



植物的表型变异是其本身遗传多样性和环境共同作用的结果, 也是植物适应环境的表现形式(Turna & Güney, 2009; Sevik et al., 2010; 徐豪等, 2019)。研究植物的表型多样性不仅能了解植物种群遗传变异的大小, 而且有助于了解植物适应性进化的方式、机制及其影响因素(贾子瑞等, 2011; Xing et al., 2014; 蔡金峰等, 2021)。多年生林木种内有着丰富的表型及遗传变异, 决定了其对环境的适应能力, 是维持森林生态系统长期稳定的基础(Jasińska et al., 2013)。松属(Pinus)隶属于松科, 是雌雄同株的常绿针叶乔木, 作为全球森林生态系统中的关键类群, 在世界上分布范围极广, 从高纬度的北极地区一直延伸到热带地区, 纬度跨度达70°以上(Jin et al., 2021)。因此, 松属物种具备高度的适应性和可塑性以应对异质环境, 对其表型变异的研究将有助于理解物种的适应性进化。针叶作为松属树种最具活力的营养器官, 通常成束生长, 且每束针叶个数较为稳定(Jasińska et al., 2013), 其形态特征在一定程度上可作为松属物种分类的重要参考依据(郑万钧和傅立国, 1978; 刘永良, 2011)。同时, 针叶树种的针叶形态变异也是适应进化的结果, 其形态和解剖特征与其生理功能(光合作用、呼吸、水分代谢、抗逆性等)密切相关, 被广泛用于地理变异、系统发育和适应性进化等相关研究(Eguchi et al., 2004; Androsiuk et al., 2011; Mao et al., 2012; Jasińska et al., 2013)。因此, 研究跨越较大地理尺度的松树种群的针叶形态解剖特征变异将有助于揭示不同地理种群的针叶表型多样性, 并从生理生态机制上探讨针叶树种的地理变异、种群动态以及对地理气候的响应机理(Xing et al., 2014; Huang et al., 2016; Jankowski et al., 2019)。
目前我国松树造林树种主要有马尾松(P. massoniana)、油松(P. tabuliformis)、华山松(P. armandii)、落叶松(Larix gmelinii)、云南松(P. yunnanensis), 以及从国外引种的湿地松(P. elliottii)和火炬松(P. taeda)等。我国当前的松树种质资源还比较单一, 遗传资源匮乏, 亟需收集全球具有重要遗传价值的松树种质资源。松属物种在热带少量分布, 但因其能够适应热带山地的独特气候环境, 具有重要的遗传价值和育种潜力。卡西亚松(P. kesiya)广泛分布于热带东南亚地区, 覆盖了泰国、老挝、越南、缅甸、菲律宾、马来西亚、印度尼西亚等多个国家, 是整个东南亚地区重要的松树用材树种(Missanjo & Matsumura, 2016; 杨雪艳等, 2019)。云南松主要分布于中国云南、四川、广西等亚热带地区(戴开结等, 2006; 陈飞等, 2012)。思茅松(P. kesiya var. langbianensis)位于滇南思茅、宁洱、景洪等热带区域, 在以往的研究中被认为是卡西亚松的一个地理变种(吴征镒等, 1987; 吴兆录, 1994)。三者的系统关系一直以来没有得到彻底解决, 早期学者认为应该将云南松、思茅松以及在滇东南和西藏分布的高山松(P. densata)都应做为卡西亚松的变种(Turnbull et al., 1980), 但后期学者研究发现高山松是云南松与油松的杂交种(Wang et al., 2011; Gao et al., 2012)。虽然云南松跟卡西亚松的关系更近, 但基于形态学和化学成分的研究认为云南松和卡西亚松有显著的区别, 应该是独立的物种(Businský et al., 2014; Zhang et al., 2014)。本研究中3种松树跨越了较大的地理区域, 分布区呈现替代性地理分布, 具有丰富的气候变异, 同时种内具有丰富的遗传变异, 种间存在一定的基因交流。显著而多样的局部适应性使卡西亚松、云南松、思茅松构成了一个具有显著生境分化和杂交渐渗的松属内近缘种复合体。
目前国内关于云南松、思茅松的表型及遗传多样性的分析还只是局限于种内的少数种群, 而针对卡西亚松天然种群变异水平的研究也仅见于少数越南种群的报道(Wang et al., 1996; Santisuk et al., 1997)。Huang等(2016)对分布于云南省的7个云南松种群的22个针叶形态和解剖特征进行了分析, 结果表明所有性状在种群间和种群内的变异均存在显著差异。许玉兰等(2018)对云南松9个天然种群的球果性状进行分析, 发现球果性状在种群间和种群内差异极显著, 存在广泛的表型变异。李帅锋等(2013)对云南省11个思茅松天然种群的种实性状进行了研究, 以此探讨思茅松种群间和种群内的表型差异, 结果表明思茅松种群间和种群内种实表型性状差异显著。而整合3种松树的表型研究仅Zhang等(2022)对卡西亚松、云南松以及思茅松的种子形态结合环境因子和纬度梯度进行了研究, 结果表明环境因素影响了松树种子的表型变异, 而细胞质基因组的遗传变异与不同种松树的种子萌发适合度显著相关。目前对于该复合体种间重要表型性状变异、遗传变异, 以及对其立地环境的适应性所知甚少。基于以上背景, 本研究关注的科学问题是: 针叶表型变异是否能解释云南松、思茅松和卡西亚松的地理替代分布格局? 本研究针对云南松、思茅松和卡西亚松3种松树进行了覆盖全分布区、跨越东南亚数个国家的大区域尺度取样, 解析了31个代表性天然种群的针叶形态和解剖学特征的变异, 旨在揭示: (1) 3种松树种间和种群间针叶性状特征的变异程度、地理分布规律; (2) 3种松树针叶性状特征与地理、环境因子的相关性。研究结果将有助于深入认识松属不同物种大区域尺度的地理种群表型变异规律, 为揭示松属植物适应性进化提供形态学和生态学的研究基础, 同时也为松属植物遗传育种、种质资源的保育与利用提供参考依据。
1 材料和方法
1.1 样本采集及气候数据获取
根据中国植物志(
表1 云南松、卡西亚松及思茅松采集信息表
Table 1
| 物种 Species | 种群地点 Population location | 种群编号 Population No. | 经度 Longitude (° E) | 纬度 Latitude (° N) | 海拔 Altitude (m) |
|---|---|---|---|---|---|
| 卡西亚松 P. kesiya | 老挝阿速坡 Attapu, Laos | ASPPk | 107.19 | 15.22 | 1 061 |
| 缅甸掸邦, 东枝 Dongzhi, Shan State, Myanmar | DZPk | 97.06 | 20.79 | 1 583 | |
| 老挝川圹, 丰沙湾 Phonsavan, Xieng Khouang, Laos | FSWPk | 103.12 | 19.50 | 1 070 | |
| 缅甸掸邦, 格劳 GeLao, Shan State, Myanmar | KLPk | 96.55 | 20.65 | 1 022 | |
| 菲律宾 Philippines | PHP1 | 120.85 | 16.19 | 1 201 | |
| 菲律宾 Philippines | PHP4 | 120.89 | 17.14 | 751 | |
| 菲律宾 Philippines | PHP8 | 120.89 | 18.10 | 50 | |
| 老挝华潘, 桑怒 Sam Neua, Houaphanh, Laos | SNPk | 103.96 | 20.22 | 1 399 | |
| 越南安沛, 拉潘滩 La Pan Tan, Yen Bai, Vietnam | V04 | 104.13 | 21.75 | 1 410 | |
| 越南得农 Dak Nong Vietnam | V08 | 107.43 | 12.15 | 847 | |
| 思茅松 P. kesiya var. langbianensis | 中国云南 Yunnan, China | ||||
| 景谷 Jinggu | JGPk | 100.50 | 23.49 | 1 707 | |
| 景洪贺建 Hejian, Jinghong | JHPk02 | 100.51 | 22.25 | 1 244 | |
| 勐腊 Mengla | MLPk | 101.67 | 21.25 | 921 | |
| 宁洱 Ning’er | NEPk | 100.97 | 23.19 | 1 004 | |
| 思茅 Simao | SMPk | 100.95 | 22.71 | 1 340 | |
| 景洪大渡岗 Dadugang, Jinghong | JHPk01 | 100.99 | 22.42 | 1 113 | |
| 勐海布朗山 Bulangshan, Menghai | MHPk01 | 100.24 | 21.66 | 885 | |
| 墨江 Mojiang | MJPk | 101.61 | 23.40 | 1 690 | |
| 镇沅 Zhengyuan | ZYPk | 101.14 | 23.88 | 1 109 | |
| 云南松 P. yunnanensis | 中国云南 Yunnan, China | ||||
| 宾川 Binchuan | BCPy | 100.35 | 25.97 | 1 809 | |
| 大理云龙 Yunlong, Dali | DLPy | 99.29 | 25.87 | 2 572 | |
| 景东 Jingdong | JDPy | 100.67 | 24.70 | 1 713 | |
| 昆明 Kunming | KMPy | 102.62 | 24.91 | 2 041 | |
| 丽江 Lijiang | LJPy | 100.22 | 26.88 | 2 651 | |
| 腾冲 Tengchong | TCPy | 98.58 | 24.91 | 1 776 | |
| 文山 Wenshan | WSPy02 | 104.63 | 23.97 | 1 573 | |
| 宜良 Yiliang | YLPy | 103.16 | 24.71 | 1 938 | |
| 玉溪 Yuxi | YXPy | 102.15 | 24.24 | 1 861 | |
| 中国广西 Guangxi, China | |||||
| 乐业 Leye | LYPy | 106.37 | 24.80 | 1 336 | |
| 西林 Xilin | XLPy | 104.62 | 24.59 | 857 | |
| 隆林 Longlin | LLPy | 104.96 | 24.69 | 1 294 |
图1
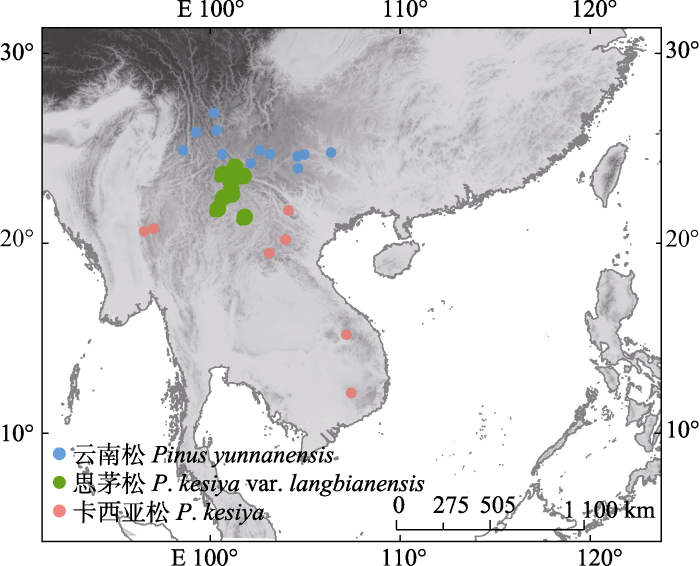
图1
云南松、卡西亚松和思茅松31个种群的地理分布。
Fig. 1
Geographic locations of 31 populations from Pinus yunnanensis, P. kesiya and P. kesiya var. langbianensis.
从WorldClim数据库(
1.2 针叶性状指标测定
利用电子游标卡尺测量针叶长度(每个种群随机选取10个个体, 每个个体随机选取10根针叶)。从针叶中部取1 cm左右的样品放入KCl、HNO3和水体积比为1:1:1配置的溶液中, 浸泡至白色; 再转移到5% NaOH溶液中继续浸泡至透明。将透明片段在体视镜下进行解剖, 沿着针叶轴面剖开, 刮去叶肉细胞和维管束, 获得完整的透明上表皮。随后滴入番红染液进行染色, 在上表皮由透明完全转为粉色后取出并用清水涮洗, 放置载玻片上展平, 滴加一滴甘油制成装片。用显微镜DM 2000 (Leica Microsystems, Wetzlar, Germany)拍摄每个装片在10倍和20倍镜下的气孔照片各3张。利用ImageJ软件测量气孔保卫细胞长度、宽度, 木质增厚层长度、宽度, 气孔腔长度、宽度共6个解剖特征(附录I)。在10倍放大镜下计数600 µm2内气孔个数用来计算气孔密度。
1.3 数据分析
首先, 采用Tukey’s HSD多重比较检测任一测量性状在3种松树间是否存在显著性差异。并根据巢式方差分析(Yijk ~ αi/βij)计算各性状在种间和种群间的差异, 其中Yijk是第i种松树第j个种群个体的第k个观测值, αi是种间效应, βij是嵌套在αi下所对应的种内群体间效应。种群间表型分化系数(VST)通过公式VST =
2 结果
2.1 3种松树群体间针叶性状的变异和变异来源
表型变异系数(CV)反映了针叶性状在3种松树种间、种群间的离散(变异)程度。结果(表2)表明: 8个针叶性状的种群间的变异系数在12.01%-34.08%之间, 其中气孔保卫细胞长度、气孔保卫细胞宽度和木质增厚层宽度的变异系数小于17%, 在种群间的变异程度较小。其余5个性状(木质增厚层长度、气孔腔长度、气孔腔宽度、针叶长度和气孔密度)的变异系数均在22%以上, 其中气孔腔长度的变异系数最大, 气孔密度和气孔腔宽度的变异系数次之, 分别为34.08%、31.66%和28.26%。比较3种松树种间的针叶性状的变异系数可知, 思茅松针叶性状的变异系数最大(10.13%-36.53%), 卡西亚松次之(10.80%- 36.70%), 云南松最小(11.26%-31.01%)。各性状在不同种内的变异程度也具有一定差异, 卡西亚松的气孔腔大小相关性状(气孔腔长度、气孔腔宽度)变异最大(36.70%、26.12%), 思茅松的针叶长度变异最大(36.53%), 云南松的气孔腔长度(31.01%)、气孔腔宽度(24.56%)以及气孔密度(25.68%)变异相对较大。通过对各性状种群水平的箱形图统计, 直观地展示出3种
松树种群水平的针叶性状的变异幅度和差异(附录II)。
表2 云南松、思茅松和卡西亚松天然种群针叶的表型性状、变异系数和种间Tukey’s HSD多重比较
Table 2
| 性状 Trait | 云南松 Pinus yunnanensis | 卡西亚松 P. kesiya | 思茅松 P. kesiya var. langbianensis | 种群间 Among populations | ||||
|---|---|---|---|---|---|---|---|---|
| 平均值±标准差 Mean ± SD | 变异系数 CV (%) | 平均值±标准差 Mean ± SD | 变异系数 CV (%) | 平均值±标准差 Mean ± SD | 变异系数 CV (%) | 平均值±标准差 Mean ± SD | 变异系数 CV (%) | |
| LSGC (μm) | 30.20 ± 3.41b | 11.30 | 35.45 ± 3.83a | 10.80 | 31.41 ± 3.86b | 12.28 | 32.28 ± 4.32 | 13.39 |
| WSGC (μm) | 24.70 ± 2.78b | 11.26 | 27.32 ± 3.17a | 11.62 | 24.94 ± 2.53ab | 10.13 | 25.63 ± 3.08 | 12.01 |
| WTLL (μm) | 16.94 ± 2.62b | 15.50 | 22.66 ± 4.64a | 20.49 | 18.31 ± 2.69b | 14.68 | 19.21 ± 4.23 | 22.01 |
| WTLW (μm) | 13.84 ± 2.20c | 15.77 | 17.65 ± 2.00a | 11.35 | 15.58 ± 2.18b | 14.01 | 15.63 ± 2.64 | 16.87 |
| LSC (μm) | 6.99 ± 2.17a | 31.01 | 8.07 ± 2.96a | 36.70 | 6.57 ± 1.90a | 28.84 | 7.23 ± 2.46 | 34.08 |
| WSC (μm) | 5.88 ± 1.45b | 24.56 | 6.95 ± 1.82a | 26.12 | 5.67 ± 1.74b | 30.70 | 6.17 ± 1.74 | 28.26 |
| NL (cm) | 21.34 ± 3.57a | 16.72 | 18.43 ± 2.87ab | 15.59 | 14.93 ± 5.45b | 36.53 | 18.53 ± 4.78 | 25.78 |
| SD (600 µm-2) | 98.43 ± 25.28a | 25.68 | 68.92 ± 16.11b | 23.38 | 66.87 ± 17.41b | 26.04 | 79.62 ± 25.21 | 31.66 |
不同小写字母表示差异显著(p < 0.05)。LSC, 气孔腔长度; LSGC, 气孔保卫细胞长度; NL, 针叶长度; SD, 气孔密度; WSC, 气孔腔宽度; WSGC, 气孔保卫细胞宽度; WTLL, 木质增厚层长度; WTLW, 木质增厚层宽度。
Different lowercase letters indicate significant difference (p < 0.05). LSC, length of the stomata cavity; LSGC, length of stomata guard cell; NL, needle length; SD, stomatal density; WSC, width of the stomata cavity; WSGC, width of the stomata guard cell; WTLL, length of the woody thickening layer; WTLW, width of the woody thickening layer.
多重比较的结果(表2)显示3种松树在气孔腔长度上无显著差异, 而木质增厚层宽度在3种松树间存在显著差异。其他性状如: 气孔保卫细胞长度、气孔保卫细胞宽度、木质增厚层长度、气孔腔宽度表现为卡西亚松和云南松差异显著, 思茅松的针叶性状更倾向于云南松。仅气孔密度例外, 表现为思茅松和卡西亚松相近, 而云南松则与这两种松树差异显著。卡西亚松针叶与气孔大小相关的性状(气孔保卫细胞长度、木质增厚层长度和气孔腔宽度)显著大于其他两种松树的性状。而云南松针叶的单位面积气孔密度是3种松树中最高的。
巢式方差分析将针叶表型性状的变异分解为种间和种内变异, 从而可以揭示性状的变异来源。结果(表3)显示: 8个针叶表型性状中, 除气孔保卫细胞宽度、气孔腔长度、气孔腔宽度外, 其他性状(气孔保卫细胞长度、木质增厚层长度、木质增厚层宽度、针叶长度、气孔密度)的变异主要存在于种间和种群间(种间和种群间方差分量之和大于50%)。同时, 种间的变异大于种内变异。本研究中, 巢式方差模型中的残差包含了种群内的方差分量和实验误差, 但是目前缺少有效的分析方法可以将种群内变异和实验误差剥离开来。
表3 云南松、思茅松和卡西亚松天然种群针叶表型性状变异方差分析
Table 3
| 性状 Trait | 均方 Mean square | F | 方差分量 Variance components (%) | VST (%) | |||||
|---|---|---|---|---|---|---|---|---|---|
| 种间 Interspecies | 种内群体间 Populations within species | 残差 Residuals | 种间 Interspecies | 种内群体间 Populations within species | 种间 Interspecies | 种内群体间 Populations within species | 残差 Residuals | ||
| LSGC (μm) | 770.50 | 43.80 | 10.40 | 17.59*** | 4.21*** | 34.66 | 16.40 | 48.94 | 67.88 |
| WSGC (μm) | 209.53 | 31.04 | 5.71 | 6.75*** | 5.44*** | 17.78 | 25.97 | 56.25 | 40.64 |
| WTLL (μm) | 914.30 | 50.20 | 7.80 | 18.21*** | 6.44*** | 41.77 | 21.06 | 37.17 | 66.48 |
| WTLW (μm) | 363.60 | 9.30 | 4.00 | 39.10*** | 2.33*** | 43.89 | 6.67 | 49.44 | 86.80 |
| LSC (μm) | 56.49 | 21.60 | 4.06 | 2.62*** | 5.32*** | 5.63 | 29.28 | 65.08 | 16.14 |
| WSC (μm) | 45.31 | 6.77 | 2.33 | 6.69*** | 2.91*** | 12.24 | 14.50 | 73.26 | 45.79 |
| NL (cm) | 1 005.90 | 61.00 | 11.50 | 16.49*** | 5.30*** | 36.54 | 19.68 | 43.78 | 64.99 |
| SD (600 µm-2) | 32 767.00 | 1 112.00 | 345.00 | 29.47*** | 3.22*** | 43.03 | 10.72 | 46.25 | 80.06 |
| 平均值 Mean | 4 516.58 | 166.96 | 48.85 | - | - | 29.44 | 18.04 | 52.52 | 58.60 |
*, p < 0.05; **, p < 0.01; ***, p < 0.001。LSC, 气孔腔长度; LSGC, 气孔保卫细胞长度; NL, 针叶长度; SD, 气孔密度; WSC, 气孔腔宽度; WSGC, 气孔保卫细胞宽度; WTLL, 木质增厚层长度; WTLW, 木质增厚层宽度; VST, 表型分化系数。-, 无数据。
*, p < 0.05; **, p < 0.01; ***, p < 0.001. LSC, length of the stomata cavity; LSGC, length of stomata guard cell; NL, needle length; SD, stomatal density; WSC, width of the stomata cavity; WSGC, width of the stomata guard cell, WTLL; length of the woody thickening layer; WTLW, width of the woody thickening layer; VST, phenotypic differentiation coefficient. -, no data.
2.2 针叶性状变异的群体结构
针叶表型解剖性状PCA结果显示, PC1和PC2的贡献率分别为50.3%和21.4%, 3种松树呈现出明显的种内聚类。其中与针叶气孔腔大小(气孔腔长度、气孔腔宽度), 保卫细胞大小(气孔保卫细胞长度、气孔保卫细胞宽度), 以及木质增厚层相关的性状(木质增厚层宽度、木质增厚层长度)在PC1上有较高的荷载。而针叶长度与单位面积针叶气孔密度在PC2上有较高的荷载(图2A)。
图2
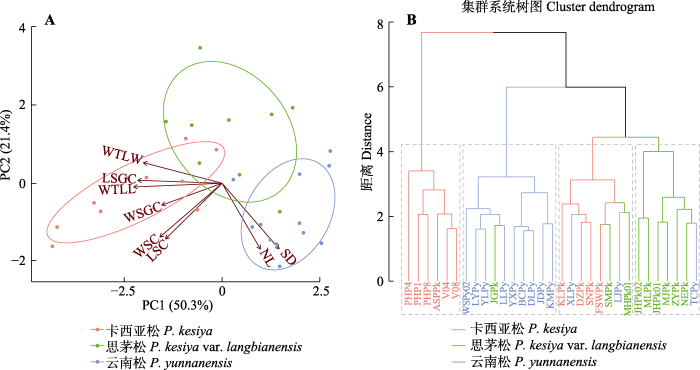
图2
云南松、思茅松和卡西亚松针叶性状指标的主成分(PC)分析(A)和31个天然群体的针叶性状聚类图(B)。LSC, 气孔腔长度; LSGC, 气孔保卫细胞长度; NL, 针叶长度; SD, 气孔密度; WSC, 气孔腔宽度; WSGC, 气孔保卫细胞宽度; WTLL, 木质增厚层长度; WTLW, 木质增厚层宽度。群体编号信息见
Fig. 2
Scatter diagram of the first two principal coordinates from principal component (PC) analysis (A) and ward cluster (B) for the 31 populations based on needle traits of Pinus yunnanensis, P. kesiya var. langbianensis and P. kesiya. LSC, length of the stomata cavity; LSGC, length of stomata guard cell; NL, needle length; SD, stomatal density; WSC, width of the stomata cavity; WSGC, width of the stomata guard cell; WTLL, length of the woody thickening layer; WTLW, width of the woody thickening layer. See
聚类分析结果表明31个天然种群分为2个聚类, 第1个聚类为来自菲律宾、老挝、越南南部的6个卡西亚松种群(PHP4、PHP1、PHP8、ASPPk、V04和V08); 第2个聚类由25个天然种群构成, 可进一步分为3个亚类群: 第1个亚类群由主要分布于云南中部和西北部的9个云南松种群(WSPy02、LYPy、YLPy、LLPy、YXPy、BCPy、DLPy、JDPy和KMPy)和1个思茅松种群(JGPk)组成, 第2个亚类群由4个来自缅甸、老挝、越南北部的卡西亚松种群(KLPk、DZPk、SNPk和FSWPk), 2个云南松种群(XLPy和LJPy)和2个思茅松种群(SMPk和MHPk01)组成, 第3个亚类群由6个思茅松种群(JHPk02、MLPk、JHPk01、MJPk、ZYPk和NEPk)和来自云南西北部的1个云南松种群(TCPy)组成(图2B)。
2.3 3种松树针叶表型性状与地理、环境因子的相关性
Spearman相关性分析结果表明3种松树针叶性状与纬度梯度相关性显著。其中针叶长度、气孔密度与纬度正相关, 而气孔保卫细胞长度、气孔保卫细胞宽度、木质增厚层长度、木质增厚层宽度、气孔腔长度和气孔腔宽度6种性状则与纬度显著负相关(图3)。多元回归分析的结果表明针叶长度、气孔密度和最干季平均气温显著负相关, 木质增厚层的长宽与最干季平均气温显著正相关。保卫细胞的大小与最湿月降水量显著正相关; 而气孔腔长度与最湿月降水量、最干月降水量显著正相关; 而与平均气温日较差和降水量季节变异系数显著负相关(图4; 表4)。图5展示了反映针叶性状的代表性主成分PC1的群体得分值, 以及多元回归分析筛选出的显著相关的气候变量在采样区域的变异趋势。
图3
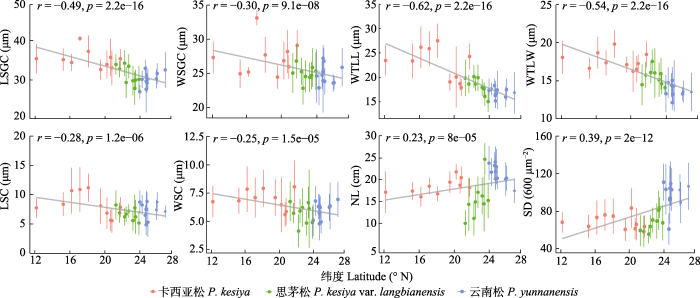
图3
云南松、思茅松和卡西亚松针叶性状与纬度梯度相关性分析。LSC, 气孔腔长度; LSGC, 气孔保卫细胞长度; NL, 针叶长度; SD, 气孔密度; WSC, 气孔腔宽度; WSGC, 气孔保卫细胞宽度; WTLL, 木质增厚层长度; WTLW, 木质增厚层宽度。
Fig. 3
Latitudinal gradients of the needle traits of Pinus yunnanensis, P. kesiya var. langbianensis and P. kesiya. LSC, length of the stomata cavity; LSGC, length of stomata guard cell; NL, needle length; SD, stomatal density; WSC, width of the stomata cavity; WSGC, width of the stomata guard cell; WTLL, length of the woody thickening layer; WTLW, width of the woody thickening layer.
图4
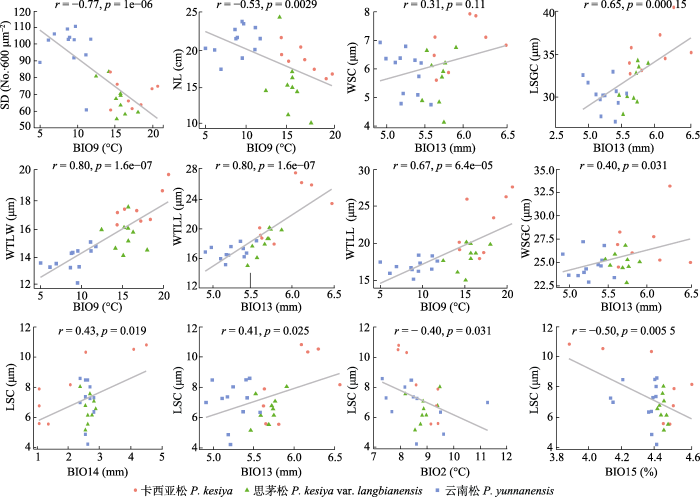
图4
多元线性模型筛选出的重要气候变量与云南松、思茅松和卡西亚松针叶性状的相关性。LSC, 气孔腔长度; LSGC, 气孔保卫细胞长度; NL, 针叶长度; SD, 气孔密度; WSC, 气孔腔宽度; WSGC, 气孔保卫细胞宽度; WTLL, 木质增厚层长度; WTLW, 木质增厚层宽度。BIO2, 平均气温日较差; BIO9, 最干季平均气温; BIO13, 最湿月降水量; BIO14, 最干月降水量; BIO15, 降水量季节变异系数。r, 相关系数。所有值为基于lg转换。
Fig. 4
Significant correlation between the needle traits and climatic variables (selected by the multiple linear regression model) of Pinus yunnanensis, P. kesiya var. langbianensis and P. kesiya. LSC, length of the stomata cavity; LSGC, length of stomata guard cell; NL, needle length; SD, stomatal density; WSC, width of the stomata cavity; WSGC, width of the stomata guard cell; WTLL, length of the woody thickening layer; WTLW, width of the woody thickening layer. BIO2, mean temperature diurnal range; BIO9, mean temperature of driest quarter; BIO13, precipitation of wettest month; BIO14, precipitation of driest month; BIO15, precipitation seasonality. r, correlation coefficient. The values was lg transformed.
图5
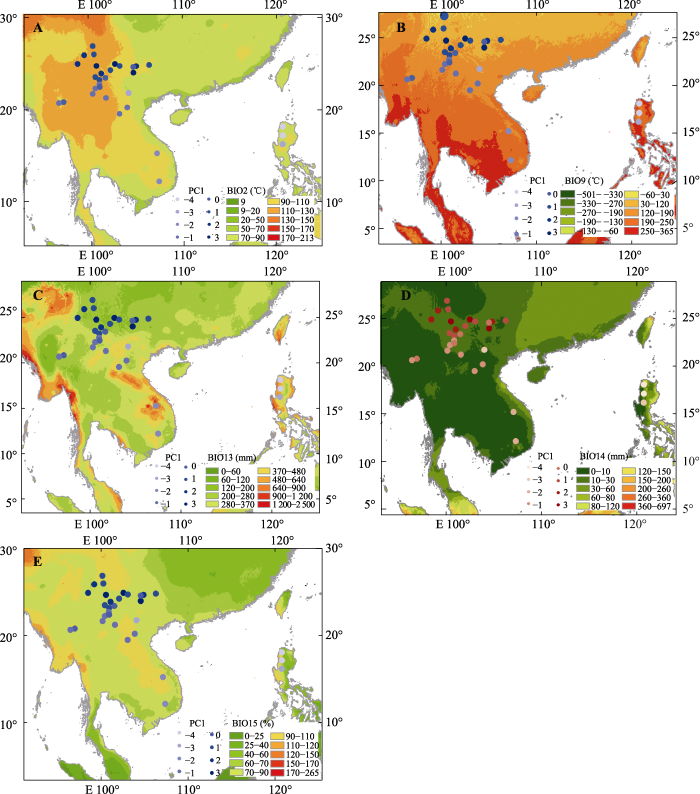
图5
云南松、思茅松和卡西亚松的针叶表型性状主成分分析中第一主成分的群体分值(PC1)随重要气候变量(多元线性模型筛选)的地理变异趋势。BIO2, 平均气温日较差; BIO9, 最干季平均气温; BIO13, 最湿月降水量; BIO14, 最干月降水量; BIO15, 降水量季节变异系数。所有值为基于lg转换后扩大100倍。
Fig. 5
Distribution patterns of the population score of the first principal component value (PC1) based on the analysis of conifer phenotypic traits of Pinus yunnanensis, P. kesiya var. langbianensis and P. kesiya was the geographical variation trend of important climatic variables (multivariate linear model screening). BIO2, mean temperature diurnal range; BIO9, mean temperature of driest quarter; BIO13, precipitation of wettest month; BIO14, precipitation of driest month; BIO15, precipitation seasonality. The values was lg transformed and multiplied by 100.
表4 多元线性回归模型筛选的影响云南松、思茅松和卡西亚松种群针叶表型性状变异的气候变量
Table 4
| 性状 Trait | 截距 Intercept | 平均气温日较差 BIO2 | 最湿季平均气温 BIO8 | 最干季平均气温 BIO9 | 最湿月降水量 BIO13 | 最干月降水量 BIO14 | 降水量季节变异系数 BIO15 |
|---|---|---|---|---|---|---|---|
| LSGC (μm) | -0.000 8 | 0.1 759 | 0.7 516 | -0.3 924 | |||
| WSGC (μm) | -0.054 6 | -0.5 432 | 0.6 846 | 0.3 786 | |||
| WTLL (μm) | 0.007 7 | 0.2 861 | 0.6 523 | -0.4 449 | |||
| WTLW (μm) | -0.008 3 | -0.3 561 | 0.9 913 | 0.1 731 | |||
| LSC (μm) | -0.031 4 | -0.4 884 | 0.7 315 | 0.7 785 | -1.3 658 | ||
| WSC (μm) | -0.027 5 | 0.4 550 | -0.3 918 | ||||
| NL (cm) | -0.007 0 | -0.3 137 | -0.5 250 | ||||
| SD ( 600 µm-2) | -0.018 1 | -0.8 011 | 0.1 918 |
LSC, 气孔腔长度; LSGC, 气孔保卫细胞长度; NL, 针叶长度; SD, 气孔密度; WSC, 气孔腔宽度; WSGC, 气孔保卫细胞宽度; WTLL, 木质增厚层长度; WTLW, 木质增厚层宽度。
LSC, length of the stomata cavity; LSGC, length of stomata guard cell; NL, needle length; SD, stomatal density; WSC, width of the stomata cavity; WSGC, width of the stomata guard cell; WTLL, length of the woody thickening layer; WTLW, width of the woody thickening layer. BIO2, mean temperature diurnal range; BIO9, mean temperature of driest quarter; BIO13, precipitation of wettest month; BIO14, precipitation of driest month; BIO15, precipitation seasonality.
3 讨论
3.1 3种松树针叶性状变异的多样性及变异来源
植物表型多样性是遗传适应性和环境异质性长期共同作用的结果, 研究不同地理环境下天然种群的表型变异丰富程度和变异规律, 能够在一定程度上反映其遗传变异水平, 同时还可揭示其对不同环境的适应能力(Willmore et al., 2007; Blackman et al., 2017)。本研究中, 针叶性状在3种松树种群间变异较大, 变异系数为12.01%-34.08%。3种松树的针叶性状变异主要存在于种间与种内群体间, 主要体现在其中5个性状(气孔保卫细胞长度、木质增厚层长度和宽度、针叶长度、气孔密度)的种间和种群间方差分量之和大于50%, 同时在这些性状上, 种间变异又大于种群间变异。尽管由于统计分析方法的限制无法将剩余变异分量中的种群内变异和实验误差剥离开, 我们认为3种松树种群内也应存在一定的遗传变异。前人针对思茅松天然种群的种实性状表型变异(李帅锋等, 2013), 以及针对云南松部分种群的针叶性状(徐杨等, 2016)、球果表型性状(许玉兰等, 2018)的研究也均发现在种群内存在丰富的变异。
松树针叶表型的丰富变异同时也反映在表型分化系数上, 3种松树的针叶表型分化系数均值为58.60%, 与黑云杉(Picea mariana)和白云杉(P. glauca)的表型分化系数(49.41%-57.04%和50%)比较接近(张含国, 2000; 罗建勋和顾万春, 2004), 但高于其他针叶树种。 例如分布较广泛的油松的表型分化系数为36% (刘永良, 2011), 川西云杉(P. likiangensis var. balfouriana)为33.03% (辜云杰等, 2009)。本研究松树的表型分化系数更是高于北美地区分布的一些针叶树种, 例如北美短叶松(Pinus banksiana)、火炬松和花旗松(Pseudotsuga menziessii)的表型分化系数分别是1.6%-18.9%、15%和11.1% (葛颂等, 1988; 辜云杰等, 2009)。这可能是因为本研究中的3种松树跨越了较大的地理分布区和气候带, 分布区域内环境异质性较高。同时与北美和中国温带分布的针叶树种相比, 本研究中的3种松树进化历史悠久, 分布地所在的中国西南和中南半岛热带区域受冰期的影响较小, 因此种群间和种群内保留了更丰富的遗传变异和表型变异。
3.2 针叶性状变异反映了云南松、思茅松和卡西亚松的地理替代分布格局
本研究表明, 木质增厚层长宽、气孔保卫细胞长宽、气孔腔宽度及气孔密度在云南松和卡西亚松间存在显著差异, 同时这些性状的表型分化系数也较高(针叶长度除外), 说明这些性状同时受到了物种系统发育关系以及各自分布环境差异的影响。云南松和卡西亚松是由亲本种P. prekesiya在新近纪因气候动荡和青藏高原隆升所引起的地形变化而分化成种(Xing et al., 2010), 两个物种分化时间较长, 在长期的进化过程中因地理隔离和适应不同的生态环境而产生了较高程度的种间分化, 从而在针叶表型上也体现出较显著的分化。云南松分布区的北部海拔较高, 在青藏高原东南侧的地理条件和错综复杂的地貌综合影响下形成了复杂多样的气候, 而分布区南部处于强季风区, 降水多、温度高, 整个区域气温和降水的差异大。卡西亚松主要的分布区位于中南半岛, 属典型的热带季风气候, 其特点为全年高温, 最冷月平均气温也在18 ℃以上, 大部分地区年降水量为1 500-2 000 mm。思茅松的表型性状则处于中间过渡状态, 大部分的针叶性状更倾向于云南松。在地域环境上, 思茅松处于3个种地理分布区的中间过渡区域, 位于云南南部的热带与亚热带地区, 年平均气温16.86-21.15 ℃, 年降水量1 082-1 402 mm之间, 有较为明显的干湿季的区分(Yang et al., 2022), 气候环境与云南松更相近。但是值得注意的是, 思茅松种群内表现出更短的针叶长度和更低的气孔密度, 与云南松、卡西亚松均不同。这可能是因为遗传组成上频繁的杂交渐渗使思茅松的基因组蕴含了两个亲本种(云南松和卡西亚松)的成分(Businský et al., 2014), 可能会表现出超亲性状。而另一方面, 思茅松的针叶表型还有可能受到极端干旱事件的影响。云南地区气候敏感性高, 近些年来干旱事件频发(付奔等, 2014), 根据Yang等(2019)研究表明思茅松分布地所处云南南部, 发生极端干旱的频率最高。频繁的极端干旱也会导致思茅松的针叶变短, 气孔密度降低。
聚类分析揭示的针叶性状的种群结构也呈现出一定的地理区域结构。聚类的第一支主要是来自卡西亚松的主分布区菲律宾岛屿种群、越南种群以及老挝南部种群, 该区域与云南松和思茅松分布区的距离较远,环境差异较大。这些卡西亚松经历长期的环境适应和自然选择可能已经形成了稳定遗传的表型, 并体现在其针叶长度适中, 气孔密度小, 气孔腔和木质增厚层较大等方面。而另一大支的种群则分为两个亚类群, 其中一个亚类群包括了来自云南中部及西北部分布的云南松种群和一个来自云南景谷的思茅松种群(JGPk)。该亚类的种群分布为3种松树分布区的最北缘, 针叶长度长, 气孔密度大。第二个亚类群主要由来自云南松和卡西亚松分布区域及分布区重叠区域的思茅松种群组成, 这一类群的特点是地理分布相邻, 环境差异较小, 同时种群间可能还存在频繁的基因交流, 体现在针叶表型上多为中间态。
3.3 3种松树天然群体针叶表型对地理及气候变量的响应
本研究中针叶性状主要呈现纬度梯度的变异趋势, 8个针叶性状均与纬度显著相关。其中针叶长度和气孔密度与纬度显著正相关, 其他性状则与纬度显著负相关。本研究发现, 热带分布的松树的针叶性状与此前报道的温带及寒带针叶树种的针叶性状变异呈现出明显不同的地理变异趋势。一般来说, 针叶树种为适应干旱或寒冷环境, 针叶长度有变短的趋势(Reich et al., 1996; Farjon & Styles, 1997; Dobbertin et al., 2010)。例如在我国北方分布的油松, 随着纬度升高, 气温降低, 针叶为适应低温环境而变短, 而针叶厚度也由西南向东北呈逐渐增加的趋势(徐化成, 1992; Zhang et al., 2017)。本研究中3种松树总体的趋势随着纬度的增加, 针叶长度增加, 气孔密度增加, 气孔腔大小减小。但是值得注意的是, 本研究发现在云南松的种内, 呈现出随着纬度增加针叶长度变短的趋势。Nobis等(2012)针对全球尺度下的针叶树种形态特征的纬向变异及其与环境和系统发育关系的研究中, 揭示了针叶长度以及针叶长宽比与纬度显著负相关, 但是在低纬度分布的松属物种中发现了明显的偏移, 并没有表现出连续的针叶长宽比随纬度的增加而下降的趋势, 本研究的研究结果与之一致。
纬度的变化伴随着一系列环境因子的变化, 植物表型性状的响应是在各种环境因子相互作用下的综合反应的结果。本研究进一步将针叶表型跟具体的环境因子进行了多元回归分析来寻找决定针叶性状变异的关键环境因子。结果发现, 气孔密度和针叶长度的最主要决定因子为最干季平均气温, 而影响气孔大小的环境因子主要是最干月降水量、最湿月降水量和降水量季节变异系数。同时最干季平均气温也影响了木质增厚层的长宽, 最湿月降水量影响气孔保卫细胞长宽。本研究确定的这些关键环境因子与其他研究的结果一致, 一项关于云南松和卡西亚松径向生长与气候的敏感性研究也表明中国西南地区云南松和卡西亚松径向生长对旱季生长期水分可利用性高度敏感(Yang et al., 2022), 而旱季平均气温在一定程度上决定了水分的可利用性。针叶长度指标反映了针叶性状的物质总量, 也体现了植物体参与光合、蒸腾作用的总器官大小。而气孔是植物与外界进行气体交换的通道, 常常被用作反映植物对干旱和温度的适应性的重要指标(Peak & Mott, 2011)。木质增厚层和保卫细胞的大小在一定程度上反映了植物的抗逆性(Li et al., 2021)。有研究表明, 相对小的叶片更利于生长在热、干、高光照和低养分的环境下(McDonald et al., 2003), 从而增强叶对水热与CO2的传导能力, 降低呼吸和蒸腾的成本。同样的, 在尾叶樱桃(Prunus dielsiana)的天然种群研究中也发现在亚热带东部地区, 夏季具有强烈的高温高湿、强风降雨、干湿交替的气候特征, 相应的尾叶樱桃表现出了叶宽度减小, 叶面积减小, 叶脉密度增加的适应性特征(朱弘等, 2018)。随着最干旱季平均气温的升高, 松树的针叶长度变短, 气孔密度降低, 木质增厚层增加; 而随着降水量的增加, 气孔大小增加; 体现了热带松树对高温、高湿以及季风气候的综合适应特征。当然值得注意的是, 针叶解剖结构的变异同时也会受到土壤、CO2、光照强度等因素的影响, 在未来的研究中还需要进一步考虑这些环境因子的影响。
4 结论
综上所述, 3种松树的针叶表型性状具有丰富的种间和种群间变异, 说明其具有丰富的遗传多样性和表型多样性。因此, 在对它们进行种质资源保存时应当尽可能收集更多种群, 同时也应注意增加种群内的样本数。3种松树的针叶性状均呈现出一定的纬度变异。针叶长度和气孔密度主要受到最干季平均气温的影响, 而气孔腔大小、保卫细胞长度和宽度以及木质增厚层等性状主要受降水相关的气候变量(最湿月降水量、 最干月降水量、 降水量季节变异系数)影响, 在一定程度上介导了松树的蒸腾和呼吸作用对环境的适应。本研究为进一步解析针叶性状的适应性分化提供了重要的理论基础, 同时也可为松树的引种、育种的地理种源选择提供参考, 对我国松属物种的种质资源保存和良种繁育具有重要的指导意义。
致谢
感谢林木遗传育种国家重点实验室开放课题(TGB2021002)的资助。诚挚地感谢佛罗里达大学董乙乂博士在针叶性状的统计分析以及文章的返修过程中提供的帮助。感谢中国科学院西双版纳热带植物园Kyle W. Tomlinson研究员在数据分析中提供的指导, 以及感谢中国科学院西双版纳热带植物园生物多样性研究组提供的实验设备。
附录I 针叶性状示意图
Supplement I Schematic diagram of needle leaf traits
附录II 云南松、卡西亚松和思茅松种群个体间性状比较
Supplement II Comparison of population individual traits among Pinus yunnanensis, P. kesiya and P. kesiya var. langbianensis
参考文献
The morphological traits of needles as markers of geographical differentiation in european Pinus sylvestris populations
Genetic adaptation and phenotypic plasticity contribute to greater leaf hydraulic tolerance in response to drought in warmer climates
DOI:10.1093/treephys/tpx005
PMID:28338733
[本文引用: 1]

The ability of plants to maintain an intact water transport system in leaves under drought conditions is intimately linked to survival and can been be seen as adaptive in shaping species climatic limits. Large differences in leaf hydraulic vulnerability to drought are known among species from contrasting climates, yet whether this trait varies among populations within a single species and, furthermore, whether it is altered by changes in growth conditions, remain unclear. We examined intraspecific variation in both leaf water transport capacity (Kleaf) and leaf hydraulic vulnerability to drought (P50leaf) among eight populations of Corymbia calophylla (R. Br.) K.D. Hill & L.A.S. Johnson (Myrtaceae) from both cool and warm climatic regions grown reciprocally under two temperature treatments representing the cool and warm edge of the species distribution. Kleaf did not vary between cool and warm-climate populations, nor was it affected by variable growth temperature. In contrast, population origin and growth temperature independently altered P50leaf. Using data pooled across growth temperatures, cool-climate populations showed significantly higher leaf hydraulic vulnerability (P50leaf = -3.55 ± 0.18 MPa) than warm-climate populations (P50leaf = -3.78 ± 0.08 MPa). Across populations, P50leaf decreased as population home-climate temperature increased, but was unrelated to rainfall and aridity. For populations from both cool and warm climatic regions, P50leaf was lower under the warmer growth conditions. These results provide evidence of trait plasticity in leaf hydraulic vulnerability to drought in response to variable growth temperature. Furthermore, they suggest that climate, and in particular temperature, may be a strong selective force in shaping intraspecific variation in leaf hydraulic vulnerability to drought.© The Author 2017. Published by Oxford University Press. All rights reserved. For Permissions, please email: journals.permissions@oup.com.
Morphological evaluation of the Pinus kesiya complex (Pinaceae)
DOI:10.1007/s00606-013-0880-0 URL [本文引用: 2]
Analysis on phenotypic variations of fruit and core traits of Melia azedarach from different seed sources
不同产地苦楝果实和果核表型多样性分析
Relationship between geographical distribution of Pinus yunnanensis and climate
云南松的地理分布与气候关系
Advances in the research on Pinus yunnanensis forest
云南松研究综述
Components of leaf-trait variation along environmental gradients
DOI:10.1111/nph.v228.1 URL [本文引用: 1]
Changes in morphology, anatomy, and photosynthetic capacity of needles of Japanese larch (Larix kaempferi) seedlings grown in high CO2 concentrations
DOI:10.1023/B:PHOT.0000040587.99518.a8 URL [本文引用: 1]
Analysis on the “normalization” of drought in Yunnan Province
云南干旱“常态化”的分析
Demography and speciation history of the homoploid hybrid pine Pinus densata on the Tibetan Plateau
DOI:10.1111/j.1365-294X.2012.05712.x URL [本文引用: 1]
Study on genetic structure of Pinus massoniana population by isozyme
用同工酶研究马尾松群体的遗传结构
Phenotypic diversity in natural populations of Picea balfouriana in Sichuan, China
川西云杉天然种群表型多样性
DOI:10.3773/j.issn.1005-264x.2009.02.006
[本文引用: 2]
为了揭示川西云杉(Picea balfouriana)天然种群表型变异程度和变异规律, 以四川甘孜藏族自治州天然分布的川西云杉为研究对象, 调查了12个种群360个单株的种实和针叶表型性状, 采用变异系数、巢式方差分析、相关分析和聚类分析等方法, 分析了种群间和种群内表型多样性。结果表明: 川西云杉表型性状在种群间和种群内存在极其丰富的多样性, 种群间表型分化系数均值为36.53%, 小于种群内变异(63.47%); 球果、针叶、种鳞和种翅的表型分化系数均值分别为47.15%、31.93%、21.89%和45.14%, 球果、针叶、种鳞和种翅4类表型性状的变异系数分别为12.56%、22.16%、12.61%和16.53%, 种鳞的变异系数和表型分化系数最小, 表明其性状最为稳定; 12个表型性状间多数呈显著或极显著正相关, 球果长、球果径、针叶长和种鳞长为川西云杉重要的表型性状。川西云杉表型变异在空间分布上呈现以经度(球果和针叶)和纬度(种鳞和种翅)并存的单向变异模式; 利用种群间欧氏距离进行聚类分析, 可以把川西云杉12个种群划分为3类。
Genetic structure of needle morphological and anatomical traits of Pinus yunnanensis
DOI:10.1007/s11676-015-0133-x URL [本文引用: 2]
Does climate-related in situ variability of Scots pine (Pinus sylvestris L.) needles have a genetic basis? Evidence from common garden experiments
DOI:10.1093/treephys/tpy145
PMID:30715504
[本文引用: 1]
The correlations of phenotypic traits with environmental drivers suggest that variability of these traits is a result of natural selection, especially if such trait correlations are based on genetic variability. We hypothesized that in situ correlations of structural needle traits of Scots pine (Pinus sylvestris L) with minimal winter temperature (Tmin) reported previously from a temperate/boreal transect would be conserved when plants are cultivated under common conditions. We tested this hypothesis by analyzing needles from two common gardens located in the temperate zone, one including adult trees and the other juvenile seedlings. The majority of adult needle traits for which correlations with Tmin were found in the field turned out to be under environmental influence. In contrast, the majority of traits studied in juvenile needles were correlated with the original Tmin suggesting the role of past natural selection in shaping their variability. Juvenile needles thus appeared to be inherently less plastic than adult needles, perhaps reflecting the stronger selective pressure acting during juvenile, as compared with adult, ontogenetic stage. Genetically based cold-climate adaptation in either juvenile or adult needles, or both, involved an increase in leaf mass per area and leaf density, decrease in needle length, reduction in the amount of xylem and phloem, increase in thickness of epidermis, decrease in tracheid diameter and increase in tracheid density, and increase in diameter and volume fraction of resin ducts. We also show that at least some traits, such as transverse xylem and phloem areas and number of fibers, scale with needle length, suggesting that climate-related trait variation may also be mediated by changes in needle length. Moreover, slopes of these allometric relationships may themselves be plastically modified. The phenotypic syndrome typical of needles from cold environments may thus be under environmental, genetic and allometric control.© The Author(s) 2019. Published by Oxford University Press. All rights reserved. For Permissions, please email: journals.permissions@oup.com.
Relationships among Cedrus libani, C. brevifolia and C. atlantica as revealed by the morphological and anatomical needle characters
DOI:10.1007/s00606-012-0700-y URL [本文引用: 3]
Genetic variation and spatial geographical trend of needles, cones and seeds traits for natural populations of Picea linzhinesis
林芝云杉天然群体针叶与种实的变异及其地理趋势
Phylogenomic and ecological analyses reveal the spatiotemporal evolution of global pines
DOI:10.1073/pnas.2022302118 [本文引用: 1]
In situ determination of guard cell ion flux underpins the mechanism of ABA-mediated stomatal closure in barley plants exposed to PEG-induced drought stress
DOI:10.1016/j.envexpbot.2021.104468 [本文引用: 1]
Phenotypic variations in cones and seeds of natural Pinus kesiya var. langbianensis populations in Yunnan Province, China
DOI:10.3724/SP.J.1258.2013.00103 URL [本文引用: 2]
思茅松天然群体种实表型变异
DOI:10.3724/SP.J.1258.2013.00103
[本文引用: 2]
以云南省思茅松(Pinus kesiya var. langbianensis)天然分布区的11个种群的16个种实性状为研究对象, 采用巢式方差分析、变异系数、相关分析和非加权配对算术平均法(UPGMA)等多种分析方法, 探讨思茅松种群间和种群内的表型变异。结果表明: 思茅松种实表型性状在种群间和种群内均存在着较丰富的差异, 种群内的变异(54.76%)大于种群间的变异(10.44%), 种群间平均分化系数为11.95%, 分化程度相对较小。球果总种子数的平均变异系数最高(35.51%), 其次是球果质量(35.1%), 种子大小的平均变异系数最小(8.86%), 成为最稳定的种实性状; 景谷县的表型多样性最丰富, 景洪市则最小。球果和种子大部分表型性状之间存在显著或极显著相关, 其中球果长和球果质量越大, 种鳞长、种子长、种翅长、千粒重、球果总种鳞数和球果总种子数就越大。生态因子中, 年平均气温对表型性状影响最大, 其次是1月平均气温和>5 ℃积温。利用种群间聚类分析可以把思茅松的11个种群分为2类4个亚类, 表型性状依据地理距离进行聚类, 聚类结果与年降水量和>5 ℃积温相关性显著。
Studies on the variation of fruiting traits in natural populations of Picea asperata
云杉天然群体种实性状变异研究
Photosynthesis and nitrogen allocation in needles in the sun and shade crowns of hybrid larch saplings: effect of nitrogen application
DOI:10.1007/s11099-012-0049-z URL [本文引用: 1]
Leaf-size divergence along rainfall and soil-nutrient gradients: Is the method of size reduction common among clades
DOI:10.1046/j.1365-2435.2003.00698.x URL [本文引用: 1]
Environmental contribution to needle variation among natural populations of Pinus tabuliformis
DOI:10.1007/s11676-018-0722-6 [本文引用: 1]
Wood density and mechanical properties of Pinus kesiya Royle ex Gordon in Malawi
DOI:10.3390/f7070135 [本文引用: 1]
Latitudinal variation in morphological traits of the genus Pinus and its relation to environmental and phylogenetic signals
A new, vapour-phase mechanism for stomatal responses to humidity and temperature
Evidence that longer needle retention of spruce and pine populations at high elevations and high latitudes is largely a phenotypic response
There is abundant evidence that evergreen conifers living at high elevations or at high latitudes have longer-lived needles than trees of the same species living elsewhere. This pattern is likely caused by the influence of low temperature in combination with related factors such as a short growing season and low nutrient availability. Because it is not known to what degree such patterns result from phenotypic versus genotypic variation, we evaluated needle longevity for common-garden-grown lowland populations of European Scots pine (Pinus sylvestris L.) of wide latitudinal origin and Norway spruce (Picea abies L.) of wide elevational origin. Nine-year-old trees of 16 Scots pine populations ranging in origin from 47 degrees to 60 degrees N were studied in Kórnik, Poland (52 degrees N) and 18-year-old trees of 18 Norway spruce populations ranging in origin from 670 to 1235 m elevation in southwestern Poland were studied near Morawina, Poland (51 degrees N, 180 m elevation). There was no tendency in either species for populations from northern or high elevation origins to retain needles longer than other populations. All of the Scots pine populations had between 2.5 to 3.0 needle age cohorts and all of the Norway spruce populations had between 6.4 and 7.2 needle age cohorts. Thus, extended needle retention in Scots pine and Norway spruce populations in low-temperature habitats at high elevations and high latitudes appears to be largely an environmentally regulated phenotypic acclimation.
Geographical and ecological distributions of the two tropical pines, Pinus kesiya and Pinus merkusii, in southeast Asia
Genetic diversity among populations in Scotch pine (Pinus silvestris L.) seed stands of Western Black Sea Region in Turkey
Altitudinal variation of some morphological characters of scots pine (Pinus sylvestris L.) in Turkey
Distribution and ecology of the Pinus kesiya complex//Armitage FB, Burley J
Colonization of the Tibetan Plateau by the homoploid hybrid pine Pinus densata
DOI:10.1111/mec.2011.20.issue-18 URL [本文引用: 1]
Genetic evidence for the presence of cytoplasmic DNA in pollen and megagametophytes and maternal inheritance of mitochondrial DNA in Pinus
Phenotypic variability: its components, measurement and underlying developmental processes
DOI:10.1007/s11692-007-9008-1 URL [本文引用: 1]
A review of the research status of Pinus kesiya var. langbianensis in southwestern China
思茅松研究现状的探讨
Needle morphological evidence of the homoploid hybrid origin of Pinus densata based on analysis of artificial hybrids and the putative parents, Pinus tabuliformis and Pinus yunnanensis
DOI:10.1002/ece3.2014.4.issue-10 URL [本文引用: 2]
Pinus prekesiya sp nov from the upper miocene of Yunnan, southwestern China and its biogeographical implications
DOI:10.1016/j.revpalbo.2009.12.008 URL [本文引用: 1]
Diversity and geographical variations of germplasm resources of Armeniaca mandshurica
DOI:10.17521/cjpe.2019.0060 URL [本文引用: 1]
东北杏种质资源多样性及其地理变化
DOI:10.17521/cjpe.2019.0060
[本文引用: 1]
东北杏(Armeniaca mandshurica)是集观赏、经济、用材于一体的重要树种, 长期以来主要处于野生和半野生状态, 鲜有相关研究报道。该研究对东北杏主要分布区内种质资源状况开展了调查, 选择来自辽宁、吉林与黑龙江的47份典型种质进行了22个定量描述性状和7个定性描述性状的观测, 旨在为该树种的种质资源收集、评价与保护提供重要参考。采用变异系数等指标分析定量描述性状多样性, 使用频率分布等指标分析定性描述性状多样性。通过趋势面方法分析定量描述性状的地理变化规律。利用99对SSR引物对47份东北杏种质进行PCR扩增, 应用遗传相似系数分析东北杏种质的遗传多样性。基于表型性状和SSR分子标记采用聚类分析方法分别对种质进行分类。东北杏不同种质间呈现出较高的表型多样性, 其中19个定量描述性状指标的变异系数在9.40%-55.98%之间, 变异系数最大的为小枝长度, 变异系数最小的为种仁宽; 7个定性描述性状的Shannon-Wiener指数在0.58-1.22之间。由于调查区域的地理位置与主要气候因子间存在着显著的相关性, 东北杏种质的定量描述性状与其地理位置有着密切的关系。其中, 小枝长度呈由东向西逐渐增大的梯度变化, 小枝粗度、种子质量呈由北向南逐渐增大的梯度变化, 果柄长呈由东北向西南逐渐增大的梯度变化; 小枝长度、果柄长与海拔呈正相关关系, 小枝粗度与海拔呈负相关关系, 种子质量与海拔相关性很小。基于26个表型性状进行系统聚类, 将47份东北杏种质分为4类, 分类结果主要体现了东北杏种质特征的差别, 同时也在一定程度上体现了种质产地的效应; 基于遗传相似系数进行聚类分析, 将47份东北杏种质也分为4类, 分类结果体现了种质产地效应; 卡方检验表明, 两种聚类结果相关性不显著, 外在环境是影响东北杏表型变异的重要因素。
Study on the phenotypic differentiation of cone traits among Pinus yunnanensis Franch. natural populations
云南松天然群体球果表型变异研究
Needle Phenotypic Variation among Populations of Pinus yunnanensis at Different Altitude
云南松不同海拔群体的针叶性状表型多样性研究
Spatial and temporal evolution characteristics of drought in Yunnan Province from 1969 to 2018 based on SPI/SPEI
DOI:10.1007/s11270-019-4287-6 [本文引用: 1]
Growth-climate sensitivity of two pine species shows species-specific changes along temperature and moisture gradients in southwest China
DOI:10.1016/j.agrformet.2022.108907 [本文引用: 2]
Growth rhythm for half-sib families of young Pinus kesiya plantations in southern Yunnan, China
卡西亚松家系幼林的生长节律
Genetic analysis of needle morphological and anatomical traits among nature populations of Pinus tabuliformis
DOI:10.5539/jps.v6n1p62
URL
[本文引用: 1]
The morphological and anatomical traits of needles are important to evaluate geographic variation and population dynamics of conifer species. Variations of morphological and anatomical needle traits in coniferous species are considered to be the consequence of genetic evolution, and be used in geographic variation and ecological studies, etc. Pinus tabuliformis is a particular native coniferous species in northern and central China. For understanding its adaptive evolution in needle traits, the needle samplings of 10 geographic populations were collected from a 30yr provenience common garden trail that might eliminate site environment effect and show genetic variation among populations and 20 needle morphological and anatomical traits were involved. The results showed that variations among and within populations were significantly different over all the measured traits and the variance components within population were generally higher than that among populations in the most measured needle traits. Population heritabilities in all measured traits were higher than 0.7 in common garden sampling among populations. Needle traits were more significantly correlated with longitude than other factors. First five principal components accounted for 81.6% of the variation with eigenvalues greater than 1; the differences among populations were mainly dependent on needle width, stomatal density, section areas of vascular bundle, total resin canals, and mesophyll, as well as area ratio traits. Ten populations were divided into two categories by Euclidean distance. Variations in needle traits among the populations have shown systematic microevolution in terms of geographic impact on P. tabuliformis. This study would provide empirical data to characterize adaptation and genetic variation of P. tabuliformis, which should be more available for ecological studies.
Discrimination of Pinus yunnanensis, P. kesiya and P. densata by FT-NIR
Geographic cline and genetic introgression effects on seed morphology variation and germination fitness in two closely related pine species in southeast Asia
DOI:10.3390/f13030374 [本文引用: 1]
Leaf phenotypic variation in natural populations of Cerasus dielsiana
DOI:10.17521/cjpe.2018.0196 URL [本文引用: 1]
尾叶樱桃天然种群叶表型性状变异研究
DOI:10.17521/cjpe.2018.0196
[本文引用: 1]
为揭示中国特有植物尾叶樱桃(Cerasus dielsiana)在现代核心分布区天然种群的叶表型地理变异规律及其生态适应性特征, 该研究通过多重比较、巢氏方差分析、相关性分析、主成分分析(PCA)、主坐标分析(PCoA)、非加权配对算术平均法(UPGMA)聚类分析等数理方法, 对来自四川、湖北、湖南、江西、台湾5省8个尾叶樱桃天然种群的11个叶表型性状进行了比较分析, 研究其不同地理单元间叶表型多样性和地理变异规律及对地理气候的响应。结果显示: 1)尾叶樱桃主要叶表型性状变异在种群内和种群间均存在显著差异, 平均变异系数为22.44%, 其中变异系数最大和最小的分别为叶面积(50.83%)与一级侧脉数(7.96%); 平均叶表型性状的分化系数为30.78%, 种群内的变异(51.55%)大于种群间的变异(22.55%)。2) PCA表明对尾叶樱桃叶表型性状变异起主要贡献作用的前三大主成分累计贡献率达到92.400%, 可以综合概括和排序为“大小性状” (73.242%)与“形状性状” (19.158%)。3)叶宽(r = -0.641)、叶面积(r = -0.658)和一级侧脉数(r = 0.659)性状均与经度呈显著负相关或正相关关系, 气温季节变化和最湿季降水量对叶表型性状变异影响较大。4)基于PCoA和UPGMA聚类分析可将8个天然种群划分为4类。尾叶樱桃天然种群叶表型性状变异丰富, 在数量上表现出一定的连续性, “大小性状”是性状间变异的主要来源, 平均表型分化处于中等程度水平, 种群内是叶表型性状变异的主要来源; 各种群间表型分化划分结果与地理位置基本一致, 在地理空间上呈现以经度为主的梯度变异模式, “气候变异性”与“展叶期降水量”是驱动叶表型性状变异的主要气候因子, 推测这是尾叶樱桃在长期进化中与亚热带季风气候环境相适应的结果。

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}