


在21世纪,随着社会经济的快速发展, 由人类活动导致的物种灭绝和生态系统结构简化不断加剧, 生物多样性在人为的驱动下以一种前所未有的速度发生改变, 使得生态系统功能向难以预测的方向发展(雷羚洁等, 2016)。因此, 生物多样性的改变对生态系统功能的影响成为越来越多研究者探索的重点方向(李静鹏等, 2016)。生态系统最为重要的价值是提供多种生态功能和服务, 如同时提供土壤有机碳蓄积、净初级生产力、碳循环和氮循环等多种不同的功能或服务, 即生态系统多功能性(ecosystem multifunctionality, EMF) (Roscher et al., 2012)。由于不同生态系统功能之间存在一定的权衡关系, 与单一生态系统功能(single ecosystem function, SEF)相比, EMF作为一个综合性考量指标, 能更全面地帮助我们从整体上理解和明确生物多样性与生态系统功能间的关系(Gamfeldt et al., 2008)。因此相较于植物多样性和单一生态系统功能之间的关系, 研究植物多样性和EMF间的关系显得更为重要。但就当下研究情况来看, 大多数研究仅关注于生物多样性(物种多样性)与SEF的关系(例如: 氮累积、生产力等), 评价和研究生态系统多种功能的研究工作鲜有报道(刘旻霞等, 2022; 孙龙等, 2022)。生态系统功能不仅依赖于物种的数目, 更依赖于物种所具有的功能性状; 相比物种多样性, 功能多样性对于生态系统功能的贡献更大, 对功能多样性的定量测定可以准确反映群落功能对生态系统多功能发展的作用(李瑞新等, 2016)。因此, 在面对和解决与生态系统功能相关的研究中, 越来越多的学者提倡用功能性状的多样性来代替物种多样性(Aiba et al., 2012; Khalil et al., 2018; Huang et al., 2019)。这是因为功能多样性作为植物多样性中的一个重要组成部分, 不仅能有效地反映与生态系统多功能间的关系, 而且可以更深入地了解功能多样性在解释生态系统功能方面的优势(刘旻霞等, 2022)。因此, 通过研究功能多样性和EMF之间的关系, 可以更清晰、细致地阐释生态系统功能的变化。
巴音布鲁克高寒草甸位于天山南坡, 对维持高海拔地区生态平衡、养分循环以及生物多样性等具有重要意义, 其在维系我国西北干旱区流域生态环境安全等方面具有重要作用。与其他地区相比, 该地区草地生态系统更为脆弱、敏感。因其海拔在2 000-3 600 m范围, 有一定的高差, 所以是研究生物多样性海拔格局的理想场所(刘旻霞等, 2017; 郭婧等, 2020)。目前, 该区域对生物多样性-生态系统功能关系研究多集中于物种多样性层次, 并且仅考虑其与单一的生态系统功能(如生产力、土壤有机碳蓄积、土壤养分循环等)的关系(常凤, 2021), 至今功能多样性与EMF在不同生态系统中的关系格局仍未得到解决, 而且在高寒草甸生态系统中的相关研究还较为缺乏。为此, 本研究选取巴音布鲁克高寒草甸海拔梯度上的植物群落为研究对象, 分析在不同海拔高度上功能多样性与EMF的分布规律及相互间的关系, 以期进一步加深对植物群落功能多样性与高寒草甸EMF间关系的认识, 为草地生物多样性及生态系统多功能保护提供科学依据。
1 材料和方法
1.1 研究区概况
巴音布鲁克高寒草甸地处83.70°-85.85° E、42.98°-43.12° N, 海拔范围为2 000-3 600 m, 属于典型的高寒草甸, 全年积雪日达150-180天, 冬季长, 夏季短, 最高气温为30.5 ℃, 最低气温可达-48 ℃, 年平均气温-4.8 ℃, 年降水量276 mm, 年蒸发量1 248 mm, 无绝对无霜期。该研究区域草地类型主要有高山草甸、亚高山草甸、亚高山草原化草甸等。
1.2 野外样地设置和植物群落调查
于2020年8-9月在巴音布鲁克山区进行植被和土壤的调查, 选择植被过渡明显的5个海拔高度(2 200、2 400、2 600、2 800、3 000 m), 每个海拔处选取坡向、土地利用强度、人为干扰尽可能相同的3个地点设置研究样地(20 m × 20 m), 在每个样地的对角线上设置3个5 m × 5 m的样方, 在每个样方内按梅花状设置5个1 m × 1 m的小样方。5个海拔共设置了15个样地, 225个样方。记录各个样方的生物信息(如: 植被类型、物种数量、盖度、高度等)和地理信息(如: 海拔、经度、纬度等) (表1)。在每个小样方内, 选取样方中盖度比较大的物种, 采集10株正常发育、无病虫害的健康植株(未达到10株的个体在样方附近采集), 在每个物种的不同高度随机采取10-15片成熟完整的叶片, 装入保鲜箱内带回实验室。将样方内植物齐地刈割, 翻土后获取植株地下部分, 将样品立即冲洗漂净并将根系分离, 在野外风干后带回实验室。在每个样方的中心区域取0-30 cm土壤样品200 g带回实验室, 在室温条件下将样地中采集的土样自然风干, 剔除土壤中杂质, 物理研磨后过60目筛, 分装密封袋中用于后续指标的测定(黄超等, 2020)。此外, 在样方内挖掘土壤剖面, 用直径为7.5 cm的环刀获取土壤样品(0-30 cm), 用以测定土壤密度及砾石比。
表1 巴音布鲁克高寒草甸各样地地理信息列表
Table 1
| 编号 Plot No. | 海拔 Altitude (m) | 经度 Longitude (° E) | 纬度 Latitude (° N) | 坡向 Aspect | 坡度 Slope (°) |
|---|---|---|---|---|---|
| 2200-1 | 2 194.12 | 83.86 | 42.35 | 阳坡 Sunny slope | 13.2 |
| 2200-2 | 2 203.71 | 83.86 | 42.34 | 阳坡 Sunny slope | 3.1 |
| 2200-3 | 2 210.63 | 83.85 | 42.34 | 阳坡 Sunny slope | 5.4 |
| 2400-1 | 2 406.20 | 83.53 | 42.68 | 阳坡 Sunny slope | 7.2 |
| 2400-2 | 2 400.90 | 83.53 | 42.68 | 阳坡 Sunny slope | 12.1 |
| 2400-3 | 2 415.20 | 84.39 | 43.05 | 阳坡 Sunny slope | 10.7 |
| 2600-1 | 2 590.42 | 84.05 | 43.05 | 阳坡 Sunny slope | 7.9 |
| 2600-2 | 2 602.60 | 82.97 | 42.86 | 阳坡 Sunny slope | 9.4 |
| 2600-3 | 2 597.20 | 84.05 | 42.85 | 阳坡 Sunny slope | 6.7 |
| 2800-1 | 2 794.62 | 82.99 | 42.86 | 阳坡 Sunny slope | 3.9 |
| 2800-2 | 2 817.50 | 82.99 | 42.87 | 阳坡 Sunny slope | 8.4 |
| 2800-3 | 2 802.00 | 82.97 | 42.87 | 阳坡 Sunny slope | 17.5 |
| 3000-1 | 3 052.70 | 83.47 | 42.51 | 阳坡 Sunny slope | 10.6 |
| 3000-2 | 3 062.10 | 83.47 | 42.51 | 阳坡 Sunny slope | 9.5 |
| 3000-3 | 3 004.12 | 83.03 | 42.51 | 阳坡 Sunny slope | 7.7 |
1.3 实验方法
采用CI-203手持式激光叶面积仪(CID, Waltham, USA)测定叶面积; 选用精度为0.000 1 g的电子天平称量叶片鲜质量, 然后将新鲜叶片样品置于60 ℃的烘箱中烘干至恒质量, 时间一般为72 h, 并称取叶片干质量(LDM)。计算比叶面积(SLA)、叶片含水量(LWC)和叶干物质含量(LDMC)。用HClO4-HsSO4消化法测量叶片氮(N)含量; 用酸溶-钼锑抗比色法测定叶片磷(P)含量; 用重铬酸钾容量-外加热法测定叶片碳(C)含量。
将获取的植株地上、地下部分放入烘箱, 在 105 ℃下杀青30 min, 之后在65 ℃下烘48 h以上直至恒质量并称量。
将处理过的土样参照土壤农化分析进行土壤养分的测定(鲍士旦,2005), 土壤有机碳(SOC)含量采用重铬酸钾容量-外加热法测定; 全氮(TN)含量采用HClO4-H2SO4消化法, 使用福斯1035全自动定氮仪(KDN-102F, 福斯, 上海)测定; 铵态氮(AN)含量采用2 mol·L-1 KCl浸提-靛酚蓝比色法测定; 硝态氮(NN)含量采用双波长紫外分光光度法测定; 全磷(TP)含量采用酸溶-钼锑抗比色法, 使用安捷伦CARY60紫外分光光度计(Agilent, Santa Clara, USA)测定; 速效磷(AP)含量采用NaOH浸提-钼锑抗比色法, 使用安捷伦CARY60紫外分光光度计测定; 全钾(TK)用NaOH溶解后采用火焰光度法测定含量; 速效钾(AK)含量采用火焰原子吸收光谱法测定。土壤密度样品在65 ℃恒温烘干后称量, 使用烘干后样品测定土壤砾石比。通过平均SOC含量与土壤密度计算单位体积土壤碳含量(USCA, kg·m-3), 计算公式如下:
式中, TOC指计算深度内土壤平均有机碳含量(%); Δd为所取土壤深度(m); ρ为土壤密度(g·cm-3); S为环刀底面积(π × 0.03752 cm2)。
1.4 功能特征数据的获取和功能多样性指数的计算
式中, SFce为群落e内性状c所占据的生态位空间, Rc为性状c的绝对值, S为物种丰富度, PEWi为物种i的加权均匀度, aj为物种j的多度, zj为物种j到加权质心的距离, dij为物种i和物种j的功能性状差异程度, Pi和Pj为物种i和j的相对多度。
1.5 生态系统功能指标的选取和EMF计算
结合以往的研究, 现在对EMF的量化评定主要有阈值法和指标权重平均值法。阈值法是将所有功能的作用都视为等效, 无法反映特定生态系统功能或服务的重要性, 因没有确定的标准阈值, 计算的生态系统功能值会随阈值的选择而变化, 容易受到研究者主观态度的影响(Byrnes et al., 2014; 孙龙等, 2022); 相比阈值法, 指标权重平均值法计算得到的EMF指数具有较好的统计学特性, 计算简便, 不会改变原始数据的变异程度, 且能够清晰直观地表征植物群落提供多种生态系统功能和服务的能力(Bynes et al., 2014), 因此本研究使用指标权重平均值法来量化EMF。选取10个生态系统功能指标综合量化多个SEF。其中采用地上生物量和地下生物量来表征植物生长功能(熊定鹏等, 2016), 单位土壤碳含量用于表征土壤有机碳蓄积功能, 土壤全氮、氨态氮、硝态氮、全磷、速效磷、全钾和速效钾含量用于表征土壤养分循环功能(Hou et al., 2021)。随后再对选取的10个生态系统功能指标参数使用指标权重平均值法来量化EMF。量化EMF前需对各样方内的各功能指标参数进行标准化处理, 然后再将15个调查样地中10种生态系统功能指标参数转化为标准Z分数。Z值计算公式如下:
式中, Zxy为样地x第y种生态系统功能参数的Z分数, x的范围在1-15, y的范围在1-10; Fxy为样地x第y种生态系统功能参数的数值; μy为第y种生态系统功能参数在15个样地间的平均值; σy为第y种生态系统功能参数在15个样地间的标准差。对每一样地10种不同生态系统功能参数的Z分数求其综合平均值, 即为各样地的EMF, EMF值计算公式如下:
1.6 数据处理
在Excel 2019中进行数据整理。计算各海拔优势种的重要值(即相对盖度、相对多度和相对高度三者之和的加权平均); 利用R 4.1.3 “FD”程序包中dbFD函数计算功能多样性指数; 采用单因素方差分析检验物种功能多样性指数在海拔梯度上是否具有显著性差异; 采用Pearson相关性分析及一元线性回归分析功能多样性指数与EMF指数间的关系。此外, 用10个生态系统功能指标间的组合权重平均值量化植物生长、养分循环和土壤有机碳蓄积这3种重要的生态系统功能, 并将这3种生态系统功能参数量化后产生的3种SEF分别与不同功能多样性指数进行相关性分析与回归分析; 考虑到功能多样性各指数与海拔间可能存在交互作用, 对EMF可能产生影响, 故采用R 4.1.3中的“plyr”包进行协方差分析, 探讨海拔与功能多样性指数间是否存在交互作用; 同时利用“lavaan”包构建结构方程模型, 分析海拔、功能多样性与EMF间的关系, 以揭示海拔对功能多样性和EMF的影响。
2 结果和分析
2.1 植物群落物种组成及丰富度沿海拔梯度的变化
在海拔2 194-3 062 m范围内的15个样地中共调
查到52种物种, 隶属于20科26属。由表2可知, 低海拔地区(2 200-2 400 m)和高海拔地区(2 800-3 000 m)植物群落物种组成差异较大。低海拔地区主要以线叶嵩草(Kobresia capillifolia)、丘陵老鹳草(Geranium collinum)、多裂委陵菜(Potentilla multifida)、茸毛委陵菜(Potentilla strigosa)为主。随着海拔的升高群落优势种发生了更替, 逐渐变为珠芽蓼(Polygonum vivipara)占优势。除此之外, 随着海拔上升, 物种丰富度呈单峰变化趋势, 峰值(16.13 ± 3.14)出现在海拔2 600 m处。
表2 巴音布鲁克高寒草甸不同海拔样地植物群落优势种组成
Table 2
| 海拔 Altitude (m) | 物种丰富度 (平均值±标准差) Specie richness (mean ± SD) | 优势种(重要值) Dominant species (importance value) |
|---|---|---|
| 2 200 | 15.00 ± 2.00a | 薹草 Carex sp. (0.212 5) |
| 线叶嵩草 Kobresia capillifolia (0.187 5) | ||
| 2 400 | 14.49 ± 2.33ab | 丘陵老鹳草 Geranium collinum (0.212 2) |
| 线叶嵩草 Kobresia capillifolia (0.186 3) | ||
| 2 600 | 16.13 ± 3.14ab | 线叶嵩草 Kobresia capillifolia (0.251 6) |
| 薹草 Carex sp. (0.145 8) | ||
| 2 800 | 15.17 ± 1.47a | 珠芽蓼 Polygonum viviparum (0.276 5) |
| 针茅 Stipa sp. (0.266 0) | ||
| 3 000 | 12.25 ± 2.87b | 珠芽蓼 Polygonum viviparum (0.349 0) |
| 丘陵老鹳草 Geranium collinum (0.188 1) |
不同小写字母表示差异显著(p < 0.05)。
Different lowercase letters indicate significant difference (p < 0.05).
2.2 植物群落功能多样性沿海拔梯度的变化
由图1可知, 随海拔升高, 功能多样性指数发生显著变化。FRic由大到小为2 800 m > 2 600 m > 2 400 m > 2 200 m > 3 000 m, 呈现出单峰变化趋势; 海拔2 200 m处与海拔2 600 m、2 800 m处的FRic存在极显著差异, 2 800m处与3 000m处存在极显著差异。FDis与FRic的变化趋势相同, 峰值出现在2 800 m处, 海拔2 800m处与3 000m处存在极显著差异。不同海拔段FEve值的大小为2 600 m > 2 800 m > 2 400 m > 2 200 m > 3 000 m, 大部分海拔段间存在显著差异。除此之外, 随着海拔的增加, Rao’Q逐渐降低, 海拔2 200 m与2 600m、2 800m、3 000m处均存在极显著差异,2 400m处与3 000m处存在极显著差异。
图1
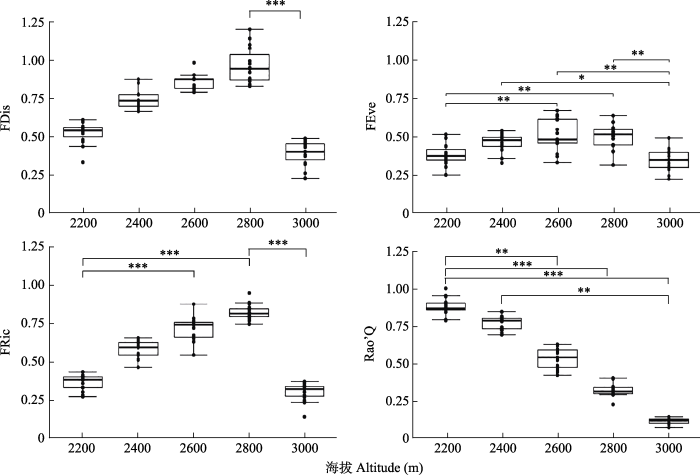
图1
巴音布鲁克高寒草甸不同海拔样地植物群落的功能多样性指数(平均值±标准差)。FDis, 功能分散度指数; FEve, 功能均匀度指数; FRic, 功能丰富度指数; Rao’Q, Rao的二次熵。*, p < 0.05; **, p < 0.01; ***, p < 0.001。
Fig. 1
Distribution of functional diversity index of plant communities at different altitudes in Bayanbulak alpine meadow (mean ± SD). FDis, functional dispersion index; FEve, functional evenness index; FRic, functional richness index; Rao’Q, Rao’s quadratic entropy. *, p < 0.05; **, p < 0.01; ***, p < 0.001.
2.3 功能多样性指数与SEF、EMF指数间的关系
图2
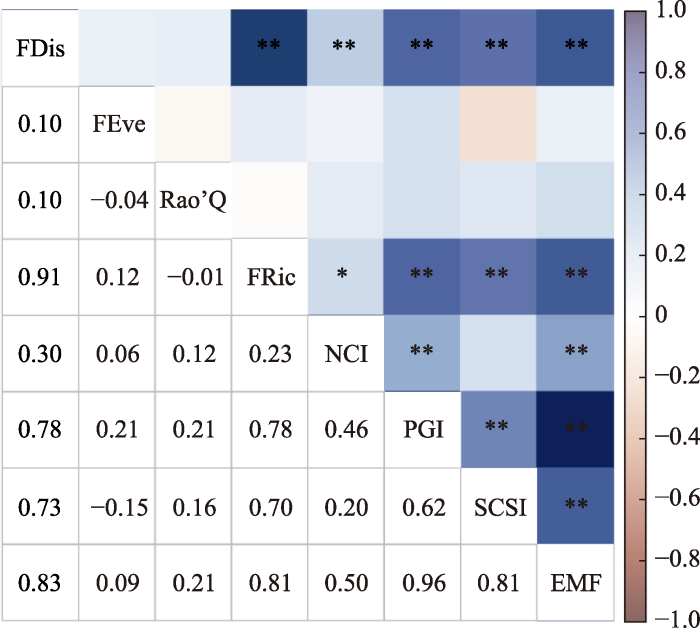
图2
巴音布鲁克高寒草甸群落功能多样性指数与单一生态系统功能(SEF)、生态系统多功能性(EMF)指数间的相关性分析。FDis, 功能分散度指数; FEve, 功能均匀度指数; FRic, 功能丰富度指数; NCI, 养分循环指数; PGI, 植物生长指数; Rao’Q, Rao的二次熵; SCSI, 土壤有机碳蓄积指数。*, p < 0.05; **, p < 0.01; ***, p < 0.001。
Fig. 2
Correlation analysis between community functional diversity index and single elosystem function (SEF) and elosystem multifuctionality (EMF) index in Bayanbulak alpine meadow. FDis, functional dispersion index; FEve, functional evenness index; FRic, functional richness index; NCI, nutrient cycling index; PGI, plant growth index; Rao’Q, Rao’s quadratic entropy; SCSI, soil carbon stock index. *, p < 0.05; **, p < 0.01; ***, p < 0.001.
图3
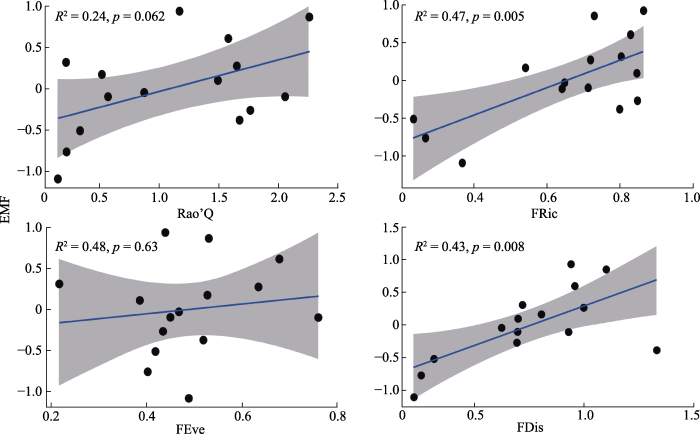
图3
巴音布鲁克高寒草甸群落功能多样性指数与生态系统多功能性指数(EMF)间的回归分析。FDis, 功能分散度指数; FEve, 功能均匀度指数; FRic, 功能丰富度指数; Rao’Q, Rao的二次熵; 灰色部分代表置信区间。
Fig. 3
Regression analysis of community functional diversity index and ecosystem multifunctional index (EMF) in Bayanbulak alpine meadow. FDis, functional dispersion index; FEve, functional uniformity index; FRic, functional richness index; Rao’Q, Rao’s quadratic entropy; the gray part represents the confidence interval.
考虑到不同海拔间各生态系统功能指数与功能多样性指标存在自相关性, 为检验上述结果, 将功能多样性指标和生态系统功能指标分海拔整理后进行相关性分析(表3)可知, 不同海拔上植物功能多样性指数与生态系统功能指数(SEF、EMF)相关性与上述结果类似, 各海拔样地FRic、FDis与大部分生态系统功能性指数间存在显著相关关系, 仅在海拔2 600 m处FEve与养分循环指数、土壤有机碳蓄积指数存在显著相关性; 海拔3 000 m处Rao’Q指数与养分循环、有机碳蓄积、EMF均存在显著相关性。
表3 巴音布鲁克高寒草甸不同海拔功能多样性指数与生态系统多功能性指数(EMF)间相关性分析
Table 3
| 海拔 (m) Altitude | 生态系统功能指数 Ecosystem function index | 功能多样性指数 Functional diversity index | |||
|---|---|---|---|---|---|
| FRic | FDis | FEve | Rao’Q | ||
| 2 200 | EMF | 0.025 0* | 0.882 9* | 0.139 4 | 0.352 2* |
| 植物生长 Plant growth | 0.984 9* | 0.947 7* | 0.904 3 | -0.705 4 | |
| 养分循环 Nutrient cycling | 0.852 0* | 0.638 2* | 0.890 6 | -0.707 8 | |
| 有机碳蓄积 Soil carbon stock | 0.918 4*** | 0.820 7* | 0.934 5 | -0.577 2 | |
| 2 400 | EMF | 0.957 3** | 0.926 7* | -0.280 9 | 0.162 8 |
| 植物生长 Plant growth | 0.938 5*** | 0.843 0*** | -0.248 2 | 0.201 0 | |
| 养分循环 Nutrient cycling | 0.867 3*** | 0.968 4*** | -0.323 6 | 0.011 4 | |
| 有机碳蓄积 Soil carbon stock | -0.568 1* | -0.642 5** | -0.186 3 | 0.137 0 | |
| 2 600 | EMF | 0.080 7* | 0.244 9 | 0.265 8 | 0.152 0 |
| 植物生长 Plant growth | 0.099 7* | 0.004 5 | -0.100 6 | 0.176 0 | |
| 养分循环 Nutrient cycling | 0.168 0 | 0.078 8** | 0.262 3** | 0.184 9 | |
| 有机碳蓄积 Soil carbon stock | 0.171 4** | 0.080 3* | 0.257 2* | 0.268 7 | |
| 2 800 | EMF | 0.585 7** | 0.222 7 | 0.373 7 | -0.449 0 |
| 植物生长 Plant growth | 0.345 3 | 0.194 2 | -0.004 2 | -0.238 8 | |
| 养分循环 Nutrient cycling | 0.961 4*** | 0.504 5 | 0.203 5 | -0.161 9 | |
| 有机碳蓄积 Soil carbon stock | 0.523 5* | 0.220 6 | 0.495 6 | -0.486 0 | |
| 3 000 | EMF | 0.868 2* | 0.918 5* | -0.044 4 | 0.695 3* |
| 植物生长 Plant growth | 0.952 1** | 0.975 1*** | 0.054 0 | 0.690 9 | |
| 养分循环 Nutrient cycling | 0.782 2** | 0.838 6*** | -0.267 6 | 0.668 2** | |
| 有机碳蓄积 Soil carbon stock | 0.856 8* | 0.927 4** | 0.005 6 | 0.734 5** | |
*, p < 0.05; **, p < 0.01; ***, p < 0.001。FDis, 功能分散度指数; FEve, 功能均匀度指数; FRic, 功能丰富度指数; Rao’Q, Rao的二次熵。
*, p < 0.05; **, p < 0.01; ***, p < 0.001. FDis, functional dispersion index; FEve, functional uniformity index; FRic, functional richness index; Rao’Q, Rao’s quadratic entropy.
2.4 海拔对植物群落功能多样性与EMF的影响
通过协方差分析可知, 除FDis与FRic间存在较为显著的交互作用(F = 5.251 2, p < 0.05)外, 其余功能多样性指数间以及与海拔变量间的交互作用并不明显。通过两种独立的结构方程模型(完全中介: 海拔对EMF的影响仅通过功能多样性指数完全传递; 部分中介: 海拔对EMF的影响分为以功能多样性指数为中介的间接影响和直接影响)同时进行海拔与植物群落功能多样性指数对EMF影响的拟合, 由结果(表4)可知, 完全中介模型的自由度(df)为15, 拟合优度指数(GFI)为0.92, 标准化均方根残差为0.08, 相比部分中介模型具有较好的拟合度。同时, 为验证海拔对植物群落功能多样性和EMF的影响程度, 构建结构方程模型进行进一步的验证(图4)。在完全中介模型中, 海拔对FRic存在极显著的正向直接效应(标准路径系数(Est.std) = 0.067), 同时FRic对EMF存在极显著积极影响(Est.std = 0.475); 海拔对Rao’Q影响也极为显著(Est.std = -0.980), 同时Rao’Q对EMF有显著积极影响(Est.std = 0.05)。这表明在本研究样地, 海拔的中介效应可通过FRic和Rao’Q传递给EMF。除此之外, 海拔对FEve和FDis表现出不显著的负效应。但FEve、FDis对EMF均具有极显著的正向直接效应, 其Est. std依次为0.127、0.283。
表4 巴音布鲁克高寒草甸海拔对功能多样性与生态系统多功能(EMF)影响的结构方程模型概述
Table 4
| 关系 Regression | 完全中介模型 Full mediation model (df = 15, SRMR = 0.08, GFI = 0.92) | 部分中介模型 Partial mediation model (df = 13, SRMR = 0.08, GFI = 0.81) | ||||
|---|---|---|---|---|---|---|
| Est. std | SE | p | Est. std | SE | p | |
| Alt-FRic | 0.067 | 0.127 | 0.008 | 0.004 | 0.059 | 0.952 |
| Alt-FEve | -0.044 | 0.111 | 0.147 | -0.015 | 0.029 | 0.606 |
| Alt-FDis | -0.049 | 0.129 | 0.309 | -0.080 | 0.058 | 0.165 |
| Alt-Rao’Q | -0.980 | 0.023 | <0.001 | -0.980 | 0.023 | 0.000 |
| FRic-EMF | 0.475 | 0.275 | <0.001 | 0.506 | 0.161 | 0.002 |
| FEve-EMF | 0.127 | 0.089 | <0.001 | 0.143 | 0.084 | 0.087 |
| FDis-EMF | 0.283 | 0.274 | <0.001 | 0.233 | 0.167 | 0.163 |
| Rao’Q-EMF | 0.050 | 0.570 | 0.017 | 0.067 | 0.068 | 0.325 |
| Alt-EMF | 0.007 | 0.400 | 0.857 | |||
Alt, 海拔; FDis, 功能分散度指数; FEve, 功能均匀度指数; FRic, 功能丰富度指数; Rao’Q, Rao的二次熵, df, 自由度; Est.std, 标准路径系数; GFI, 拟合优度指数; p, 给定模型的数据概率; SE, 标准误; SRMR, 标准化均方根残差。
Alt, altitude; FDis, functional dispersion index; FEve, functional uniformity indes; FRic, functional richness index; Rao’Q, Rao’s quadratic entropy; df, degree of freedom; Est.std, standard path coefficient; GFI, goodness-of-fit index; p, probability for a given model; SE, standard error; SRMR, standardized root mean square residual.
图4
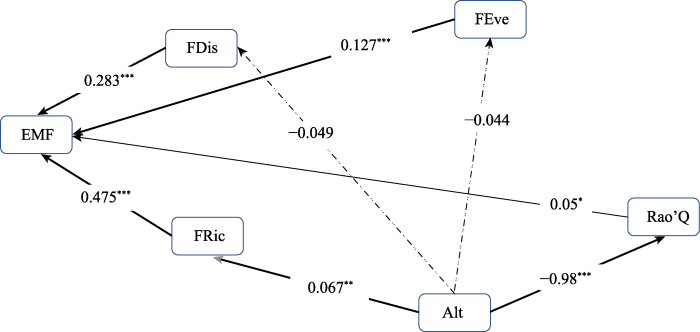
图4
巴音布鲁克高寒草甸海拔对功能多样性指数和态系统多功能性指数(EMF)影响的结构方程模型图。Alt, 海拔; FDis, 功能分散度; FEve, 功能均匀度; FRic, 功能丰富度; Rao’Q, Rao的二次熵。箭头旁边的数字为标准化路径系数(正值代表正效应, 负值表示负效应)。实线代表存在显著相关性,虚线代表不存在显著相关性。箭头的粗细代表显著性差异(粗代表显著性高, 细代表显著性低)。*, p < 0.05; **, p < 0.01; ***, p < 0.001。
Fig. 4
Structural equation model diagram of the effect of altitude on functional diversity index and ecosystem multifunctional index (EMF) in Bayanbulak alpine meadow. Alt, altitude; FDis, functional dispersion; FEve, functional uniformity; FRic, functional richness; Rao’Q, Rao’s quadratic entropy. The number next to the arrow in the plot is the normalized path coefficient (positive values represent positive effects, negative values represent negative effects). A solid line indicates a significant correlation, and a dashed line indicates no significant correlation. The thickness of the arrow represents significant difference (thick represents high significance, thin represents low significance). *, p < 0.05; **, p < 0.01; ***, p < 0.001.
3 讨论
3.1 植物群落组成与功能多样性沿海拔梯度的变化
海拔是影响山地植物分布的重要因子, 海拔的变化通常会引起水热条件、土壤养分等多种环境因子发生变化, 造成环境的异质性, 进一步影响了植物群落的结构、群落稳定性和多样性。在本研究中, 不同海拔样方内植物群落类型的物种组成和结构存在差异。物种丰富度指数沿海拔变化显著, 呈现“单峰型”变化趋势, 在中海拔2 600 m处物种丰富度指数达到最大, 这与刘旻霞等(2022)对海拔梯度上甘南高寒草甸功能多样性与EMF关系研究结果一致。主要是因为低海拔热量足够, 环境条件相对较好, 但存在水分蒸发量较高现象; 同时, 随着优势物种的出现, 群落中大量的生态位被其占据; 此外, 低海拔段(2 200-2 400 m)存在相对较强的人为干扰, 造成物种缺失; 高海拔段(2 800-3 000 m)虽然水分含量有所增加, 但是风速大,温度低, 生境较为严苛, 环境筛选对该地区群落构建起到决定性作用, 呈现出物种丰富度较低的现象; 而在中海拔段(2 600-2 800 m), 人为干扰程度较低, 水热条件相较于高、低海拔更为均衡, 适宜植物生长, 因此物种丰富度相对较高。
功能多样性可清晰直观地表征群落中物种的分布与生态系统功能间的关系, 为生态系统平衡提供理论基础(Klimešáová et al., 2008)。FRic通过量化群落中现有物种占据的生态位空间大小来反映植被对资源的利用率(Aiba et al., 2012)。本研究表明, 由于环境因子与群落物种组成间相互作用, 导致随着海拔高度的增加, FRic总体呈现先上升后下降的“单峰”趋势。中低海拔范围内(2 200-2 600 m)随着物种丰富度增加, 种间竞争越发激烈, 导致生态位分化, 物种性状分化的范围也随之增加。高海拔段物种丰富度降低, 种间竞争强度下降; 同时由于严苛的生境条件, 环境筛的作用愈发显著, 促使物种的功能性状发生趋同化现象, 功能丰富度指数降低, 出现不同程度的生态位重叠现象, 这在一定程度上会降低植物群落物种的功能多样性。
FEve可衡量物种在空间中的特征分布, 是衡量生产力恢复、抵御外来物种入侵的重要指标(Zhang et al., 2021)。本研究中, FEve随着海拔的上升呈现出“单峰”变化趋势, 这与秦浩等(2019)在山西关帝山森林群落物种、谱系和功能多样性海拔格局的研究中得到的结论相同。出现这种现象可能是因为在海拔2 600 m处物种丰富度达到最高, 同时, 该海拔段处于过渡段, 群落组成兼有适应高海拔和低海拔不同环境的物种, 表现出较高的均匀度; 对于其他海拔段而言, 低海拔段(2 200-2 400 m)由于水热条件和人为干扰的影响, 高海拔段(2 800-3 000 m)由于有环境筛的影响, 导致FEve均较低。
Rao’Q指数表示群落生态位的重叠程度。物种的生态位重叠程度越强, 资源竞争越强, Rao’Q指数较低(董世魁等, 2019)。本研究中, Rao’Q指数随海拔上升呈现下降的趋势, 这与刘旻霞等(2022)针对甘南高寒草甸海拔梯度上功能多样性与生态系统多功能关系的研究结果相同。低海拔段(2 200-2 400 m) Rao’Q指数较高是因为该地区植物具有较高的资源利用效率, 所以种间、种内资源竞争强度较弱; 相较于中、低海拔段(2 200-2 600 m), 高海拔段(2 800-3 000 m)环境相对恶劣, 物种丰富度相对较低, 同时为了适应该地区的环境, 各种植物产生趋同进化效应, 演变出相似的形态结构和功能特征, 导致互补性降低, 植物群落物种趋于单一, 对资源的利用效率减弱, 资源竞争性增强。
3.2 植物群落功能多样性与EMF的关系
在巴音布鲁克高寒草甸, 不同海拔功能多样性指数与EMF间关系格局存在部分差异。本研究发现, 功能多样性指数均与EMF呈正相关关系。其中FRic与EMF间相关性最强且线性回归关系最显著, 是维持较高EMF的一个重要元素。除此之外, FDis与EMF也存在显著正相关关系。研究结果表明, 巴音布鲁克高寒草甸EMF主要受群落功能数量和功能分散程度的影响, 而非功能性状分布的均匀程度影响。本研究结果与刘旻霞等(2022)针对甘南高寒草甸海拔梯度上功能多样性与EMF的关系所得结果相一致, 表明群落功能多样性越高, 植物功能性状的中间变异程度越高, 功能性状越分散, 生态位分化越强, 这使得植物群落物种能够充分利用环境资源; 而FRic越高, 个体物种特征叠加后, 群落整体功能特征越丰富, 生态位将被完全占据, 生态系统功能将更加稳定。研究结果验证了在干旱草地生态系统中互补效应作为植物群落功能多样性与EMF关系维持的一种重要机制, 同时也支持了Huang等(2019)在对功能多样性如何驱动云南松(Pinus yunnanensis)次生林EMF的研究中得到的结论: FRic相比于其他功能多样性指数可以更好地预测EMF。FEve可以衡量植物性状在所占用空间中的分布, 功能均匀度越高, 植物性状的分布规律性越强, 各种自然资源的利用程度越接近。在本研究中, 在2 600 m处FEve与有机碳蓄积之间存在极显著的相关关系, 表明群落功能多样性对生态系统功能有积极的影响, 这与艾比湖流域功能丰富度与有机碳蓄积显著相关的结果(许彧, 2017)相一致; 研究结果表明, 无论是高寒草甸还是热带雨林区, 功能均匀度对土壤有机碳蓄积都有显著影响。
3.3 功能多样性介导的海拔对EMF的影响
结构方程模型是一种功能强大的多元统计模型, 越来越多地出现在生态学研究中, 用于测试和评估多变量间的因果关系。结合Wondimu等(2021)的研究, 本研究假设海拔对EMF的影响可以通过功能多样性来介导, 因此使用了完全中介和部分中介两种模型来测试验证。本研究通过比较两种结构方程模型分析结果发现, 在考虑了环境驱动因素(海拔)的影响后, 相比于完全中介模型, 部分中介模型的拟合效果较差, 同时功能多样性指数对EMF的影响发生改变, 即减弱了两者之间的关系, 这与刘阳等(2021)针对生物多样性与环境驱动因素对内蒙古草原EMF变化的研究结果相同; 同时研究结果也验证了Wondimu等(2021)的物种多样性通过功能多样性对埃塞俄比亚常绿林中地上碳储量影响的结果, 即环境因素的引入会降低多样性对EMF的关系。这可能是因为功能多样性对EMF的部分影响被海拔变化所引起的环境因素与气候因素的改变所掩盖(Lepš, 2004), 因此, 在区域水平上, 海拔可能是巴音布鲁克草原EMF的关键驱动因素。通过结构方程模型也验证了该研究区EMF受生物因子中的FRic影响最为显著; 同时也表现出海拔可通过对FRic与Rao’Q施加影响, 进而间接影响到EMF。这表明海拔的改变会通过功能多样性这一媒介作用于EMF。研究结果同时也验证了生态位互补效应是维持EMF的重要机制(Cardinale et al., 2011; Wondimu et al., 2021)。
4 结论
综上所述, 在巴音布鲁克高寒草甸, 功能多样性会沿海拔梯度发生明显变化, 进而影响SEF和EMF; 功能多样性对于维持EMF具有重要意义, 除FEve与土壤有机碳蓄积间呈现出不显著的负相关外, 其他功能多样性指数均与SEF和EMF呈正相关关系, 其中FRic和FDis是影响EMF的重要因子, 也是维持EMF的关键元素; 表明维持EMF需要更丰富的物种功能性状特性和分散程度, 互补效应在高寒草甸EMF维持中更为重要。
致谢
感谢新疆师范大学博士启动基金项目(XJNUBS1813)和干旱区植物逆境生物学实验室的资助。
参考文献
Interspecific differences in determinants of plant species distribution and the relationships with functional traits
DOI:10.1111/jec.2012.100.issue-4 URL [本文引用: 2]
Investigating the relationship between biodiversity and ecosystem multifunctionality: challenges and solutions
DOI:10.1111/2041-210X.12143 URL [本文引用: 1]
The functional role of producer diversity in ecosystems
DOI:10.3732/ajb.1000364
PMID:21613148
[本文引用: 1]

Over the past several decades, a rapidly expanding field of research known as biodiversity and ecosystem functioning has begun to quantify how the world's biological diversity can, as an independent variable, control ecological processes that are both essential for, and fundamental to, the functioning of ecosystems. Research in this area has often been justified on grounds that (1) loss of biological diversity ranks among the most pronounced changes to the global environment and that (2) reductions in diversity, and corresponding changes in species composition, could alter important services that ecosystems provide to humanity (e.g., food production, pest/disease control, water purification). Here we review over two decades of experiments that have examined how species richness of primary producers influences the suite of ecological processes that are controlled by plants and algae in terrestrial, marine, and freshwater ecosystems. Using formal meta-analyses, we assess the balance of evidence for eight fundamental questions and corresponding hypotheses about the functional role of producer diversity in ecosystems. These include questions about how primary producer diversity influences the efficiency of resource use and biomass production in ecosystems, how primary producer diversity influences the transfer and recycling of biomass to other trophic groups in a food web, and the number of species and spatial /temporal scales at which diversity effects are most apparent. After summarizing the balance of evidence and stating our own confidence in the conclusions, we outline several new questions that must now be addressed if this field is going to evolve into a predictive science that can help conserve and manage ecological processes in ecosystems.
A handbook of protocols for standardised and easy measurement of plant functional traits worldwide
DOI:10.1071/BT02124
URL
[本文引用: 1]
There is growing recognition that classifying terrestrial plant species on the basis of their function (into 'functional types') rather than their higher taxonomic identity, is a promising way forward for tackling important ecological questions at the scale of ecosystems, landscapes or biomes. These questions include those on vegetation responses to and vegetation effects on, environmental changes (e.g. changes in climate, atmospheric chemistry, land use or other disturbances). There is also growing consensus about a shortlist of plant traits that should underlie such functional plant classifications, because they have strong predictive power of important ecosystem responses to environmental change and/or they themselves have strong impacts on ecosystem processes. The most favoured traits are those that are also relatively easy and inexpensive to measure for large numbers of plant species. Large international research efforts, promoted by the IGBP–GCTE Programme, are underway to screen predominant plant species in various ecosystems and biomes worldwide for such traits. This paper provides an international methodological protocol aimed at standardising this research effort, based on consensus among a broad group of scientists in this field. It features a practical handbook with step-by-step recipes, with relatively brief information about the ecological context, for 28 functional traits recognised as critical for tackling large-scale ecological questions.
Relationship between plant species diversity and functional diversity in alpine grasslands
高寒草地植物物种多样性与功能多样性的关系
Multiple functions increase the importance of biodiversity for overall ecosystem functioning
DOI:10.1890/06-2091.1
PMID:18543617
[本文引用: 1]
Biodiversity is proposed to be important for the rate of ecosystem functions. Most biodiversity-ecosystem function studies, however, consider only one response variable at a time, and even when multiple variables are examined they are analyzed separately. This means that a very important aspect of biodiversity is overlooked: the possibility for different species to carry out different functions at any one time. We propose a conceptual model to explore the effects of species loss on overall ecosystem functioning, where overall functioning is defined as the joint effect of many ecosystem functions. We show that, due to multifunctional complementarity among species, overall functioning is more susceptible to species loss than are single functions. Modeled relationships between species richness and overall ecosystem functioning using five empirical data sets on monocultures reflected the range of effects of species loss on multiple functions predicted by the model. Furthermore, an exploration of the correlations across functions and the degree of redundancy within functions revealed that multifunctional redundancy was generally lower than single-function redundancy in these empirical data sets. We suggest that by shifting the focus to the variety of functions maintained by a diversity of species, the full importance of biodiversity for the functioning of ecosystems can be uncovered. Our results are thus important for conservation and management of biota and ecosystem services.
Status and function improvement technology of the grassland ecosystem in the upper Yellow River Basin
DOI:10.11733/j.issn.1007-0435.2020.05.001
[本文引用: 1]
Grassland ecosystem is not only an important natural security barrier of natural system,but also the key support of animal husbandry development. The ecological quality of grassland in the upper reaches of the Yellow River is related to the ecological protection and regional sustainable development of the whole Yellow River Basin. However,in recent years,the grassland ecological environment in the upper reaches of the Yellow River has evolved in a direction that is not conducive to human development. Grassland degradation has accelerated the process of land desertification. At the same time,it has caused natural disasters and river cutoff,which has restricted the healthy and stable development of regional economy. In this paper,the present situation,causes and main characteristics of grassland degradation in important water conservation areas in the upper reaches of the Yellow River were summarized. The technical model of grassland ecological function improvement and grassland adaptive management mechanism were put forward,and the new technologies of grassland restoration and management in the future were prospected. The purpose of this study was to provide theoretical and technical support for ecological restoration and governance of typical water conservation areas such as Sanjiangyuan,Qilian Mountain and Gannan,so as to provide scientific reference for effectively promoting ecological protection and high-quality development of the Yellow River Basin and helping the country to build a sound ecological security system.
黄河上游草地生态现状及功能提升技术
DOI:10.11733/j.issn.1007-0435.2020.05.001
[本文引用: 1]
草地生态系统不仅是自然系统重要的天然安全屏障,也是畜牧业发展的关键依托。黄河上游草地生态质量关系到整个黄河流域的生态保护和区域可持续发展。然而,近年来黄河上游草地生态环境向着不利于人类发展的方向演化,草地退化问题日趋严重,加速了土地荒漠化进程,同时引发自然灾害、河流断流等问题,制约了地区经济健康稳定的发展。本文综述了黄河上游重要水源涵养区域草地退化现状、成因和草地退化的主要特征,提出了草地生态功能提升技术模式及草地适应性管理机制,并对未来草地恢复治理新技术进行了展望,旨在为三江源、祁连山和甘南等典型水源涵养区的生态恢复和治理提供理论和技术支撑,以期对有效推动黄河流域生态保护和高质量发展及助力国家构建完善的生态安全体系提供科学参考。
Biodiversity and ecosystem multifunctionality
Functional diversity can predict ecosystem functions better than dominant species: the case of desert plants in the Ebinur Lake Basin
DOI:10.3390/su13052858 [本文引用: 1]
The functional divernity of understory plants during the trausomaion from Pinus massoniana to Cinnamomum camphora forest
马尾松林向香樟林改造林下植物功能多样性研究
Functional diversity drives ecosystem multifunctionality in a Pinus yunnanensis natural secondary forest
DOI:10.1038/s41598-019-43475-1 [本文引用: 2]
Functional diversity is more sensitive to biotic filters than phylogenetic diversity during community assembly
DOI:10.1002/ecs2.2164 [本文引用: 1]
Plant functional traits in studies of vegetation changes in response to grazing and mowing: towards a use of more specific traits
Plant functional traits, functional diversity, and ecosystem functioning: current knowledge and perspectives
DOI:10.17520/biods.2015295
[本文引用: 1]
Increasing attention has recently been focused on the linkages between plant functional traits and ecosystem functioning. A comprehensive understanding of these linkages can facilitate to address the ecological consequences of plant species loss induced by human activities and climate change, and provide theoretical support for ecological restoration and ecosystem management. In recent twenty years, the evidence of strong correlations between plant functional traits and changes in ecosystem processes is growing. More importantly, ecosystem functioning can be predicted more precisely, using plant functional trait diversity (i.e., functional diversity) than species diversity. In this paper, we first defined plant functional traits and their important roles in determining ecosystem processes. Then, we review recent advances in the relationships between ecosystem functions and plant functional traits and their diversity. Finally, we propose several important future research directions, including (1) exploration of the relationships between aboveground and belowground plant traits and their roles in determining ecosystem functioning, (2) incorporation of the impacts of consumer and global environmental change into the correlation between plant functional traits and ecosystem functioning, (3) effects of functional diversity on ecosystem multifunctionality, and (4) examination of the functional diversity-ecosystem functioning relationship at different temporal and spatial scales.
植物功能性状、功能多样性与生态系统功能: 进展与展望
DOI:10.17520/biods.2015295
[本文引用: 1]
植物功能性状与生态系统功能是生态学研究的一个重要领域和热点问题。开展植物功能性状与生态系统功能的研究不仅有助于人类更好地应对全球变化情景下生物多样性丧失的生态学后果,而且能为生态恢复实践提供理论基础。近二十年来,该领域的研究迅速发展,并取得了一系列的重要研究成果,增强了人们对植物功能性状-生态系统功能关系的认识和理解。本文首先明确了植物功能性状的概念, 评述了近年来植物功能性状-生态系统功能关系领域的重要研究结果, 尤其是植物功能性状多样性-生态系统功能关系研究现状; 提出了未来植物功能性状与生态系统功能关系研究中应加强植物地上和地下性状之间关系及其与生态系统功能、植物功能性状与生态系统多功能性、不同时空尺度上植物功能性状与生态系统功能, 以及全球变化和消费者的影响等方面。
What do the biodiversity experiments tell us about consequences of plant species loss in the real world
DOI:10.1016/j.baae.2004.06.003 URL [本文引用: 1]
Responses of leaf functional traits of Zanthoxylum planispinum var. dintanensis to premature senescence
顶坛花椒叶片功能性状对早衰的响应
Relationship between ecosystem multifuntionality and species diversity in grassland ecosystems under land-use types of clipping, enclosure and grazing
DOI:10.17521/cjpe.2015.0457 URL [本文引用: 1]
刈割、围封、放牧三种利用方式下草原生态系统的多功能性与植物物种多样性之间的关系
DOI:10.17521/cjpe.2015.0457
[本文引用: 1]
随着全球变化对生物多样性的影响不断加剧, 生物多样性与生态系统功能之间相互关系(BEF)的研究显得极为重要。过去的20多年, BEF的研究大多集中在对物种多样性与单一或少数生态系统功能之间关系的探讨, 但生态系统最为重要的价值是同时维持多种服务和功能的能力, 基于此, 该文首次在国内引入近年来不断完善的生态系统多功能性(multifunctionality)的概念, 并对目前主流的评价方法进行了改进, 从而对内蒙古三种利用方式(刈割、围封、放牧)下的草地群落进行了多功能性评价, 并探讨了多功能性与物种多样性之间的关系。结果显示本研究改进的方法和目前主流方法评价得出的多功能性指数在样方和样地尺度上都有很高的相关性(R<sup>2</sup> = 0.6956, p < 0.0001; R<sup>2</sup> = 0.9231, p < 0.0001), 表明该文作者改进后的方法是可靠的。重度放牧的草地群落物种多样性水平最低, 绝大多数土壤功能指标较差, 表现出退化特征; 7年的围封和刈割群落均有较高的物种多样性水平和改善的土壤功能指标; 三者的多功能性指数为刈割(0.2178) >围封(0.0704) >放牧(-0.8031)。植被样方主要沿水肥梯度分布; 多样性指数中, 均匀度指数(Pielou index)和丰富度指数(Margelf index)对多功能性的影响作用最大, 均为样方尺度(R<sup>2 </sup>= 0.1871, p < 0.0001; R<sup>2 </sup>= 0.1601, p < 0.0001)小于样地尺度(R<sup>2 </sup>= 0.5921, p = 0.0093; R<sup>2 </sup>= 0.7499, p = 0.0007), 有尺度依赖性; 多功能性在样方和样地尺度上均与物种均匀度呈线性正相关关系, 而与物种丰富度呈单峰曲线关系。该文研究结果表明, 相对于重度放牧和围封, 刈割更有利于维持该地区生态系统的多功能性; 物种丰富度适中且物种分布均匀的生态系统可能有更好的多功能性。
Research advances in plant functional diversity and its relationship with ecosystem functions
植物功能多样性及其与生态系统功能关系研究进展
DOI:10.16258/j.cnki.1674-5906.2016.06.022 [本文引用: 2]
Relationship between functional diversity and ecosystem multifunctionality of alpine meadow along an altitude gradient in Gannan, China
甘南高寒草甸海拔梯度上功能多样性与生态系统多功能的关系
DOI:10.13287/j.1001-9332.202205.003
[本文引用: 5]
植物群落功能多样性与生态系统多功能的关系是近年来生态学研究的新视角,以往的研究多关注植物群落功能多样性与单一生态系统功能的关系,缺乏对生态系统多功能的理解。本研究以甘南高寒草甸植物群落为对象,选取地上生物量、土壤有机碳、土壤全氮、土壤全磷、土壤速效氮和土壤速效磷6个功能指标,运用Bartlett球形度检验与多阈值分析法,分析甘南高寒草甸海拔梯度上植物群落功能多样性与生态系统多功能的关系。结果表明:不同海拔植物群落组成差异显著,且海拔3500 m物种丰富度、植物盖度显著高于其他海拔。一元、多元功能多样性随海拔升高总体呈降低趋势,且海拔间差异显著。冗余分析发现,一元、多元功能丰富度、功能均匀度、Rao的二次熵均与土壤温度、土壤含水量、土壤容重呈显著正相关,与土壤pH、土壤电导率呈显著负相关。在较大阈值范围(6%~89%)内,功能多样性对生态系统多功能具有显著正效应;基于相关分析、最优回归模型和随机森林模型分析发现,功能多样性中多元功能丰富度指数与生态系统多功能具有显著正线性关系,同时多元功能丰富度也是生态系统多功能的主要驱动因素。总体来看,功能丰富度对青藏高原高寒草甸生态系统多功能的影响最为显著。
Responses of physiological parameters in plants on sub-alpine meadow to slope aspects
亚高寒草甸植物叶片生理指标对坡向的响应
DOI:10.13287/j.1001-9332.201709.030
[本文引用: 1]
研究了甘南亚高寒草甸不同坡向条件下矮嵩草、狼毒和棘豆叶片的叶绿素、游离脯氨酸和可溶性糖含量,以及稳定碳同位素(δ<sup>13</sup>C)的变化,分析干旱胁迫条件下,植物适应干旱胁迫的生理机制.结果表明: 随着坡向由北坡-西北坡-西坡-西南坡到南坡的变化,土壤含水量(北坡0.36 g·g<sup>-1</sup>,南坡0.15 g·g<sup>-1</sup>)呈降低趋势,土壤温度(北坡14.76 ℃,南坡24.85 ℃)和光照度(北坡540.34 lx,南坡744.12 lx)呈增加趋势;植物物种的组成也随之发生了变化,北坡主要有灌木金露梅及杂类草,而南坡主要有禾草类物种.3种植物叶片的脯氨酸、可溶性糖、叶绿素含量及稳定碳同位素(δ<sup>13</sup>C)随着坡向的变化均有不同程度的变化,且物种不同,各物种的生理指标变化幅度也有差异.在坡向梯度上,3种植物的脯氨酸、可溶性糖含量和稳定碳同位素与土壤含水量均呈显著负相关,与温度和光照强度呈显著正相关;植物叶片叶绿素与土壤含水量呈显著正相关,与温度和光照强度呈显著负相关.其中,土壤含水量是坡向梯度上影响植物生长的关键因子.植物叶片生理指标(脯氨酸、可溶性糖及叶绿素等)可以作为衡量植物抗逆性的因素,3种植物的抗性大小顺序为:矮嵩草>狼毒>棘豆.
Altitudinal patterns of taxonomic, phylogenetic and functional diversity of forest communities in Mount Guandi, Shanxi, China
DOI:10.17521/cjpe.2018.0088
[本文引用: 1]
Aims Understanding the spatial patterns and maintenance mechanisms of biodiversity is one of the central issues in ecology and biogeography. Specifically, altitudinal patterns of biodiversity have been widely explored to represent to mimic the latitudinal patterns. However, previous studies on altitudinal patterns of plant diversity have focused mainly on the taxonomic diversity, with less attention paid to the comprehensive information of taxonomic, phylogenetic evolution and functional traits. In this study, we explored maintenance mechanisms of diversity of forest communities in Mount Guandi, by comparing the altitudinal patterns of taxonomic, phylogenetic and functional diversity, based on the systematic investigation of forest communities.Methods 52 forest plots, each with an area of 30 m × 20 m and divided into six subplots (10 m × 10 m), were investigated along the altitudinal gradient (1 409-2 150 m) in Mount Guandi, Shanxi. Name, diameter at breast height (DBH) and height were identified and measured for each tree stem with DBH ≥3 cm at each plot; name, average height, coverage and basal diameter for each shrub species were identified and measured in two of the six subplots; name, abundance, coverage and average height for each herbaceous species were identified and measured for one quadrat of 1 m × 1 m in each subplot. The indices of taxonomic, phylogenetic and functional α and β diversity were then calculated.Important findings Species richness (S), phylogenetic diversity (PD) and functional richness (FRic) increased along the altitudinal gradient, especially in altitudes above 1 800 m. Total β diversity (βtotal) and replacement β diversity (βrepl) increased, while the richness difference (βrich) decreased, along the altitudinal gradient. The patterns of taxonomic, phylogenetic and functional diversity were significantly different between woody and herbaceous plants. S and Shannon-Wiener diversity (H′) of herbaceous plants increased more obvious along the altitude than those of woody plants. Topographic factors regulated the pattern of the herbaceous plant richness, while historical process regulated the woody plant richness. Along the altitude, βtotal increased more obvious for the woody plants than for the herbaceous plants. βrepl and βrich of woody plant showed a unimodal pattern and U-shaped pattern, while those of herbaceous plants increased or decreased, respectively. Taxonomic, phylogenetic and functional β diversity of both woody and herbaceous plants among communities increased with the environmental and geographical distances. Environmental distance had a relatively stronger effects than geographical distance on β diversity of woody plants, while environmental distance and geographical distance jointly influenced β diversity of the herbaceous plants.
山西关帝山森林群落物种、谱系和功能多样性海拔格局
DOI:10.17521/cjpe.2018.0088
[本文引用: 1]
探索和揭示生物多样性的空间格局和维持机制是生态学和生物地理学研究的热点内容, 但综合物种、系统进化和功能属性等方面的多样性海拔格局研究很少。该文以关帝山森林群落为研究对象, 综合物种、谱系和功能α和β多样性指数, 旨在初步探讨关帝山森林群落多样性海拔格局及其维持机制。研究结果表明: 随着海拔的升高(1 409-2 150 m), 关帝山森林群落物种丰富度指数(S)、谱系多样性指数(PD)和功能丰富度指数(FRic)整体上表现出上升的趋势, 特别是海拔1 800 m以上区域。随着海拔的升高, 总β多样性(β<sub>total</sub>)和更替(β<sub>repl</sub>)上升趋势明显, 而丰富度差异(β<sub>rich</sub>)则逐渐下降。不同生活型植物的物种、谱系和功能多样性海拔格局差异较大。随着海拔的升高, 草本植物S和物种多样性指数(H′)上升趋势高于木本植物。影响草本植物S分布的主要因素是地形因子, 而影响木本植物S分布的主要因素是历史过程。随着海拔的升高, 木本植物β<sub>total</sub>上升趋势要比草本植物明显。随着海拔的升高, 木本植物β<sub>repl</sub>和β<sub>rich</sub>分别表现出单峰格局和“U”形格局, 而草本植物β<sub>repl</sub>和β<sub>rich</sub>则分别表现出单调递增和单调递减的格局。随着环境差异和地理距离的增加, 群落间物种、谱系和功能β多样性显著增加。环境差异(环境过滤)对木本植物的β多样性具有相对较强的作用; 而环境差异(环境过滤)和地理距离(扩散限制)共同作用于草本植物的β多样性。
Using plant functional traits to explain diversity-productivity relationships
DOI:10.1371/journal.pone.0036760 [本文引用: 1]
Research progress on the effects of disturbance on ecosystem multifunctionality
干扰对生态系统多功能性的影响研究进展
Tree species diversity predicts aboveground carbon storage through functional diversity and functional dominance in the dry evergreen Afromontane forest of Hararghe highland, Southeast Ethiopia
DOI:10.1186/s13717-021-00322-4 [本文引用: 3]
The relationship between species diversity and ecosystem multifunctionality in alpine grasslands on the Tibetan Changtang Plateau
羌塘高寒草地物种多样性与生态系统多功能关系格局
Predictability of functional diversity depends on the number of traits
DOI:10.5814/j.issn.1674-764x.2021.03.003
[本文引用: 1]
Analysis of functional diversity, based on plant traits and community structure, provides a promising approach for exploration of the adaptive strategies of plants and the relationship between plant traits and ecosystem functioning. However, it is unclear how the number of plant traits included influences functional diversity, and whether or not there are quantitatively dependent traits. This information is fundamental to the correct use of functional diversity metrics. Here, we measured 34 traits of 366 plant species in nine forests from the tropical to boreal zones in China. These traits were used to calculate seven functional diversity metrics: functional richness (functional attribute diversity (FAD), modified FAD (MFAD), convex hull hypervolume (FRic)), functional evenness (FEve), and functional divergence (functional divergence (FDiv), functional dispersion (FDis), quadratic entropy (RaoQ)). Functional richness metrics increased with an increase in trait number, whereas the relationships between the trait divergence indexes (FDiv and FDis) and trait number were inconsistent. Four of the seven functional diversity indexes (FAD, MFAD, FRic, and RaoQ) were comparable with those in previous studies, showing predictable trends with a change in trait number. We verified our hypothesis that the number of traits strongly influences functional diversity. The relationships between these predictable functional diversity metrics and the number of traits facilitated the development of a standard protocol to enhance comparability across different studies. These findings can support integration of functional diversity index data from different studies at the site to the regional scale, and they focus attention on the influence of quantitative selection of traits on functional diversity analysis.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}