野扁桃(Amygdalus ledebouriana )也称矮扁桃, 属蔷薇科李亚科桃属扁桃亚属植物, 是一古老的物种(朱京琳, 1983 ), 已被列入国家重点保护野生植物名录(鲁兆莉等, 2021 )。野扁桃不仅抗干旱还耐严寒。其果实有较高营养价值, 同时在药用方面也有研究价值(李疆等, 2006 ), 野扁桃还可以用作果树嫁接的优良矮化砧木材料, 也是选育扁桃抗性杂交新品种的最佳材料(张敏等, 1995 )。新疆的野扁桃在原生境下受到人类活动及放牧的影响, 种群数量和质量逐年下降, 蓄积量锐减, 其种群的天然更新存在问题, 因此深入了解其天然更新规律, 对该物种的保护和野果林生态恢复至关重要。
植物种群的年龄结构可以反映种群内不同年龄阶段个体的比例(李博, 2000 )。通过对植物种群在不同生境、不同时期的个体数量变化的研究, 可以反映植物种群生物学特性受环境的影响(卢晶等, 2022 )。分析植物种群年龄结构状态, 探索种群动态变化, 可以对目前种群的生存状态和种群与生境间的影响关系有所了解(袁宏波等, 2011 ), 且对于了解种群干扰情况和预测种群未来发展具有重要意义(何斌等, 2020 ; 张金峰等, 2022 )。对于植物种群年龄结构及动态分析已有不少研究报道: 濒危植物鹅掌楸(Liriodendron chinense )天然种群中中小龄级个体数量相对较多, 未来一段时间内将发展良好, 但幼龄与老龄个体严重缺乏(罗西等, 2021 )。江西南方红豆杉(Taxus wallichiana var. mairei )天然种群空间结构整体呈聚集分布, 幼苗、幼树极少, 自然更新能力差, 种群结构均呈两头小、中间大的纺锤型, 种群呈衰退趋势(喻龙华等, 2021 )。因此, 可以通过对种群年龄结构的研究, 深入探讨种群更新过程, 以及结构与动态的内在联系。新疆天山野果林准噶尔山楂(Crataegus songarica )种群现阶段的生长呈增长趋势, 但种群的长期稳定维持存在困难, 建议适当补充幼苗、幼株, 减少对成年树的砍伐破坏, 从而为种群的保育和恢复提供保障(杨蕾等, 2018 )。在研究新疆野苹果(Malus sieversii )种群年龄结构及其动态特征中, 新疆野苹果受虫害、旅游、放牧干扰影响, 时间序列分析预测野苹果种群老龄个体数量增加, 中、幼龄个体缺乏补充, 呈现不同程度衰退, 应采取人工促萌或补植幼苗, 调控林地密度, 进而加强野苹果种质资源保护(马闯等, 2018 )。结合以上研究可知, 对野果林优势种种群年龄结构的调查结合野果林受人类活动及放牧干扰的现状可以为野果林的生态恢复提供理论依据。
种群生态学对于研究种群内部各成员之间, 种群与生境的相互作用规律有重要意义。而目前有关新疆野扁桃的研究主要集中在生物信息学(王波, 2018 )、抗逆性(宋宏伟等, 2013 )、繁殖生物学(程靖, 2016 )、果实营养成分分析(朱绪春等, 2016 )、遗传多样性(马松梅等, 2019 )等方面, 但是在新疆野扁桃的种群生态学领域方面的研究, 目前尚未见报道。本研究以新疆巴尔鲁克山天然野扁桃种群为研究对象, 利用典型样方法进行种群数量统计, 结合存活曲线及静态生命表, 进行生存分析, 旨在探索野扁桃年龄结构特征及更新受限机制, 以期为新疆野扁桃种群的保护管理及恢复提供一定的理论依据及研究基础。
1 材料和方法
1.1 研究区概况
巴尔鲁克山(45.42°-46.98° N, 82.26°-83.38° E)位于塔城地区裕民县和托里县境内, 年平均气温为4.9-6.9 ℃, 年降水量241 mm, 该地区土壤类型为栗钙土, 有机质含量为5% (曾斌, 2008 ), 野扁桃的分布海拔主要在800-1 300 m, 研究区植被的乔木层主要有野扁桃、野苹果、准噶尔山楂、野杏(Armeniaca vulgaris )等; 灌木层主要有天山樱桃(Cerasus tianshanica )、栒子(Cotoneaster hissaricus )、金丝桃叶绣线菊(Spiraea hypericifolia )等; 草本植物主要有菥蓂(Thlaspi arvense )、苦马豆(Sphaerophysa salsula )、塔城棘豆(Oxytropis schrenkii )等。
1.2 研究方法
1.2.1 样地调查
2021年8月, 经过充分踏查, 通过典型样方法, 选取新疆野扁桃种群分布集中且保存较好的地段设置样方, 调查区内野扁桃呈斑块状分布, 故设置不同坡向的规格为5 m × 5 m的21个样方(表1 )。采用每木检尺法, 调查记录样地内野扁桃植株的基径(BD)、高度、冠幅以及幼苗(BD ≤ 0.3 cm)的高度和个体数量, 记录灌木植物的种类、数量、高度和冠幅等信息, 同时记录各样地的放牧程度、中心点坐标、坡向、坡位、坡度以及海拔等生境特征。
1.2.2 种群径级结构划分
本研究采用“空间代替时间”的方法(谢晋阳, 1992 )。由于野扁桃生长缓慢, 株高和冠幅的生物学特性差异不大, 加之存在放牧和牲畜啃食的情况, 故利用野扁桃基径的大小级代替年龄结构来研究其组成结构(高婷婷等, 2010 ), 将野扁桃种群划分为13个龄级, 第I龄级的基径≤0.3 cm, 第II到第XII龄级每级间隔0.3 cm, 即第II龄级的基径为0.3-0.6 cm, 0.6-0.9 cm为第III龄级, 0.9-1.2 cm为第IV龄级, 1.2-1.5 cm为第V龄级, 1.5-1.8 cm为第VI龄级, 1.8-2.1 cm为第VII龄级, 2.1-2.4 cm为第VIII龄级, 2.4-2.7 cm为第IX龄级, 2.7-3.0 cm为第X龄级, 3.0-3.3 cm为第XI龄级, 3.3-3.6 cm为第XII龄级, 第XIII龄级的基径>3.6 cm。统计各龄级野扁桃株数, 绘制野扁桃年龄结构图。
1.2.3 匀化修正数据
在静态生命表的编制过程中, 针对种群死亡率为负值的情况, 采用匀滑技术对统计数据进行处理(江洪, 1992 )。
根据野外调査资料, 新疆野扁桃的第I、II龄级的数据发生波动, 第I、II龄级新疆野扁桃的存活数小于第III龄级。根据特定生命表假设, 其各年龄组合是稳定的, 各年龄个体数的比例不变。修匀可以针对一个参数或一个参数系列。因此, 将数据分为一个区段, 即I-VII龄级。计算这一区段存活数的累积:
(1) $N=\sum\limits_{i=1}^{7}{axi}=4955$
(2) $ax=\frac{N}{n}=\frac{4955}{7}=707.86\approx 708$
式中, N 为样地内调查的野扁桃总株数, ax 为需匀滑修正各龄级的平均株数, 且认为这个平均数是该区段的组中值, i 为龄级数, n 为需匀滑修正的龄级数。根据这个区段最多存活数与最少存活数差值为600, 区段间隔数量为7, 可以确定这个区段每一相邻年龄组存活数之间的差为86。ax 经匀滑修正后得a'x (表2 )。
1.2.4 静态生命表的编制
静态生命表可以反映多年生植物种群的出生率和死亡率的变化规律, 因此可用静态生命表对多年生植物的种群动态进行分析(杨小平等, 2020 )。通过野扁桃种群各龄级的株数可编制野扁桃种群的静态生命表(程子敏等, 2018 )。计算公式如下:
(3) ${{l}_{x}}=\left( \frac{{{a}_{x}}}{{{a}_{0}}} \right)\times 1000$
(4) ${{d}_{x}}={{l}_{x}}-{{l}_{x+1}}$
(5) ${{q}_{x}}=\left( \frac{{{d}_{x}}}{{{l}_{x}}} \right)\times 100%$
(6) ${{L}_{x}}=\left( \frac{{{l}_{x}}+{{l}_{x+1}}}{2} \right)$
(7) ${{T}_{x}}=\sum\limits_{x}^{\infty }{{{L}_{x}}}$
(8) ${{e}_{x}}=\frac{{{T}_{x}}}{{{l}_{x}}}$
(9) ${{K}_{x}}=\ln {{l}_{x}}-\ln {{l}_{x+1}}$
(10) ${{S}_{x}}=\frac{{{l}_{x+1}}}{lx}$
式中, x 为龄级, a 0 为种群初始个体数; ax 表示在x 龄级的存活数, lx 表示在x 龄级开始时标准化存活个体数, dx 是从x 到x + 1龄级间隔期内标准化死亡个体数, qx 是从x 到x + 1龄级间隔期间的死亡率, Lx 为从x 到x + 1龄级间隔期间还存活的个体数, Tx 是从x 龄级到超过x 龄级的总个体数, ex 是进入x 龄级个体的期望寿命, Kx 是x 龄级的消失率, Sx 是x 龄级的存活率。
1.2.5 种群动态量化分析
本研究采用动态量化分析法定量描述野扁桃种群结构以及对外界环境干扰的敏感性(陈晓德, 1998 ), 量化公式如下:
(11) ${{V}_{x}}=\frac{{{S}_{x}}-{{S}_{x-1}}}{\max ({{S}_{x}},{{S}_{x+1}})}\times 100%$
(12) ${{V}_{pi}}=(\frac{1}{\sum\limits_{x=1}^{k-1}{{{S}_{x}}}})\times \sum\limits_{x=1}^{k-1}{({{S}_{x}}\times {{V}_{x}})}$
(13) ${{{V}'}_{pi}}=\frac{\sum\limits_{x=1}^{k-1}{({{S}_{x}}\times {{V}_{x}})}}{k\times \min ({{S}_{1}},{{S}_{2}},{{S}_{3}},\cdots,{{S}_{k}})\times \sum\limits_{x=1}^{k-1}{{{S}_{x}}}}$
(14) ${{P}_{\max }}=\frac{1}{k\times \min ({{S}_{1}},{{S}_{2}},{{S}_{3}},\cdots,{{S}_{k}})}$
式中, Vx 为种群内两个相邻龄级之间个体数量变化的动态指数, Vpi 、V′pi 分别为忽略外部环境干扰和考虑到存在未来外部干扰时整个种群的数量动态变化指数; Sx 、Sx+ 1 分别为第x 、x + 1龄级种群个体数; k 为龄级数量; P max 表示种群在外部环境干扰下承担的风险概率最大值。
Vx 、Vpi 、V′pi 取正值、0、负值时分别对应种群内相邻的两个邻级(或整个种群)个体数量增长、稳定和衰退的趋势。
1.2.5 死亡率、消失率曲线及存活曲线拟合
以龄级为横坐标, 死亡率、消失率为纵坐标, 分别绘制种群死亡率曲线和消失率曲线。以龄级为横坐标, 以野扁桃标准个体存活数的对数(lnlx )为纵坐标绘制种群存活曲线。Deevey存活曲线一般有3种基本类型, I型存活曲线种群内的大多数个体均能达到生理寿命, II型存活曲线种群内各龄级具有相同的死亡率, III型存活曲线种群内幼株死亡率高, 幼苗期之后, 种群死亡率低且稳定。本研究采用Hett和Loucks (1976 )提出的数学模型对野扁桃种群存活曲线进行检验, 即用指数方程(公式15)和幂函数方程(公式16)来验证种群属于上述哪种类型。方程如下:
(15) ax ' = a 0 e– bx
(16) ax ' = a 0 x – b
式中, ax ' 为匀滑后x 龄级内存活数, a 0 为种群初始个体数, b 为死亡率。
1.2.6 生存分析方法
为阐明种群的生存规律, 本研究以野扁桃静态生命表为基础, 引入种群生存率函数(Si )、累计死亡率函数(Fi )、死亡密度函数(fti )和危险率函数(λti ), 4个函数的计算采用下式进行。
(17) ${{S}_{i}}={{S}_{1}}\times {{S}_{2}}\times {{S}_{3}}\cdots {{S}_{x}}$
(18) ${{F}_{i}}=1-{{S}_{i}}$
(19) ${{f}_{ti}}=\frac{{{S}_{i-1}}-{{S}_{i}}}{{{h}_{i}}}$
(20) ${{\lambda }_{ti}}=\frac{2(1-{{S}_{i}})}{{{h}_{i}}(1+{{S}_{i}})}$
其中Si 为存活率, hi 为龄级宽度(杨凤翔等, 1991 ), 通过绘制野扁桃种群生存分析函数曲线图, 进一步分析野扁桃的种群动态。
1.2.7 种群数量动态的时间序列预测
采用时间序列分析中的一次平均推移法对新疆野扁桃种群未来2、3、4、5、6、7、8、9、10个龄级时间内不同龄级个体数量进行预测。计算公式如下:
(21) $M{{t}^{(1)}}=\frac{1}{t}\sum\limits_{k=x-t-1}^{x}{Xk}$
式中, t 为预测时间, x 为龄级, Xk 为k 龄级内的个体数, Mt (1) 为经过t 个龄级时间后x 龄级个体数的预测值。
1.3 数据分析
对照《天山野果林资源: 伊犁野果林综合研究》(林培钧和崔乃然, 2000 ), 核对样地出现的植物物种学名及拉丁名, 本研究所有统计分析在Excel 2007和SPSS 21中进行, 用Origin 2019软件绘图。
2 结果和分析
2.1 种群年龄结构
以龄级为横坐标, 以野扁桃个体数为纵坐标作 图, 得到野扁桃种群的年龄结构图(图1 )。野扁桃种群的个体数随着龄级的增大表现出先增大后减小的趋势, 总体呈现为“纺锤”型; 野扁桃种群主要分布在第III龄级, 以中龄株为主, 个体数为1 065株, 占调查总数的19.19%; 第I、II龄级的个体数为1 085株, 占种群个体总数的19.55%, 其中第I龄级为幼苗, 占种群个体总数的8.38%; 第IV-X龄级属于中龄株, 其个体数为3 290株, 占种群个体总数的59.28%; 第XI龄级之后以老龄株为主, 个体数为110株, 占种群个体总数的1.98%。种群内中龄级个体较为充足, 幼、老龄株个体数占比较小, 所以野扁桃种群年龄结构为衰退型。
图1
图1
新疆野扁桃种群的年龄结构。龄级划分见表2。
Fig. 1
An age structure of Amygdalus ledebouriana population in Xinjiang. Age class see Table 2.
2.2 种群静态生命表
由野扁桃种群静态生命表(表3 )可知, 野扁桃种群幼龄株个体数量稳定, 中龄株个体数量丰富, 老龄株数量较少。野扁桃种群的个体存活数随着龄级的增大逐级减少; 其中, 第VIII龄级出现了较高的死亡率, 达到0.58, 表明野扁桃植株在这个龄级的生活状态已经衰退, 即野扁桃个体基径达到2.1 cm之后便老化严重, 适应性差, 生活能力也快速下降。期望寿命(ex )在第I龄级达到最大, 为5.24, 自第I龄级之后, 野扁桃种群ex 值随着龄级的增长逐级下降, 表明野扁桃种群在幼苗期生长最旺盛, 随着年龄的增长, 生存力与对环境的适应能力逐渐下降, 尤其在第VIII龄级以后, 随着个体生长的过熟, 数量明显减少。
2.3 种群动态分析
2.3.1 种群数量动态
种群数量动态量化分析结果见表4 。野扁桃种群V I -V II 小于0, 而V III -V XII 大于0, 呈现出动态变化, 由此可看出野扁桃种群在I-XII龄级范围内呈现出“衰退-增长-衰退-增长-衰退-增长”的变化趋势。随机干扰风险极值(P max )大于0但数值极小。排除外部干扰, 野扁桃种群的数量变化动态指数Vpi > 0, 可知野扁桃种群为增长型。然而, V′pi 趋近于0, 说明该种群受外界随机干扰时增长趋势不明显。
2.3.2 死亡率与消失率曲线
种群数量随龄级增加的动态变化过程可通过种群的死亡率和消失率反映。图2 显示, 野扁桃种群两条曲线变化趋势基本一致, 随着龄级的增加二者都出现了先增后降, 再增再降的复杂动态变化趋势, 在第VIII龄级死亡率和消失率都达到了最大(0.58和0.87), 说明野扁桃种群在该龄级数量发生锐减。结合种群静态生命表以及种群动态量化分析的结果可知, 该种群处于不稳定的状态。
图2
图2
新疆野扁桃种群死亡率(qx )与消失率(Kx )曲线。龄级划分见表2。
Fig. 2
Curve of mortality rate (qx ) and disappearance rate (Kx ) of Amygdalus ledebouriana populations in Xinjiang. Age class see Table 2.
2.3.3 种群存活曲线
由图3 可以看出野扁桃种群的存活曲线趋势处于Deevey-II型与Deevey-III型之间, 为了检验野扁桃种群存活曲线更趋于Deevey-II型还是Deevey-III型, 采用指数方程和幂函数方程进行验证。
图3
图3
新疆野扁桃种群存活曲线。龄级划分见表2。
Fig. 3
Survival curve of Amygdalus ledebouriana populations in Xinjiang. Age class see Table 2.
依据Hett和Loucks的数学模型得出的检验方程及参数见表5 , 模型检验结果显示, 指数模型的F 值及相关系数均大于幂函数模型的F 检验值和相关系数, 说明野扁桃种群的存活曲线趋近于Deevey-II型。野扁桃种群存活曲线变化平缓, 第I龄级个体存活数量最高, 随着龄级递增, 存活数量逐渐减少, 第I-VII龄级的种群处于平稳状态, 存活率较高, 到第VIII龄级之后, 种群存活数量下降较明显, 在这个阶段的种群处于衰退状态。
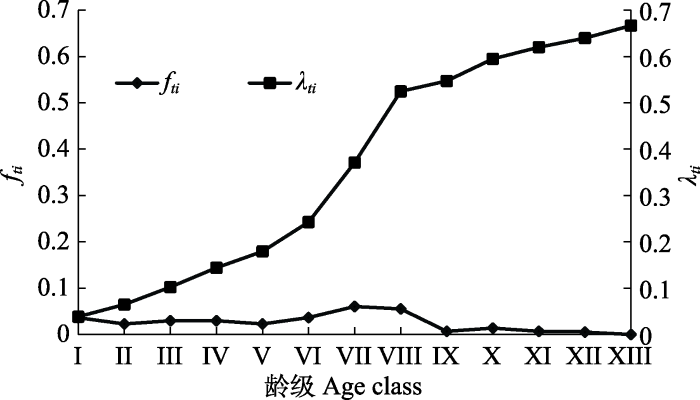
2.4 生存分析
以龄级为横坐标, 分别以4个函数值为纵坐标作图, 绘制生存率曲线、累计死亡率曲线、死亡密度曲线和危险率曲线, 得到图4 、图5 。由图4 得知, 野扁桃种群呈现出随龄级的增大生存率减小、累计死亡率增加的趋势。种群生存率曲线和累计死亡率曲线在第I-VIII龄级升降幅度较大, 在第VIII龄级开始升降比较平缓。在第VI龄级时, 即野扁桃种群个体基径达到1.5 cm时, 种群生存率近乎等于累计死亡率, 说明野扁桃种群在第VI龄级后各项生理功能都逐渐进入衰老期, 种群呈现衰退的趋势。由图5 可以得知, 野扁桃种群的死亡密度曲线在第VI-IX龄级的变化幅度较大, 其余龄级变化很小, 整体趋势平稳, 而危险率曲线在第I-VIII龄级变化波动大, 之后较平稳, 和累计死亡率曲线变化趋势基本一致, 综上, 野扁桃种群第I-VIII龄级生命状态不稳定, 死亡数量较多, 种群内数量呈下降趋势, 自第VIII龄级以后为衰退趋势, 这与静态生命表、存活曲线的分析结果是相符的。
图4
图4
新疆野扁桃种群生存率(Si )与累计死亡率(Fi )曲线。龄级划分见表2。
Fig. 4
Survival rate (Si ) and cumulative mortality rate (Fi ) curve of Amygdalus ledebouriana populations in Xinjiang. Age class see Table 2.
图5
图5
新疆野扁桃种群死亡密度(fti )与危险率(λti )曲线。龄级划分见表2。
Fig. 5
Mortality density (fti ) and risk rate (λti ) curve of Amygdalus ledebouriana populations in Xinjiang. Age class see Table 2.
2.5 时间序列预测
通过野扁桃种群时间序列预测表(表6 )可知, II、III龄级个体数都在未来2个龄级时间后减少; IV、V、VI、VII和VIII龄级分别在经过2、3、4、5、6个龄级时间后出现峰值, 随后个体数开始减少。以上结果说明野扁桃种群在未来会出现幼、中龄个体减少的趋势; 而IX、X、XI、XII和XIII龄级个体数在未来是以稳定增加的趋势呈现的。由此可推测, 未来野扁桃种群幼、中龄个体缺乏, 老龄个体占主导地位, 但是随着龄级越高, 经历的时间越长, 种群个体数量增加速率逐渐变缓; 第VIII龄级之后的个体数增长速率明显缓于第VIII龄级之前的(9.8%和12.04%), 这也说明新疆野扁桃自第VIII龄级开始生理能力出现衰退。
3 讨论
3.1 新疆野扁桃种群年龄结构特征
种群年龄结构特征同时受到生物学特性和环境因素共同影响, 且种群年龄结构特征能体现种群动态变化及未来发展趋势(Svensson & Jeglum, 2001 ; 李俊清, 2006 )。通过对新疆野扁桃种群的年龄结构进行研究, 可以揭示种群生存现状并提出更新策略。不同灌木种群的年龄结构及其发展趋势是各不相同的, 贾喆亭等(2021 )研究发现阿拉善高原珍珠猪毛菜(Salsola passerina )种群结构以稳定型为主, 其次是增长型, 成年个体占比高, 种群整体趋于稳定。李清河等(2009 )发现半日花(Helianthemum songaricum )种群的个体数量在不同坡位存在下降趋势, 中龄级个体多, 幼龄级和老龄级个体少。本研究发现, 野扁桃种群不同龄级个体数呈现为由中龄级向幼龄级与老龄级两侧递减的趋势, 中龄级稳定生长, 占据优势, 而幼龄级缺乏, 可以看出野扁桃种群总体趋于衰退型种群结构特征。当种群处在第VIII龄级, 即达到老龄阶段之后种群数量发生锐减, 原因可能是, 野扁桃作为一种灌木, 在到达老龄阶段时, 其根部萌生的幼苗与老龄株争夺养分, 加之老龄个体的生理能力及对环境的适应性都降低, 故野扁桃种群老龄个体数量减少。在未来的发展中, 野扁桃种群虽然能暂时适应当地的生存环境, 但由于幼龄级个体储备不足, 无法维持中、老龄个体数量, 因此种群整体的长期稳定难以维持。这与牛皮杜鹃(Rhododendron aureum )及水曲柳(Fraxinus mandshurica )的种群年龄结构特征(易雪梅等, 2015 ; 金慧等, 2017 )相似。野扁桃的种群结构是受其自然生境及繁殖方式等因素共同影响的, 裕民野巴旦杏保护区内冬季长时间被深厚积雪覆盖, 受严寒气候影响, 部分老龄株死亡; 野外实地调查发现野扁桃主要以萌蘖繁殖为主, 虽然结实量大, 但是生境内土壤贫瘠、有大量的碎石岩块, 自然下落的种子很难进入土层中萌发, 并且野果林内放牧情况严重, 灌丛下萌发的幼苗及散落的种子很容易遭到牲畜的啃食, 故幼苗在建成过程中受到阻碍, 这在一定程度上限制了野扁桃种群的自然更新, 使野扁桃种群更新和发展比较困难。
3.2 新疆野扁桃种群动态趋势
存活曲线的走势反映了生存率随龄级的变化, 种群动态的基本趋势可以通过种群的存活曲线进行直接分析。对于野生果树而言, 其存活曲线的表现形式并不是单一的。田润炜等(2016 )发现塔城地区野苹果种群呈Deevey-I型, 野苹果老龄个体在较差的自然环境中加快死亡, 而幼龄个体补充不足, 所以整体衰退。杨小平等(2020 )发现野生樱桃李(Prunus cerasifera )种群生存曲线属于Deevey-II型, 由于野生樱桃李从幼苗期过渡到中龄株的时期存在严重的天然更新障碍, 所以幼苗大量死亡。本研究中, 新疆野扁桃种群存活曲线显示其基本属于Deevey-II型, 结合总体数量动态变化指数Vpi > 0, 可知种群为增长型。新疆野扁桃种群“早期-中期-后期”表现为“减少-稳定-衰退”的特征, 与姜在民等(2018 )对濒危植物羽叶丁香(Syringa pinnatifolia )种群结构与动态特征的研究结果相同。新疆野扁桃种群在第II-V龄级的死亡率较低且稳定, 可能是因为随着植株个体的发育, 对空间和资源的需求逐渐增大, 种内和种间竞争作用增强, 但在第I以及第V龄级以后出现较高的死亡率, 在第VIII龄级的死亡率最高, 说明老龄株个体发展遇到了瓶颈, 这可能是种群生理衰老导致的, 由此可以看出野扁桃种群在未来会走向衰退, 这也是小范围、片段化生境下野生植物种群必然经历的发展过程(蔡新斌和吴俊侠, 2016 ; 刘海燕等, 2016 )。通过引入生存分析的4个函数可以明显地看出野扁桃种群的结构和动态变化, 其中生存率曲线和累计死亡率曲线的变化幅度大, 在到第V龄级阶段时, 种群生存率和累计死亡率几乎相等, 种群达到平衡状态, 此后种群逐渐向衰退方向发展, 生存空间、资源环境有限以及个体衰老导致种群进入生理衰老期, 呈现衰退的趋势, 这与存活曲线反映的结果一致。综合以上分析, 野扁桃种群中幼龄个体的缺乏会导致种群逐渐走向濒危, 因此保护幼龄个体的生态环境以及增强种群自身繁育和扩散能力是阻止野扁桃种群衰退和进行种群恢复的关键。
从时间序列预测分析上看, 在经过2-10个龄级时间之后, 野扁桃种群第II-VIII龄级个体数逐渐减少, 这说明从幼龄株转变为中龄株有一定难度; 第VIII龄级之后, 个体数在未来逐渐减少, 由此可以反映野扁桃在进入中龄阶段后生存能力下降, 并且野扁桃种群内幼龄株个体数量占比较小, 整个野扁桃种群个体数量集中在中龄级, 在不受外界环境干扰的前提下, 种群会呈现增长趋势, 然而, 一旦植株生理衰退, 种群数量便会急剧减少。综合以上分析, 认为幼龄个体的缺乏是导致野扁桃种群更新受限的主要原因, 因此, 补充幼龄个体即对野扁桃繁殖的保护是阻止新疆野扁桃种群衰退和进行种群恢复的关键。
3.3 新疆野扁桃种群适应性保护及恢复对策
新疆野扁桃种群幼龄个体偏少, 目前种群的稳定主要靠中龄个体维持, 且龄级越大的植株危险率越高, 越容易死亡。植物种群数量的减少, 不仅包括幼苗供给量不足的情况。还包括成活个体存在死亡的情况, 可以看出, 野扁桃种群现状是其自身生态学特性及外在环境影响共同作用的结果, 因此要开展实生苗繁育及回归工作提高幼苗存活率及存活质量, 为防止远交衰退带来的遗传没化的影响, 应采集原生种群的种子进行实生苗的培育; 新疆野扁桃根蘖繁殖的幼苗距离母株的最佳生存范围仍需进一步探索; 针对环境因素的干扰, 应该在新疆野扁桃的自然分布区采取封育保护措施, 避免对野扁桃生境的破坏, 以维持野扁桃种群的自然繁殖演替; 同时, 在近地保护的基础上深入研究其人工植苗更新技术及迁地栽培技术, 从而促进人工干扰下种群的恢复, 扩大种群数量及其分布面积, 辅助种群壮大, 增强种群自身繁育及扩散能力。
参考文献
View Option
[1]
Cai XB Wu JX (2016 ). Characteristics and dynamics analysis of Haloxylon persicum populations in the Nature Reserve of Gan Jia Lake
Journal of Arid Land Resources and Environment 30 (7 ), 90 -94 .
[本文引用: 1]
[蔡新斌 , 吴俊侠 (2016 ). 甘家湖自然保护区白梭梭种群特征与动态分析
干旱区资源与环境 , 30 (7 ), 90 -94 .]
[本文引用: 1]
[2]
Chen XD (1998 ). Study on dynamic quantitative analysis method of plant population and community structure
Acta Ecologica Sinica 18 , 214 -217 .
[本文引用: 1]
[陈晓德 (1998 ). 植物种群与群落结构动态量化分析方法研究
生态学报 , 18 : 214 -217 .]
[本文引用: 1]
[3]
Cheng J (2016 ). Study on Reproductive Biology of Amygdalus ledebouriana in Xinjiang
Master degree dissertation, Xinjiang Agricultural University, Ürümqi .
[本文引用: 1]
[程靖 (2016 ). 新疆野扁桃繁殖生物学研究
硕士学位论文, 新疆农业大学, 乌鲁木齐 .]
[本文引用: 1]
[4]
Cheng ZM Wang N Mu F (2018 ). Structure and dynamic of natural Armeninca sibirica populations at different altitudes in mountain areas of northern Hebei
Acta Botanica Boreali-Occidentalia Sinica 38 , 2303 -2313 .
[本文引用: 1]
[程子敏 , 王南 , 穆枫 (2018 ). 冀北山地不同海拔天然山杏种群的结构与动态
西北植物学报 , 38 , 2303 -2313 .]
[本文引用: 1]
[5]
Gao TT Li QH Xu J Xin ZM Chu JM (2010 ). Sexual reproduction system characteristics of rare eremophyte Helianthemum soongoricum
Acta Botanica Boreali- Occidentalia Sinica 30 , 1982 -1988 .
[本文引用: 1]
[高婷婷 , 李清河 , 徐军 , 辛智鸣 , 褚建民 (2010 ). 荒漠珍稀灌木半日花繁殖生物学特性研究
西北植物学报 , 30 , 1982 -1988 .]
[本文引用: 1]
[6]
He B Li Q Chen QL Xue XH Li WJ (2020 ). Structure and dynamic characteristics of Pinus armandii population in karst region, northwest of Guizhou Province
Journal of Ecology and Rural Environment 36 , 788 -795 .
[本文引用: 1]
[何斌 , 李青 , 陈群利 , 薛晓辉 , 李望军 (2020 ). 黔西北岩溶区华山松(Pinus armandii )种群结构及动态特征
生态与农村环境学报 , 36 , 788 -795 .]
[本文引用: 1]
[7]
Hett JM Loucks OL (1976 ). Age structure models of balsam fir and eastern hemlock
Journal of Ecology 64 , 1029 -1044 .
DOI:10.2307/2258822
URL
[本文引用: 1]
[8]
Jia ZT Yang JY Sun YX Chen Q Yan RL (2021 ). Spatial distribution pattern of Salsola passerina population in Alashan Plateau
Journal of Desert Research 41 (1 ), 119 -128 .
[本文引用: 1]
[贾喆亭 , 杨九艳 , 孙艳霞 , 陈琪 , 闫瑞玲 (2021 ). 阿拉善高原珍珠猪毛菜(Salsola passerina )种群空间分布格局
中国沙漠 , 41 (1 ), 119 -128 .]
DOI:10.7522/j.issn.1000-694X.2020.00041
[本文引用: 1]
为探究阿拉善高原珍珠猪毛菜(Salsola passerina)种群生长习性和空间格局分布机制,采用体积划分龄级方法和Ripley’s K函数对具有代表性的珍珠猪毛菜种群年龄结构、空间分布格局及不同龄级间的空间关联性进行研究。结果表明:珍珠猪毛菜种群结构以稳定型为主,其次是增长型,成年个体占比高,种群整体趋于稳定。随干旱程度增加,珍珠猪毛菜种群及各龄级空间分布格局由复杂向单一转变,即由聚集分布或均匀分布向随机分布转变。在降水较多的种群内,各龄级间的空间关联性相对复杂,随尺度增大由正关联转变为无关联,再转变为负关联。在较干旱地区,整体上表现为无关联。在阿拉善高原,珍珠猪毛菜种群空间分布特性由其本身生物学特性与环境因素共同作用,在生态建设中应遵循种群与生境间相互关系,发挥其饲用及防风固沙价值。
[9]
Jiang H (1992 ). Population Ecology of Spruce China Forestry Publishing House , Beijing . 7 -13 .
[本文引用: 1]
[江洪 (1992 ). 云杉种群生态学 . 中国林业出版社 , 北京 . 7 -13 .]
[本文引用: 1]
[10]
Jiang ZM He ZS Su H Zhao H Cai J (2018 ). Population structure and dynamic characteristics of endangered Syringa pinnatifolia Hemsl
Acta Ecologica Sinica 38 , 2471 -2480 .
[本文引用: 1]
[姜在民 , 和子森 , 宿昊 , 赵涵 , 蔡靖 (2018 ). 濒危植物羽叶丁香种群结构与动态特征
生态学报 , 38 , 2471 -2480 .]
[本文引用: 1]
[11]
Jin H Zhao Y Yin H Qin LW Liu LJ Wang C Jia X Li BY (2017 ). Quantitative characteristics and dynamic analysis of the endangered species Rhododendron chrysanthum population in Changbai Mountain
Chinese Journal of Ecology 36 , 3123 -3130 .
[本文引用: 1]
[金慧 , 赵莹 , 尹航 , 秦立武 , 刘丽杰 , 王超 , 贾翔 , 李冰岩 (2017 ). 长白山濒危植物牛皮杜鹃(Rhododendron chrysanthum )种群数量特征与动态分析
生态学杂志 , 36 , 3123 -3130 .]
[本文引用: 1]
牛皮杜鹃为国家Ⅲ级重点保护植物,种群数量有限,处于濒危状态。本文运用种群静态生命表、存活曲线、个体数量动态预测等方法分析了长白山高山苔原带不同海拔牛皮杜鹃种群数量特征及动态发展趋势。结果表明:1)牛皮杜鹃种群大小级结构呈纺锤形,属衰退型种群;2)随海拔升高,种群增长潜力逐渐减小,非完全随机干扰下,种群增长潜力有限,低海拔的V<sub>pi</sub>″接近于0;3)种群存活曲线趋于Deevey Ⅲ型,高海拔出现较高的死亡率和致死力,种群退化明显;4)在经历1、2、4个龄级后,Ⅳ龄级个体数量会不同程度增加,种群在生长后期个体数量得到补充;5)牛皮杜鹃种群数量主要靠自身匍匐生根形成分株来维持。整个种群向大龄级个体集中,高海拔面临较强的环境胁迫,会比中低海拔更早进入衰退期。导致其濒危的可能原因是牛皮杜鹃的生物学特性及人为干扰。
[12]
Li B (2000 ). Ecology Higher Education Press , Beijing . 46 .
[本文引用: 1]
[李博 (2000 ). 生态学 . 高等教育出版社 , 北京 . 46 .]
[本文引用: 1]
[13]
Li J Zeng B Luo SP Li HL Wulazihan M (2006 ). Protection and propagation of Amygdalus Ledebouriana in China
Xinjiang Agriculture Science 43 , 61 -62 .
[本文引用: 1]
[李疆 , 曾斌 , 罗淑萍 , 李海龙 , 马达尼亚提·吾拉孜汗 , (2006 ). 我国新疆野扁桃资源的保护及引种繁育
新疆农业科学 , 43 , 61 -62 .]
[本文引用: 1]
[14]
Li JQ (2006 ). Forest Ecology Higher Education Press , Beijing . 186 -192 .
[本文引用: 1]
[李俊清 (2006 ). 森林生态学 . 高等教育出版社 , 北京 . 186 -192 .]
[本文引用: 1]
[15]
Li QH Gao TT Liu JF Xin ZM Xu J (2009 ). The age structure and life table of rare eremophyte Helianthemum ordosicum population
Bulletin of Botanical Research 29 , 176 -181 .
[本文引用: 1]
[李清河 , 高婷婷 , 刘建锋 , 辛智鸣 , 徐军 (2009 ). 荒漠珍稀灌木半日花种群的年龄结构与生命表分析
植物研究 , 29 , 176 -181 .]
DOI:10.7525/j.issn.1673-5102.2009.02.009
[本文引用: 1]
根据对不同坡位的半日花种群的调查资料,分析了其年龄结构,编制了静态生命表,分析了存活曲线和死亡率曲线、损失度曲线,并利用生存分析理论进行了函数分析。结果表明:不同坡位的半日花种群存在下降趋势。幼龄级和老龄级个体少,中龄级个体多。不同坡位的半日花种群密度存在差异,坡上部种群密度最大,坡下部最小,坡中部居中。不同坡位半日花种群在Ⅰ龄级时期望寿命达到最大,并在Ⅶ、Ⅷ龄级时期望寿命出现波动;不同坡位的半日花种群的存活曲线整体上趋于Deevy Ⅱ~Ⅲ型之间;在第Ⅵ~Ⅷ龄级半日花种群死亡率较高,且坡中部和坡下部种群死亡率峰值比坡上部种群滞后一个龄级。生存分析引入生命表中的4个函数能较好地说明不同坡位半日花种群的结构和动态变化。
[16]
Liu HY Yang NK Li YY Hong J Zou TC (2016 ). Population structure and dynamic analysis of the rare and endangered plant Camellia longistyla
Plant Science Journal 34 , 89 -98 .
[本文引用: 1]
[刘海燕 , 杨乃坤 , 李媛媛 , 洪江 , 邹天才 (2016 ). 稀有濒危植物长柱红山茶种群特征及数量动态研究
植物科学学报 , 34 , 89 -98 .]
[本文引用: 1]
[17]
Lin PJ Cui NR (2000 ). Wild Fruit Forests Resources in Tianshan Mountains—Comprehensive Research on Wild Fruit Forests in ILi, Xinjiang, China Chinese Forestry Publishing House , Beijing .
[本文引用: 1]
[林培钧 , 崔乃然 (2000 ). 天山野果林资源: 伊犁野果林综合研究 . 中国林业出版社 , 北京 .]
[本文引用: 1]
[18]
Lu J Ma ZQ Gao PF Fan BL Sun K (2022 ). Changes in the Hippophae tibetana population structure and dynamics, a pioneer species of succession, to altitudinal gradients in the Qilian Mountains, China
Chinese Journal of Plant Ecology 46 , 569 -579 .
DOI:10.17521/cjpe.2021.0419
URL
[本文引用: 1]
[卢晶 , 马宗祺 , 高鹏斐 , 樊宝丽 , 孙坤 (2022 ). 祁连山区演替先锋物种西藏沙棘的种群结构及动态对海拔梯度的响应
植物生态学报 , 46 , 569 -579 .]
DOI:10.17521/cjpe.2021.0419
[本文引用: 1]
西藏沙棘(Hippophae tibetana)是青藏高原高寒区特有的低矮灌木和植被演替先锋物种, 具有优良的水土保持功效, 对高海拔环境表现出良好的生态适应性, 但有关其种群结构及动态对海拔梯度的响应规律少有研究, 阻碍了对西藏沙棘适应高寒生境生态策略的理解。青藏高原东北缘的祁连山区生态脆弱, 乡土物种西藏沙棘常在海拔2 700-3 300 m的高寒退化草地呈斑块状分布, 对该区水源涵养林的维持具有重要作用。该研究以祁连山区3个海拔(2 868、3 012、3 244 m)的西藏沙棘为研究对象, 通过编制静态生命表及绘制存活曲线, 分析西藏沙棘的种群结构特征和种群动态, 并利用种群动态量化分析和时间序列模型定量研究其未来发展趋势。结果显示: 1)西藏沙棘种群的基径、株高和冠幅均随海拔的升高而降低; 3个海拔种群均呈中龄期个体丰富, 老幼龄期个体较少的纺锤型年龄结构, 充足的中龄期个体可以维持种群短期稳定。2)种群存活曲线均为Deevey-II型, 存活能力为低海拔>中海拔>高海拔; 死亡率和消失率均较高, 呈高海拔>低海拔>中海拔的趋势, 3个海拔种群均缺乏幼苗, 未来均将走向衰退, 高海拔种群较其他海拔提前进入衰退期; 各海拔老龄期生命期望值为中海拔>低海拔>高海拔。3)各海拔种群动态指数(V′<sub>pi</sub>)均趋于0, 表明各种群均趋于稳定, 中海拔种群受随机干扰风险概率的极大值(P<sub>max</sub>)最小, 表明受随机干扰时中海拔种群最稳定, 中海拔更适宜西藏沙棘的生存。4)未来2、4、6个龄级时间后, 3个海拔的中、老龄期苗木增多, 小龄期苗木数目减少, 且各海拔种群均面临幼苗补充不及时的风险, 各种群将由稳定向衰退发展, 幼苗的缺少, 种内、种间竞争及环境胁迫是造成种群走向衰退的重要原因。
[19]
Lu ZL Qin HN Jin XH Zhang ZX Yang QW Hong DY Li DZ Li KF Yuan LC Zhou ZH (2021 ). On the necessity, principle and process of updating the List of National Key Protected Wild Plants
Biodiversity Science 29 , 1577 -1582 .
DOI:10.17520/biods.2021394
URL
[本文引用: 1]
[鲁兆莉 , 覃海宁 , 金效华 , 张志翔 , 杨庆文 , 洪德元 , 李德铢 , 李开凡 , 袁良琛 , 周志华 (2021 ). 《国家重点保护野生植物名录》调整的必要性、原则和程序
生物多样性 , 29 , 1577 -1582 .]
DOI:10.17520/biods.2021394
[本文引用: 1]
我国是世界上植物多样性最丰富的国家之一, 1999年发布的《国家重点保护野生植物名录》(下称《名录》(第一批))明确了国家重点保护野生植物的范围, 为依法强化保护、规范无序开发利用、提高公众保护意识奠定了基础。20多年来, 我国野生植物多样性保护形势发生了很大变化, 需要对《名录》进行调整。2018年, 国家林业和草原局、农业农村部启动《名录》调整工作, 物种的遴选遵循了5条基本原则和4条补充性原则, 这些原则主要涉及中国珍稀濒危物种, 具有重要经济、文化、科研、生态等价值物种的入选以及部分物种的排除。经国务院批准, 2021年9月7日, 国家林业和草原局、农业农村部发布了调整后的《名录》, 包括真菌类、藻类、苔藓、石松类和蕨类植物、裸子植物和被子植物, 共计约1,101种(455种和40类)野生植物列入其中。本文简要介绍了《名录》调整的必要性、原则和程序及调整后的情况。
[20]
Luo X Guo QJ Yao L Ai XR Zhu J Huang YX Xue WX Li WY (2021 ). Characteristics of natural population structure of endangered plant Liriodendron chinense
Journal of Central South University of Forestry & Technology 41 (7 ), 115 -123 .
[本文引用: 1]
[罗西 , 郭秋菊 , 姚兰 , 艾训儒 , 朱江 , 黄阳祥 , 薛卫星 , 李玮宜 (2021 ). 濒危植物鹅掌楸的天然种群结构特征
中南林业科技大学学报 , 41 (7 ), 115 -123 .]
[本文引用: 1]
[21]
Ma C Yang ML Zhang YX Yan GR Xu Z (2018 ). Age composition and dynamic characteristics of the main populations of endangered Malus sieversii
Arid Zone Research 35 , 156 -164 .
[本文引用: 1]
[马闯 , 杨美玲 , 张云秀 , 阎国荣 , 许正 (2018 ). 新疆野苹果(Malus sieversii )种群年龄结构及其动态特征
干旱区研究 , 35 , 156 -164 .]
[本文引用: 1]
[22]
Ma SM Wang CC Sun FF Wei B Nie YB (2019 ). Genetic diversity of an endangered plant Amygdalus ledebouriana in Xinjiang
Scientia Silvae Sinicae 55 (9 ), 71 -80 .
[本文引用: 1]
[马松梅 , 王春成 , 孙芳芳 , 魏博 , 聂迎彬 (2019 ). 濒危植物新疆野扁桃的遗传多样性
林业科学 , 55 (9 ), 71 -80 .]
[本文引用: 1]
[23]
Song HW Tian J Zeng B (2013 ). Determination of physiological index of drought resistance of Xinjiang wild almond
Chinese Horticulture Abstracts 29 (12 ), 22 -23 .
[本文引用: 1]
[宋宏伟 , 田嘉 , 曾斌 (2013 ). 新疆野扁桃抗旱性生理指标的测定
中国园艺文摘 , 29 (12 ), 22 -23 .]
[本文引用: 1]
[24]
Svensson JS Jeglum JK (2001 ). Structure and dynamics of an undisturbed old-growth Norway spruce forest on the rising Bothnian coastline
Forest Ecology and Management 151 , 67 -79 .
DOI:10.1016/S0378-1127(00)00697-6
URL
[本文引用: 1]
[25]
Tian RW Cai XB Liu LY Jiang XH Buzaolamu T Maryamgul A (2016 ). Characteristics of the age structure a dynamics of Malus sieversii population
Acta Botanica Boreali- Occidentalia Sinica 36 , 811 -817 .
[本文引用: 1]
[田润炜 , 蔡新斌 , 刘丽燕 , 江晓珩 , 布早拉木·吐尔逊 , 买尔燕古丽·阿不都热合曼 (2016 ). 新疆野苹果种群年龄结构特征与动态分析
西北植物学报 , 36 , 811 -817 .]
[本文引用: 1]
[26]
Wang B (2018 ). Cloning and Bioinformatics Analysis of PetCullin1 Gene in Wild Almond in Xinjiang
Master degree dissertation, Xinjiang Agricultural University, Ürümqi .
[本文引用: 1]
[王波 (2018 ). 新疆野扁桃PetCullin1 基因的克隆与生物信息学分析
硕士学位论文, 新疆农业大学, 乌鲁木齐 .]
[本文引用: 1]
[27]
Xie JY (1992 ). A method of founding the life table of forest plants
Journal of Nanchang University (Natural Science )16 , 383 -386 .
[本文引用: 1]
[谢晋阳 (1992 ). 森林植物立木种群静态生命表的一种编制方法
南昌大学学报(理科版) , 16 , 383 -386 .]
[本文引用: 1]
[28]
Yang FX Wang SQ Xu HG Li BQ (1991 ). The theory of survival analysis and its application to life table
Acta Ecologica Sinica 11 , 153 -158 .
[本文引用: 1]
[杨凤翔 , 王顺庆 , 徐海根 , 李邦庆 (1991 ). 生存分析理论及其在研究生命表中的应用
生态学报 , 11 , 153 -158 .]
[本文引用: 1]
[29]
Yang L Lü HY Li J Ma X Akebar Y (2018 ). Structure and dynamic analysis of Crataegus songarica K. Koch population in Tianshan wild fruit forest of Xinjiang
Acta Botanica Boreali-Occidentalia Sinica 38 , 2314 -2323 .
[本文引用: 1]
[杨蕾 , 吕海英 , 李进 , 马雪 , 艾克拜尔·依米提 , (2018 ). 新疆天山野果林准噶尔山楂种群结构与动态分析
西北植物学报 , 38 , 2314 -2323 .]
[本文引用: 1]
[30]
Yang XP Zhou L Jiang LL Maola A Wang J (2020 ). Population statistics and survival analysis of Prunus divaricate
Journal of Arid Land Resources and Environment 34 , 154 -160 .
[本文引用: 2]
[杨小平 , 周龙 , 蒋丽丽 , 艾克拜尔·毛拉 , 王瑾 (2020 ). 野生樱桃李的种群统计与生存分析
干旱区资源与环境 , 34 , 154 -160 .]
[本文引用: 2]
[31]
Yi XM Zhang Y Wang YX Ji LZ Wu PL (2015 ). Population structure and dynamics of Fraxinus mandshurica in Changbai Mountain
Acta Ecologica Sinica 35 , 91 -97 .
[本文引用: 1]
[易雪梅 , 张悦 , 王远遐 , 姬兰柱 , 吴培莉 (2015 ). 长白山水曲柳种群动态
生态学报 , 35 , 91 -97 .]
[本文引用: 1]
[32]
Yu LH Li YQ Chen ZM Zeng PS Zhang LL Yao JB Zhou XH Liu SZ Deng ZF (2021 ). Community and population structure of natural Taxus chinensis var. mairei in Jiangxi Province
Journal of Central South University of Forestry & Technology 41 (11 ), 164 -172 .
[本文引用: 1]
[喻龙华 , 厉月桥 , 陈珍明 , 曾平生 , 张利利 , 姚甲宝 , 周新华 , 刘素贞 , 邓宗付 (2021 ). 江西天然南方红豆杉群落及种群结构特征
中南林业科技大学学报 , 41 (11 ), 164 -172 .]
[本文引用: 1]
[33]
Yuan HB Zhang JC Chu JM Liu SJ Tang JN Ding F (2011 ). Characteristics of the age structures of three typical plant populations in Kumtag desert
Acta Botanica Boreali- Occidentalia Sinica 31 , 2304 -2309 .
[本文引用: 1]
[袁宏波 , 张锦春 , 褚建民 , 刘淑娟 , 唐进年 , 丁峰 (2011 ). 库姆塔格沙漠典型植物种群年龄结构特征
西北植物学报 , 31 , 2304 -2309 .]
[本文引用: 1]
[34]
Zeng B (2008 ). A Study on Propagate Biological Characteristics and Genetic Diversity of Germplasm Resources of Amygdalus ledebouriana Schlecht
PhD dissertation, Xinjiang Agricultural University, Ürümqi .
[本文引用: 1]
[曾斌 (2008 ). 新疆野扁桃繁殖生物学特性及种质资源遗传多样性研究
博士学位论文, 新疆农业大学, 乌鲁木齐 .]
[本文引用: 1]
[35]
Zhang JF Ge SS Liang JH Li JQ (2022 ). Population age structure and dynamics of Pinus koraiensis in broadleaved Korean pine forest in Changbai Mountain, China
Chinese Journal of Plant Ecology 46 , 667 -677 .
DOI:10.17521/cjpe.2021.0498
URL
[本文引用: 1]
[张金峰 , 葛树森 , 梁金花 , 李俊清 (2022 ). 长白山阔叶红松林红松种群年龄结构与数量动态特征
植物生态学报 , 46 , 667 -677 .]
DOI:10.17521/cjpe.2021.0498
[本文引用: 1]
该研究以分布于长白山阔叶红松林内的红松(Pinus koraiensis)种群为对象, 通过编制种群静态生命表, 计算数量动态指数, 绘制存活曲线、死亡率曲线和消失率曲线, 应用4个生存函数并引入谱分析和时间序列预测模型, 分析红松种群年龄结构, 揭示其天然更新过程及未来发展趋势, 以期为野生红松种群的保护和恢复提供科学依据。结果显示: 红松种群数量变化具有阶段性, 幼龄(I-III龄级)和成龄(VII-X龄级)个体数量多, 中龄(IV-VI龄级)和老龄(XI-XIV龄级)个体数量少, 形成明显间断的两个优势年龄分布区。种群存活曲线趋近于Deevey-Ⅲ型, 表明幼龄个体死亡率高。忽略外部干扰时的总体数量动态变化指数大于>0, 表明红松种群为增长型; 考虑未来外部干扰时的种群动态变化指数趋近于0, 结合死亡率和消失率曲线呈现出连续先增后降的复杂动态变化趋势, 可知该种群受外界随机干扰时增长不明显。生存函数分析显示, 红松种群具有前期锐减、中期稳定、后期衰退的特点。谱分析表明红松种群天然更新呈周期性波动。未来2、3、4、5、6、7、8、9、10个龄级时间后, 红松幼、中龄个体数逐渐减少, 而成、老龄个体数量将逐渐增加。幼龄个体死亡率高、生存空间和资源条件有限, 老龄个体生理衰老明显是限制红松种群增长的主要原因。建议加强幼龄个体的抚育工作, 提高其存活率和生存质量; 保护和改善生存环境, 从而促进红松种群的自然更新和恢复。
[36]
Zhang M Pa HT Yan PX (1995 ). Analysis of amino acid components in Xinjiang almond
Special Wild Economic Animal and Plant Research 4 ), 47 .
[本文引用: 1]
[张敏 , 帕哈提 , 晏佩霞 (1995 ). 新疆巴旦杏氨基酸组分分析
特产研究 , (4 ), 47 .]
[本文引用: 1]
[37]
Zhu JL (1983 ). Xinjiang Paddan Almond Xinjiang People’s Publishing House , Ürümqi . 137 -155 .
[本文引用: 1]
[朱京琳 (1983 ). 新疆巴旦杏 . 新疆人民出版社 , 乌鲁木齐 . 137 -155 .]
[本文引用: 1]
[38]
Zhu XC Wu YTN Jiang ZM Huang MZ (2016 ). Physicochemical properties and fatty acid compositions of oils from three kinds of wild almond
China Oils and Fats 41 (3 ), 93 -95 .
[本文引用: 1]
[朱绪春 , 乌云塔娜 , 姜仲茂 , 黄梦真 (2016 ). 3种野生扁桃油脂的理化性质及脂肪酸组成研究
中国油脂 , 41 (3 ), 93 -95 .]
[本文引用: 1]
甘家湖自然保护区白梭梭种群特征与动态分析
1
2016
... 存活曲线的走势反映了生存率随龄级的变化, 种群动态的基本趋势可以通过种群的存活曲线进行直接分析.对于野生果树而言, 其存活曲线的表现形式并不是单一的.田润炜等(2016 )发现塔城地区野苹果种群呈Deevey-I型, 野苹果老龄个体在较差的自然环境中加快死亡, 而幼龄个体补充不足, 所以整体衰退.杨小平等(2020 )发现野生樱桃李(Prunus cerasifera )种群生存曲线属于Deevey-II型, 由于野生樱桃李从幼苗期过渡到中龄株的时期存在严重的天然更新障碍, 所以幼苗大量死亡.本研究中, 新疆野扁桃种群存活曲线显示其基本属于Deevey-II型, 结合总体数量动态变化指数Vpi > 0, 可知种群为增长型.新疆野扁桃种群“早期-中期-后期”表现为“减少-稳定-衰退”的特征, 与姜在民等(2018 )对濒危植物羽叶丁香(Syringa pinnatifolia )种群结构与动态特征的研究结果相同.新疆野扁桃种群在第II-V龄级的死亡率较低且稳定, 可能是因为随着植株个体的发育, 对空间和资源的需求逐渐增大, 种内和种间竞争作用增强, 但在第I以及第V龄级以后出现较高的死亡率, 在第VIII龄级的死亡率最高, 说明老龄株个体发展遇到了瓶颈, 这可能是种群生理衰老导致的, 由此可以看出野扁桃种群在未来会走向衰退, 这也是小范围、片段化生境下野生植物种群必然经历的发展过程(蔡新斌和吴俊侠, 2016 ; 刘海燕等, 2016 ).通过引入生存分析的4个函数可以明显地看出野扁桃种群的结构和动态变化, 其中生存率曲线和累计死亡率曲线的变化幅度大, 在到第V龄级阶段时, 种群生存率和累计死亡率几乎相等, 种群达到平衡状态, 此后种群逐渐向衰退方向发展, 生存空间、资源环境有限以及个体衰老导致种群进入生理衰老期, 呈现衰退的趋势, 这与存活曲线反映的结果一致.综合以上分析, 野扁桃种群中幼龄个体的缺乏会导致种群逐渐走向濒危, 因此保护幼龄个体的生态环境以及增强种群自身繁育和扩散能力是阻止野扁桃种群衰退和进行种群恢复的关键. ...
甘家湖自然保护区白梭梭种群特征与动态分析
1
2016
... 存活曲线的走势反映了生存率随龄级的变化, 种群动态的基本趋势可以通过种群的存活曲线进行直接分析.对于野生果树而言, 其存活曲线的表现形式并不是单一的.田润炜等(2016 )发现塔城地区野苹果种群呈Deevey-I型, 野苹果老龄个体在较差的自然环境中加快死亡, 而幼龄个体补充不足, 所以整体衰退.杨小平等(2020 )发现野生樱桃李(Prunus cerasifera )种群生存曲线属于Deevey-II型, 由于野生樱桃李从幼苗期过渡到中龄株的时期存在严重的天然更新障碍, 所以幼苗大量死亡.本研究中, 新疆野扁桃种群存活曲线显示其基本属于Deevey-II型, 结合总体数量动态变化指数Vpi > 0, 可知种群为增长型.新疆野扁桃种群“早期-中期-后期”表现为“减少-稳定-衰退”的特征, 与姜在民等(2018 )对濒危植物羽叶丁香(Syringa pinnatifolia )种群结构与动态特征的研究结果相同.新疆野扁桃种群在第II-V龄级的死亡率较低且稳定, 可能是因为随着植株个体的发育, 对空间和资源的需求逐渐增大, 种内和种间竞争作用增强, 但在第I以及第V龄级以后出现较高的死亡率, 在第VIII龄级的死亡率最高, 说明老龄株个体发展遇到了瓶颈, 这可能是种群生理衰老导致的, 由此可以看出野扁桃种群在未来会走向衰退, 这也是小范围、片段化生境下野生植物种群必然经历的发展过程(蔡新斌和吴俊侠, 2016 ; 刘海燕等, 2016 ).通过引入生存分析的4个函数可以明显地看出野扁桃种群的结构和动态变化, 其中生存率曲线和累计死亡率曲线的变化幅度大, 在到第V龄级阶段时, 种群生存率和累计死亡率几乎相等, 种群达到平衡状态, 此后种群逐渐向衰退方向发展, 生存空间、资源环境有限以及个体衰老导致种群进入生理衰老期, 呈现衰退的趋势, 这与存活曲线反映的结果一致.综合以上分析, 野扁桃种群中幼龄个体的缺乏会导致种群逐渐走向濒危, 因此保护幼龄个体的生态环境以及增强种群自身繁育和扩散能力是阻止野扁桃种群衰退和进行种群恢复的关键. ...
植物种群与群落结构动态量化分析方法研究
1
1998
... 本研究采用动态量化分析法定量描述野扁桃种群结构以及对外界环境干扰的敏感性(陈晓德, 1998 ), 量化公式如下: ...
植物种群与群落结构动态量化分析方法研究
1
1998
... 本研究采用动态量化分析法定量描述野扁桃种群结构以及对外界环境干扰的敏感性(陈晓德, 1998 ), 量化公式如下: ...
新疆野扁桃繁殖生物学研究
1
2016
... 种群生态学对于研究种群内部各成员之间, 种群与生境的相互作用规律有重要意义.而目前有关新疆野扁桃的研究主要集中在生物信息学(王波, 2018 )、抗逆性(宋宏伟等, 2013 )、繁殖生物学(程靖, 2016 )、果实营养成分分析(朱绪春等, 2016 )、遗传多样性(马松梅等, 2019 )等方面, 但是在新疆野扁桃的种群生态学领域方面的研究, 目前尚未见报道.本研究以新疆巴尔鲁克山天然野扁桃种群为研究对象, 利用典型样方法进行种群数量统计, 结合存活曲线及静态生命表, 进行生存分析, 旨在探索野扁桃年龄结构特征及更新受限机制, 以期为新疆野扁桃种群的保护管理及恢复提供一定的理论依据及研究基础. ...
新疆野扁桃繁殖生物学研究
1
2016
... 种群生态学对于研究种群内部各成员之间, 种群与生境的相互作用规律有重要意义.而目前有关新疆野扁桃的研究主要集中在生物信息学(王波, 2018 )、抗逆性(宋宏伟等, 2013 )、繁殖生物学(程靖, 2016 )、果实营养成分分析(朱绪春等, 2016 )、遗传多样性(马松梅等, 2019 )等方面, 但是在新疆野扁桃的种群生态学领域方面的研究, 目前尚未见报道.本研究以新疆巴尔鲁克山天然野扁桃种群为研究对象, 利用典型样方法进行种群数量统计, 结合存活曲线及静态生命表, 进行生存分析, 旨在探索野扁桃年龄结构特征及更新受限机制, 以期为新疆野扁桃种群的保护管理及恢复提供一定的理论依据及研究基础. ...
冀北山地不同海拔天然山杏种群的结构与动态
1
2018
... 静态生命表可以反映多年生植物种群的出生率和死亡率的变化规律, 因此可用静态生命表对多年生植物的种群动态进行分析(杨小平等, 2020 ).通过野扁桃种群各龄级的株数可编制野扁桃种群的静态生命表(程子敏等, 2018 ).计算公式如下: ...
冀北山地不同海拔天然山杏种群的结构与动态
1
2018
... 静态生命表可以反映多年生植物种群的出生率和死亡率的变化规律, 因此可用静态生命表对多年生植物的种群动态进行分析(杨小平等, 2020 ).通过野扁桃种群各龄级的株数可编制野扁桃种群的静态生命表(程子敏等, 2018 ).计算公式如下: ...
荒漠珍稀灌木半日花繁殖生物学特性研究
1
2010
... 本研究采用“空间代替时间”的方法(谢晋阳, 1992 ).由于野扁桃生长缓慢, 株高和冠幅的生物学特性差异不大, 加之存在放牧和牲畜啃食的情况, 故利用野扁桃基径的大小级代替年龄结构来研究其组成结构(高婷婷等, 2010 ), 将野扁桃种群划分为13个龄级, 第I龄级的基径≤0.3 cm, 第II到第XII龄级每级间隔0.3 cm, 即第II龄级的基径为0.3-0.6 cm, 0.6-0.9 cm为第III龄级, 0.9-1.2 cm为第IV龄级, 1.2-1.5 cm为第V龄级, 1.5-1.8 cm为第VI龄级, 1.8-2.1 cm为第VII龄级, 2.1-2.4 cm为第VIII龄级, 2.4-2.7 cm为第IX龄级, 2.7-3.0 cm为第X龄级, 3.0-3.3 cm为第XI龄级, 3.3-3.6 cm为第XII龄级, 第XIII龄级的基径>3.6 cm.统计各龄级野扁桃株数, 绘制野扁桃年龄结构图. ...
荒漠珍稀灌木半日花繁殖生物学特性研究
1
2010
... 本研究采用“空间代替时间”的方法(谢晋阳, 1992 ).由于野扁桃生长缓慢, 株高和冠幅的生物学特性差异不大, 加之存在放牧和牲畜啃食的情况, 故利用野扁桃基径的大小级代替年龄结构来研究其组成结构(高婷婷等, 2010 ), 将野扁桃种群划分为13个龄级, 第I龄级的基径≤0.3 cm, 第II到第XII龄级每级间隔0.3 cm, 即第II龄级的基径为0.3-0.6 cm, 0.6-0.9 cm为第III龄级, 0.9-1.2 cm为第IV龄级, 1.2-1.5 cm为第V龄级, 1.5-1.8 cm为第VI龄级, 1.8-2.1 cm为第VII龄级, 2.1-2.4 cm为第VIII龄级, 2.4-2.7 cm为第IX龄级, 2.7-3.0 cm为第X龄级, 3.0-3.3 cm为第XI龄级, 3.3-3.6 cm为第XII龄级, 第XIII龄级的基径>3.6 cm.统计各龄级野扁桃株数, 绘制野扁桃年龄结构图. ...
黔西北岩溶区华山松(Pinus armandii )种群结构及动态特征
1
2020
... 植物种群的年龄结构可以反映种群内不同年龄阶段个体的比例(李博, 2000 ).通过对植物种群在不同生境、不同时期的个体数量变化的研究, 可以反映植物种群生物学特性受环境的影响(卢晶等, 2022 ).分析植物种群年龄结构状态, 探索种群动态变化, 可以对目前种群的生存状态和种群与生境间的影响关系有所了解(袁宏波等, 2011 ), 且对于了解种群干扰情况和预测种群未来发展具有重要意义(何斌等, 2020 ; 张金峰等, 2022 ).对于植物种群年龄结构及动态分析已有不少研究报道: 濒危植物鹅掌楸(Liriodendron chinense )天然种群中中小龄级个体数量相对较多, 未来一段时间内将发展良好, 但幼龄与老龄个体严重缺乏(罗西等, 2021 ).江西南方红豆杉(Taxus wallichiana var. mairei )天然种群空间结构整体呈聚集分布, 幼苗、幼树极少, 自然更新能力差, 种群结构均呈两头小、中间大的纺锤型, 种群呈衰退趋势(喻龙华等, 2021 ).因此, 可以通过对种群年龄结构的研究, 深入探讨种群更新过程, 以及结构与动态的内在联系.新疆天山野果林准噶尔山楂(Crataegus songarica )种群现阶段的生长呈增长趋势, 但种群的长期稳定维持存在困难, 建议适当补充幼苗、幼株, 减少对成年树的砍伐破坏, 从而为种群的保育和恢复提供保障(杨蕾等, 2018 ).在研究新疆野苹果(Malus sieversii )种群年龄结构及其动态特征中, 新疆野苹果受虫害、旅游、放牧干扰影响, 时间序列分析预测野苹果种群老龄个体数量增加, 中、幼龄个体缺乏补充, 呈现不同程度衰退, 应采取人工促萌或补植幼苗, 调控林地密度, 进而加强野苹果种质资源保护(马闯等, 2018 ).结合以上研究可知, 对野果林优势种种群年龄结构的调查结合野果林受人类活动及放牧干扰的现状可以为野果林的生态恢复提供理论依据. ...
黔西北岩溶区华山松(Pinus armandii )种群结构及动态特征
1
2020
... 植物种群的年龄结构可以反映种群内不同年龄阶段个体的比例(李博, 2000 ).通过对植物种群在不同生境、不同时期的个体数量变化的研究, 可以反映植物种群生物学特性受环境的影响(卢晶等, 2022 ).分析植物种群年龄结构状态, 探索种群动态变化, 可以对目前种群的生存状态和种群与生境间的影响关系有所了解(袁宏波等, 2011 ), 且对于了解种群干扰情况和预测种群未来发展具有重要意义(何斌等, 2020 ; 张金峰等, 2022 ).对于植物种群年龄结构及动态分析已有不少研究报道: 濒危植物鹅掌楸(Liriodendron chinense )天然种群中中小龄级个体数量相对较多, 未来一段时间内将发展良好, 但幼龄与老龄个体严重缺乏(罗西等, 2021 ).江西南方红豆杉(Taxus wallichiana var. mairei )天然种群空间结构整体呈聚集分布, 幼苗、幼树极少, 自然更新能力差, 种群结构均呈两头小、中间大的纺锤型, 种群呈衰退趋势(喻龙华等, 2021 ).因此, 可以通过对种群年龄结构的研究, 深入探讨种群更新过程, 以及结构与动态的内在联系.新疆天山野果林准噶尔山楂(Crataegus songarica )种群现阶段的生长呈增长趋势, 但种群的长期稳定维持存在困难, 建议适当补充幼苗、幼株, 减少对成年树的砍伐破坏, 从而为种群的保育和恢复提供保障(杨蕾等, 2018 ).在研究新疆野苹果(Malus sieversii )种群年龄结构及其动态特征中, 新疆野苹果受虫害、旅游、放牧干扰影响, 时间序列分析预测野苹果种群老龄个体数量增加, 中、幼龄个体缺乏补充, 呈现不同程度衰退, 应采取人工促萌或补植幼苗, 调控林地密度, 进而加强野苹果种质资源保护(马闯等, 2018 ).结合以上研究可知, 对野果林优势种种群年龄结构的调查结合野果林受人类活动及放牧干扰的现状可以为野果林的生态恢复提供理论依据. ...
Age structure models of balsam fir and eastern hemlock
1
1976
... 以龄级为横坐标, 死亡率、消失率为纵坐标, 分别绘制种群死亡率曲线和消失率曲线.以龄级为横坐标, 以野扁桃标准个体存活数的对数(lnlx )为纵坐标绘制种群存活曲线.Deevey存活曲线一般有3种基本类型, I型存活曲线种群内的大多数个体均能达到生理寿命, II型存活曲线种群内各龄级具有相同的死亡率, III型存活曲线种群内幼株死亡率高, 幼苗期之后, 种群死亡率低且稳定.本研究采用Hett和Loucks (1976 )提出的数学模型对野扁桃种群存活曲线进行检验, 即用指数方程(公式15)和幂函数方程(公式16)来验证种群属于上述哪种类型.方程如下: ...
阿拉善高原珍珠猪毛菜(Salsola passerina )种群空间分布格局
1
2021
... 种群年龄结构特征同时受到生物学特性和环境因素共同影响, 且种群年龄结构特征能体现种群动态变化及未来发展趋势(Svensson & Jeglum, 2001 ; 李俊清, 2006 ).通过对新疆野扁桃种群的年龄结构进行研究, 可以揭示种群生存现状并提出更新策略.不同灌木种群的年龄结构及其发展趋势是各不相同的, 贾喆亭等(2021 )研究发现阿拉善高原珍珠猪毛菜(Salsola passerina )种群结构以稳定型为主, 其次是增长型, 成年个体占比高, 种群整体趋于稳定.李清河等(2009 )发现半日花(Helianthemum songaricum )种群的个体数量在不同坡位存在下降趋势, 中龄级个体多, 幼龄级和老龄级个体少.本研究发现, 野扁桃种群不同龄级个体数呈现为由中龄级向幼龄级与老龄级两侧递减的趋势, 中龄级稳定生长, 占据优势, 而幼龄级缺乏, 可以看出野扁桃种群总体趋于衰退型种群结构特征.当种群处在第VIII龄级, 即达到老龄阶段之后种群数量发生锐减, 原因可能是, 野扁桃作为一种灌木, 在到达老龄阶段时, 其根部萌生的幼苗与老龄株争夺养分, 加之老龄个体的生理能力及对环境的适应性都降低, 故野扁桃种群老龄个体数量减少.在未来的发展中, 野扁桃种群虽然能暂时适应当地的生存环境, 但由于幼龄级个体储备不足, 无法维持中、老龄个体数量, 因此种群整体的长期稳定难以维持.这与牛皮杜鹃(Rhododendron aureum )及水曲柳(Fraxinus mandshurica )的种群年龄结构特征(易雪梅等, 2015 ; 金慧等, 2017 )相似.野扁桃的种群结构是受其自然生境及繁殖方式等因素共同影响的, 裕民野巴旦杏保护区内冬季长时间被深厚积雪覆盖, 受严寒气候影响, 部分老龄株死亡; 野外实地调查发现野扁桃主要以萌蘖繁殖为主, 虽然结实量大, 但是生境内土壤贫瘠、有大量的碎石岩块, 自然下落的种子很难进入土层中萌发, 并且野果林内放牧情况严重, 灌丛下萌发的幼苗及散落的种子很容易遭到牲畜的啃食, 故幼苗在建成过程中受到阻碍, 这在一定程度上限制了野扁桃种群的自然更新, 使野扁桃种群更新和发展比较困难. ...
阿拉善高原珍珠猪毛菜(Salsola passerina )种群空间分布格局
1
2021
... 种群年龄结构特征同时受到生物学特性和环境因素共同影响, 且种群年龄结构特征能体现种群动态变化及未来发展趋势(Svensson & Jeglum, 2001 ; 李俊清, 2006 ).通过对新疆野扁桃种群的年龄结构进行研究, 可以揭示种群生存现状并提出更新策略.不同灌木种群的年龄结构及其发展趋势是各不相同的, 贾喆亭等(2021 )研究发现阿拉善高原珍珠猪毛菜(Salsola passerina )种群结构以稳定型为主, 其次是增长型, 成年个体占比高, 种群整体趋于稳定.李清河等(2009 )发现半日花(Helianthemum songaricum )种群的个体数量在不同坡位存在下降趋势, 中龄级个体多, 幼龄级和老龄级个体少.本研究发现, 野扁桃种群不同龄级个体数呈现为由中龄级向幼龄级与老龄级两侧递减的趋势, 中龄级稳定生长, 占据优势, 而幼龄级缺乏, 可以看出野扁桃种群总体趋于衰退型种群结构特征.当种群处在第VIII龄级, 即达到老龄阶段之后种群数量发生锐减, 原因可能是, 野扁桃作为一种灌木, 在到达老龄阶段时, 其根部萌生的幼苗与老龄株争夺养分, 加之老龄个体的生理能力及对环境的适应性都降低, 故野扁桃种群老龄个体数量减少.在未来的发展中, 野扁桃种群虽然能暂时适应当地的生存环境, 但由于幼龄级个体储备不足, 无法维持中、老龄个体数量, 因此种群整体的长期稳定难以维持.这与牛皮杜鹃(Rhododendron aureum )及水曲柳(Fraxinus mandshurica )的种群年龄结构特征(易雪梅等, 2015 ; 金慧等, 2017 )相似.野扁桃的种群结构是受其自然生境及繁殖方式等因素共同影响的, 裕民野巴旦杏保护区内冬季长时间被深厚积雪覆盖, 受严寒气候影响, 部分老龄株死亡; 野外实地调查发现野扁桃主要以萌蘖繁殖为主, 虽然结实量大, 但是生境内土壤贫瘠、有大量的碎石岩块, 自然下落的种子很难进入土层中萌发, 并且野果林内放牧情况严重, 灌丛下萌发的幼苗及散落的种子很容易遭到牲畜的啃食, 故幼苗在建成过程中受到阻碍, 这在一定程度上限制了野扁桃种群的自然更新, 使野扁桃种群更新和发展比较困难. ...
1
1992
... 在静态生命表的编制过程中, 针对种群死亡率为负值的情况, 采用匀滑技术对统计数据进行处理(江洪, 1992 ). ...
1
1992
... 在静态生命表的编制过程中, 针对种群死亡率为负值的情况, 采用匀滑技术对统计数据进行处理(江洪, 1992 ). ...
濒危植物羽叶丁香种群结构与动态特征
1
2018
... 存活曲线的走势反映了生存率随龄级的变化, 种群动态的基本趋势可以通过种群的存活曲线进行直接分析.对于野生果树而言, 其存活曲线的表现形式并不是单一的.田润炜等(2016 )发现塔城地区野苹果种群呈Deevey-I型, 野苹果老龄个体在较差的自然环境中加快死亡, 而幼龄个体补充不足, 所以整体衰退.杨小平等(2020 )发现野生樱桃李(Prunus cerasifera )种群生存曲线属于Deevey-II型, 由于野生樱桃李从幼苗期过渡到中龄株的时期存在严重的天然更新障碍, 所以幼苗大量死亡.本研究中, 新疆野扁桃种群存活曲线显示其基本属于Deevey-II型, 结合总体数量动态变化指数Vpi > 0, 可知种群为增长型.新疆野扁桃种群“早期-中期-后期”表现为“减少-稳定-衰退”的特征, 与姜在民等(2018 )对濒危植物羽叶丁香(Syringa pinnatifolia )种群结构与动态特征的研究结果相同.新疆野扁桃种群在第II-V龄级的死亡率较低且稳定, 可能是因为随着植株个体的发育, 对空间和资源的需求逐渐增大, 种内和种间竞争作用增强, 但在第I以及第V龄级以后出现较高的死亡率, 在第VIII龄级的死亡率最高, 说明老龄株个体发展遇到了瓶颈, 这可能是种群生理衰老导致的, 由此可以看出野扁桃种群在未来会走向衰退, 这也是小范围、片段化生境下野生植物种群必然经历的发展过程(蔡新斌和吴俊侠, 2016 ; 刘海燕等, 2016 ).通过引入生存分析的4个函数可以明显地看出野扁桃种群的结构和动态变化, 其中生存率曲线和累计死亡率曲线的变化幅度大, 在到第V龄级阶段时, 种群生存率和累计死亡率几乎相等, 种群达到平衡状态, 此后种群逐渐向衰退方向发展, 生存空间、资源环境有限以及个体衰老导致种群进入生理衰老期, 呈现衰退的趋势, 这与存活曲线反映的结果一致.综合以上分析, 野扁桃种群中幼龄个体的缺乏会导致种群逐渐走向濒危, 因此保护幼龄个体的生态环境以及增强种群自身繁育和扩散能力是阻止野扁桃种群衰退和进行种群恢复的关键. ...
濒危植物羽叶丁香种群结构与动态特征
1
2018
... 存活曲线的走势反映了生存率随龄级的变化, 种群动态的基本趋势可以通过种群的存活曲线进行直接分析.对于野生果树而言, 其存活曲线的表现形式并不是单一的.田润炜等(2016 )发现塔城地区野苹果种群呈Deevey-I型, 野苹果老龄个体在较差的自然环境中加快死亡, 而幼龄个体补充不足, 所以整体衰退.杨小平等(2020 )发现野生樱桃李(Prunus cerasifera )种群生存曲线属于Deevey-II型, 由于野生樱桃李从幼苗期过渡到中龄株的时期存在严重的天然更新障碍, 所以幼苗大量死亡.本研究中, 新疆野扁桃种群存活曲线显示其基本属于Deevey-II型, 结合总体数量动态变化指数Vpi > 0, 可知种群为增长型.新疆野扁桃种群“早期-中期-后期”表现为“减少-稳定-衰退”的特征, 与姜在民等(2018 )对濒危植物羽叶丁香(Syringa pinnatifolia )种群结构与动态特征的研究结果相同.新疆野扁桃种群在第II-V龄级的死亡率较低且稳定, 可能是因为随着植株个体的发育, 对空间和资源的需求逐渐增大, 种内和种间竞争作用增强, 但在第I以及第V龄级以后出现较高的死亡率, 在第VIII龄级的死亡率最高, 说明老龄株个体发展遇到了瓶颈, 这可能是种群生理衰老导致的, 由此可以看出野扁桃种群在未来会走向衰退, 这也是小范围、片段化生境下野生植物种群必然经历的发展过程(蔡新斌和吴俊侠, 2016 ; 刘海燕等, 2016 ).通过引入生存分析的4个函数可以明显地看出野扁桃种群的结构和动态变化, 其中生存率曲线和累计死亡率曲线的变化幅度大, 在到第V龄级阶段时, 种群生存率和累计死亡率几乎相等, 种群达到平衡状态, 此后种群逐渐向衰退方向发展, 生存空间、资源环境有限以及个体衰老导致种群进入生理衰老期, 呈现衰退的趋势, 这与存活曲线反映的结果一致.综合以上分析, 野扁桃种群中幼龄个体的缺乏会导致种群逐渐走向濒危, 因此保护幼龄个体的生态环境以及增强种群自身繁育和扩散能力是阻止野扁桃种群衰退和进行种群恢复的关键. ...
长白山濒危植物牛皮杜鹃(Rhododendron chrysanthum )种群数量特征与动态分析
1
2017
... 种群年龄结构特征同时受到生物学特性和环境因素共同影响, 且种群年龄结构特征能体现种群动态变化及未来发展趋势(Svensson & Jeglum, 2001 ; 李俊清, 2006 ).通过对新疆野扁桃种群的年龄结构进行研究, 可以揭示种群生存现状并提出更新策略.不同灌木种群的年龄结构及其发展趋势是各不相同的, 贾喆亭等(2021 )研究发现阿拉善高原珍珠猪毛菜(Salsola passerina )种群结构以稳定型为主, 其次是增长型, 成年个体占比高, 种群整体趋于稳定.李清河等(2009 )发现半日花(Helianthemum songaricum )种群的个体数量在不同坡位存在下降趋势, 中龄级个体多, 幼龄级和老龄级个体少.本研究发现, 野扁桃种群不同龄级个体数呈现为由中龄级向幼龄级与老龄级两侧递减的趋势, 中龄级稳定生长, 占据优势, 而幼龄级缺乏, 可以看出野扁桃种群总体趋于衰退型种群结构特征.当种群处在第VIII龄级, 即达到老龄阶段之后种群数量发生锐减, 原因可能是, 野扁桃作为一种灌木, 在到达老龄阶段时, 其根部萌生的幼苗与老龄株争夺养分, 加之老龄个体的生理能力及对环境的适应性都降低, 故野扁桃种群老龄个体数量减少.在未来的发展中, 野扁桃种群虽然能暂时适应当地的生存环境, 但由于幼龄级个体储备不足, 无法维持中、老龄个体数量, 因此种群整体的长期稳定难以维持.这与牛皮杜鹃(Rhododendron aureum )及水曲柳(Fraxinus mandshurica )的种群年龄结构特征(易雪梅等, 2015 ; 金慧等, 2017 )相似.野扁桃的种群结构是受其自然生境及繁殖方式等因素共同影响的, 裕民野巴旦杏保护区内冬季长时间被深厚积雪覆盖, 受严寒气候影响, 部分老龄株死亡; 野外实地调查发现野扁桃主要以萌蘖繁殖为主, 虽然结实量大, 但是生境内土壤贫瘠、有大量的碎石岩块, 自然下落的种子很难进入土层中萌发, 并且野果林内放牧情况严重, 灌丛下萌发的幼苗及散落的种子很容易遭到牲畜的啃食, 故幼苗在建成过程中受到阻碍, 这在一定程度上限制了野扁桃种群的自然更新, 使野扁桃种群更新和发展比较困难. ...
长白山濒危植物牛皮杜鹃(Rhododendron chrysanthum )种群数量特征与动态分析
1
2017
... 种群年龄结构特征同时受到生物学特性和环境因素共同影响, 且种群年龄结构特征能体现种群动态变化及未来发展趋势(Svensson & Jeglum, 2001 ; 李俊清, 2006 ).通过对新疆野扁桃种群的年龄结构进行研究, 可以揭示种群生存现状并提出更新策略.不同灌木种群的年龄结构及其发展趋势是各不相同的, 贾喆亭等(2021 )研究发现阿拉善高原珍珠猪毛菜(Salsola passerina )种群结构以稳定型为主, 其次是增长型, 成年个体占比高, 种群整体趋于稳定.李清河等(2009 )发现半日花(Helianthemum songaricum )种群的个体数量在不同坡位存在下降趋势, 中龄级个体多, 幼龄级和老龄级个体少.本研究发现, 野扁桃种群不同龄级个体数呈现为由中龄级向幼龄级与老龄级两侧递减的趋势, 中龄级稳定生长, 占据优势, 而幼龄级缺乏, 可以看出野扁桃种群总体趋于衰退型种群结构特征.当种群处在第VIII龄级, 即达到老龄阶段之后种群数量发生锐减, 原因可能是, 野扁桃作为一种灌木, 在到达老龄阶段时, 其根部萌生的幼苗与老龄株争夺养分, 加之老龄个体的生理能力及对环境的适应性都降低, 故野扁桃种群老龄个体数量减少.在未来的发展中, 野扁桃种群虽然能暂时适应当地的生存环境, 但由于幼龄级个体储备不足, 无法维持中、老龄个体数量, 因此种群整体的长期稳定难以维持.这与牛皮杜鹃(Rhododendron aureum )及水曲柳(Fraxinus mandshurica )的种群年龄结构特征(易雪梅等, 2015 ; 金慧等, 2017 )相似.野扁桃的种群结构是受其自然生境及繁殖方式等因素共同影响的, 裕民野巴旦杏保护区内冬季长时间被深厚积雪覆盖, 受严寒气候影响, 部分老龄株死亡; 野外实地调查发现野扁桃主要以萌蘖繁殖为主, 虽然结实量大, 但是生境内土壤贫瘠、有大量的碎石岩块, 自然下落的种子很难进入土层中萌发, 并且野果林内放牧情况严重, 灌丛下萌发的幼苗及散落的种子很容易遭到牲畜的啃食, 故幼苗在建成过程中受到阻碍, 这在一定程度上限制了野扁桃种群的自然更新, 使野扁桃种群更新和发展比较困难. ...
1
2000
... 植物种群的年龄结构可以反映种群内不同年龄阶段个体的比例(李博, 2000 ).通过对植物种群在不同生境、不同时期的个体数量变化的研究, 可以反映植物种群生物学特性受环境的影响(卢晶等, 2022 ).分析植物种群年龄结构状态, 探索种群动态变化, 可以对目前种群的生存状态和种群与生境间的影响关系有所了解(袁宏波等, 2011 ), 且对于了解种群干扰情况和预测种群未来发展具有重要意义(何斌等, 2020 ; 张金峰等, 2022 ).对于植物种群年龄结构及动态分析已有不少研究报道: 濒危植物鹅掌楸(Liriodendron chinense )天然种群中中小龄级个体数量相对较多, 未来一段时间内将发展良好, 但幼龄与老龄个体严重缺乏(罗西等, 2021 ).江西南方红豆杉(Taxus wallichiana var. mairei )天然种群空间结构整体呈聚集分布, 幼苗、幼树极少, 自然更新能力差, 种群结构均呈两头小、中间大的纺锤型, 种群呈衰退趋势(喻龙华等, 2021 ).因此, 可以通过对种群年龄结构的研究, 深入探讨种群更新过程, 以及结构与动态的内在联系.新疆天山野果林准噶尔山楂(Crataegus songarica )种群现阶段的生长呈增长趋势, 但种群的长期稳定维持存在困难, 建议适当补充幼苗、幼株, 减少对成年树的砍伐破坏, 从而为种群的保育和恢复提供保障(杨蕾等, 2018 ).在研究新疆野苹果(Malus sieversii )种群年龄结构及其动态特征中, 新疆野苹果受虫害、旅游、放牧干扰影响, 时间序列分析预测野苹果种群老龄个体数量增加, 中、幼龄个体缺乏补充, 呈现不同程度衰退, 应采取人工促萌或补植幼苗, 调控林地密度, 进而加强野苹果种质资源保护(马闯等, 2018 ).结合以上研究可知, 对野果林优势种种群年龄结构的调查结合野果林受人类活动及放牧干扰的现状可以为野果林的生态恢复提供理论依据. ...
1
2000
... 植物种群的年龄结构可以反映种群内不同年龄阶段个体的比例(李博, 2000 ).通过对植物种群在不同生境、不同时期的个体数量变化的研究, 可以反映植物种群生物学特性受环境的影响(卢晶等, 2022 ).分析植物种群年龄结构状态, 探索种群动态变化, 可以对目前种群的生存状态和种群与生境间的影响关系有所了解(袁宏波等, 2011 ), 且对于了解种群干扰情况和预测种群未来发展具有重要意义(何斌等, 2020 ; 张金峰等, 2022 ).对于植物种群年龄结构及动态分析已有不少研究报道: 濒危植物鹅掌楸(Liriodendron chinense )天然种群中中小龄级个体数量相对较多, 未来一段时间内将发展良好, 但幼龄与老龄个体严重缺乏(罗西等, 2021 ).江西南方红豆杉(Taxus wallichiana var. mairei )天然种群空间结构整体呈聚集分布, 幼苗、幼树极少, 自然更新能力差, 种群结构均呈两头小、中间大的纺锤型, 种群呈衰退趋势(喻龙华等, 2021 ).因此, 可以通过对种群年龄结构的研究, 深入探讨种群更新过程, 以及结构与动态的内在联系.新疆天山野果林准噶尔山楂(Crataegus songarica )种群现阶段的生长呈增长趋势, 但种群的长期稳定维持存在困难, 建议适当补充幼苗、幼株, 减少对成年树的砍伐破坏, 从而为种群的保育和恢复提供保障(杨蕾等, 2018 ).在研究新疆野苹果(Malus sieversii )种群年龄结构及其动态特征中, 新疆野苹果受虫害、旅游、放牧干扰影响, 时间序列分析预测野苹果种群老龄个体数量增加, 中、幼龄个体缺乏补充, 呈现不同程度衰退, 应采取人工促萌或补植幼苗, 调控林地密度, 进而加强野苹果种质资源保护(马闯等, 2018 ).结合以上研究可知, 对野果林优势种种群年龄结构的调查结合野果林受人类活动及放牧干扰的现状可以为野果林的生态恢复提供理论依据. ...
我国新疆野扁桃资源的保护及引种繁育
1
2006
... 野扁桃(Amygdalus ledebouriana )也称矮扁桃, 属蔷薇科李亚科桃属扁桃亚属植物, 是一古老的物种(朱京琳, 1983 ), 已被列入国家重点保护野生植物名录(鲁兆莉等, 2021 ).野扁桃不仅抗干旱还耐严寒.其果实有较高营养价值, 同时在药用方面也有研究价值(李疆等, 2006 ), 野扁桃还可以用作果树嫁接的优良矮化砧木材料, 也是选育扁桃抗性杂交新品种的最佳材料(张敏等, 1995 ).新疆的野扁桃在原生境下受到人类活动及放牧的影响, 种群数量和质量逐年下降, 蓄积量锐减, 其种群的天然更新存在问题, 因此深入了解其天然更新规律, 对该物种的保护和野果林生态恢复至关重要. ...
我国新疆野扁桃资源的保护及引种繁育
1
2006
... 野扁桃(Amygdalus ledebouriana )也称矮扁桃, 属蔷薇科李亚科桃属扁桃亚属植物, 是一古老的物种(朱京琳, 1983 ), 已被列入国家重点保护野生植物名录(鲁兆莉等, 2021 ).野扁桃不仅抗干旱还耐严寒.其果实有较高营养价值, 同时在药用方面也有研究价值(李疆等, 2006 ), 野扁桃还可以用作果树嫁接的优良矮化砧木材料, 也是选育扁桃抗性杂交新品种的最佳材料(张敏等, 1995 ).新疆的野扁桃在原生境下受到人类活动及放牧的影响, 种群数量和质量逐年下降, 蓄积量锐减, 其种群的天然更新存在问题, 因此深入了解其天然更新规律, 对该物种的保护和野果林生态恢复至关重要. ...
1
2006
... 种群年龄结构特征同时受到生物学特性和环境因素共同影响, 且种群年龄结构特征能体现种群动态变化及未来发展趋势(Svensson & Jeglum, 2001 ; 李俊清, 2006 ).通过对新疆野扁桃种群的年龄结构进行研究, 可以揭示种群生存现状并提出更新策略.不同灌木种群的年龄结构及其发展趋势是各不相同的, 贾喆亭等(2021 )研究发现阿拉善高原珍珠猪毛菜(Salsola passerina )种群结构以稳定型为主, 其次是增长型, 成年个体占比高, 种群整体趋于稳定.李清河等(2009 )发现半日花(Helianthemum songaricum )种群的个体数量在不同坡位存在下降趋势, 中龄级个体多, 幼龄级和老龄级个体少.本研究发现, 野扁桃种群不同龄级个体数呈现为由中龄级向幼龄级与老龄级两侧递减的趋势, 中龄级稳定生长, 占据优势, 而幼龄级缺乏, 可以看出野扁桃种群总体趋于衰退型种群结构特征.当种群处在第VIII龄级, 即达到老龄阶段之后种群数量发生锐减, 原因可能是, 野扁桃作为一种灌木, 在到达老龄阶段时, 其根部萌生的幼苗与老龄株争夺养分, 加之老龄个体的生理能力及对环境的适应性都降低, 故野扁桃种群老龄个体数量减少.在未来的发展中, 野扁桃种群虽然能暂时适应当地的生存环境, 但由于幼龄级个体储备不足, 无法维持中、老龄个体数量, 因此种群整体的长期稳定难以维持.这与牛皮杜鹃(Rhododendron aureum )及水曲柳(Fraxinus mandshurica )的种群年龄结构特征(易雪梅等, 2015 ; 金慧等, 2017 )相似.野扁桃的种群结构是受其自然生境及繁殖方式等因素共同影响的, 裕民野巴旦杏保护区内冬季长时间被深厚积雪覆盖, 受严寒气候影响, 部分老龄株死亡; 野外实地调查发现野扁桃主要以萌蘖繁殖为主, 虽然结实量大, 但是生境内土壤贫瘠、有大量的碎石岩块, 自然下落的种子很难进入土层中萌发, 并且野果林内放牧情况严重, 灌丛下萌发的幼苗及散落的种子很容易遭到牲畜的啃食, 故幼苗在建成过程中受到阻碍, 这在一定程度上限制了野扁桃种群的自然更新, 使野扁桃种群更新和发展比较困难. ...
1
2006
... 种群年龄结构特征同时受到生物学特性和环境因素共同影响, 且种群年龄结构特征能体现种群动态变化及未来发展趋势(Svensson & Jeglum, 2001 ; 李俊清, 2006 ).通过对新疆野扁桃种群的年龄结构进行研究, 可以揭示种群生存现状并提出更新策略.不同灌木种群的年龄结构及其发展趋势是各不相同的, 贾喆亭等(2021 )研究发现阿拉善高原珍珠猪毛菜(Salsola passerina )种群结构以稳定型为主, 其次是增长型, 成年个体占比高, 种群整体趋于稳定.李清河等(2009 )发现半日花(Helianthemum songaricum )种群的个体数量在不同坡位存在下降趋势, 中龄级个体多, 幼龄级和老龄级个体少.本研究发现, 野扁桃种群不同龄级个体数呈现为由中龄级向幼龄级与老龄级两侧递减的趋势, 中龄级稳定生长, 占据优势, 而幼龄级缺乏, 可以看出野扁桃种群总体趋于衰退型种群结构特征.当种群处在第VIII龄级, 即达到老龄阶段之后种群数量发生锐减, 原因可能是, 野扁桃作为一种灌木, 在到达老龄阶段时, 其根部萌生的幼苗与老龄株争夺养分, 加之老龄个体的生理能力及对环境的适应性都降低, 故野扁桃种群老龄个体数量减少.在未来的发展中, 野扁桃种群虽然能暂时适应当地的生存环境, 但由于幼龄级个体储备不足, 无法维持中、老龄个体数量, 因此种群整体的长期稳定难以维持.这与牛皮杜鹃(Rhododendron aureum )及水曲柳(Fraxinus mandshurica )的种群年龄结构特征(易雪梅等, 2015 ; 金慧等, 2017 )相似.野扁桃的种群结构是受其自然生境及繁殖方式等因素共同影响的, 裕民野巴旦杏保护区内冬季长时间被深厚积雪覆盖, 受严寒气候影响, 部分老龄株死亡; 野外实地调查发现野扁桃主要以萌蘖繁殖为主, 虽然结实量大, 但是生境内土壤贫瘠、有大量的碎石岩块, 自然下落的种子很难进入土层中萌发, 并且野果林内放牧情况严重, 灌丛下萌发的幼苗及散落的种子很容易遭到牲畜的啃食, 故幼苗在建成过程中受到阻碍, 这在一定程度上限制了野扁桃种群的自然更新, 使野扁桃种群更新和发展比较困难. ...
荒漠珍稀灌木半日花种群的年龄结构与生命表分析
1
2009
... 种群年龄结构特征同时受到生物学特性和环境因素共同影响, 且种群年龄结构特征能体现种群动态变化及未来发展趋势(Svensson & Jeglum, 2001 ; 李俊清, 2006 ).通过对新疆野扁桃种群的年龄结构进行研究, 可以揭示种群生存现状并提出更新策略.不同灌木种群的年龄结构及其发展趋势是各不相同的, 贾喆亭等(2021 )研究发现阿拉善高原珍珠猪毛菜(Salsola passerina )种群结构以稳定型为主, 其次是增长型, 成年个体占比高, 种群整体趋于稳定.李清河等(2009 )发现半日花(Helianthemum songaricum )种群的个体数量在不同坡位存在下降趋势, 中龄级个体多, 幼龄级和老龄级个体少.本研究发现, 野扁桃种群不同龄级个体数呈现为由中龄级向幼龄级与老龄级两侧递减的趋势, 中龄级稳定生长, 占据优势, 而幼龄级缺乏, 可以看出野扁桃种群总体趋于衰退型种群结构特征.当种群处在第VIII龄级, 即达到老龄阶段之后种群数量发生锐减, 原因可能是, 野扁桃作为一种灌木, 在到达老龄阶段时, 其根部萌生的幼苗与老龄株争夺养分, 加之老龄个体的生理能力及对环境的适应性都降低, 故野扁桃种群老龄个体数量减少.在未来的发展中, 野扁桃种群虽然能暂时适应当地的生存环境, 但由于幼龄级个体储备不足, 无法维持中、老龄个体数量, 因此种群整体的长期稳定难以维持.这与牛皮杜鹃(Rhododendron aureum )及水曲柳(Fraxinus mandshurica )的种群年龄结构特征(易雪梅等, 2015 ; 金慧等, 2017 )相似.野扁桃的种群结构是受其自然生境及繁殖方式等因素共同影响的, 裕民野巴旦杏保护区内冬季长时间被深厚积雪覆盖, 受严寒气候影响, 部分老龄株死亡; 野外实地调查发现野扁桃主要以萌蘖繁殖为主, 虽然结实量大, 但是生境内土壤贫瘠、有大量的碎石岩块, 自然下落的种子很难进入土层中萌发, 并且野果林内放牧情况严重, 灌丛下萌发的幼苗及散落的种子很容易遭到牲畜的啃食, 故幼苗在建成过程中受到阻碍, 这在一定程度上限制了野扁桃种群的自然更新, 使野扁桃种群更新和发展比较困难. ...
荒漠珍稀灌木半日花种群的年龄结构与生命表分析
1
2009
... 种群年龄结构特征同时受到生物学特性和环境因素共同影响, 且种群年龄结构特征能体现种群动态变化及未来发展趋势(Svensson & Jeglum, 2001 ; 李俊清, 2006 ).通过对新疆野扁桃种群的年龄结构进行研究, 可以揭示种群生存现状并提出更新策略.不同灌木种群的年龄结构及其发展趋势是各不相同的, 贾喆亭等(2021 )研究发现阿拉善高原珍珠猪毛菜(Salsola passerina )种群结构以稳定型为主, 其次是增长型, 成年个体占比高, 种群整体趋于稳定.李清河等(2009 )发现半日花(Helianthemum songaricum )种群的个体数量在不同坡位存在下降趋势, 中龄级个体多, 幼龄级和老龄级个体少.本研究发现, 野扁桃种群不同龄级个体数呈现为由中龄级向幼龄级与老龄级两侧递减的趋势, 中龄级稳定生长, 占据优势, 而幼龄级缺乏, 可以看出野扁桃种群总体趋于衰退型种群结构特征.当种群处在第VIII龄级, 即达到老龄阶段之后种群数量发生锐减, 原因可能是, 野扁桃作为一种灌木, 在到达老龄阶段时, 其根部萌生的幼苗与老龄株争夺养分, 加之老龄个体的生理能力及对环境的适应性都降低, 故野扁桃种群老龄个体数量减少.在未来的发展中, 野扁桃种群虽然能暂时适应当地的生存环境, 但由于幼龄级个体储备不足, 无法维持中、老龄个体数量, 因此种群整体的长期稳定难以维持.这与牛皮杜鹃(Rhododendron aureum )及水曲柳(Fraxinus mandshurica )的种群年龄结构特征(易雪梅等, 2015 ; 金慧等, 2017 )相似.野扁桃的种群结构是受其自然生境及繁殖方式等因素共同影响的, 裕民野巴旦杏保护区内冬季长时间被深厚积雪覆盖, 受严寒气候影响, 部分老龄株死亡; 野外实地调查发现野扁桃主要以萌蘖繁殖为主, 虽然结实量大, 但是生境内土壤贫瘠、有大量的碎石岩块, 自然下落的种子很难进入土层中萌发, 并且野果林内放牧情况严重, 灌丛下萌发的幼苗及散落的种子很容易遭到牲畜的啃食, 故幼苗在建成过程中受到阻碍, 这在一定程度上限制了野扁桃种群的自然更新, 使野扁桃种群更新和发展比较困难. ...
稀有濒危植物长柱红山茶种群特征及数量动态研究
1
2016
... 存活曲线的走势反映了生存率随龄级的变化, 种群动态的基本趋势可以通过种群的存活曲线进行直接分析.对于野生果树而言, 其存活曲线的表现形式并不是单一的.田润炜等(2016 )发现塔城地区野苹果种群呈Deevey-I型, 野苹果老龄个体在较差的自然环境中加快死亡, 而幼龄个体补充不足, 所以整体衰退.杨小平等(2020 )发现野生樱桃李(Prunus cerasifera )种群生存曲线属于Deevey-II型, 由于野生樱桃李从幼苗期过渡到中龄株的时期存在严重的天然更新障碍, 所以幼苗大量死亡.本研究中, 新疆野扁桃种群存活曲线显示其基本属于Deevey-II型, 结合总体数量动态变化指数Vpi > 0, 可知种群为增长型.新疆野扁桃种群“早期-中期-后期”表现为“减少-稳定-衰退”的特征, 与姜在民等(2018 )对濒危植物羽叶丁香(Syringa pinnatifolia )种群结构与动态特征的研究结果相同.新疆野扁桃种群在第II-V龄级的死亡率较低且稳定, 可能是因为随着植株个体的发育, 对空间和资源的需求逐渐增大, 种内和种间竞争作用增强, 但在第I以及第V龄级以后出现较高的死亡率, 在第VIII龄级的死亡率最高, 说明老龄株个体发展遇到了瓶颈, 这可能是种群生理衰老导致的, 由此可以看出野扁桃种群在未来会走向衰退, 这也是小范围、片段化生境下野生植物种群必然经历的发展过程(蔡新斌和吴俊侠, 2016 ; 刘海燕等, 2016 ).通过引入生存分析的4个函数可以明显地看出野扁桃种群的结构和动态变化, 其中生存率曲线和累计死亡率曲线的变化幅度大, 在到第V龄级阶段时, 种群生存率和累计死亡率几乎相等, 种群达到平衡状态, 此后种群逐渐向衰退方向发展, 生存空间、资源环境有限以及个体衰老导致种群进入生理衰老期, 呈现衰退的趋势, 这与存活曲线反映的结果一致.综合以上分析, 野扁桃种群中幼龄个体的缺乏会导致种群逐渐走向濒危, 因此保护幼龄个体的生态环境以及增强种群自身繁育和扩散能力是阻止野扁桃种群衰退和进行种群恢复的关键. ...
稀有濒危植物长柱红山茶种群特征及数量动态研究
1
2016
... 存活曲线的走势反映了生存率随龄级的变化, 种群动态的基本趋势可以通过种群的存活曲线进行直接分析.对于野生果树而言, 其存活曲线的表现形式并不是单一的.田润炜等(2016 )发现塔城地区野苹果种群呈Deevey-I型, 野苹果老龄个体在较差的自然环境中加快死亡, 而幼龄个体补充不足, 所以整体衰退.杨小平等(2020 )发现野生樱桃李(Prunus cerasifera )种群生存曲线属于Deevey-II型, 由于野生樱桃李从幼苗期过渡到中龄株的时期存在严重的天然更新障碍, 所以幼苗大量死亡.本研究中, 新疆野扁桃种群存活曲线显示其基本属于Deevey-II型, 结合总体数量动态变化指数Vpi > 0, 可知种群为增长型.新疆野扁桃种群“早期-中期-后期”表现为“减少-稳定-衰退”的特征, 与姜在民等(2018 )对濒危植物羽叶丁香(Syringa pinnatifolia )种群结构与动态特征的研究结果相同.新疆野扁桃种群在第II-V龄级的死亡率较低且稳定, 可能是因为随着植株个体的发育, 对空间和资源的需求逐渐增大, 种内和种间竞争作用增强, 但在第I以及第V龄级以后出现较高的死亡率, 在第VIII龄级的死亡率最高, 说明老龄株个体发展遇到了瓶颈, 这可能是种群生理衰老导致的, 由此可以看出野扁桃种群在未来会走向衰退, 这也是小范围、片段化生境下野生植物种群必然经历的发展过程(蔡新斌和吴俊侠, 2016 ; 刘海燕等, 2016 ).通过引入生存分析的4个函数可以明显地看出野扁桃种群的结构和动态变化, 其中生存率曲线和累计死亡率曲线的变化幅度大, 在到第V龄级阶段时, 种群生存率和累计死亡率几乎相等, 种群达到平衡状态, 此后种群逐渐向衰退方向发展, 生存空间、资源环境有限以及个体衰老导致种群进入生理衰老期, 呈现衰退的趋势, 这与存活曲线反映的结果一致.综合以上分析, 野扁桃种群中幼龄个体的缺乏会导致种群逐渐走向濒危, 因此保护幼龄个体的生态环境以及增强种群自身繁育和扩散能力是阻止野扁桃种群衰退和进行种群恢复的关键. ...
1
2000
... 对照《天山野果林资源: 伊犁野果林综合研究》(林培钧和崔乃然, 2000 ), 核对样地出现的植物物种学名及拉丁名, 本研究所有统计分析在Excel 2007和SPSS 21中进行, 用Origin 2019软件绘图. ...
1
2000
... 对照《天山野果林资源: 伊犁野果林综合研究》(林培钧和崔乃然, 2000 ), 核对样地出现的植物物种学名及拉丁名, 本研究所有统计分析在Excel 2007和SPSS 21中进行, 用Origin 2019软件绘图. ...
祁连山区演替先锋物种西藏沙棘的种群结构及动态对海拔梯度的响应
1
2022
... 植物种群的年龄结构可以反映种群内不同年龄阶段个体的比例(李博, 2000 ).通过对植物种群在不同生境、不同时期的个体数量变化的研究, 可以反映植物种群生物学特性受环境的影响(卢晶等, 2022 ).分析植物种群年龄结构状态, 探索种群动态变化, 可以对目前种群的生存状态和种群与生境间的影响关系有所了解(袁宏波等, 2011 ), 且对于了解种群干扰情况和预测种群未来发展具有重要意义(何斌等, 2020 ; 张金峰等, 2022 ).对于植物种群年龄结构及动态分析已有不少研究报道: 濒危植物鹅掌楸(Liriodendron chinense )天然种群中中小龄级个体数量相对较多, 未来一段时间内将发展良好, 但幼龄与老龄个体严重缺乏(罗西等, 2021 ).江西南方红豆杉(Taxus wallichiana var. mairei )天然种群空间结构整体呈聚集分布, 幼苗、幼树极少, 自然更新能力差, 种群结构均呈两头小、中间大的纺锤型, 种群呈衰退趋势(喻龙华等, 2021 ).因此, 可以通过对种群年龄结构的研究, 深入探讨种群更新过程, 以及结构与动态的内在联系.新疆天山野果林准噶尔山楂(Crataegus songarica )种群现阶段的生长呈增长趋势, 但种群的长期稳定维持存在困难, 建议适当补充幼苗、幼株, 减少对成年树的砍伐破坏, 从而为种群的保育和恢复提供保障(杨蕾等, 2018 ).在研究新疆野苹果(Malus sieversii )种群年龄结构及其动态特征中, 新疆野苹果受虫害、旅游、放牧干扰影响, 时间序列分析预测野苹果种群老龄个体数量增加, 中、幼龄个体缺乏补充, 呈现不同程度衰退, 应采取人工促萌或补植幼苗, 调控林地密度, 进而加强野苹果种质资源保护(马闯等, 2018 ).结合以上研究可知, 对野果林优势种种群年龄结构的调查结合野果林受人类活动及放牧干扰的现状可以为野果林的生态恢复提供理论依据. ...
祁连山区演替先锋物种西藏沙棘的种群结构及动态对海拔梯度的响应
1
2022
... 植物种群的年龄结构可以反映种群内不同年龄阶段个体的比例(李博, 2000 ).通过对植物种群在不同生境、不同时期的个体数量变化的研究, 可以反映植物种群生物学特性受环境的影响(卢晶等, 2022 ).分析植物种群年龄结构状态, 探索种群动态变化, 可以对目前种群的生存状态和种群与生境间的影响关系有所了解(袁宏波等, 2011 ), 且对于了解种群干扰情况和预测种群未来发展具有重要意义(何斌等, 2020 ; 张金峰等, 2022 ).对于植物种群年龄结构及动态分析已有不少研究报道: 濒危植物鹅掌楸(Liriodendron chinense )天然种群中中小龄级个体数量相对较多, 未来一段时间内将发展良好, 但幼龄与老龄个体严重缺乏(罗西等, 2021 ).江西南方红豆杉(Taxus wallichiana var. mairei )天然种群空间结构整体呈聚集分布, 幼苗、幼树极少, 自然更新能力差, 种群结构均呈两头小、中间大的纺锤型, 种群呈衰退趋势(喻龙华等, 2021 ).因此, 可以通过对种群年龄结构的研究, 深入探讨种群更新过程, 以及结构与动态的内在联系.新疆天山野果林准噶尔山楂(Crataegus songarica )种群现阶段的生长呈增长趋势, 但种群的长期稳定维持存在困难, 建议适当补充幼苗、幼株, 减少对成年树的砍伐破坏, 从而为种群的保育和恢复提供保障(杨蕾等, 2018 ).在研究新疆野苹果(Malus sieversii )种群年龄结构及其动态特征中, 新疆野苹果受虫害、旅游、放牧干扰影响, 时间序列分析预测野苹果种群老龄个体数量增加, 中、幼龄个体缺乏补充, 呈现不同程度衰退, 应采取人工促萌或补植幼苗, 调控林地密度, 进而加强野苹果种质资源保护(马闯等, 2018 ).结合以上研究可知, 对野果林优势种种群年龄结构的调查结合野果林受人类活动及放牧干扰的现状可以为野果林的生态恢复提供理论依据. ...
《国家重点保护野生植物名录》调整的必要性、原则和程序
1
2021
... 野扁桃(Amygdalus ledebouriana )也称矮扁桃, 属蔷薇科李亚科桃属扁桃亚属植物, 是一古老的物种(朱京琳, 1983 ), 已被列入国家重点保护野生植物名录(鲁兆莉等, 2021 ).野扁桃不仅抗干旱还耐严寒.其果实有较高营养价值, 同时在药用方面也有研究价值(李疆等, 2006 ), 野扁桃还可以用作果树嫁接的优良矮化砧木材料, 也是选育扁桃抗性杂交新品种的最佳材料(张敏等, 1995 ).新疆的野扁桃在原生境下受到人类活动及放牧的影响, 种群数量和质量逐年下降, 蓄积量锐减, 其种群的天然更新存在问题, 因此深入了解其天然更新规律, 对该物种的保护和野果林生态恢复至关重要. ...
《国家重点保护野生植物名录》调整的必要性、原则和程序
1
2021
... 野扁桃(Amygdalus ledebouriana )也称矮扁桃, 属蔷薇科李亚科桃属扁桃亚属植物, 是一古老的物种(朱京琳, 1983 ), 已被列入国家重点保护野生植物名录(鲁兆莉等, 2021 ).野扁桃不仅抗干旱还耐严寒.其果实有较高营养价值, 同时在药用方面也有研究价值(李疆等, 2006 ), 野扁桃还可以用作果树嫁接的优良矮化砧木材料, 也是选育扁桃抗性杂交新品种的最佳材料(张敏等, 1995 ).新疆的野扁桃在原生境下受到人类活动及放牧的影响, 种群数量和质量逐年下降, 蓄积量锐减, 其种群的天然更新存在问题, 因此深入了解其天然更新规律, 对该物种的保护和野果林生态恢复至关重要. ...
濒危植物鹅掌楸的天然种群结构特征
1
2021
... 植物种群的年龄结构可以反映种群内不同年龄阶段个体的比例(李博, 2000 ).通过对植物种群在不同生境、不同时期的个体数量变化的研究, 可以反映植物种群生物学特性受环境的影响(卢晶等, 2022 ).分析植物种群年龄结构状态, 探索种群动态变化, 可以对目前种群的生存状态和种群与生境间的影响关系有所了解(袁宏波等, 2011 ), 且对于了解种群干扰情况和预测种群未来发展具有重要意义(何斌等, 2020 ; 张金峰等, 2022 ).对于植物种群年龄结构及动态分析已有不少研究报道: 濒危植物鹅掌楸(Liriodendron chinense )天然种群中中小龄级个体数量相对较多, 未来一段时间内将发展良好, 但幼龄与老龄个体严重缺乏(罗西等, 2021 ).江西南方红豆杉(Taxus wallichiana var. mairei )天然种群空间结构整体呈聚集分布, 幼苗、幼树极少, 自然更新能力差, 种群结构均呈两头小、中间大的纺锤型, 种群呈衰退趋势(喻龙华等, 2021 ).因此, 可以通过对种群年龄结构的研究, 深入探讨种群更新过程, 以及结构与动态的内在联系.新疆天山野果林准噶尔山楂(Crataegus songarica )种群现阶段的生长呈增长趋势, 但种群的长期稳定维持存在困难, 建议适当补充幼苗、幼株, 减少对成年树的砍伐破坏, 从而为种群的保育和恢复提供保障(杨蕾等, 2018 ).在研究新疆野苹果(Malus sieversii )种群年龄结构及其动态特征中, 新疆野苹果受虫害、旅游、放牧干扰影响, 时间序列分析预测野苹果种群老龄个体数量增加, 中、幼龄个体缺乏补充, 呈现不同程度衰退, 应采取人工促萌或补植幼苗, 调控林地密度, 进而加强野苹果种质资源保护(马闯等, 2018 ).结合以上研究可知, 对野果林优势种种群年龄结构的调查结合野果林受人类活动及放牧干扰的现状可以为野果林的生态恢复提供理论依据. ...
濒危植物鹅掌楸的天然种群结构特征
1
2021
... 植物种群的年龄结构可以反映种群内不同年龄阶段个体的比例(李博, 2000 ).通过对植物种群在不同生境、不同时期的个体数量变化的研究, 可以反映植物种群生物学特性受环境的影响(卢晶等, 2022 ).分析植物种群年龄结构状态, 探索种群动态变化, 可以对目前种群的生存状态和种群与生境间的影响关系有所了解(袁宏波等, 2011 ), 且对于了解种群干扰情况和预测种群未来发展具有重要意义(何斌等, 2020 ; 张金峰等, 2022 ).对于植物种群年龄结构及动态分析已有不少研究报道: 濒危植物鹅掌楸(Liriodendron chinense )天然种群中中小龄级个体数量相对较多, 未来一段时间内将发展良好, 但幼龄与老龄个体严重缺乏(罗西等, 2021 ).江西南方红豆杉(Taxus wallichiana var. mairei )天然种群空间结构整体呈聚集分布, 幼苗、幼树极少, 自然更新能力差, 种群结构均呈两头小、中间大的纺锤型, 种群呈衰退趋势(喻龙华等, 2021 ).因此, 可以通过对种群年龄结构的研究, 深入探讨种群更新过程, 以及结构与动态的内在联系.新疆天山野果林准噶尔山楂(Crataegus songarica )种群现阶段的生长呈增长趋势, 但种群的长期稳定维持存在困难, 建议适当补充幼苗、幼株, 减少对成年树的砍伐破坏, 从而为种群的保育和恢复提供保障(杨蕾等, 2018 ).在研究新疆野苹果(Malus sieversii )种群年龄结构及其动态特征中, 新疆野苹果受虫害、旅游、放牧干扰影响, 时间序列分析预测野苹果种群老龄个体数量增加, 中、幼龄个体缺乏补充, 呈现不同程度衰退, 应采取人工促萌或补植幼苗, 调控林地密度, 进而加强野苹果种质资源保护(马闯等, 2018 ).结合以上研究可知, 对野果林优势种种群年龄结构的调查结合野果林受人类活动及放牧干扰的现状可以为野果林的生态恢复提供理论依据. ...
新疆野苹果(Malus sieversii )种群年龄结构及其动态特征
1
2018
... 植物种群的年龄结构可以反映种群内不同年龄阶段个体的比例(李博, 2000 ).通过对植物种群在不同生境、不同时期的个体数量变化的研究, 可以反映植物种群生物学特性受环境的影响(卢晶等, 2022 ).分析植物种群年龄结构状态, 探索种群动态变化, 可以对目前种群的生存状态和种群与生境间的影响关系有所了解(袁宏波等, 2011 ), 且对于了解种群干扰情况和预测种群未来发展具有重要意义(何斌等, 2020 ; 张金峰等, 2022 ).对于植物种群年龄结构及动态分析已有不少研究报道: 濒危植物鹅掌楸(Liriodendron chinense )天然种群中中小龄级个体数量相对较多, 未来一段时间内将发展良好, 但幼龄与老龄个体严重缺乏(罗西等, 2021 ).江西南方红豆杉(Taxus wallichiana var. mairei )天然种群空间结构整体呈聚集分布, 幼苗、幼树极少, 自然更新能力差, 种群结构均呈两头小、中间大的纺锤型, 种群呈衰退趋势(喻龙华等, 2021 ).因此, 可以通过对种群年龄结构的研究, 深入探讨种群更新过程, 以及结构与动态的内在联系.新疆天山野果林准噶尔山楂(Crataegus songarica )种群现阶段的生长呈增长趋势, 但种群的长期稳定维持存在困难, 建议适当补充幼苗、幼株, 减少对成年树的砍伐破坏, 从而为种群的保育和恢复提供保障(杨蕾等, 2018 ).在研究新疆野苹果(Malus sieversii )种群年龄结构及其动态特征中, 新疆野苹果受虫害、旅游、放牧干扰影响, 时间序列分析预测野苹果种群老龄个体数量增加, 中、幼龄个体缺乏补充, 呈现不同程度衰退, 应采取人工促萌或补植幼苗, 调控林地密度, 进而加强野苹果种质资源保护(马闯等, 2018 ).结合以上研究可知, 对野果林优势种种群年龄结构的调查结合野果林受人类活动及放牧干扰的现状可以为野果林的生态恢复提供理论依据. ...
新疆野苹果(Malus sieversii )种群年龄结构及其动态特征
1
2018
... 植物种群的年龄结构可以反映种群内不同年龄阶段个体的比例(李博, 2000 ).通过对植物种群在不同生境、不同时期的个体数量变化的研究, 可以反映植物种群生物学特性受环境的影响(卢晶等, 2022 ).分析植物种群年龄结构状态, 探索种群动态变化, 可以对目前种群的生存状态和种群与生境间的影响关系有所了解(袁宏波等, 2011 ), 且对于了解种群干扰情况和预测种群未来发展具有重要意义(何斌等, 2020 ; 张金峰等, 2022 ).对于植物种群年龄结构及动态分析已有不少研究报道: 濒危植物鹅掌楸(Liriodendron chinense )天然种群中中小龄级个体数量相对较多, 未来一段时间内将发展良好, 但幼龄与老龄个体严重缺乏(罗西等, 2021 ).江西南方红豆杉(Taxus wallichiana var. mairei )天然种群空间结构整体呈聚集分布, 幼苗、幼树极少, 自然更新能力差, 种群结构均呈两头小、中间大的纺锤型, 种群呈衰退趋势(喻龙华等, 2021 ).因此, 可以通过对种群年龄结构的研究, 深入探讨种群更新过程, 以及结构与动态的内在联系.新疆天山野果林准噶尔山楂(Crataegus songarica )种群现阶段的生长呈增长趋势, 但种群的长期稳定维持存在困难, 建议适当补充幼苗、幼株, 减少对成年树的砍伐破坏, 从而为种群的保育和恢复提供保障(杨蕾等, 2018 ).在研究新疆野苹果(Malus sieversii )种群年龄结构及其动态特征中, 新疆野苹果受虫害、旅游、放牧干扰影响, 时间序列分析预测野苹果种群老龄个体数量增加, 中、幼龄个体缺乏补充, 呈现不同程度衰退, 应采取人工促萌或补植幼苗, 调控林地密度, 进而加强野苹果种质资源保护(马闯等, 2018 ).结合以上研究可知, 对野果林优势种种群年龄结构的调查结合野果林受人类活动及放牧干扰的现状可以为野果林的生态恢复提供理论依据. ...
濒危植物新疆野扁桃的遗传多样性
1
2019
... 种群生态学对于研究种群内部各成员之间, 种群与生境的相互作用规律有重要意义.而目前有关新疆野扁桃的研究主要集中在生物信息学(王波, 2018 )、抗逆性(宋宏伟等, 2013 )、繁殖生物学(程靖, 2016 )、果实营养成分分析(朱绪春等, 2016 )、遗传多样性(马松梅等, 2019 )等方面, 但是在新疆野扁桃的种群生态学领域方面的研究, 目前尚未见报道.本研究以新疆巴尔鲁克山天然野扁桃种群为研究对象, 利用典型样方法进行种群数量统计, 结合存活曲线及静态生命表, 进行生存分析, 旨在探索野扁桃年龄结构特征及更新受限机制, 以期为新疆野扁桃种群的保护管理及恢复提供一定的理论依据及研究基础. ...
濒危植物新疆野扁桃的遗传多样性
1
2019
... 种群生态学对于研究种群内部各成员之间, 种群与生境的相互作用规律有重要意义.而目前有关新疆野扁桃的研究主要集中在生物信息学(王波, 2018 )、抗逆性(宋宏伟等, 2013 )、繁殖生物学(程靖, 2016 )、果实营养成分分析(朱绪春等, 2016 )、遗传多样性(马松梅等, 2019 )等方面, 但是在新疆野扁桃的种群生态学领域方面的研究, 目前尚未见报道.本研究以新疆巴尔鲁克山天然野扁桃种群为研究对象, 利用典型样方法进行种群数量统计, 结合存活曲线及静态生命表, 进行生存分析, 旨在探索野扁桃年龄结构特征及更新受限机制, 以期为新疆野扁桃种群的保护管理及恢复提供一定的理论依据及研究基础. ...
新疆野扁桃抗旱性生理指标的测定
1
2013
... 种群生态学对于研究种群内部各成员之间, 种群与生境的相互作用规律有重要意义.而目前有关新疆野扁桃的研究主要集中在生物信息学(王波, 2018 )、抗逆性(宋宏伟等, 2013 )、繁殖生物学(程靖, 2016 )、果实营养成分分析(朱绪春等, 2016 )、遗传多样性(马松梅等, 2019 )等方面, 但是在新疆野扁桃的种群生态学领域方面的研究, 目前尚未见报道.本研究以新疆巴尔鲁克山天然野扁桃种群为研究对象, 利用典型样方法进行种群数量统计, 结合存活曲线及静态生命表, 进行生存分析, 旨在探索野扁桃年龄结构特征及更新受限机制, 以期为新疆野扁桃种群的保护管理及恢复提供一定的理论依据及研究基础. ...
新疆野扁桃抗旱性生理指标的测定
1
2013
... 种群生态学对于研究种群内部各成员之间, 种群与生境的相互作用规律有重要意义.而目前有关新疆野扁桃的研究主要集中在生物信息学(王波, 2018 )、抗逆性(宋宏伟等, 2013 )、繁殖生物学(程靖, 2016 )、果实营养成分分析(朱绪春等, 2016 )、遗传多样性(马松梅等, 2019 )等方面, 但是在新疆野扁桃的种群生态学领域方面的研究, 目前尚未见报道.本研究以新疆巴尔鲁克山天然野扁桃种群为研究对象, 利用典型样方法进行种群数量统计, 结合存活曲线及静态生命表, 进行生存分析, 旨在探索野扁桃年龄结构特征及更新受限机制, 以期为新疆野扁桃种群的保护管理及恢复提供一定的理论依据及研究基础. ...
Structure and dynamics of an undisturbed old-growth Norway spruce forest on the rising Bothnian coastline
1
2001
... 种群年龄结构特征同时受到生物学特性和环境因素共同影响, 且种群年龄结构特征能体现种群动态变化及未来发展趋势(Svensson & Jeglum, 2001 ; 李俊清, 2006 ).通过对新疆野扁桃种群的年龄结构进行研究, 可以揭示种群生存现状并提出更新策略.不同灌木种群的年龄结构及其发展趋势是各不相同的, 贾喆亭等(2021 )研究发现阿拉善高原珍珠猪毛菜(Salsola passerina )种群结构以稳定型为主, 其次是增长型, 成年个体占比高, 种群整体趋于稳定.李清河等(2009 )发现半日花(Helianthemum songaricum )种群的个体数量在不同坡位存在下降趋势, 中龄级个体多, 幼龄级和老龄级个体少.本研究发现, 野扁桃种群不同龄级个体数呈现为由中龄级向幼龄级与老龄级两侧递减的趋势, 中龄级稳定生长, 占据优势, 而幼龄级缺乏, 可以看出野扁桃种群总体趋于衰退型种群结构特征.当种群处在第VIII龄级, 即达到老龄阶段之后种群数量发生锐减, 原因可能是, 野扁桃作为一种灌木, 在到达老龄阶段时, 其根部萌生的幼苗与老龄株争夺养分, 加之老龄个体的生理能力及对环境的适应性都降低, 故野扁桃种群老龄个体数量减少.在未来的发展中, 野扁桃种群虽然能暂时适应当地的生存环境, 但由于幼龄级个体储备不足, 无法维持中、老龄个体数量, 因此种群整体的长期稳定难以维持.这与牛皮杜鹃(Rhododendron aureum )及水曲柳(Fraxinus mandshurica )的种群年龄结构特征(易雪梅等, 2015 ; 金慧等, 2017 )相似.野扁桃的种群结构是受其自然生境及繁殖方式等因素共同影响的, 裕民野巴旦杏保护区内冬季长时间被深厚积雪覆盖, 受严寒气候影响, 部分老龄株死亡; 野外实地调查发现野扁桃主要以萌蘖繁殖为主, 虽然结实量大, 但是生境内土壤贫瘠、有大量的碎石岩块, 自然下落的种子很难进入土层中萌发, 并且野果林内放牧情况严重, 灌丛下萌发的幼苗及散落的种子很容易遭到牲畜的啃食, 故幼苗在建成过程中受到阻碍, 这在一定程度上限制了野扁桃种群的自然更新, 使野扁桃种群更新和发展比较困难. ...
新疆野苹果种群年龄结构特征与动态分析
1
2016
... 存活曲线的走势反映了生存率随龄级的变化, 种群动态的基本趋势可以通过种群的存活曲线进行直接分析.对于野生果树而言, 其存活曲线的表现形式并不是单一的.田润炜等(2016 )发现塔城地区野苹果种群呈Deevey-I型, 野苹果老龄个体在较差的自然环境中加快死亡, 而幼龄个体补充不足, 所以整体衰退.杨小平等(2020 )发现野生樱桃李(Prunus cerasifera )种群生存曲线属于Deevey-II型, 由于野生樱桃李从幼苗期过渡到中龄株的时期存在严重的天然更新障碍, 所以幼苗大量死亡.本研究中, 新疆野扁桃种群存活曲线显示其基本属于Deevey-II型, 结合总体数量动态变化指数Vpi > 0, 可知种群为增长型.新疆野扁桃种群“早期-中期-后期”表现为“减少-稳定-衰退”的特征, 与姜在民等(2018 )对濒危植物羽叶丁香(Syringa pinnatifolia )种群结构与动态特征的研究结果相同.新疆野扁桃种群在第II-V龄级的死亡率较低且稳定, 可能是因为随着植株个体的发育, 对空间和资源的需求逐渐增大, 种内和种间竞争作用增强, 但在第I以及第V龄级以后出现较高的死亡率, 在第VIII龄级的死亡率最高, 说明老龄株个体发展遇到了瓶颈, 这可能是种群生理衰老导致的, 由此可以看出野扁桃种群在未来会走向衰退, 这也是小范围、片段化生境下野生植物种群必然经历的发展过程(蔡新斌和吴俊侠, 2016 ; 刘海燕等, 2016 ).通过引入生存分析的4个函数可以明显地看出野扁桃种群的结构和动态变化, 其中生存率曲线和累计死亡率曲线的变化幅度大, 在到第V龄级阶段时, 种群生存率和累计死亡率几乎相等, 种群达到平衡状态, 此后种群逐渐向衰退方向发展, 生存空间、资源环境有限以及个体衰老导致种群进入生理衰老期, 呈现衰退的趋势, 这与存活曲线反映的结果一致.综合以上分析, 野扁桃种群中幼龄个体的缺乏会导致种群逐渐走向濒危, 因此保护幼龄个体的生态环境以及增强种群自身繁育和扩散能力是阻止野扁桃种群衰退和进行种群恢复的关键. ...
新疆野苹果种群年龄结构特征与动态分析
1
2016
... 存活曲线的走势反映了生存率随龄级的变化, 种群动态的基本趋势可以通过种群的存活曲线进行直接分析.对于野生果树而言, 其存活曲线的表现形式并不是单一的.田润炜等(2016 )发现塔城地区野苹果种群呈Deevey-I型, 野苹果老龄个体在较差的自然环境中加快死亡, 而幼龄个体补充不足, 所以整体衰退.杨小平等(2020 )发现野生樱桃李(Prunus cerasifera )种群生存曲线属于Deevey-II型, 由于野生樱桃李从幼苗期过渡到中龄株的时期存在严重的天然更新障碍, 所以幼苗大量死亡.本研究中, 新疆野扁桃种群存活曲线显示其基本属于Deevey-II型, 结合总体数量动态变化指数Vpi > 0, 可知种群为增长型.新疆野扁桃种群“早期-中期-后期”表现为“减少-稳定-衰退”的特征, 与姜在民等(2018 )对濒危植物羽叶丁香(Syringa pinnatifolia )种群结构与动态特征的研究结果相同.新疆野扁桃种群在第II-V龄级的死亡率较低且稳定, 可能是因为随着植株个体的发育, 对空间和资源的需求逐渐增大, 种内和种间竞争作用增强, 但在第I以及第V龄级以后出现较高的死亡率, 在第VIII龄级的死亡率最高, 说明老龄株个体发展遇到了瓶颈, 这可能是种群生理衰老导致的, 由此可以看出野扁桃种群在未来会走向衰退, 这也是小范围、片段化生境下野生植物种群必然经历的发展过程(蔡新斌和吴俊侠, 2016 ; 刘海燕等, 2016 ).通过引入生存分析的4个函数可以明显地看出野扁桃种群的结构和动态变化, 其中生存率曲线和累计死亡率曲线的变化幅度大, 在到第V龄级阶段时, 种群生存率和累计死亡率几乎相等, 种群达到平衡状态, 此后种群逐渐向衰退方向发展, 生存空间、资源环境有限以及个体衰老导致种群进入生理衰老期, 呈现衰退的趋势, 这与存活曲线反映的结果一致.综合以上分析, 野扁桃种群中幼龄个体的缺乏会导致种群逐渐走向濒危, 因此保护幼龄个体的生态环境以及增强种群自身繁育和扩散能力是阻止野扁桃种群衰退和进行种群恢复的关键. ...
新疆野扁桃PetCullin1 基因的克隆与生物信息学分析
1
2018
... 种群生态学对于研究种群内部各成员之间, 种群与生境的相互作用规律有重要意义.而目前有关新疆野扁桃的研究主要集中在生物信息学(王波, 2018 )、抗逆性(宋宏伟等, 2013 )、繁殖生物学(程靖, 2016 )、果实营养成分分析(朱绪春等, 2016 )、遗传多样性(马松梅等, 2019 )等方面, 但是在新疆野扁桃的种群生态学领域方面的研究, 目前尚未见报道.本研究以新疆巴尔鲁克山天然野扁桃种群为研究对象, 利用典型样方法进行种群数量统计, 结合存活曲线及静态生命表, 进行生存分析, 旨在探索野扁桃年龄结构特征及更新受限机制, 以期为新疆野扁桃种群的保护管理及恢复提供一定的理论依据及研究基础. ...
新疆野扁桃PetCullin1 基因的克隆与生物信息学分析
1
2018
... 种群生态学对于研究种群内部各成员之间, 种群与生境的相互作用规律有重要意义.而目前有关新疆野扁桃的研究主要集中在生物信息学(王波, 2018 )、抗逆性(宋宏伟等, 2013 )、繁殖生物学(程靖, 2016 )、果实营养成分分析(朱绪春等, 2016 )、遗传多样性(马松梅等, 2019 )等方面, 但是在新疆野扁桃的种群生态学领域方面的研究, 目前尚未见报道.本研究以新疆巴尔鲁克山天然野扁桃种群为研究对象, 利用典型样方法进行种群数量统计, 结合存活曲线及静态生命表, 进行生存分析, 旨在探索野扁桃年龄结构特征及更新受限机制, 以期为新疆野扁桃种群的保护管理及恢复提供一定的理论依据及研究基础. ...
森林植物立木种群静态生命表的一种编制方法
1
1992
... 本研究采用“空间代替时间”的方法(谢晋阳, 1992 ).由于野扁桃生长缓慢, 株高和冠幅的生物学特性差异不大, 加之存在放牧和牲畜啃食的情况, 故利用野扁桃基径的大小级代替年龄结构来研究其组成结构(高婷婷等, 2010 ), 将野扁桃种群划分为13个龄级, 第I龄级的基径≤0.3 cm, 第II到第XII龄级每级间隔0.3 cm, 即第II龄级的基径为0.3-0.6 cm, 0.6-0.9 cm为第III龄级, 0.9-1.2 cm为第IV龄级, 1.2-1.5 cm为第V龄级, 1.5-1.8 cm为第VI龄级, 1.8-2.1 cm为第VII龄级, 2.1-2.4 cm为第VIII龄级, 2.4-2.7 cm为第IX龄级, 2.7-3.0 cm为第X龄级, 3.0-3.3 cm为第XI龄级, 3.3-3.6 cm为第XII龄级, 第XIII龄级的基径>3.6 cm.统计各龄级野扁桃株数, 绘制野扁桃年龄结构图. ...
森林植物立木种群静态生命表的一种编制方法
1
1992
... 本研究采用“空间代替时间”的方法(谢晋阳, 1992 ).由于野扁桃生长缓慢, 株高和冠幅的生物学特性差异不大, 加之存在放牧和牲畜啃食的情况, 故利用野扁桃基径的大小级代替年龄结构来研究其组成结构(高婷婷等, 2010 ), 将野扁桃种群划分为13个龄级, 第I龄级的基径≤0.3 cm, 第II到第XII龄级每级间隔0.3 cm, 即第II龄级的基径为0.3-0.6 cm, 0.6-0.9 cm为第III龄级, 0.9-1.2 cm为第IV龄级, 1.2-1.5 cm为第V龄级, 1.5-1.8 cm为第VI龄级, 1.8-2.1 cm为第VII龄级, 2.1-2.4 cm为第VIII龄级, 2.4-2.7 cm为第IX龄级, 2.7-3.0 cm为第X龄级, 3.0-3.3 cm为第XI龄级, 3.3-3.6 cm为第XII龄级, 第XIII龄级的基径>3.6 cm.统计各龄级野扁桃株数, 绘制野扁桃年龄结构图. ...
生存分析理论及其在研究生命表中的应用
1
1991
... 其中Si 为存活率, hi 为龄级宽度(杨凤翔等, 1991 ), 通过绘制野扁桃种群生存分析函数曲线图, 进一步分析野扁桃的种群动态. ...
生存分析理论及其在研究生命表中的应用
1
1991
... 其中Si 为存活率, hi 为龄级宽度(杨凤翔等, 1991 ), 通过绘制野扁桃种群生存分析函数曲线图, 进一步分析野扁桃的种群动态. ...
新疆天山野果林准噶尔山楂种群结构与动态分析
1
2018
... 植物种群的年龄结构可以反映种群内不同年龄阶段个体的比例(李博, 2000 ).通过对植物种群在不同生境、不同时期的个体数量变化的研究, 可以反映植物种群生物学特性受环境的影响(卢晶等, 2022 ).分析植物种群年龄结构状态, 探索种群动态变化, 可以对目前种群的生存状态和种群与生境间的影响关系有所了解(袁宏波等, 2011 ), 且对于了解种群干扰情况和预测种群未来发展具有重要意义(何斌等, 2020 ; 张金峰等, 2022 ).对于植物种群年龄结构及动态分析已有不少研究报道: 濒危植物鹅掌楸(Liriodendron chinense )天然种群中中小龄级个体数量相对较多, 未来一段时间内将发展良好, 但幼龄与老龄个体严重缺乏(罗西等, 2021 ).江西南方红豆杉(Taxus wallichiana var. mairei )天然种群空间结构整体呈聚集分布, 幼苗、幼树极少, 自然更新能力差, 种群结构均呈两头小、中间大的纺锤型, 种群呈衰退趋势(喻龙华等, 2021 ).因此, 可以通过对种群年龄结构的研究, 深入探讨种群更新过程, 以及结构与动态的内在联系.新疆天山野果林准噶尔山楂(Crataegus songarica )种群现阶段的生长呈增长趋势, 但种群的长期稳定维持存在困难, 建议适当补充幼苗、幼株, 减少对成年树的砍伐破坏, 从而为种群的保育和恢复提供保障(杨蕾等, 2018 ).在研究新疆野苹果(Malus sieversii )种群年龄结构及其动态特征中, 新疆野苹果受虫害、旅游、放牧干扰影响, 时间序列分析预测野苹果种群老龄个体数量增加, 中、幼龄个体缺乏补充, 呈现不同程度衰退, 应采取人工促萌或补植幼苗, 调控林地密度, 进而加强野苹果种质资源保护(马闯等, 2018 ).结合以上研究可知, 对野果林优势种种群年龄结构的调查结合野果林受人类活动及放牧干扰的现状可以为野果林的生态恢复提供理论依据. ...
新疆天山野果林准噶尔山楂种群结构与动态分析
1
2018
... 植物种群的年龄结构可以反映种群内不同年龄阶段个体的比例(李博, 2000 ).通过对植物种群在不同生境、不同时期的个体数量变化的研究, 可以反映植物种群生物学特性受环境的影响(卢晶等, 2022 ).分析植物种群年龄结构状态, 探索种群动态变化, 可以对目前种群的生存状态和种群与生境间的影响关系有所了解(袁宏波等, 2011 ), 且对于了解种群干扰情况和预测种群未来发展具有重要意义(何斌等, 2020 ; 张金峰等, 2022 ).对于植物种群年龄结构及动态分析已有不少研究报道: 濒危植物鹅掌楸(Liriodendron chinense )天然种群中中小龄级个体数量相对较多, 未来一段时间内将发展良好, 但幼龄与老龄个体严重缺乏(罗西等, 2021 ).江西南方红豆杉(Taxus wallichiana var. mairei )天然种群空间结构整体呈聚集分布, 幼苗、幼树极少, 自然更新能力差, 种群结构均呈两头小、中间大的纺锤型, 种群呈衰退趋势(喻龙华等, 2021 ).因此, 可以通过对种群年龄结构的研究, 深入探讨种群更新过程, 以及结构与动态的内在联系.新疆天山野果林准噶尔山楂(Crataegus songarica )种群现阶段的生长呈增长趋势, 但种群的长期稳定维持存在困难, 建议适当补充幼苗、幼株, 减少对成年树的砍伐破坏, 从而为种群的保育和恢复提供保障(杨蕾等, 2018 ).在研究新疆野苹果(Malus sieversii )种群年龄结构及其动态特征中, 新疆野苹果受虫害、旅游、放牧干扰影响, 时间序列分析预测野苹果种群老龄个体数量增加, 中、幼龄个体缺乏补充, 呈现不同程度衰退, 应采取人工促萌或补植幼苗, 调控林地密度, 进而加强野苹果种质资源保护(马闯等, 2018 ).结合以上研究可知, 对野果林优势种种群年龄结构的调查结合野果林受人类活动及放牧干扰的现状可以为野果林的生态恢复提供理论依据. ...
野生樱桃李的种群统计与生存分析
2
2020
... 静态生命表可以反映多年生植物种群的出生率和死亡率的变化规律, 因此可用静态生命表对多年生植物的种群动态进行分析(杨小平等, 2020 ).通过野扁桃种群各龄级的株数可编制野扁桃种群的静态生命表(程子敏等, 2018 ).计算公式如下: ...
... 存活曲线的走势反映了生存率随龄级的变化, 种群动态的基本趋势可以通过种群的存活曲线进行直接分析.对于野生果树而言, 其存活曲线的表现形式并不是单一的.田润炜等(2016 )发现塔城地区野苹果种群呈Deevey-I型, 野苹果老龄个体在较差的自然环境中加快死亡, 而幼龄个体补充不足, 所以整体衰退.杨小平等(2020 )发现野生樱桃李(Prunus cerasifera )种群生存曲线属于Deevey-II型, 由于野生樱桃李从幼苗期过渡到中龄株的时期存在严重的天然更新障碍, 所以幼苗大量死亡.本研究中, 新疆野扁桃种群存活曲线显示其基本属于Deevey-II型, 结合总体数量动态变化指数Vpi > 0, 可知种群为增长型.新疆野扁桃种群“早期-中期-后期”表现为“减少-稳定-衰退”的特征, 与姜在民等(2018 )对濒危植物羽叶丁香(Syringa pinnatifolia )种群结构与动态特征的研究结果相同.新疆野扁桃种群在第II-V龄级的死亡率较低且稳定, 可能是因为随着植株个体的发育, 对空间和资源的需求逐渐增大, 种内和种间竞争作用增强, 但在第I以及第V龄级以后出现较高的死亡率, 在第VIII龄级的死亡率最高, 说明老龄株个体发展遇到了瓶颈, 这可能是种群生理衰老导致的, 由此可以看出野扁桃种群在未来会走向衰退, 这也是小范围、片段化生境下野生植物种群必然经历的发展过程(蔡新斌和吴俊侠, 2016 ; 刘海燕等, 2016 ).通过引入生存分析的4个函数可以明显地看出野扁桃种群的结构和动态变化, 其中生存率曲线和累计死亡率曲线的变化幅度大, 在到第V龄级阶段时, 种群生存率和累计死亡率几乎相等, 种群达到平衡状态, 此后种群逐渐向衰退方向发展, 生存空间、资源环境有限以及个体衰老导致种群进入生理衰老期, 呈现衰退的趋势, 这与存活曲线反映的结果一致.综合以上分析, 野扁桃种群中幼龄个体的缺乏会导致种群逐渐走向濒危, 因此保护幼龄个体的生态环境以及增强种群自身繁育和扩散能力是阻止野扁桃种群衰退和进行种群恢复的关键. ...
野生樱桃李的种群统计与生存分析
2
2020
... 静态生命表可以反映多年生植物种群的出生率和死亡率的变化规律, 因此可用静态生命表对多年生植物的种群动态进行分析(杨小平等, 2020 ).通过野扁桃种群各龄级的株数可编制野扁桃种群的静态生命表(程子敏等, 2018 ).计算公式如下: ...
... 存活曲线的走势反映了生存率随龄级的变化, 种群动态的基本趋势可以通过种群的存活曲线进行直接分析.对于野生果树而言, 其存活曲线的表现形式并不是单一的.田润炜等(2016 )发现塔城地区野苹果种群呈Deevey-I型, 野苹果老龄个体在较差的自然环境中加快死亡, 而幼龄个体补充不足, 所以整体衰退.杨小平等(2020 )发现野生樱桃李(Prunus cerasifera )种群生存曲线属于Deevey-II型, 由于野生樱桃李从幼苗期过渡到中龄株的时期存在严重的天然更新障碍, 所以幼苗大量死亡.本研究中, 新疆野扁桃种群存活曲线显示其基本属于Deevey-II型, 结合总体数量动态变化指数Vpi > 0, 可知种群为增长型.新疆野扁桃种群“早期-中期-后期”表现为“减少-稳定-衰退”的特征, 与姜在民等(2018 )对濒危植物羽叶丁香(Syringa pinnatifolia )种群结构与动态特征的研究结果相同.新疆野扁桃种群在第II-V龄级的死亡率较低且稳定, 可能是因为随着植株个体的发育, 对空间和资源的需求逐渐增大, 种内和种间竞争作用增强, 但在第I以及第V龄级以后出现较高的死亡率, 在第VIII龄级的死亡率最高, 说明老龄株个体发展遇到了瓶颈, 这可能是种群生理衰老导致的, 由此可以看出野扁桃种群在未来会走向衰退, 这也是小范围、片段化生境下野生植物种群必然经历的发展过程(蔡新斌和吴俊侠, 2016 ; 刘海燕等, 2016 ).通过引入生存分析的4个函数可以明显地看出野扁桃种群的结构和动态变化, 其中生存率曲线和累计死亡率曲线的变化幅度大, 在到第V龄级阶段时, 种群生存率和累计死亡率几乎相等, 种群达到平衡状态, 此后种群逐渐向衰退方向发展, 生存空间、资源环境有限以及个体衰老导致种群进入生理衰老期, 呈现衰退的趋势, 这与存活曲线反映的结果一致.综合以上分析, 野扁桃种群中幼龄个体的缺乏会导致种群逐渐走向濒危, 因此保护幼龄个体的生态环境以及增强种群自身繁育和扩散能力是阻止野扁桃种群衰退和进行种群恢复的关键. ...
长白山水曲柳种群动态
1
2015
... 种群年龄结构特征同时受到生物学特性和环境因素共同影响, 且种群年龄结构特征能体现种群动态变化及未来发展趋势(Svensson & Jeglum, 2001 ; 李俊清, 2006 ).通过对新疆野扁桃种群的年龄结构进行研究, 可以揭示种群生存现状并提出更新策略.不同灌木种群的年龄结构及其发展趋势是各不相同的, 贾喆亭等(2021 )研究发现阿拉善高原珍珠猪毛菜(Salsola passerina )种群结构以稳定型为主, 其次是增长型, 成年个体占比高, 种群整体趋于稳定.李清河等(2009 )发现半日花(Helianthemum songaricum )种群的个体数量在不同坡位存在下降趋势, 中龄级个体多, 幼龄级和老龄级个体少.本研究发现, 野扁桃种群不同龄级个体数呈现为由中龄级向幼龄级与老龄级两侧递减的趋势, 中龄级稳定生长, 占据优势, 而幼龄级缺乏, 可以看出野扁桃种群总体趋于衰退型种群结构特征.当种群处在第VIII龄级, 即达到老龄阶段之后种群数量发生锐减, 原因可能是, 野扁桃作为一种灌木, 在到达老龄阶段时, 其根部萌生的幼苗与老龄株争夺养分, 加之老龄个体的生理能力及对环境的适应性都降低, 故野扁桃种群老龄个体数量减少.在未来的发展中, 野扁桃种群虽然能暂时适应当地的生存环境, 但由于幼龄级个体储备不足, 无法维持中、老龄个体数量, 因此种群整体的长期稳定难以维持.这与牛皮杜鹃(Rhododendron aureum )及水曲柳(Fraxinus mandshurica )的种群年龄结构特征(易雪梅等, 2015 ; 金慧等, 2017 )相似.野扁桃的种群结构是受其自然生境及繁殖方式等因素共同影响的, 裕民野巴旦杏保护区内冬季长时间被深厚积雪覆盖, 受严寒气候影响, 部分老龄株死亡; 野外实地调查发现野扁桃主要以萌蘖繁殖为主, 虽然结实量大, 但是生境内土壤贫瘠、有大量的碎石岩块, 自然下落的种子很难进入土层中萌发, 并且野果林内放牧情况严重, 灌丛下萌发的幼苗及散落的种子很容易遭到牲畜的啃食, 故幼苗在建成过程中受到阻碍, 这在一定程度上限制了野扁桃种群的自然更新, 使野扁桃种群更新和发展比较困难. ...
长白山水曲柳种群动态
1
2015
... 种群年龄结构特征同时受到生物学特性和环境因素共同影响, 且种群年龄结构特征能体现种群动态变化及未来发展趋势(Svensson & Jeglum, 2001 ; 李俊清, 2006 ).通过对新疆野扁桃种群的年龄结构进行研究, 可以揭示种群生存现状并提出更新策略.不同灌木种群的年龄结构及其发展趋势是各不相同的, 贾喆亭等(2021 )研究发现阿拉善高原珍珠猪毛菜(Salsola passerina )种群结构以稳定型为主, 其次是增长型, 成年个体占比高, 种群整体趋于稳定.李清河等(2009 )发现半日花(Helianthemum songaricum )种群的个体数量在不同坡位存在下降趋势, 中龄级个体多, 幼龄级和老龄级个体少.本研究发现, 野扁桃种群不同龄级个体数呈现为由中龄级向幼龄级与老龄级两侧递减的趋势, 中龄级稳定生长, 占据优势, 而幼龄级缺乏, 可以看出野扁桃种群总体趋于衰退型种群结构特征.当种群处在第VIII龄级, 即达到老龄阶段之后种群数量发生锐减, 原因可能是, 野扁桃作为一种灌木, 在到达老龄阶段时, 其根部萌生的幼苗与老龄株争夺养分, 加之老龄个体的生理能力及对环境的适应性都降低, 故野扁桃种群老龄个体数量减少.在未来的发展中, 野扁桃种群虽然能暂时适应当地的生存环境, 但由于幼龄级个体储备不足, 无法维持中、老龄个体数量, 因此种群整体的长期稳定难以维持.这与牛皮杜鹃(Rhododendron aureum )及水曲柳(Fraxinus mandshurica )的种群年龄结构特征(易雪梅等, 2015 ; 金慧等, 2017 )相似.野扁桃的种群结构是受其自然生境及繁殖方式等因素共同影响的, 裕民野巴旦杏保护区内冬季长时间被深厚积雪覆盖, 受严寒气候影响, 部分老龄株死亡; 野外实地调查发现野扁桃主要以萌蘖繁殖为主, 虽然结实量大, 但是生境内土壤贫瘠、有大量的碎石岩块, 自然下落的种子很难进入土层中萌发, 并且野果林内放牧情况严重, 灌丛下萌发的幼苗及散落的种子很容易遭到牲畜的啃食, 故幼苗在建成过程中受到阻碍, 这在一定程度上限制了野扁桃种群的自然更新, 使野扁桃种群更新和发展比较困难. ...
江西天然南方红豆杉群落及种群结构特征
1
2021
... 植物种群的年龄结构可以反映种群内不同年龄阶段个体的比例(李博, 2000 ).通过对植物种群在不同生境、不同时期的个体数量变化的研究, 可以反映植物种群生物学特性受环境的影响(卢晶等, 2022 ).分析植物种群年龄结构状态, 探索种群动态变化, 可以对目前种群的生存状态和种群与生境间的影响关系有所了解(袁宏波等, 2011 ), 且对于了解种群干扰情况和预测种群未来发展具有重要意义(何斌等, 2020 ; 张金峰等, 2022 ).对于植物种群年龄结构及动态分析已有不少研究报道: 濒危植物鹅掌楸(Liriodendron chinense )天然种群中中小龄级个体数量相对较多, 未来一段时间内将发展良好, 但幼龄与老龄个体严重缺乏(罗西等, 2021 ).江西南方红豆杉(Taxus wallichiana var. mairei )天然种群空间结构整体呈聚集分布, 幼苗、幼树极少, 自然更新能力差, 种群结构均呈两头小、中间大的纺锤型, 种群呈衰退趋势(喻龙华等, 2021 ).因此, 可以通过对种群年龄结构的研究, 深入探讨种群更新过程, 以及结构与动态的内在联系.新疆天山野果林准噶尔山楂(Crataegus songarica )种群现阶段的生长呈增长趋势, 但种群的长期稳定维持存在困难, 建议适当补充幼苗、幼株, 减少对成年树的砍伐破坏, 从而为种群的保育和恢复提供保障(杨蕾等, 2018 ).在研究新疆野苹果(Malus sieversii )种群年龄结构及其动态特征中, 新疆野苹果受虫害、旅游、放牧干扰影响, 时间序列分析预测野苹果种群老龄个体数量增加, 中、幼龄个体缺乏补充, 呈现不同程度衰退, 应采取人工促萌或补植幼苗, 调控林地密度, 进而加强野苹果种质资源保护(马闯等, 2018 ).结合以上研究可知, 对野果林优势种种群年龄结构的调查结合野果林受人类活动及放牧干扰的现状可以为野果林的生态恢复提供理论依据. ...
江西天然南方红豆杉群落及种群结构特征
1
2021
... 植物种群的年龄结构可以反映种群内不同年龄阶段个体的比例(李博, 2000 ).通过对植物种群在不同生境、不同时期的个体数量变化的研究, 可以反映植物种群生物学特性受环境的影响(卢晶等, 2022 ).分析植物种群年龄结构状态, 探索种群动态变化, 可以对目前种群的生存状态和种群与生境间的影响关系有所了解(袁宏波等, 2011 ), 且对于了解种群干扰情况和预测种群未来发展具有重要意义(何斌等, 2020 ; 张金峰等, 2022 ).对于植物种群年龄结构及动态分析已有不少研究报道: 濒危植物鹅掌楸(Liriodendron chinense )天然种群中中小龄级个体数量相对较多, 未来一段时间内将发展良好, 但幼龄与老龄个体严重缺乏(罗西等, 2021 ).江西南方红豆杉(Taxus wallichiana var. mairei )天然种群空间结构整体呈聚集分布, 幼苗、幼树极少, 自然更新能力差, 种群结构均呈两头小、中间大的纺锤型, 种群呈衰退趋势(喻龙华等, 2021 ).因此, 可以通过对种群年龄结构的研究, 深入探讨种群更新过程, 以及结构与动态的内在联系.新疆天山野果林准噶尔山楂(Crataegus songarica )种群现阶段的生长呈增长趋势, 但种群的长期稳定维持存在困难, 建议适当补充幼苗、幼株, 减少对成年树的砍伐破坏, 从而为种群的保育和恢复提供保障(杨蕾等, 2018 ).在研究新疆野苹果(Malus sieversii )种群年龄结构及其动态特征中, 新疆野苹果受虫害、旅游、放牧干扰影响, 时间序列分析预测野苹果种群老龄个体数量增加, 中、幼龄个体缺乏补充, 呈现不同程度衰退, 应采取人工促萌或补植幼苗, 调控林地密度, 进而加强野苹果种质资源保护(马闯等, 2018 ).结合以上研究可知, 对野果林优势种种群年龄结构的调查结合野果林受人类活动及放牧干扰的现状可以为野果林的生态恢复提供理论依据. ...
库姆塔格沙漠典型植物种群年龄结构特征
1
2011
... 植物种群的年龄结构可以反映种群内不同年龄阶段个体的比例(李博, 2000 ).通过对植物种群在不同生境、不同时期的个体数量变化的研究, 可以反映植物种群生物学特性受环境的影响(卢晶等, 2022 ).分析植物种群年龄结构状态, 探索种群动态变化, 可以对目前种群的生存状态和种群与生境间的影响关系有所了解(袁宏波等, 2011 ), 且对于了解种群干扰情况和预测种群未来发展具有重要意义(何斌等, 2020 ; 张金峰等, 2022 ).对于植物种群年龄结构及动态分析已有不少研究报道: 濒危植物鹅掌楸(Liriodendron chinense )天然种群中中小龄级个体数量相对较多, 未来一段时间内将发展良好, 但幼龄与老龄个体严重缺乏(罗西等, 2021 ).江西南方红豆杉(Taxus wallichiana var. mairei )天然种群空间结构整体呈聚集分布, 幼苗、幼树极少, 自然更新能力差, 种群结构均呈两头小、中间大的纺锤型, 种群呈衰退趋势(喻龙华等, 2021 ).因此, 可以通过对种群年龄结构的研究, 深入探讨种群更新过程, 以及结构与动态的内在联系.新疆天山野果林准噶尔山楂(Crataegus songarica )种群现阶段的生长呈增长趋势, 但种群的长期稳定维持存在困难, 建议适当补充幼苗、幼株, 减少对成年树的砍伐破坏, 从而为种群的保育和恢复提供保障(杨蕾等, 2018 ).在研究新疆野苹果(Malus sieversii )种群年龄结构及其动态特征中, 新疆野苹果受虫害、旅游、放牧干扰影响, 时间序列分析预测野苹果种群老龄个体数量增加, 中、幼龄个体缺乏补充, 呈现不同程度衰退, 应采取人工促萌或补植幼苗, 调控林地密度, 进而加强野苹果种质资源保护(马闯等, 2018 ).结合以上研究可知, 对野果林优势种种群年龄结构的调查结合野果林受人类活动及放牧干扰的现状可以为野果林的生态恢复提供理论依据. ...
库姆塔格沙漠典型植物种群年龄结构特征
1
2011
... 植物种群的年龄结构可以反映种群内不同年龄阶段个体的比例(李博, 2000 ).通过对植物种群在不同生境、不同时期的个体数量变化的研究, 可以反映植物种群生物学特性受环境的影响(卢晶等, 2022 ).分析植物种群年龄结构状态, 探索种群动态变化, 可以对目前种群的生存状态和种群与生境间的影响关系有所了解(袁宏波等, 2011 ), 且对于了解种群干扰情况和预测种群未来发展具有重要意义(何斌等, 2020 ; 张金峰等, 2022 ).对于植物种群年龄结构及动态分析已有不少研究报道: 濒危植物鹅掌楸(Liriodendron chinense )天然种群中中小龄级个体数量相对较多, 未来一段时间内将发展良好, 但幼龄与老龄个体严重缺乏(罗西等, 2021 ).江西南方红豆杉(Taxus wallichiana var. mairei )天然种群空间结构整体呈聚集分布, 幼苗、幼树极少, 自然更新能力差, 种群结构均呈两头小、中间大的纺锤型, 种群呈衰退趋势(喻龙华等, 2021 ).因此, 可以通过对种群年龄结构的研究, 深入探讨种群更新过程, 以及结构与动态的内在联系.新疆天山野果林准噶尔山楂(Crataegus songarica )种群现阶段的生长呈增长趋势, 但种群的长期稳定维持存在困难, 建议适当补充幼苗、幼株, 减少对成年树的砍伐破坏, 从而为种群的保育和恢复提供保障(杨蕾等, 2018 ).在研究新疆野苹果(Malus sieversii )种群年龄结构及其动态特征中, 新疆野苹果受虫害、旅游、放牧干扰影响, 时间序列分析预测野苹果种群老龄个体数量增加, 中、幼龄个体缺乏补充, 呈现不同程度衰退, 应采取人工促萌或补植幼苗, 调控林地密度, 进而加强野苹果种质资源保护(马闯等, 2018 ).结合以上研究可知, 对野果林优势种种群年龄结构的调查结合野果林受人类活动及放牧干扰的现状可以为野果林的生态恢复提供理论依据. ...
新疆野扁桃繁殖生物学特性及种质资源遗传多样性研究
1
2008
... 巴尔鲁克山(45.42°-46.98° N, 82.26°-83.38° E)位于塔城地区裕民县和托里县境内, 年平均气温为4.9-6.9 ℃, 年降水量241 mm, 该地区土壤类型为栗钙土, 有机质含量为5% (曾斌, 2008 ), 野扁桃的分布海拔主要在800-1 300 m, 研究区植被的乔木层主要有野扁桃、野苹果、准噶尔山楂、野杏(Armeniaca vulgaris )等; 灌木层主要有天山樱桃(Cerasus tianshanica )、栒子(Cotoneaster hissaricus )、金丝桃叶绣线菊(Spiraea hypericifolia )等; 草本植物主要有菥蓂(Thlaspi arvense )、苦马豆(Sphaerophysa salsula )、塔城棘豆(Oxytropis schrenkii )等. ...
新疆野扁桃繁殖生物学特性及种质资源遗传多样性研究
1
2008
... 巴尔鲁克山(45.42°-46.98° N, 82.26°-83.38° E)位于塔城地区裕民县和托里县境内, 年平均气温为4.9-6.9 ℃, 年降水量241 mm, 该地区土壤类型为栗钙土, 有机质含量为5% (曾斌, 2008 ), 野扁桃的分布海拔主要在800-1 300 m, 研究区植被的乔木层主要有野扁桃、野苹果、准噶尔山楂、野杏(Armeniaca vulgaris )等; 灌木层主要有天山樱桃(Cerasus tianshanica )、栒子(Cotoneaster hissaricus )、金丝桃叶绣线菊(Spiraea hypericifolia )等; 草本植物主要有菥蓂(Thlaspi arvense )、苦马豆(Sphaerophysa salsula )、塔城棘豆(Oxytropis schrenkii )等. ...
长白山阔叶红松林红松种群年龄结构与数量动态特征
1
2022
... 植物种群的年龄结构可以反映种群内不同年龄阶段个体的比例(李博, 2000 ).通过对植物种群在不同生境、不同时期的个体数量变化的研究, 可以反映植物种群生物学特性受环境的影响(卢晶等, 2022 ).分析植物种群年龄结构状态, 探索种群动态变化, 可以对目前种群的生存状态和种群与生境间的影响关系有所了解(袁宏波等, 2011 ), 且对于了解种群干扰情况和预测种群未来发展具有重要意义(何斌等, 2020 ; 张金峰等, 2022 ).对于植物种群年龄结构及动态分析已有不少研究报道: 濒危植物鹅掌楸(Liriodendron chinense )天然种群中中小龄级个体数量相对较多, 未来一段时间内将发展良好, 但幼龄与老龄个体严重缺乏(罗西等, 2021 ).江西南方红豆杉(Taxus wallichiana var. mairei )天然种群空间结构整体呈聚集分布, 幼苗、幼树极少, 自然更新能力差, 种群结构均呈两头小、中间大的纺锤型, 种群呈衰退趋势(喻龙华等, 2021 ).因此, 可以通过对种群年龄结构的研究, 深入探讨种群更新过程, 以及结构与动态的内在联系.新疆天山野果林准噶尔山楂(Crataegus songarica )种群现阶段的生长呈增长趋势, 但种群的长期稳定维持存在困难, 建议适当补充幼苗、幼株, 减少对成年树的砍伐破坏, 从而为种群的保育和恢复提供保障(杨蕾等, 2018 ).在研究新疆野苹果(Malus sieversii )种群年龄结构及其动态特征中, 新疆野苹果受虫害、旅游、放牧干扰影响, 时间序列分析预测野苹果种群老龄个体数量增加, 中、幼龄个体缺乏补充, 呈现不同程度衰退, 应采取人工促萌或补植幼苗, 调控林地密度, 进而加强野苹果种质资源保护(马闯等, 2018 ).结合以上研究可知, 对野果林优势种种群年龄结构的调查结合野果林受人类活动及放牧干扰的现状可以为野果林的生态恢复提供理论依据. ...
长白山阔叶红松林红松种群年龄结构与数量动态特征
1
2022
... 植物种群的年龄结构可以反映种群内不同年龄阶段个体的比例(李博, 2000 ).通过对植物种群在不同生境、不同时期的个体数量变化的研究, 可以反映植物种群生物学特性受环境的影响(卢晶等, 2022 ).分析植物种群年龄结构状态, 探索种群动态变化, 可以对目前种群的生存状态和种群与生境间的影响关系有所了解(袁宏波等, 2011 ), 且对于了解种群干扰情况和预测种群未来发展具有重要意义(何斌等, 2020 ; 张金峰等, 2022 ).对于植物种群年龄结构及动态分析已有不少研究报道: 濒危植物鹅掌楸(Liriodendron chinense )天然种群中中小龄级个体数量相对较多, 未来一段时间内将发展良好, 但幼龄与老龄个体严重缺乏(罗西等, 2021 ).江西南方红豆杉(Taxus wallichiana var. mairei )天然种群空间结构整体呈聚集分布, 幼苗、幼树极少, 自然更新能力差, 种群结构均呈两头小、中间大的纺锤型, 种群呈衰退趋势(喻龙华等, 2021 ).因此, 可以通过对种群年龄结构的研究, 深入探讨种群更新过程, 以及结构与动态的内在联系.新疆天山野果林准噶尔山楂(Crataegus songarica )种群现阶段的生长呈增长趋势, 但种群的长期稳定维持存在困难, 建议适当补充幼苗、幼株, 减少对成年树的砍伐破坏, 从而为种群的保育和恢复提供保障(杨蕾等, 2018 ).在研究新疆野苹果(Malus sieversii )种群年龄结构及其动态特征中, 新疆野苹果受虫害、旅游、放牧干扰影响, 时间序列分析预测野苹果种群老龄个体数量增加, 中、幼龄个体缺乏补充, 呈现不同程度衰退, 应采取人工促萌或补植幼苗, 调控林地密度, 进而加强野苹果种质资源保护(马闯等, 2018 ).结合以上研究可知, 对野果林优势种种群年龄结构的调查结合野果林受人类活动及放牧干扰的现状可以为野果林的生态恢复提供理论依据. ...
新疆巴旦杏氨基酸组分分析
1
1995
... 野扁桃(Amygdalus ledebouriana )也称矮扁桃, 属蔷薇科李亚科桃属扁桃亚属植物, 是一古老的物种(朱京琳, 1983 ), 已被列入国家重点保护野生植物名录(鲁兆莉等, 2021 ).野扁桃不仅抗干旱还耐严寒.其果实有较高营养价值, 同时在药用方面也有研究价值(李疆等, 2006 ), 野扁桃还可以用作果树嫁接的优良矮化砧木材料, 也是选育扁桃抗性杂交新品种的最佳材料(张敏等, 1995 ).新疆的野扁桃在原生境下受到人类活动及放牧的影响, 种群数量和质量逐年下降, 蓄积量锐减, 其种群的天然更新存在问题, 因此深入了解其天然更新规律, 对该物种的保护和野果林生态恢复至关重要. ...
新疆巴旦杏氨基酸组分分析
1
1995
... 野扁桃(Amygdalus ledebouriana )也称矮扁桃, 属蔷薇科李亚科桃属扁桃亚属植物, 是一古老的物种(朱京琳, 1983 ), 已被列入国家重点保护野生植物名录(鲁兆莉等, 2021 ).野扁桃不仅抗干旱还耐严寒.其果实有较高营养价值, 同时在药用方面也有研究价值(李疆等, 2006 ), 野扁桃还可以用作果树嫁接的优良矮化砧木材料, 也是选育扁桃抗性杂交新品种的最佳材料(张敏等, 1995 ).新疆的野扁桃在原生境下受到人类活动及放牧的影响, 种群数量和质量逐年下降, 蓄积量锐减, 其种群的天然更新存在问题, 因此深入了解其天然更新规律, 对该物种的保护和野果林生态恢复至关重要. ...
1
1983
... 野扁桃(Amygdalus ledebouriana )也称矮扁桃, 属蔷薇科李亚科桃属扁桃亚属植物, 是一古老的物种(朱京琳, 1983 ), 已被列入国家重点保护野生植物名录(鲁兆莉等, 2021 ).野扁桃不仅抗干旱还耐严寒.其果实有较高营养价值, 同时在药用方面也有研究价值(李疆等, 2006 ), 野扁桃还可以用作果树嫁接的优良矮化砧木材料, 也是选育扁桃抗性杂交新品种的最佳材料(张敏等, 1995 ).新疆的野扁桃在原生境下受到人类活动及放牧的影响, 种群数量和质量逐年下降, 蓄积量锐减, 其种群的天然更新存在问题, 因此深入了解其天然更新规律, 对该物种的保护和野果林生态恢复至关重要. ...
1
1983
... 野扁桃(Amygdalus ledebouriana )也称矮扁桃, 属蔷薇科李亚科桃属扁桃亚属植物, 是一古老的物种(朱京琳, 1983 ), 已被列入国家重点保护野生植物名录(鲁兆莉等, 2021 ).野扁桃不仅抗干旱还耐严寒.其果实有较高营养价值, 同时在药用方面也有研究价值(李疆等, 2006 ), 野扁桃还可以用作果树嫁接的优良矮化砧木材料, 也是选育扁桃抗性杂交新品种的最佳材料(张敏等, 1995 ).新疆的野扁桃在原生境下受到人类活动及放牧的影响, 种群数量和质量逐年下降, 蓄积量锐减, 其种群的天然更新存在问题, 因此深入了解其天然更新规律, 对该物种的保护和野果林生态恢复至关重要. ...
3种野生扁桃油脂的理化性质及脂肪酸组成研究
1
2016
... 种群生态学对于研究种群内部各成员之间, 种群与生境的相互作用规律有重要意义.而目前有关新疆野扁桃的研究主要集中在生物信息学(王波, 2018 )、抗逆性(宋宏伟等, 2013 )、繁殖生物学(程靖, 2016 )、果实营养成分分析(朱绪春等, 2016 )、遗传多样性(马松梅等, 2019 )等方面, 但是在新疆野扁桃的种群生态学领域方面的研究, 目前尚未见报道.本研究以新疆巴尔鲁克山天然野扁桃种群为研究对象, 利用典型样方法进行种群数量统计, 结合存活曲线及静态生命表, 进行生存分析, 旨在探索野扁桃年龄结构特征及更新受限机制, 以期为新疆野扁桃种群的保护管理及恢复提供一定的理论依据及研究基础. ...
3种野生扁桃油脂的理化性质及脂肪酸组成研究
1
2016
... 种群生态学对于研究种群内部各成员之间, 种群与生境的相互作用规律有重要意义.而目前有关新疆野扁桃的研究主要集中在生物信息学(王波, 2018 )、抗逆性(宋宏伟等, 2013 )、繁殖生物学(程靖, 2016 )、果实营养成分分析(朱绪春等, 2016 )、遗传多样性(马松梅等, 2019 )等方面, 但是在新疆野扁桃的种群生态学领域方面的研究, 目前尚未见报道.本研究以新疆巴尔鲁克山天然野扁桃种群为研究对象, 利用典型样方法进行种群数量统计, 结合存活曲线及静态生命表, 进行生存分析, 旨在探索野扁桃年龄结构特征及更新受限机制, 以期为新疆野扁桃种群的保护管理及恢复提供一定的理论依据及研究基础. ...










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}