


种群是指在一定时间内占据一定空间的同种生物个体的集合, 是物种生存、发展和进化的基本单位, 也是连接生物个体、群落和生态系统的关键纽带(赵阳等, 2020)。种群结构与动态是种群生态学研究的两个核心内容(Helt & Loucks, 1976; Chhetri et al., 2016), 前者是指种群内个体的年龄、大小、数量及其配置情况(Nunes & Petrere Jr, 2012; 张亚芳等, 2015); 后者则是种群数量的时空变化(谭一波等, 2010)。基于静态生命表所构建的存活曲线、数量动态指数、生存函数、谱分析以及时间序列预测模型是研究植物种群结构与动态的重要方法(Harcombe, 1987; 李艳丽等, 2014; 杨立荣等, 2018; 秦爱丽, 2020), 不仅可以用来分析种群当前的生存状态及其与环境间的适合度(Wang et al., 2004; 肖宜安等, 2004), 还可以评估种群的天然更新规律、受干扰状况、预测未来的发展趋势, 对植物资源的保护和利用具有重要意义(陈晓德, 1998; 刘海洋等, 2012)。
红松(Pinus koraiensis)是第四纪冰川孑遗种(傅俊卿, 2009; 董雪等, 2020), 主要分布于中国东北、 俄罗斯远东南部、朝鲜半岛和日本(马建路等, 1992; 杨传平等, 2005)。该种为我国东北地区阔叶红松林的优势树种和主要建群种, 在水源涵养、水土保持及维持生态系统平衡等方面发挥着重要作用(张悦等, 2015)。19世纪末以来, 由于过度采伐和利用, 大范围的原始阔叶红松林遭到破坏, 红松种群数量和质量逐年下降, 蓄积量锐减, 现被列为国家二级重点保护野生植物(于大炮等, 2019)。目前, 得益于“天保工程”的实施以及红松不再作为主要用材树种被采伐, 其种群恢复明显(于大炮等, 2015)。例如, 张悦等(2015)、陈科屹等(2021)报道了皆伐林、天然次生林内红松种群为增长型。然而, 红松种群的天然更新问题依然存在。例如, 张晓鹏等(2021)发现辽东山区3种次生林内的红松种群为衰退型; 在原始阔叶红松林中更是出现“只见幼苗不见幼树”、“红松林下无红松”等现象(李俊清和王业蘧, 1986; 张群等, 2003; 于大炮等, 2015)。因此, 深入了解其天然更新规律, 对该物种的保护和阔叶红松林的恢复至关重要。
红松种群结构的若干文献指出, 该种具有年龄集中分布和几个世代同林的特点(李俊清和王业蘧, 1986; 伍业钢和韩进轩, 1988; 乌弘奇, 1989; 葛剑平等, 1990)。天然更新过程则是年龄集中分布的群体代替另一群体向前发展的过程(伍业钢和韩进轩, 1988)。因此, 通过年龄结构的研究, 进一步探讨种群更新过程, 以及结构与动态的内在联系是可行的。目前, 关于该种天然更新的研究多集中于幼龄期, 认为该阶段决定了种群的演替过程及其在群落中的地位(肖翠等, 2015; 孟令君等, 2018; 张晓鹏等, 2021)。然而, 从物种结构特征来看, 中、成和老龄阶段林木个体也不容忽视, 因为这些年龄的个体一般占据林内上层空间, 这可能会对林下幼苗及幼树产生影响(李俊清和王业蘧, 1986); 另一方面, 种群内不同年龄个体的差异可能影响种内和种间竞争进而决定物种的生命周期(伍业钢和韩进轩, 1988)。因此, 对红松种群完整年龄结构的分析可以清晰地揭示种群的自我维持机制及其动态发展规律。
本研究基于长白山阔叶红松林植被调查数据, 以建群种红松为研究对象, 利用统计学方法, 分析种群年龄结构与数量动态特征, 认识红松种群的生存现状及天然更新规律, 以期为野生红松种群的保护提出适应性对策, 进而为长白山区森林生态系统恢复和生物多样性保护提供科学依据。
1 材料和方法
1.1 研究区概况
研究区位于吉林省东南部长白山国家级自然保护区北坡海拔1 150 m以下的阔叶红松林(127.55°- 128.27° E, 41.07°-42.75° N)。受温带大陆性山地气候影响, 该区夏季短暂多雨, 冬季漫长寒冷; 年降水量630-780 mm, 其中60%集中于6-8月份; 年平均气温3.6 ℃, 最热月(7月)平均气温为19.7 ℃, 最冷月(1月)平均气温为-18.6 ℃; 土壤为发育在火山上的暗棕色森林土, 土层厚度20-100 cm。研究区内群落主林层林龄约300 a、平均高约为26 m, 是典型的复层异龄林。乔木层除红松外, 还有紫椴(Tilia amurensis)、蒙古栎(Quercus mongolica)、五角枫(Acer pictum subsp. mono)、春榆(Ulmus davidiana var. japonica)和水曲柳(Fraxinus mandshurica)等; 灌木主要包括毛榛(Corylus mandshurica)、长白忍冬(Lonicera ruprechtiana)、东北茶藨子(Ribes mandshuricum)、东北溲疏(Deutzia parviflora var. amurensis)、刺五加(Eleutherococcus senticosus)和卫矛(Euonymus alatus)等; 草本层主要有山茄子(Brachybotrys paridiformis)、蚊子草(Filipendula palmata)、荨麻叶龙头草(Meehania urticifolia)、山尖子(Parasenecio hastatus)和中华蹄盖蕨(Athyrium sinense)等。
1.2 研究方法
1.2.1 样地设置
2020年6-8月, 以长白山北坡的阔叶红松林为研究对象, 采用样地调查法, 沿海拔高度每间隔50 m设置5个样地作为5个重复, 样地大小为30 m × 30 m, 共40个样地(图1), 总样地面积 36 000 m2。采用每木检尺法, 对样地内胸径(DBH) ≥1 cm的乔木进行调查, 记录每个乔木的胸径、高度和冠幅等。
图1
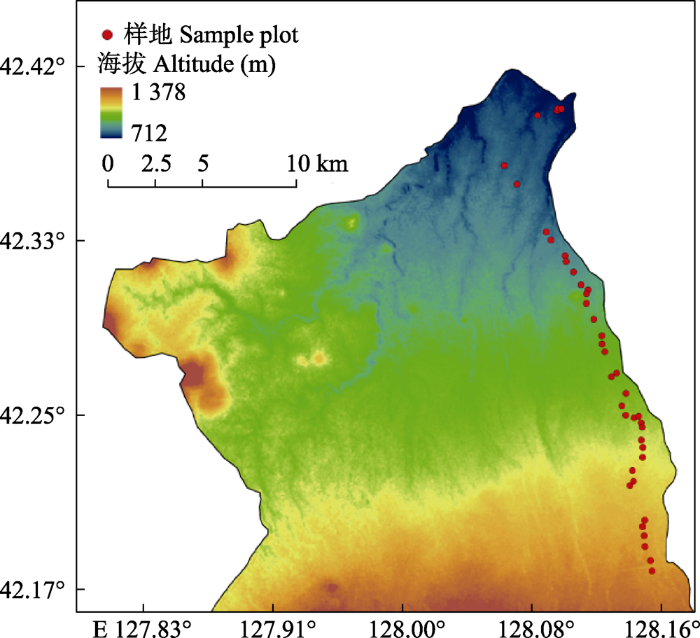
图1
长白山阔叶红松林红松种群样地分布图。
Fig. 1
Map of sampling locations for Pinus koraiensis population in a broadleaved Korean pine forest in Changbai Mountain.
1.2.2 红松种群龄级结构划分
红松为稀有物种, 其种群数量有限, 不宜利用解析木或生长锥进行年龄测定。因此, 本研究依据同一生境内, 同种植物的龄级和径级对环境具有相同的反应规律这一特点, 采用径级代替龄级的方法来分析种群动态变化(李艳丽等, 2014; 秦爱丽等, 2020)。根据样地内调查情况和红松的生长规律, 并参考董雪等(2020)对红松种群龄级结构的划分标准, 将其划分为14个龄级, 具体如下: I, 0-5 cm; II, 5-10 cm; III, 10-15 cm; IV, 15-20 cm; V, 20-25 cm; VI, 25-30 cm; VII, 30-35 cm; VIII, 35-40 cm; IX, 40-45 cm; X, 45-50 cm; XI, 50-55 cm; XII, 55-60 cm; XIII, 60-65 cm; XIV, >60 cm; 同时, 将14个龄级分为4个阶段, 即幼龄阶段(I-III龄级)、中龄阶段(IV-VI龄级)、成龄阶段(VII-X龄级)和老龄阶段(XI-XIV龄级)。
1.2.3 红松种群静态生命表
以红松种群各龄级的株数为依据, 编制其静态生命表。表中参数如下: 1) x, 龄级; 2) Ax, x龄级内存活个体数。由于静态生命表反映的是多个世代重叠的植物种群在某一特定时刻的年龄分布, 而不是对同龄出生群的全部生活史的追踪, 因此, 编制生命表过程中会出现死亡率为负值的情况, 故采用匀滑技术(刘海洋等, 2012; 姜在民等, 2018)对x龄级内的存活个体数Ax进行处理, 得到修正后的数据ax, a0为ax的初始值; 3) lx, x龄级的标准化存活个体数, lx = ax/a0 × 1000; 4) lnlx, lx的自然对数; 5) dx, 从x龄级到x + 1龄级的死亡个体数, dx = lx - lx+1; 6) qx, 从x龄级到 x + 1龄级的种群个体死亡率, qx = dx/lx × 100%; 7) Lx, 从x到x + 1龄级的平均存活个体数, Lx = (lx + lx+1)/2; 8) Tx, 大于等于x龄级的存活个体总数, Tx = ∑Lx; 9) ex, 进入x龄级个体的生命期望或平均期望寿命, ex = Tx/Lx; 10) Kx, 各龄级内的消失率, Kx = lnlx - lnlx+1; 11) Sx, 种群存活率, Sx = lx+1/lx。
1.2.4 种群动态量化分析
本研究采用陈晓德(1998)的动态量化分析法定量描述红松种群结构:
式中, Vx为种群内相邻两龄级间个体数量变化动态指数, x为龄级; Vpi、V′pi分别为忽略外部干扰和考虑到存在未来外部干扰时整个种群的数量动态变化指数; Ax、Ax+1分别为第x、x + 1龄级种群个体数; K为年龄级数量。
Vx、Vpi、V′pi取正、0、负值时分别反映种群内两相邻级(或整个种群)个体数量的增长、稳定和衰退。
1.2.5 存活曲线、死亡率曲线和消失率曲线拟合
本研究以龄级x为横坐标, 标准化存活数lx为纵坐标, 绘制种群存活曲线。根据存活曲线的变化特征, Deevey Jr (1947)将其分为3种类型: Deevey-I型(凸型), 该类型种群幼龄个体死亡率较低且大多数都能活到其平均生理寿命, 但达到平均生理寿命时, 短期内几乎全部死亡; Deevey-II型(直线型), 该类型种群各龄级死亡率大致相同; Deevey-III型(凹型), 该类型种群中幼龄个体死亡率较高。本研究采用指数方程ax = a0e-bx和幂函数方程ax = a0e-b (ax, 匀滑后x龄级内存活数; a0, 种群初始个体数; b, 死亡率)来验证种群符合上述哪种类型。
以龄级x为横坐标, 死亡率qx、消失率Kx为纵坐标, 分别绘制种群死亡率曲线和消失率曲线。
1.2.6 生存分析
应用生存分析理论中的生存函数Si、累计死亡率函数Fi、死亡密度函数fti和危险率函数λti对红松种群进行定量分析。计算公式见杨凤翔等(1991)的研究。
1.2.7 谱分析
谱分析被广泛应用于种群天然更新过程的研究, 常用来揭示种群内树木年龄更替过程的周期性以及结构的波动性(伍业钢和韩进轩, 1988)。谱分析是Fouirier级数的展开, 其周期现象由不同振幅和相对应的谐波构成, 可写成正弦波形式:
式中, A0为周期变化的平均值; t为时间系列, 即种群的各年龄分布; n为时间系列总长度, 即种群的龄级数(本研究中n = 14); Ak (k = 1, 2, 3, …, p)为各谐波的振幅, 其值反映了各周期所起作用的大小, 且p = n/2为谐波个数; ωk和θk分别为谐波频率与相角; Nt为t时刻种群大小。A0、Ak、ωk、θk等参数由下式估算, 即:
式中, ak、bk为参数估计值; Xt为t年龄序列的个体数, 该值与生命表中ax栏内数值相对应。考虑到本研究中红松种群ax相差较大, 因此在计算前首先对Xt进行对数处理, 即用ln(Xt + 1)代替Xt。利用谱分析中的公式计算各种情况下各个波形的振幅(Ak), A1为基波, A1-Ak为谐波。每个谐波的周期分别是基本周期的1/2, 1/3, …, 1/p。
1.2.8 时间序列模型预测
采用时间序列分析中的一次平均推移法对红松种群未来2、3、4、5、6、7、8、9、10个龄级的年龄结构动态进行模拟和预测。种群存活数计算参照肖宜安等(2004)的方法。
1.3 数据处理
本研究所有统计分析在Excel 2007和SPSS 21中进行, 用Origin 2019软件绘图。
2 结果和分析
2.1 红松种群年龄结构
本次共调查到红松个体280株, 最大胸径 135.52 cm。静态生命表(表1)显示, 红松种群不同龄级个体存活数差别大, II龄级个体数最多(52株); 前3个龄级个体数为123株, 占个体总数的43.9%, 表明幼苗数量充足, 种群具有一定的增长潜力。IV龄级后种群个体数量降低趋势明显, 说明幼龄个体很难向中龄阶段过渡。随龄级增加, 成龄(VII-X龄级)阶段种群个体数呈增加趋势(分别为15、19、19、23株), 反映出林木个体度过幼、中龄级后生存能力显著增强。老龄(XI-XIV龄级)阶段种群个体数为33株, 占个体总数的11.8%。随龄级增加, 红松种群标准化存活数逐渐减小; 生命期望值随龄级增加呈先增后降的趋势, IV龄级种群具最大生命期望, 说明在该龄级种群生长最旺盛, 生存质量最高。
表1 长白山阔叶红松林红松种群静态生命表
Table 1
| 龄级 Age class (x) | 径级 DBH class (cm) | Ax | ax | lx | dx | qx | Lx | Tx | ex | lnlx | Kx | Sx |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| I | 0-5 | 30 | 51 | 1 000 | 196 | 0.20 | 902 | 4 971 | 4.97 | 6.91 | 0.22 | 0.78 |
| II | 5-10 | 52 | 41 | 804 | 196 | 0.24 | 706 | 4 069 | 5.06 | 6.69 | 0.28 | 0.76 |
| III | 10-15 | 41 | 31 | 608 | 137 | 0.23 | 539 | 3 422 | 5.63 | 6.41 | 0.26 | 0.84 |
| IV | 15-20 | 23 | 24 | 471 | 39 | 0.08 | 451 | 2 882 | 6.13 | 6.15 | 0.09 | 0.91 |
| V | 20-25 | 14 | 22 | 431 | 39 | 0.09 | 412 | 2 431 | 5.64 | 6.07 | 0.10 | 0.90 |
| VI | 25-30 | 11 | 20 | 392 | 39 | 0.10 | 373 | 2 020 | 5.15 | 5.97 | 0.11 | 0.89 |
| VII | 30-35 | 15 | 18 | 353 | 39 | 0.11 | 333 | 1 647 | 4.67 | 5.87 | 0.12 | 0.88 |
| VIII | 35-40 | 19 | 16 | 314 | 39 | 0.13 | 294 | 1 313 | 4.19 | 5.75 | 0.13 | 0.87 |
| IX | 40-45 | 19 | 14 | 275 | 39 | 0.14 | 255 | 1 020 | 3.71 | 5.61 | 0.15 | 0.85 |
| X | 45-50 | 23 | 12 | 235 | 39 | 0.17 | 216 | 765 | 3.25 | 5.46 | 0.18 | 0.86 |
| XI | 50-55 | 10 | 10 | 196 | 19 | 0.10 | 186 | 549 | 2.80 | 5.28 | 0.11 | 0.89 |
| XII | 55-60 | 9 | 9 | 176 | 19 | 0.11 | 167 | 363 | 2.06 | 5.17 | 0.12 | 0.82 |
| XIII | 60-65 | 8 | 8 | 157 | 39 | 0.25 | 137 | 196 | 1.25 | 5.06 | 0.29 | 0.43 |
| XIV | >65 | 6 | 6 | 118 | - | - | 59 | - | - | 4.77 | - | - |
Ax, x龄级内存活个体数; ax, 匀滑后x龄级内的存活个体数; lx, x龄级的标准化存活个体数; dx, 从x龄级到x + 1龄级的死亡个体数; qx, 从x龄级到x + 1龄级的种群个体死亡率; Lx, 从x到x + 1龄级的平均存活个体数; Tx, 大于等于x龄级的存活个体总数; ex, 平均期望寿命; Kx, 种群消失率; Sx, 种群存活率。
Ax, actual survival number; ax, the correction value of Ax; lx, logarithmic standard surviving number in x age class; dx, standardized deaths from x to x + 1 age class; qx, mortality rate from x to x + 1 age class; Lx, survived individuals from x to x + 1 age class; Tx, total survival number of individuals over x age class; ex, average life expectancy; Kx, vanish rate; Sx, survival rate; DBH, diameter at breast height.
2.2 红松种群动态分析
2.2.1 红松种群数量动态
种群数量动态量化分析结果见表2。红松种群VI、VVI-VVII、VIX小于0, VVIII等于0, 而VII-VV、VX-VXIII大于0, 表明在I-XIII龄级范围内红松种群数量呈现出“衰退-增长-衰退-稳定-衰退-增长”的变化趋势。忽略外部干扰时, 整个种群的数量变化动态指数Vpi > 0, 可知红松种群为增长型。然而, V′pi趋近于0, 说明该种群受外界随机干扰时增长趋势不明显。
表2 长白山阔叶红松林红松种群动态变化指数
Table 2
| VI | VII | VIII | VIV | VV | VVI | VVII | VVIII | VIX | VX | VXI | VXII | VXIII | Vpi | V′pi |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| -42.31 | 21.15 | 43.90 | 39.13 | 21.43 | -26.67 | -21.05 | 0 | -17.39 | 56.52 | 10.00 | 11.11 | 25.00 | 13.11 | 0.12 |
VI-VXIII, 种群内相邻两龄级间个体数量变化动态指数, 龄级同
VI-VXIII, dynamic index of of quantitative change between adjacent age classes in population, age class see
2.2.2 死亡率和消失率曲线
图2
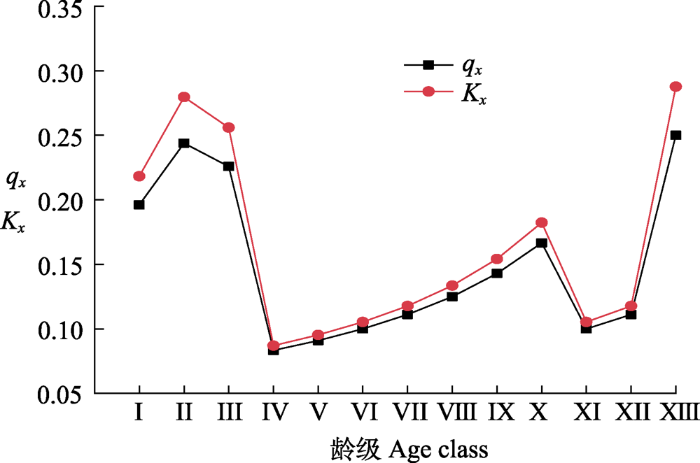
图2
长白山阔叶红松林红松种群死亡率(qx)和消失率(Kx)曲线。龄级同
Fig. 2
Curves of mortality rate (qx) and vanish rate (Kx) for Pinus koraiensis population in broadleaved Korean pine forest in Changbai Mountain. Age class see
2.2.3 存活曲线
图3
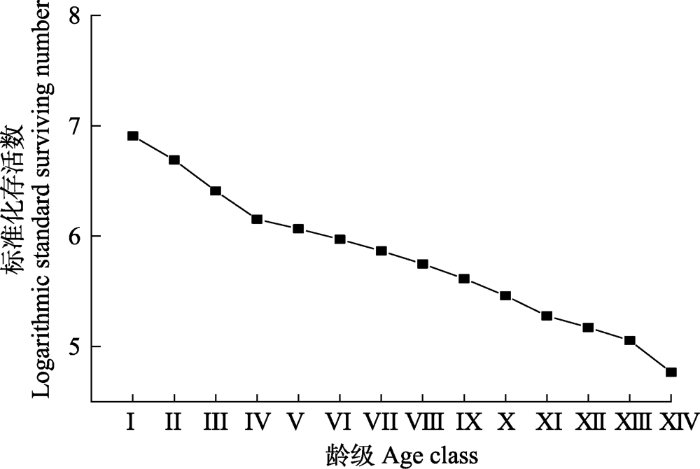
图3
长白山阔叶红松林红松种群存活曲线。龄级同
Fig. 3
Survival curves of Pinus koraiensis population in a broadleaved Korean pine forest in Changbai Mountain. Age class see
表3 长白山阔叶红松林红松种群存活曲线的检验模型
Table 3
| 存活曲线类型 Type | 拟合模型 Test model | 方程 Equation | R2 | F | p |
|---|---|---|---|---|---|
| Deevey-II | ax = a0e-bx | ax = 7.742e-0.038x | 0.623 | 18.918 | 0.001 |
| Deevey-III | ax = a0x-b | ax = 1.496x-0.853 | 0.718 | 30.505 | 0.000 |
ax, 匀滑后x龄级内存活数; N0, 种群初始个体数; b, 死亡率。
ax, the correction value of actual survival number of x age class; a0, initial number of individuals in population; b, mortality rate.
2.3 生存分析
图4显示, 随龄级增加, 红松种群生存率单调递减, 累积死亡率单调递增, 二者互补。I-III龄级间二者的变化趋势较IV-XII龄级更明显, 这可能跟水热条件和生长空间有限而导致的强竞争作用有关。在第III龄级, 生存率和累计死亡率相等, 表明种群达到平衡状态, 尔后累计死亡率大于生存率, 种群开始衰退。XII龄级后, 生存率骤降而累计死亡率骤升, 这可能跟老龄个体的生理衰老有关。
图4
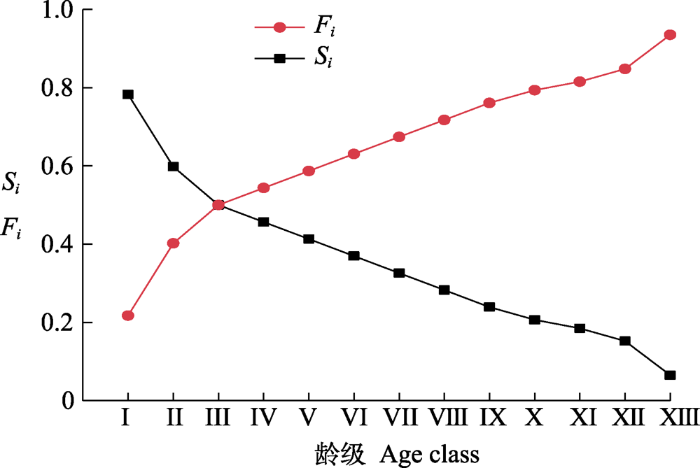
图4
长白山阔叶红松林红松种群生存率(Si)和累计死亡率(Fi)曲线。龄级同
Fig. 4
Curves of survival rate (Si) and cumulative mortality rate (Fi) for Pinus koraiensis population in a broadleaved Korean pine forest in Changbai Mountain. Age class see
图5显示, 红松种群危险率曲线整体呈单调递减。其中, 斜率随龄级增加逐渐减小, 表明危险率前期大后期小。死亡密度曲线在第I-II径级间急剧下降, 在第V龄级出现最小值(0)。4个生存函数曲线共同表明, 长白山阔叶红松林内红松种群具有前半期锐减、中期稳定、后期衰退的特征。
图5
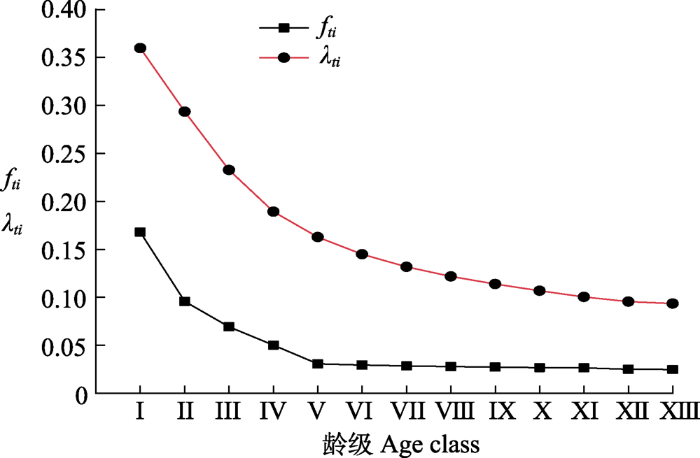
图5
长白山阔叶红松林红松种群死亡密度(fti)与危险率(λti)曲线。龄级同
Fig. 5
Mortality density (fti) and hazard rate curves (λti) of Pinus koraiensis population in a broadleaved Korean pine forest in Changbai Mountain. Age class see
2.4 谱分析
由表4可以看出, 整个种群的基波最大(A1 = 1.73), 说明其在红松种群天然更新过程中所起的作用最大, 表现为红松种群波动具有大周期。从谐波A2-A6振幅值的变化来看, 红松种群生长过程也容易受到小周期波动的影响。其中, 比较明显的小周期主要分布在基本周期的1/4和1/10处, 二者分别对应空间序列径级IV和X, 即中龄阶段早期和成龄阶段末期, 这种小周期的波动可能与红松种群生物学特性以及种内、种间竞争有关, 反映出种群天然更新过程的数量动态特征。
表4 长白山阔叶红松林红松种群的周期性波动
Table 4
| 谐波 Resonance spectra | A0 | A1 | A2 | A3 | A4 | A5 | A6 | A7 |
|---|---|---|---|---|---|---|---|---|
| 振幅值 Amplitude value | 3.18 | 1.73 | 0.41 | 0.15 | 0.23 | 0.36 | 0.13 | 0.02 |
2.5 时间序列模拟和预测
本研究按照一次平均推移法, 预测种群在未来2、3、4、5、6、7、8、9、10个龄级时间后的数量动态变化趋势(表5)。结果显示, II龄级个体数在未来2个龄级时间后减少; III、IV、V龄级个体数分别经过2、3、4个龄级时间后达到峰值, 随后开始下降, 上述结果共同说明幼、中龄个体数在未来呈减少趋势。VII、VIII、IX、X、XI、XII、XIII和XIV龄级个体数在未来总体呈现增加的趋势, 推测未来种群以大树、老树占主导地位, 整个红松种群向高龄级个体集中。然而, 老龄阶段(XI-XIV龄级)个体的平均增长速率低于成龄阶段(VII-X龄级)(分别为10.86%和19.26%), 这可能跟生理衰老有关。
表5 长白山阔叶红松林红松种群数量动态时间序列预测
Table 5
| 龄级 Age class | >初始个体数 Initial number of individuals | Mt | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| M2 | M3 | M4 | M5 | M6 | M7 | M8 | M9 | M10 | ||||||||||
| I | 30 | |||||||||||||||||
| II | 52 | 41 | ||||||||||||||||
| III | 41 | 47 | 41 | |||||||||||||||
| IV | 23 | 32 | 39 | 37 | ||||||||||||||
| V | 14 | 19 | 26 | 33 | 32 | |||||||||||||
| VI | 11 | 13 | 18 | 22 | 28 | 29 | ||||||||||||
| VII | 15 | 13 | 14 | 16 | 21 | 26 | 27 | |||||||||||
| VIII | 19 | 17 | 14 | 15 | 16 | 21 | 25 | 26 | ||||||||||
| IX | 19 | 19 | 17 | 16 | 16 | 17 | 20 | 24 | 25 | |||||||||
| X | 23 | 21 | 16 | 19 | 17 | 17 | 18 | 21 | 24 | 25 | ||||||||
| XI | 10 | 17 | 11 | 18 | 17 | 16 | 16 | 17 | 19 | 23 | ||||||||
| XII | 9 | 10 | 9 | 15 | 16 | 16 | 15 | 15 | 16 | 18 | ||||||||
| XIII | 8 | 9 | 9 | 13 | 14 | 15 | 15 | 14 | 14 | 15 | ||||||||
| XIV | 8 | 8 | 8 | 9 | 12 | 13 | 14 | 14 | 14 | 14 | ||||||||
Mt, 经过2、3、4、5、6、7、8、9、10个龄级时间后种群的大小。龄级同
Mt, the number of individuals in each age class after the population has developed from 2 to 10 age class in this study. Age class see
3 讨论和结论
3.1 红松种群年龄结构特征
长白山阔叶红松林红松种群数量变化具有阶段性, 幼龄(I-III龄级)个体较多, 占总数的43.9%, 表明植株具有一定的自然更新能力, 结合总体数量动态变化指数Vpi > 0, 可知种群为增长型。然而, 在IV-VI龄级间, 种群个体数量锐减, 说明幼龄个体难以向中龄阶段过度, 种群缺乏幼树。与之类似, 葛剑平等(1990)发现小兴安岭天然红松种群幼龄个体具有明显的数量优势, 而小树分布极少。这可能是其生物学特性和环境因子共同作用的结果。红松果实体积和质量较大, 其成熟后多散落于母树周围, 这种生物学特性使幼苗在空间上易形成聚集分布的特征(李俊清和祝宁, 1990)。然而受密度制约影响, 聚集分布的幼龄个体之间势必存在激烈的种内竞争, 相互之间争夺光照、水分和养分等资源以满足其生存需要, 导致了大量个体死亡(邱华等, 2020); 除种内竞争外, 幼龄个体还受到邻近其他植物以及主林层大树对养分和空间资源的不对称竞争, 增加了死亡率(肖翠等, 2015; 刘帅等, 2016)。此外, 尽管有研究指出红松幼龄个体具有较强的耐阴性, 但其向大径级林木转化仍需充足光照(李俊清和祝宁, 1990)。实地调查发现, 阔叶红松林郁闭度大, 光照弱, 凋落层较厚, 加之该区冬季寒冷漫长, 强烈的环境筛也会导致幼龄个体生长不良甚至死亡(邱华等, 2020; 哈努拉·塔斯肯等, 2021)。由此可见, 红松幼龄个体要进入中龄阶段, 必须经过竞争以及环境因子的多重筛选, 致使幼龄个体不能向下一级林木大量转化, 说明幼龄阶段是红松种群更新和发展的“瓶颈”。
3.2 红松种群的动态特征
植物种群的静态生命表、存活曲线、死亡率曲线和消失率曲线能够揭示种群的生存现状和种群对环境的适应机制(Wu et al., 2010; Kang et al., 2014; 秦爱丽等, 2020)。研究发现, 随龄级的增加, 红松种群的生命期望值呈先增后降的趋势, IV龄级种群具最大生命期望值, 说明该龄级种群对环境的适应能力最强, 生存质量最高; V龄级后, 随着植株个体的发育, 对空间和资源的需求逐渐增大, 种内和种间竞争作用增强, 期望寿命开始下降, 这与周赛霞等(2020)对庐山黄山松(P. taiwanensis)种群以及张亚芳等(2015)对黄土高原不同地区杜松(Juniperus rigida)种群的研究结果类似。从存活曲线可以看出, 红松种群属于Deevey-III型, 即种群内幼龄个体死亡率高, 这与上述种群年龄结构特征分析的结果相对应, 然而该研究结果与张悦等(2015)对长白山西北麓东升林场红松种群存活曲线趋近于Deevey-II型有所不同, 这可能与不同研究区域的生境差异、外界干扰以及调查范围有关。本研究中, 种群死亡率和消失率波动较为频繁, 呈现出双峰型, 即死亡率和消失率分别在II和XIII龄级达到高峰, 前者主要受生存空间和资源条件的限制, 而后者可能是种群生理衰老所致。
生存函数能够更直观、具体地反映种群增长与衰亡的动态过程(谭一波等, 2010)。研究发现, I-III龄级间种群生存率大于累计死亡率, 这是因为幼龄个体数量充足, 基本上能及时补充由竞争作用而导致的该龄级个体迁出或者死亡所造成的数量缺失; 在III龄级, 生存率等于累计死亡率, 种群达到平衡状态, 此后种群逐渐向衰退方向发展, 生存空间和资源条件有限以及个体自然衰老是红松种群进入衰退期的主要因素(张悦等, 2015; 董雪等, 2020)。此外, 本研究中, I-III龄级间种群生存率和累计死亡率的变化趋势较IV-XII龄级更明显, 死亡密度和危险率也具有前期大后期小的特点。4个生存函数曲线共同表明, 红松种群的更新过程具有阶段性, 即前期锐减、中期稳定、后期衰退。这与种群静态生命表及与其相关的存活曲线、死亡率曲线和消失率曲线的分析结果相同。
3.3 红松种群的谱分析与时间序列预测
种群更新的动态是通过该种群不同龄级个体数量的分布波动来表现(吴其超等, 2021)。伍业钢和韩进轩(1998)对小兴安岭、张广才岭和长白山次生林红松种群的研究显示, 该物种的天然更新具有320 a左右的大周期, 但种群个体数量存在小周期(70-107 a)和中等周期(140-160 a)的波动, 并认为这种周期性波动可能是维持红松种群稳定的一个机制。本研究结果与上述研究者的结论基本一致, 即红松种群天然更新过程呈现出基本周期内的小周期波动。从本研究结果来看, 红松种群的更新受基波A1影响较大, 基波的周期长度为种群本身所固有, 由种群波动的特性所决定, 表现了种群基本周期的波动(吴其超等, 2021)。因此, 红松种群的更新过程主要受物种生物学特性的控制, 具有明显的大周期。除受到基波的影响外, 红松种群还具有小周期波动现象。其中, 发生在IV和X龄级的小周期波动较为明显, 大量幼龄个体死亡和生理衰老可能是造成小周期波动明显的主要原因。在红松种群的生长过程中, 这种小周期的波动能够促进种群的自然稀疏及其数量调节, 从而维持其自身的稳定。
时间序列预测分析表明, 经历2个龄级时间后, 第II龄级种群个体数量减少, III-V龄级个体数在经过2、3、4个龄级时间后达到峰值, 随后开始下降, 表明红松幼龄个体难以向中龄阶段发展, 这在一定程度上限制了红松种群的自然更新。徐玮泽等(2018)报道的“长白山阔叶红松林红松种群在过去35年内几乎无进阶幼树产生”的结论可从另一侧面为上述结果提供佐证。VI-XIV龄级个体数在未来总体呈现增加的趋势, 整个红松种群向高龄级个体集中, 在外界环境保持良好的前提下, 种群会呈现增长趋势。然而, 一旦植株达到生理年龄后, 种群开始走向衰退。另外, 本研究中, 红松老龄个体数量虽然少但占据较大的上层空间, 因此, 从种群整体发展角度来看, 少数老龄个体的消亡可促使幼龄个体进入上层空间, 有利于种群更新和繁衍(李俊清和祝宁, 1990)。
3.4 红松种群的保护对策
红松是长白山阔叶红松林的主要建群种, 对维系该地区生物多样性、调节气候和维持生态平衡等具有至关重要的作用。过去一个多世纪, 受采伐的影响, 长白山红松分布区呈锐减趋势(于大炮等, 2019), 因此, 亟待对红松进行有效的保护、恢复和重建。根据本研究结果, 长白山阔叶红松林红松种群更新与恢复存在以下问题: 一是种群幼龄个体死亡率高且难以成林; 二是在干扰情况下种群增长缓慢; 三是种群老龄个体衰亡明显。因此, 建议采取以下保护措施: 1)开展实生苗繁育和回归工作, 提高幼苗质量和存活率; 同时, 为防止远交衰退所带来的遗传没化, 实生苗的种子须从该原生种群采集(丁剑敏等, 2018)。2)采取封育保护措施, 避免对红松种群栖息地造成破坏。在封育保护的同时进行科学的经营和管理, 从而促进林下长期“蹲守”的小径级植株进入林冠层。3)实行近地保护, 选择与红松原生境具有相同气候条件的近地点进行人工育苗, 从而促进人工种群的恢复。4)目前已建立的长白山国家级自然保护区对该区植物资源保护起到了一定作用, 但诸如采集食用菌、药用植物等人为干扰因素仍较多, 加之近年来长白山地区旅游业逐渐兴起(马琳和李俊清, 2019), 建议加强对保护区的管理力度, 杜绝人为破坏。
参考文献
Population structure and dynamic characteristics of typical constructive species in natural secondary forest on the northern slope of Changbai Mountain
长白山北坡天然次生林典型建群种的种群结构及动态特征
A study on the method of quantitative analysis for plant population and community structural dynamics
植物种群与群落结构动态量化分析方法研究
Population structure and dynamics of Abies spectabilis at treeline ecotone of Barun Valley, Makalu Barun National Park, Nepal
DOI:10.1016/j.chnaes.2016.05.003 URL [本文引用: 1]
Life tables for natural populations of animals
DOI:10.1086/395888 URL [本文引用: 1]
Genetic considerations in recovery of endangered plants
濒危植物居群恢复的遗传学考量
Spatial pattern and intraspecific association of natural Korean pine population under the influence of habitat gradient
生境梯度影响下的天然红松种群空间格局与种内关联
Current situation of natural Korean pine forests in northeast China and their management countermeasures
东北天然红松林资源现状与保护经营对策
Study on age structure and spatial pattern of old-growth Korean pine forest in Lesser Xingan Mountain
小兴安岭天然红松林种群结构的研究
Age structure models of balsam fir and eastern hemlock
DOI:10.2307/2258822 URL [本文引用: 1]
Population structure and dynamic characteristics of endangered Syringa pinnatifolia Hemsl
濒危植物羽叶丁香种群结构与动态特征
Population structure and spatial pattern of main tree species in secondary Betula platyphylla forest in Ziwuling Mountains, China
DOI:10.1038/srep06873
PMID:25362993
[本文引用: 1]

This study investigated a typical secondary Betula platyphylla forest in the Ziwuling Mountains, Loess Plateau, China. In the sample plot, the DBH (diameter at breast height) class structure of B. platyphylla was bimodal. Individuals with small and large DBH values were abundant. The DBH structures of Quercus wutaishanica and Pinus tabulaeformis were close to that of the logistic model, thus suggesting the increasing population of these species. B. platyphylla and Populus davidiana showed random spatial distributions at almost all scales. However, Q. wutaishanica and P. tabulaeformis were significantly clumped at small scales. B. platyphylla had a negative spatial relation with Q. wutaishanica at small spatial scales. P. tabulaeformis and Q. wutaishanica showed negative spatial correlations at small scales, but they had positive correlations at large scales. These results suggest that P. tabulaeformis and Q. wutaishanica shared habitat preferences at these scales. In the future, the secondary B. platyphylla forest in the Ziwuling Mountains in the Loess Plateau will probably change into a multi-species mixed forest (Quercus-Pinus mixed forest). Assisted restoration strategies must be employed to improve the regeneration dynamics of the forest in the long term.
Wave features of population changes of Pinus koraiensis in natural forest
天然林内红松种群数量变化的波动性
Structure of Korean pine and its dynamics
红松的种群结构与动态过程
Population structures and dynamics of Abies nephrolepis and Picea koraiensis in the Changbai Mountains of northeastern China
长白山云冷杉种群结构和动态分析
Population quantitative characteristics and dynamics of rare and endangered plant Davidia involucrata in Hunan Province
DOI:10.5846/stxb201203270425 URL [本文引用: 2]
湖南珍稀濒危植物--珙桐种群数量动态
Local biotic neighborhood and habitat heterogeneity drive tree seedling survival dynamics in a temperate forest
生物邻体和生境异质性共同驱动乔木幼苗存活动态
Geographic distribution of Pinus koraiensis in the world
红松的地理分布
Systematic conservation planning optimization for the broad-leaved Korean pine mixed forest conservation network system in the Changbai Mountain Region, China
基于系统保护规划的长白山阔叶红松林保护网络优化研究
Drivers of composition and density pattern of tree seedlings in a secondary mixed conifer and broad-leaved forest, Jiaohe, Jilin, China
DOI:10.17521/cjpe.2018.0025 URL [本文引用: 1]
吉林蛟河针阔混交林乔木幼苗组成及其密度格局影响因素
DOI:10.17521/cjpe.2018.0025
[本文引用: 1]
以吉林蛟河次生针阔混交林42 hm <sup>2</sup>固定监测样地中209个幼苗监测站内的乔木幼苗为研究对象, 基于2016和2017年幼苗调查数据, 探究幼苗物种组成、数量动态等特征, 并运用广义线性混合效应模型分析了幼苗密度与生物邻体及其生境因素的相关性。结果表明: (1)该样地内所有幼苗监测站共调查到幼苗4 245株, 分属10科12属18种, 新生幼苗的数量在物种和调查年份间均有明显差异, 其中水曲柳(Fraxinus mandschurica)和紫椴(Tilia amurensis)幼苗表现出大量出生和死亡的动态特征。(2)群落水平上, 幼苗密度与局域同种成体胸高断面积之和、土壤全磷和有效钾等养分含量显著正相关, 适宜大树生长的生境同样适宜幼苗的生长。(3)水曲柳幼苗密度的影响因素与群落水平一致, 红松(Pinus koraiensis)存在明显的生境偏好, 在湿度较小、土壤全磷、有效氮和有效磷含量较低的生境中密度更高。相对于多年生幼苗, 同种成年邻体对当年生幼苗密度影响更加显著。该研究证实了扩散限制和生境过滤共同影响幼苗密度格局, 生物邻体和生境异质性的相对重要性随幼苗物种种类和年龄级变化。
Structure and dynamics of a Cariniana estrellensis (Lecythidaceae) population in a fragment of Atlantic Forest in Minas Gerais, Brazil
DOI:10.1590/S2175-78602012000200002 URL [本文引用: 1]
Population structure and dynamic characteristics of a rare and endangered tree species Michelia wilsonii Finet et Gagn
珍稀濒危树种峨眉含笑种群结构与动态特征
Influencing factors of composition and abundance pattern of tree seedlings in broad-leaved Korean pine (Pinus koraiensis) mixed forest, Changbai Mountain, China
长白山阔叶红松林乔木幼苗组成及多度格局的影响因素
Population structure and dynamic characteristics of Machilus chinensis in Nan'ao Island, Guangdong Province
广东南澳岛中华楠种群结构及动态特征
Effects of canopy structure on productivity in a typical mixed broadleaved Korean pine forest
DOI:10.17521/cjpe.2020.0176 URL [本文引用: 1]
树冠结构对典型阔叶红松林生产力的影响
Age structure of Picea schrenkiana forest along an altitudinal gradient in the central Tianshan Mountains, northwestern China
DOI:10.1016/j.foreco.2004.02.063 URL [本文引用: 1]
Dynamics of Korean pine population in birch forests of the Changbai Mountain
长白山白桦林中红松种群动态的研究
Characteristics and dynamics of Populus euphratica population at lower reaches of Tarim River
塔里木河下游胡杨种群特征与动态分析
Population structure and dynamics of endangered Populus wulianensis
濒危树种五莲杨种群结构与动态特征
A spectral analysis of the population dynamics of Korean pine in the mixed broad-leaved Pinus koraiensis forest
阔叶红松林红松种群动态的谱分析
Effects of biotic and abiotic factors on tree seedling survival in a broad leaved Korean pine (Pinus koraiensis) mixed forest on Changbai Mountain
长白山阔叶红松林中影响乔木幼苗存活的关键因素
Study on numeric dynamics of natural populations of the endangered species Disanthus cercidifolius var.longipes
濒危植物长柄双花木自然种群数量动态
DOI:10.17521/cjpe.2004.0037
[本文引用: 2]
运用种群生命表、生殖力表、Leslie矩阵模型和时间序列预测分析方法,研究了濒危植物长柄双花木(Disanthus cercidifolius var. longipes)种群的动态变化过程,揭示了长柄双花木各龄级植株的数量动态规律。结果表明:长柄双花木为缓慢负增长型种群;种群的净增殖率、内禀增长率以及周限增长率都较低,世代平均周期较长;Leslie矩阵模型和时间序列预测分析均表明在未来50年内长柄双花木各年龄级种群数量会出现波动性的消长,但其种群总数将逐步下降。导致种群下降的主要原因可能是人为砍伐及由此造成的生境破碎化等。
Long-term monitoring of tree population dynamics of broad-leaved Korean pine forest in Changbai Mountains, China
长白山阔叶红松林树木种群动态的长期监测
DOI:10.13287/j.1001-9332.201810.004
[本文引用: 1]
基于1981—2016年对长白山阔叶红松林固定样地(面积 1 hm<sup>2</sup>)4次调查的监测数据,研究了阔叶红松林树种组成与结构的变化特征,分析了各树种的径阶分布、空间分布、数量变化等.结果表明: 35年间,林分的树种组成变化不大,林分总密度呈下降趋势.总胸高断面积和蓄积量分别为 43~45 m<sup>2</sup>·hm<sup>-2</sup>和474~496 m<sup>3</sup>·hm<sup>-2</sup>,其中,红松的优势度最大,胸高断面积和蓄积量分别占57.8%~59.7%和62.5%~65.4%.所有优势树种的空间分布无明显变化,均呈随机分布.其中,红松的径级结构呈近似正态分布,乔木层全部树种的径级结构呈倒 “J” 型分布.小径级(10~26 cm)死亡个体的绝对数量最多,死亡比例为30%,而中径级(30~50 cm)死亡比例最高,为30%~50%.乔木层中除色木槭外的主要树种均没有明显进阶,死亡率高于更新率,特别是红松,完全没有更新或进阶,种群处于衰退状态.原始阔叶红松林中红松、紫椴等优势树种的更新受到阻碍,这些树种的种群维持可能需要自然干扰.
Analysis of genetic diversity for nineteen populations of Pinus sibirica Du Tour with technique of ISSR
应用ISSR PCR对西伯利亚红松19个种源的遗传多样性分析
The theory of survival analysis and its application to life table
生存分析理论及其在研究生命表中的应用
The population structure and dynamics of Dracaena cambodiana, an endangered tree on Hainan Island
濒危植物海南龙血树的种群结构与动态
Forest management of Korean pine and broadleaf mixed forest in Northeast China since the implementation of Natural Forest Protection Project
DOI:10.1016/j.chnaes.2014.12.004 URL [本文引用: 2]
天保工程实施以来东北阔叶红松林的可持续经营
Exploring the history of the management theory and technology of broad leaved Korean pine (Pinus koraiensis Sieb. et Zucc.) forest in Changbai Mountain Region, Northeast China
长白山区阔叶红松林经营历史与研究历程
DOI:10.13287/j.1001-9332.201905.004
[本文引用: 3]
长白山区位于中国东北东部的山地森林区,地带性植被是阔叶红松林,是我国珍贵木材生产和东北森林带的重要载体,孕育着丰富的物种资源.短短120年来,长白山区广袤的原始森林几乎消失殆尽,被次生林、过伐林和人工林等替代.建国以来,东北的阔叶红松林经营理论与技术在红松更新规律、皆伐与择伐的争论中,在人们对阔叶红松林结构和功能认识不断深入的过程中逐渐完善.但目前红松更新问题依然存在,森林经营依然十分粗放.本文梳理了过去70年来阔叶红松林的经营历史及研究历程,分析当前森林资源类型和经营制度的形成过程,以期为该区天然林可持续经营和质量的精准提升等提供借鉴.
Research and development on the growth environment of the young tree of Pinus koraiensis in Pinus koraiensis-broadleaved mixed forest
红松混交林中红松幼树生长环境的研究进展及展望
Population structure and dynamics of Pinus koraiensis seedlings regenerated from seeds in a montane region of eastern Liaoning Province, China
辽东山区天然更新红松幼苗种群结构与动态
Impact of tree harvesting on the population structure and dynamics of Pinus koraiensis (Pinaceae)
采伐对红松种群结构与动态的影响
Population structure and dynamics of Juniperus rigida in different regions of Loess Plateau
黄土高原不同地区杜松种群结构与动态
Population structure characteristics of Picea purpurea in the upstream of Taohe River
DOI:10.17521/cjpe.2019.0077
[本文引用: 1]
<p id="C2"><strong><i>Aims</i></strong> Natural population of <i>Picea purpurea</i> in Gahai-Zecha Nature Reserve and Kache and Yeliguan forest regions in the Taohe River upstream was selected, to reveal the demographic status of <i>P. purpurea </i>populations. The results from the current study will provide theoretical basis for the protection, management and structural restoration of <i>P. purpurea</i>.<br><strong><i>Methods</i></strong> Based on field surveys and data analysis, population structure diagram was drawn, static life table was designed, and survival curve of <i>P. purpurea</i> was fitted and analyzed. Quantitative methods were used to study the population dynamics. The population development trend was predicted by time-sequence model.<br><strong><i>Important findings</i></strong> In the three forest regions, <i>P. purpurea</i> population showed signs of strong natural regeneration ability with abundant seedling and sapling reserves, yet the mortality rate of saplings was high. The population in Gahai-Zecha forest manifested a Deevey-III type survival curve representing a stable population with a complete population structure, and the population in Kache forest and Yeliguan forest manifested Deevey-II type survival curves with partial declines present in both forests. With population dynamics indexes (<i>V<sub>pi</sub></i>) > 0, the populations of all three forests were in growth. The growth potentials ranking from highest to lowest were Gahai-Zecha > Kache > Yeliguan. The <i>P. purpurea</i> population in Kache forest showed the highest sensitivity to random interference followed by that in Yeliguan forest while Gahai-Zecha population was the most stable. This study showed that competition and self-thinning were the common causes of low survival rate of <i>P. purpurea</i> saplings and that different living situation in the three forests reflected the difference of survivability of <i>P. purpurea</i> populations in different habitats and life histories. The key to regeneration and development of <i>P. purpurea</i> populations was to protect the habitat of seedlings and to improve their quality and survival rate. The <i>P. purpurea</i> population in Gahai-Zecha forest grew well and the survivorship of that population was mainly affected by its own biological characteristics and environmental factors. The population structure of Kache forest was damaged under human interference. The population structure of Yeliguan forest was unstable due to its limited distribution. Appropriate anthropogenic interference measures should be adopted to promote population renewal and growth.</p>
洮河上游紫果云杉种群结构特征
DOI:10.17521/cjpe.2019.0077
[本文引用: 1]
该研究以洮河上游尕海-则岔自然保护区、卡车林区和冶力关林区的紫果云杉(Picea purpurea)天然种群为研究对象, 通过样地调查和数据统计, 绘制种群结构图, 编制静态生命表, 拟合并分析存活曲线, 运用数量化方法研究种群动态, 揭示种群生存现状, 预测种群发展趋势, 以期为该物种的保护、管理及结构恢复提供理论依据。结果显示: 3个林区紫果云杉种群自然更新能力强, 幼苗、幼树储量丰富, 幼小龄期死亡率普遍偏高; 尕海-则岔种群存活曲线符合Deevey-III型, 种群稳定结构完整, 卡车林区和冶力关林区种群存活曲线均符合Deevey-II型, 且均出现了局部衰退; 3个林区种群动态指数均大于0, 说明种群均属于增长型, 增长潜力为尕海-则岔>卡车>冶力关; 受随机干扰时卡车林区紫果云杉最敏感, 冶力关次之, 尕海-则岔种群最稳定。该研究表明: 竞争和自疏作用是造成紫果云杉幼小龄级个体存活率偏低的普遍因素, 3个林区不同的生存状况反映了紫果云杉种群在不同生境及生活史下生存能力的差异, 保护幼苗生存环境并提高幼苗质量和存活率是种群更新和发展的关键。尕海-则岔紫果云杉生存良好, 种群生存状况主要受自身生物学特性和环境因子的影响; 卡车林区主要受人为影响, 种群结构遭到破坏; 冶力关林区受分布限制, 造成种群结构不稳定, 须采取一定的人工措施来促进种群更新与增长。

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}