2021a), 这一数据相当于人类活动每年排放量的0.6% (王法明等, 2021).在未来气候变化和海平面上升情境下, 盐沼湿地蓝碳的沉积和埋藏潜力如何? ...
Plant root exudation under drought: implications for ecosystem functioning
1
2020
... 植物光合碳分配也是植物采用的一种应对环境胁迫的适应性策略(Stefan et al., 2018; Wang et al., 2021b).例如随着植物可用养分在干旱胁迫环境下变得越来越有限, 植物可能向根部分配更多的光合碳用来吸收水分, 从而引发土壤有机质分解并增加植物养分的可利用性(Williams & de Vries, 2020).同时干旱还可以通过调节根际分泌物的质量和数量等加快或减缓土壤碳循环(Preece et al., 2018).相应地, 滨海湿地生态系统水文过程主要受周期性潮汐作用以及地表淡水与地下咸水之间的相互作用, 导致湿地土壤盐碱化或地表淹水(Han et al., 2018), 也可能改变植被光合碳分配策略.目前, 国内外学者已经使用13C或14C标记对农田、草地和森林等旱地生态系统的光合碳及其后续转化进行了定量分析(Pausch & Kuzyakov, 2018; Wang et al., 2019a).由于盐沼湿地长期或周期性淹水和厌氧, 其植物光合碳分配途径有别于其他生态系统, 因此对旱地生态系统的归纳不一定能够阐明湿地生态系统碳循环的关键过程(Liu et al., 2019).因此, 在全球气候变化背景下, 迫切需要了解盐沼湿地植物-土壤中光合碳分配过程及比例, 这对于理解和精确评估盐沼湿地碳汇功能及构建模型具有重要意义. ...
Sedimentary records of mangrove evolution during the past one hundred years based on stable carbon isotope and pollen evidences in Maowei, SW China
1
2016
... 盐沼湿地碳埋藏速率通常通过沉积速率与沉积物碳密度计算而来.沉积物中放射性核素的含量被广泛应用于河口、海岸及陆架区域沉积物年龄的确定, 沉积过程的揭示和物质来源判断等.常用的天然核素为210Pb, 其半衰期为22.3 a, 定年上限为150 a.除天然核素外, 人类核试验等活动还产生了137Cs等人工核素, 137Cs半衰期为30.17 a, 在沉积物垂直分布中通常会出现峰值, 对应特定的核爆年份, 具有重要的纪年意义(Xia et al., 2016).沉积速率研究中一般将137Cs与210Pb纪年结果相互印证(Peck et al., 2020).例如, Ye等(2018)利用210Pb纪年方法研究了黄河三角洲不同植被类型湿地的沉积速率及碳埋藏速率, 发现黄河三角洲盐沼湿地的沉积速率在3.87-9.68 cm·a-1, 而碳埋藏速率则在586-1 784 g·m-2·a-1, 远高于全球盐沼湿地固碳速率的平均值(168 g·m-2·a-1), 显示出黄河三角洲滨海湿地极高的碳埋藏能力. ...
Distribution, sources, and decomposition of soil organic matter along a salinity gradient in estuarine wetlands characterized by C:N ratio, δ13C-δ15N, and lignin biomarker
2
2021a
... 除了植被光合固碳外, 滨海盐沼湿地还具有较高的碳沉积埋藏速率, 成为盐沼湿地蓝碳功能的重要组成部分, 主要包括内源有机碳和外源有机碳.解析沉积物中碳的埋藏特征及来源过程是认知盐沼湿地固碳功能的基础和重点, 也是当前滨海湿地碳循环研究的热点和难点.沉积到滨海湿地的有机质有一部分经微生物降解和矿化作用生成CO2和CH4, 而另一部分将被埋藏在沉积物中, 成为滨海湿地固碳功能的重要组成部分(Xia et al., 2021a).滨海湿地的碳埋藏主要有两个来源, 首先是内源有机碳, 主要指湿地植被凋落物的累积; 其次是外源有机碳, 主要是潮汐淹没过程中海水中悬浮颗粒物有机碳的沉积, 包括海水中各种藻类和其他生物死亡残体以及从河流上游运输而来的各类颗粒有机质(Xiong et al., 2018).滨海湿地的碳埋藏速率因不同植被类型而变化较大, 同时受海平面上升速率的影响.最新的数据估算了全球尺度上盐沼湿地和红树林的碳埋藏速率约为53.65 Tg·a-1, 换算成CO2当量为196.71 Tg·a-1 (Wang et al., 2021a), 相当于人类活动每年排放量的0.6%.如果从单位面积碳埋藏速率估算, 滨海湿地蓝碳生态系统的碳埋藏速率是陆地生态系统固碳速率的15倍, 是海洋生态系统固碳速率的50倍左右, 显示出极强的碳汇功能.但是在河口地区, 外源有机碳会贡献更多的埋藏碳.在我国黄河口以及长江口的滨海湿地的沉积速率远超其他非河口地区(Ye et al., 2018), 其主要原因是河水的搬移作用携带了大量的上游颗粒物质在河口地区沉积下来, 因此, 这些河水中的颗粒物贡献了河口地区滨海湿地土壤有机碳的主要部分. ...
... 解析滨海湿地埋藏碳的来源所使用的方法主要是碳氮比值法(C:N)、稳定同位素组成法和生物标记物法(Xia et al., 2021a).综合利用碳、氮稳定性同位素丰度(δ)可以解析湿地沉积物有机质的来源(Xia et al., 2021b).通过对沉积物C:N的测定可以区分其碳的来源是水生植被还是陆生植被, 水生植物C:N一般在4-10之间, 而陆生维管植物和挺水植物C:N一般大于20.同时, 沉积物有机质中δ13C和δ15N与有机质来源紧密相关, 通过对滨海湿地地上植被和周围水体主要植物体δ13C的测定可以区分其沉积物中内源有机碳的来源(黄梅等, 2018).除了δ13C, δ15N也被经常用于解析有机质的来源.典型陆地植物的δ15N变化范围为0-11‰, 海草δ15N变化范围为6‰-12‰, 而海洋中藻类的δ15N变化范围为0-4‰ (Samper- Villarreal et al., 2016).另外, 生物标志物是环境和地质体中记载着原始生物母质分子结构信息的有机化合物, 被广泛用于研究埋藏碳的来源和转化特征(冯晓娟等, 2020).例如, 正构烷烃广泛分布于低等细菌藻类与高等植物等不同类型的生物体的蜡质脂类中, 来源于不同生物体的正构烷烃, 其碳数组成与分布特征不同(冯晓娟等, 2020), 可以用来探究沉积物有机质内源菌藻、水生植物、草本植物与木本植物的相对贡献.氨基糖在土壤中更稳定而且保存更长时间(Joergensen, 2018), 是土壤中微生物残体碳的主要标记物, 被广泛应用于研究微生物残体对土壤有机碳和各种微环境流通量的贡献(Joergensen, 2018).综合运用C:N法、稳定同位素组成法和生物标记物法可以有效地解析湿地沉积物中有机碳的来源及其转化过程, 是当前湿地碳循环研究的热点方向(冯晓娟等, 2020; Xia et al., 2021b; Yuan et al., 2021). ...
Spartina alterniflora invasion controls organic carbon stocks in coastal marsh and mangrove soils across tropics and subtropics
2
2021b
... 解析滨海湿地埋藏碳的来源所使用的方法主要是碳氮比值法(C:N)、稳定同位素组成法和生物标记物法(Xia et al., 2021a).综合利用碳、氮稳定性同位素丰度(δ)可以解析湿地沉积物有机质的来源(Xia et al., 2021b).通过对沉积物C:N的测定可以区分其碳的来源是水生植被还是陆生植被, 水生植物C:N一般在4-10之间, 而陆生维管植物和挺水植物C:N一般大于20.同时, 沉积物有机质中δ13C和δ15N与有机质来源紧密相关, 通过对滨海湿地地上植被和周围水体主要植物体δ13C的测定可以区分其沉积物中内源有机碳的来源(黄梅等, 2018).除了δ13C, δ15N也被经常用于解析有机质的来源.典型陆地植物的δ15N变化范围为0-11‰, 海草δ15N变化范围为6‰-12‰, 而海洋中藻类的δ15N变化范围为0-4‰ (Samper- Villarreal et al., 2016).另外, 生物标志物是环境和地质体中记载着原始生物母质分子结构信息的有机化合物, 被广泛用于研究埋藏碳的来源和转化特征(冯晓娟等, 2020).例如, 正构烷烃广泛分布于低等细菌藻类与高等植物等不同类型的生物体的蜡质脂类中, 来源于不同生物体的正构烷烃, 其碳数组成与分布特征不同(冯晓娟等, 2020), 可以用来探究沉积物有机质内源菌藻、水生植物、草本植物与木本植物的相对贡献.氨基糖在土壤中更稳定而且保存更长时间(Joergensen, 2018), 是土壤中微生物残体碳的主要标记物, 被广泛应用于研究微生物残体对土壤有机碳和各种微环境流通量的贡献(Joergensen, 2018).综合运用C:N法、稳定同位素组成法和生物标记物法可以有效地解析湿地沉积物中有机碳的来源及其转化过程, 是当前湿地碳循环研究的热点方向(冯晓娟等, 2020; Xia et al., 2021b; Yuan et al., 2021). ...
... ., 2021b; Yuan et al., 2021). ...
Mangrove vegetation enhances soil carbon storage primarily through in situ inputs rather than increasing allochthonous sediments
1
2018
... 除了植被光合固碳外, 滨海盐沼湿地还具有较高的碳沉积埋藏速率, 成为盐沼湿地蓝碳功能的重要组成部分, 主要包括内源有机碳和外源有机碳.解析沉积物中碳的埋藏特征及来源过程是认知盐沼湿地固碳功能的基础和重点, 也是当前滨海湿地碳循环研究的热点和难点.沉积到滨海湿地的有机质有一部分经微生物降解和矿化作用生成CO2和CH4, 而另一部分将被埋藏在沉积物中, 成为滨海湿地固碳功能的重要组成部分(Xia et al., 2021a).滨海湿地的碳埋藏主要有两个来源, 首先是内源有机碳, 主要指湿地植被凋落物的累积; 其次是外源有机碳, 主要是潮汐淹没过程中海水中悬浮颗粒物有机碳的沉积, 包括海水中各种藻类和其他生物死亡残体以及从河流上游运输而来的各类颗粒有机质(Xiong et al., 2018).滨海湿地的碳埋藏速率因不同植被类型而变化较大, 同时受海平面上升速率的影响.最新的数据估算了全球尺度上盐沼湿地和红树林的碳埋藏速率约为53.65 Tg·a-1, 换算成CO2当量为196.71 Tg·a-1 (Wang et al., 2021a), 相当于人类活动每年排放量的0.6%.如果从单位面积碳埋藏速率估算, 滨海湿地蓝碳生态系统的碳埋藏速率是陆地生态系统固碳速率的15倍, 是海洋生态系统固碳速率的50倍左右, 显示出极强的碳汇功能.但是在河口地区, 外源有机碳会贡献更多的埋藏碳.在我国黄河口以及长江口的滨海湿地的沉积速率远超其他非河口地区(Ye et al., 2018), 其主要原因是河水的搬移作用携带了大量的上游颗粒物质在河口地区沉积下来, 因此, 这些河水中的颗粒物贡献了河口地区滨海湿地土壤有机碳的主要部分. ...
Human eutrophication drives biogeographic salt marsh productivity patterns in China
1
2020
... 盐沼湿地的蓝碳功能是光合碳吸收、碳沉积埋藏、碳输出等多过程相互作用和平衡的结果(图1), 其关键过程主要包括: (1)植被光合固碳.盐沼湿地通常具有高的净初级生产力, 尤其在河口区, 植被净初级生产力可达1 745 g C·m-2·a-1 (Sousa et al., 2010).(2)光合碳分配.植物通过调节碳在不同部位的分配模式来满足自身生长与繁殖的需求或者应对周边环境变化(Wang et al., 2019a).光合固定的碳向根系和土壤碳库的分配和转移规律, 直接影响着土壤碳库的走向(Xu et al., 2020).盐沼植被根冠比可达1.4-50, 有大量的初级生产力所固定的碳被储存在地下生物量中, 通过根系周转进入土壤碳库(王秀君等, 2016).(3)碳沉积埋藏.由于滨海盐沼湿地不断向下沉积, 土壤碳库很难达到饱和, 储存的碳可以在土壤中保存数千年(Radabaugh et al., 2018).因此, 盐沼湿地的碳封存效率远高于森林或其他陆地生态系统(Wang et al., 2021a).(4)土壤碳流失.潮汐作用导致盐沼湿地间歇性厌氧好氧交替, 氧化还原过程频繁发生, 陆地和海洋微生物交互作用; 同时, 周期性潮汐携带大量的SO42-阻碍CH4产生, 从而降低盐沼湿地碳排放量(Choi & Wang, 2004).另外, 潮汐作用下盐沼湿地可溶性有机碳(DOC)、可溶性无机碳(DIC)、颗粒有机碳(POC)交换及其输出也是其碳流失中不容忽视的组成部分(韩广轩, 2017). ...
Carbon sequestration and its controlling factors in the temperate wetland communities along the Bohai Sea, China
2
2018
... 除了植被光合固碳外, 滨海盐沼湿地还具有较高的碳沉积埋藏速率, 成为盐沼湿地蓝碳功能的重要组成部分, 主要包括内源有机碳和外源有机碳.解析沉积物中碳的埋藏特征及来源过程是认知盐沼湿地固碳功能的基础和重点, 也是当前滨海湿地碳循环研究的热点和难点.沉积到滨海湿地的有机质有一部分经微生物降解和矿化作用生成CO2和CH4, 而另一部分将被埋藏在沉积物中, 成为滨海湿地固碳功能的重要组成部分(Xia et al., 2021a).滨海湿地的碳埋藏主要有两个来源, 首先是内源有机碳, 主要指湿地植被凋落物的累积; 其次是外源有机碳, 主要是潮汐淹没过程中海水中悬浮颗粒物有机碳的沉积, 包括海水中各种藻类和其他生物死亡残体以及从河流上游运输而来的各类颗粒有机质(Xiong et al., 2018).滨海湿地的碳埋藏速率因不同植被类型而变化较大, 同时受海平面上升速率的影响.最新的数据估算了全球尺度上盐沼湿地和红树林的碳埋藏速率约为53.65 Tg·a-1, 换算成CO2当量为196.71 Tg·a-1 (Wang et al., 2021a), 相当于人类活动每年排放量的0.6%.如果从单位面积碳埋藏速率估算, 滨海湿地蓝碳生态系统的碳埋藏速率是陆地生态系统固碳速率的15倍, 是海洋生态系统固碳速率的50倍左右, 显示出极强的碳汇功能.但是在河口地区, 外源有机碳会贡献更多的埋藏碳.在我国黄河口以及长江口的滨海湿地的沉积速率远超其他非河口地区(Ye et al., 2018), 其主要原因是河水的搬移作用携带了大量的上游颗粒物质在河口地区沉积下来, 因此, 这些河水中的颗粒物贡献了河口地区滨海湿地土壤有机碳的主要部分. ...
... 盐沼湿地碳埋藏速率通常通过沉积速率与沉积物碳密度计算而来.沉积物中放射性核素的含量被广泛应用于河口、海岸及陆架区域沉积物年龄的确定, 沉积过程的揭示和物质来源判断等.常用的天然核素为210Pb, 其半衰期为22.3 a, 定年上限为150 a.除天然核素外, 人类核试验等活动还产生了137Cs等人工核素, 137Cs半衰期为30.17 a, 在沉积物垂直分布中通常会出现峰值, 对应特定的核爆年份, 具有重要的纪年意义(Xia et al., 2016).沉积速率研究中一般将137Cs与210Pb纪年结果相互印证(Peck et al., 2020).例如, Ye等(2018)利用210Pb纪年方法研究了黄河三角洲不同植被类型湿地的沉积速率及碳埋藏速率, 发现黄河三角洲盐沼湿地的沉积速率在3.87-9.68 cm·a-1, 而碳埋藏速率则在586-1 784 g·m-2·a-1, 远高于全球盐沼湿地固碳速率的平均值(168 g·m-2·a-1), 显示出黄河三角洲滨海湿地极高的碳埋藏能力. ...
Phosphorus addition decreases microbial residual contribution to soil organic carbon pool in a tropical coastal forest
1
2021
... 解析滨海湿地埋藏碳的来源所使用的方法主要是碳氮比值法(C:N)、稳定同位素组成法和生物标记物法(Xia et al., 2021a).综合利用碳、氮稳定性同位素丰度(δ)可以解析湿地沉积物有机质的来源(Xia et al., 2021b).通过对沉积物C:N的测定可以区分其碳的来源是水生植被还是陆生植被, 水生植物C:N一般在4-10之间, 而陆生维管植物和挺水植物C:N一般大于20.同时, 沉积物有机质中δ13C和δ15N与有机质来源紧密相关, 通过对滨海湿地地上植被和周围水体主要植物体δ13C的测定可以区分其沉积物中内源有机碳的来源(黄梅等, 2018).除了δ13C, δ15N也被经常用于解析有机质的来源.典型陆地植物的δ15N变化范围为0-11‰, 海草δ15N变化范围为6‰-12‰, 而海洋中藻类的δ15N变化范围为0-4‰ (Samper- Villarreal et al., 2016).另外, 生物标志物是环境和地质体中记载着原始生物母质分子结构信息的有机化合物, 被广泛用于研究埋藏碳的来源和转化特征(冯晓娟等, 2020).例如, 正构烷烃广泛分布于低等细菌藻类与高等植物等不同类型的生物体的蜡质脂类中, 来源于不同生物体的正构烷烃, 其碳数组成与分布特征不同(冯晓娟等, 2020), 可以用来探究沉积物有机质内源菌藻、水生植物、草本植物与木本植物的相对贡献.氨基糖在土壤中更稳定而且保存更长时间(Joergensen, 2018), 是土壤中微生物残体碳的主要标记物, 被广泛应用于研究微生物残体对土壤有机碳和各种微环境流通量的贡献(Joergensen, 2018).综合运用C:N法、稳定同位素组成法和生物标记物法可以有效地解析湿地沉积物中有机碳的来源及其转化过程, 是当前湿地碳循环研究的热点方向(冯晓娟等, 2020; Xia et al., 2021b; Yuan et al., 2021). ...
The transfer and allocation of newly fixed C by invasive Spartina alterniflora and native Phragmites australis to soil microbiota
1
2017
... 植物光合碳是植物-土壤系统碳循环的起点, 解析植物-土壤系统中光合碳的分配过程及机制是估算盐沼湿地固碳功能的先决条件.植物主要通过光合作用将CO2转化为多糖等光合产物, 由叶片通过枝、干向下运输, 分配到植物体各个组织器官.在生态系统碳循环过程中, 植物会通过调节碳在不同部位的分配来满足自身生长与繁殖的需求或应对周边环境变化, 从而间接影响生物地球化学循环(Wang et al., 2019a).气候条件、土壤理化性质、微生物活性和植物生长发育等均会影响植物对光合碳分配的模式, 改变植物向地上、地下部分的碳输入(Diao et al., 2020).此外, 光合碳固定是碳分配的主要驱动力, 环境胁迫还可以通过影响光合作用影响碳吸收, 从而间接影响光同化物的分配(Pausch & Kuzyakov, 2018).目前, 碳同位素技术是研究植物光合碳分配、周转和固定的重要手段.光合碳在植物体各组织器官内的迁移和转化十分迅速, 碳同化后的第1天从叶片到根系的分配就达到峰值(Pausch & Kuzyakov, 2018).同时, 新合成的光合产物可以通过根系分泌物的形式刺激土壤有机质(老碳)的分解, 即土壤碳输入的激发效应(Street et al., 2020).另外, 光合碳分配在物种间也存在差异.Meta分析发现, 与禾本科植物相比, 莎草科植物可以通过根际沉积将更多的碳转移至根和土壤中, 增加地下碳固存(Mou et al., 2018); 采用稳定碳同位素方法, 发现互花米草(Spartina alterniflora)可以将33%和7%的光合同化碳分配到茎和根, 显著高于芦苇(Phragmites australis)的分配比例(Zhang et al., 2017). ...
中国海岸带蓝碳生态系统碳汇潜力的初步分析
2
2016
... 全球滨海湿地面积约为20.3 × 104 km2, 我国滨海湿地面积约占全球滨海湿地总面积的1/4, 其中滨海盐沼湿地是我国最主要的滨海湿地类型(曹磊等, 2013).据估算, 我国蓝碳生态系统的碳年埋藏量为0.349-0.835 Tg, 其中盐沼湿地约占80%, 远高于红树林和海草床, 是我国蓝碳碳汇的主要贡献者(周晨昊等, 2016).模型模拟结果表明, 未来气候变暖和海平面上升可能使得盐沼湿地能够更迅速捕获和埋藏大气中的碳(Kirwan & Mudd, 2012).另外, 加强全球滨海湿地的保护和修复, 每年能够吸收、储存和减少0.32-0.89 Tg CO2排放(Hoegh-Guldberg et al., 2019).2010年, 保护国际基金会、世界自然保护联盟、联合国教科文组织政府间海洋学委员会共同发起“蓝碳倡议” (The Blue Carbon Initiative), 旨在支撑全球蓝碳的科学研究、项目实施和政策制定等, 通过恢复和保护受损蓝碳生态系统工作而产生碳效益, 减缓气候变化. ...
... 综上所述, 盐沼等滨海湿地蓝碳潜力巨大, 但是其固碳的关键机理尚不清楚, 碳汇通量、封存量、增汇潜力的数据还很少, 目前尚无一个全球公认的机制来正确认知蓝碳的重要性(王秀君等, 2016; 周晨昊等, 2016).同时, 以往的研究多关注单一过程对盐沼湿地固碳功能的影响, 较少考虑多过程的系统影响及其相对贡献, 缺乏综合性机理探讨.另外, 气候变化和海平面上升背景下盐沼湿地蓝碳的增汇潜力如何, 目前尚无定论, 这在很大程度上制约着对盐沼湿地碳汇功能的整体理解和准确评估. ...
中国海岸带蓝碳生态系统碳汇潜力的初步分析
2
2016
... 全球滨海湿地面积约为20.3 × 104 km2, 我国滨海湿地面积约占全球滨海湿地总面积的1/4, 其中滨海盐沼湿地是我国最主要的滨海湿地类型(曹磊等, 2013).据估算, 我国蓝碳生态系统的碳年埋藏量为0.349-0.835 Tg, 其中盐沼湿地约占80%, 远高于红树林和海草床, 是我国蓝碳碳汇的主要贡献者(周晨昊等, 2016).模型模拟结果表明, 未来气候变暖和海平面上升可能使得盐沼湿地能够更迅速捕获和埋藏大气中的碳(Kirwan & Mudd, 2012).另外, 加强全球滨海湿地的保护和修复, 每年能够吸收、储存和减少0.32-0.89 Tg CO2排放(Hoegh-Guldberg et al., 2019).2010年, 保护国际基金会、世界自然保护联盟、联合国教科文组织政府间海洋学委员会共同发起“蓝碳倡议” (The Blue Carbon Initiative), 旨在支撑全球蓝碳的科学研究、项目实施和政策制定等, 通过恢复和保护受损蓝碳生态系统工作而产生碳效益, 减缓气候变化. ...
... 综上所述, 盐沼等滨海湿地蓝碳潜力巨大, 但是其固碳的关键机理尚不清楚, 碳汇通量、封存量、增汇潜力的数据还很少, 目前尚无一个全球公认的机制来正确认知蓝碳的重要性(王秀君等, 2016; 周晨昊等, 2016).同时, 以往的研究多关注单一过程对盐沼湿地固碳功能的影响, 较少考虑多过程的系统影响及其相对贡献, 缺乏综合性机理探讨.另外, 气候变化和海平面上升背景下盐沼湿地蓝碳的增汇潜力如何, 目前尚无定论, 这在很大程度上制约着对盐沼湿地碳汇功能的整体理解和准确评估. ...






{kind=link}
{kind=link}
{kind=link}
{kind=link}