
植物生态学报 ›› 2016, Vol. 40 ›› Issue (7): 702-710.DOI: 10.17521/cjpe.2016.0064
所属专题: 植物功能性状
金鹰, 王传宽*( )
)
收稿日期:2016-02-14
接受日期:2016-05-09
出版日期:2016-07-10
发布日期:2016-07-07
通讯作者:
王传宽
基金资助:
Ying JIN, Chuan-Kuan WANG*()
Received:2016-02-14
Accepted:2016-05-09
Online:2016-07-10
Published:2016-07-07
Contact:
Chuan-Kuan WANG
摘要:
不同材性树种的解剖、叶脉分布等结构性状差异会影响树木的水分运输效率和水分利用策略, 进而限制树木的生存、生长和分布。然而, 材性对叶导水率、水力脆弱性及其潜在的权衡关系的影响尚不清楚。该研究选择东北温带森林中不同材性的9种树种(散孔材: 山杨(Populus davidiana)、紫椴(Tilia amurensis)、白桦(Betula platyphylla); 环孔材: 蒙古栎(Quercus mongolica)、水曲柳(Fraxinus mandshurica)、胡桃楸(Juglans mandshurica); 无孔材: 红皮云杉(Picea koraiensis)、樟子松(Pinus sylvestris var. mongolica)、红松(Pinus koraiensis), 测量其基于叶面积和叶质量的叶导水率(Karea和Kmass)、水力脆弱性(P50)、膨压丧失点水势(TLP)及叶结构性状, 以比较不同材性树种叶水力性状的差异, 并探索叶水力效率与安全的权衡关系。结果表明: 3种材性树种的Karea、Kmass和P50均差异显著(p < 0.05)。无孔材树种的Karea和Kmass最低, 而散孔材和环孔材树种差异不显著; 环孔材树种P50最高, 而散孔材和无孔材树种差异不显著。Karea和Kmass均与P50显著负相关(p < 0.05), 但散孔材、环孔材和无孔材树种的相关关系分别呈线性、幂函数和指数函数关系。这表明叶水力效率与安全之间存在一定的权衡关系, 但该关系受树木材性的影响。Kmass与TLP显著负相关(p < 0.01), 其中散孔材和环孔材树种呈线性负相关, 无孔材树种呈负指数函数关系; P50随TLP的增加而增加, 这表明树木在面临水分胁迫时, 其质外体和共质体抗旱阻力共同协调保护叶片活细胞, 防止其水分状况到达临界阈值。Kmass与叶干物质含量、叶密度、比叶重均显著负相关, 而P50与之显著正相关(p < 0.01, P50与比叶重的关系除外), 表明树木叶水力特性的变化受相同叶结构特性驱动, 树木增加对水力失调的容忍需要在叶水力系统构建上增加碳投资。
金鹰, 王传宽. 九种不同材性的温带树种叶水力性状及其权衡关系. 植物生态学报, 2016, 40(7): 702-710. DOI: 10.17521/cjpe.2016.0064
Ying JIN, Chuan-Kuan WANG. Leaf hydraulic traits and their trade-offs for nine Chinese temperate tree species with different wood properties. Chinese Journal of Plant Ecology, 2016, 40(7): 702-710. DOI: 10.17521/cjpe.2016.0064
| 材性(代码) Wood property (code) | 树种(代码) Species (code) | 叶习性 Leaf habit | 生境 Habitat | 胸径 DBH (cm) |
|---|---|---|---|---|
| 散孔材 | 白桦 Betula platyphylla (BH) | 落叶阔叶 Deciduous-broadleaved | 山坡中部 Mid slope | 24.46 ± 1.05 |
| Diffuse-porous (DP) | 山杨 Populus davidiana (SY) | 落叶阔叶 Deciduous-broadleaved | 山坡上部 Upper slope | 31.15 ± 0.55 |
| 紫椴 Tilia amurensis (ZD) | 落叶阔叶 Deciduous-broadleaved | 山坡上部 Upper slope | 25.75 ± 1.30 | |
| 环孔材 | 水曲柳 Fraxinus mandshurica (SQL) | 落叶阔叶 Deciduous-broadleaved | 山坡下部 Toe slope | 34.30 ± 0.45 |
| Ring-porous (RP) | 蒙古栎 Quercus mongolica (MGL) | 落叶阔叶 Deciduous-broadleaved | 山坡上部 Upper slope | 27.86 ± 1.19 |
| 胡桃楸 Juglans mandshurica (HTQ) | 落叶阔叶 Deciduous-broadleaved | 山谷 Valley bottom | 34.60 ± 1.38 | |
| 无孔材 | 红松 Pinus koraiensis (HS) | 常绿针叶 Evergreen-coniferous | 山坡中部 Mid slope | 28.17 ± 0.91 |
| Non-porous (NP) | 云杉 Picea koraiensis (YS) | 常绿针叶 Evergreen-coniferous | 山谷 Valley bottom | 30.05 ± 0.65 |
| 樟子松 Pinus sylvestris var. mongolica (ZZS) | 常绿针叶 Evergreen-coniferous | 山坡中部 Mid slope | 26.32 ± 1.15 |
表1 9种不同材性的温带树种样木的基本特性(平均值±标准误差, n = 4)
Table 1 Basic characteristics of the sampled trees for the nine temperate tree species with different wood properties (mean ± SE, n = 4)
| 材性(代码) Wood property (code) | 树种(代码) Species (code) | 叶习性 Leaf habit | 生境 Habitat | 胸径 DBH (cm) |
|---|---|---|---|---|
| 散孔材 | 白桦 Betula platyphylla (BH) | 落叶阔叶 Deciduous-broadleaved | 山坡中部 Mid slope | 24.46 ± 1.05 |
| Diffuse-porous (DP) | 山杨 Populus davidiana (SY) | 落叶阔叶 Deciduous-broadleaved | 山坡上部 Upper slope | 31.15 ± 0.55 |
| 紫椴 Tilia amurensis (ZD) | 落叶阔叶 Deciduous-broadleaved | 山坡上部 Upper slope | 25.75 ± 1.30 | |
| 环孔材 | 水曲柳 Fraxinus mandshurica (SQL) | 落叶阔叶 Deciduous-broadleaved | 山坡下部 Toe slope | 34.30 ± 0.45 |
| Ring-porous (RP) | 蒙古栎 Quercus mongolica (MGL) | 落叶阔叶 Deciduous-broadleaved | 山坡上部 Upper slope | 27.86 ± 1.19 |
| 胡桃楸 Juglans mandshurica (HTQ) | 落叶阔叶 Deciduous-broadleaved | 山谷 Valley bottom | 34.60 ± 1.38 | |
| 无孔材 | 红松 Pinus koraiensis (HS) | 常绿针叶 Evergreen-coniferous | 山坡中部 Mid slope | 28.17 ± 0.91 |
| Non-porous (NP) | 云杉 Picea koraiensis (YS) | 常绿针叶 Evergreen-coniferous | 山谷 Valley bottom | 30.05 ± 0.65 |
| 樟子松 Pinus sylvestris var. mongolica (ZZS) | 常绿针叶 Evergreen-coniferous | 山坡中部 Mid slope | 26.32 ± 1.15 |
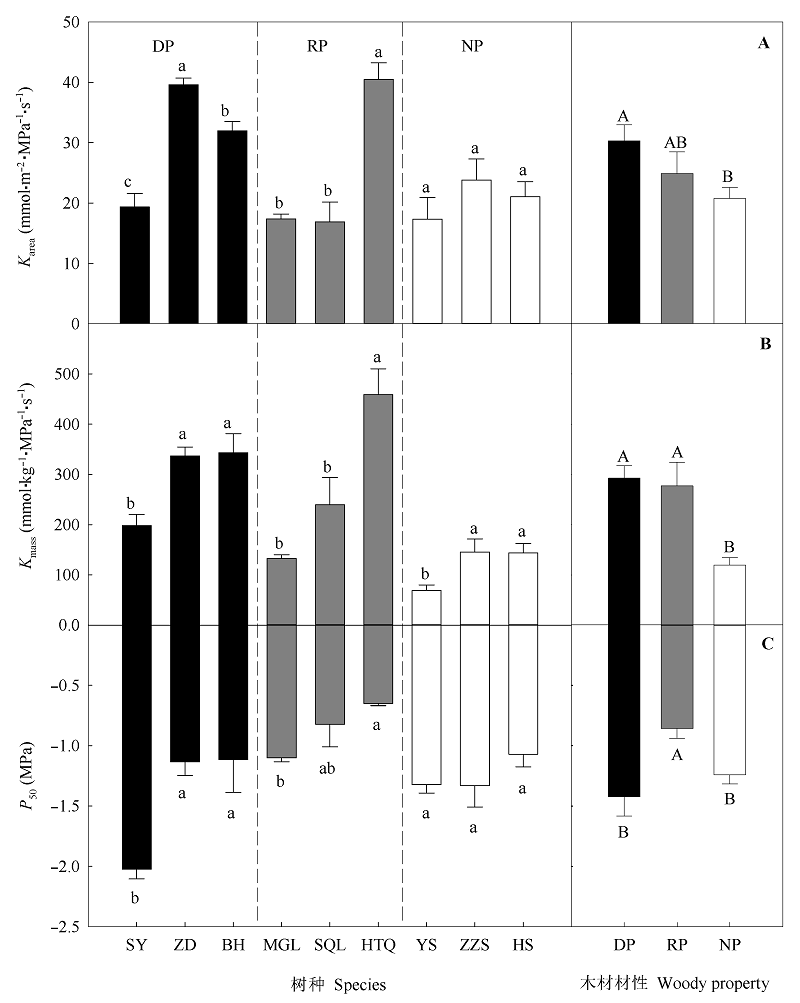
图1 不同材性树种叶水力性状的比较(平均值±标准误差)。不同大、小写字母分别代表不同材性之间和同一材性不同树种之间差异显著(p < 0.05)。Karea和Kmass分别表示基于叶面积和叶质量的叶导水率; P50表示叶水力脆弱性。材性和树种代码同表1。
Fig. 1 Comparisons of leaf hydraulic traits among the tree species with different wood properties (mean ± SE). Different uppercase and lowercase letters above columns indicate significant differences among different wood properties and among different tree species with the same wood property, respectively (p < 0.05). Karea and Kmass, leaf hydraulic conductance per leaf area and dry mass, respectively; P50, leaf hydraulic vulnerability. See Table 1 for the codes of tree species and wood properties.
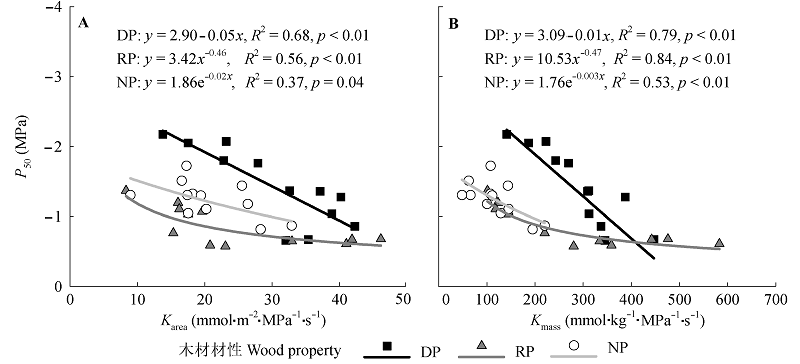
图2 3种材性树种叶水力效率与水力脆弱性(P50)的关系。Karea和Kmass分别表示基于叶面积和叶质量的叶导水率。材性代码同表1。
Fig. 2 Relationships between leaf hydraulic efficiency and hydraulic vulnerability (P50) of the trees with wood properties. Karea and Kmass, leaf hydraulic conductance per leaf area and dry mass, respectively. The codes of wood properties are listed in Table 1.
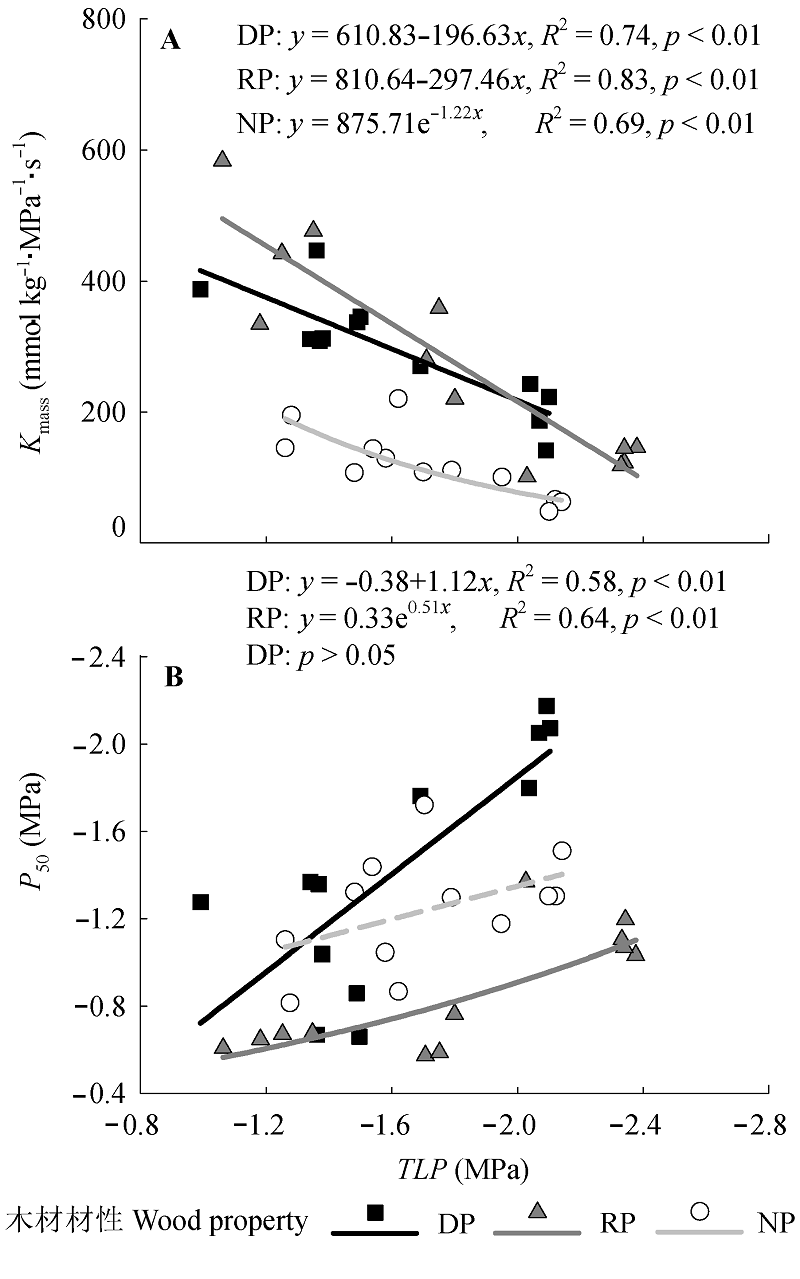
图3 3种材性树种膨压丧失点水势(TLP)与(A)基于叶质量的叶导水率(Kmass)和(B)叶水力脆弱性(P50)的关系。虚线表示关系不显著(p > 0.05)。材性代码同表1。
Fig. 3 Relationships between leaf water potential at turgor loss point (TLP) and (A) leaf-mass-based hydraulic conductance (Kmass) or (B) leaf hydraulic vulnerability (P50) of the trees with wood properties. The dash line denotes non-significant (p > 0.05). The codes of wood properties are listed in Table 1.
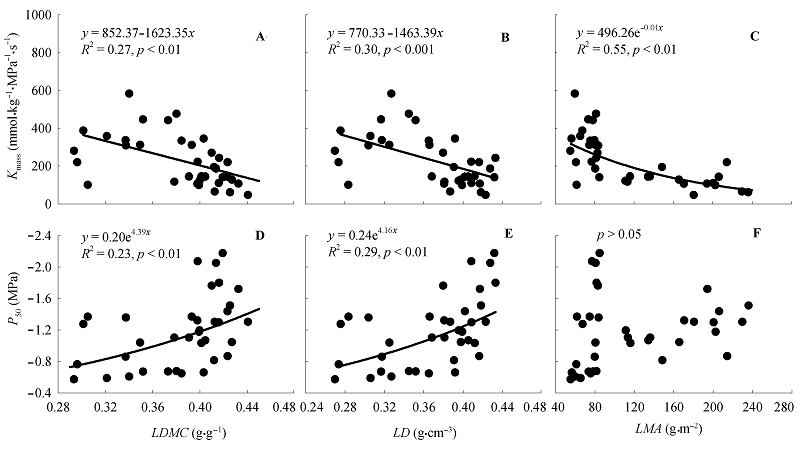
图4 基于所有树种的叶水力性状与叶结构性状的关系。Kmass, 基于叶质量的叶导水率; LD, 叶密度; LDMC, 叶干物质含量; LMA, 比叶重; P50, 叶水力脆弱性。
Fig. 4 The relationships between leaf hydraulics and structural traits of all tree species. Kmass, leaf hydraulic conductance per dry mass; LD, leaf density; LDMC, leaf dry mass content; LMA, leaf mass per unit area; P50, leaf hydraulic vulnerability.
| 1 | Blackman CJ, Brodribb TJ, Jordan GJ (2010). Leaf hydraulic vulnerability is related to conduit dimensions and drought resistance across a diverse range of woody angiosperms.New Phytologist, 188, 1113-1123. |
| 2 | Brodribb TJ, Holbrook NM (2003). Stomatal closure during leaf dehydration, correlation with other leaf physiological traits.Plant Physiology, 132, 2166-2173. |
| 3 | Brodribb TJ, Holbrook NM, Zwieniecki MA, Palma B (2005). Leaf hydraulic capacity in ferns, conifers and angiosperms: Impacts on photosynthetic maxima.New Phytologist, 165, 839-846. |
| 4 | Bucci SJ, Scholz FG, Campanello PI, Montti L, Jimenez- Castillo M, Rockwell FA, Manna LL, Guerra P, Bernal PL, Troncoso O, Enricci J, Holbrook MN, Goldstein G (2012). Hydraulic differences along the water transport system of South American Nothofagus species: Do leaves protect the stem functionality?Tree Physiology, 32, 880-893. |
| 5 | Bucci SJ, Scholz FG, Goldstein G, Meinzer FC, Franco AC, Campanello P, Villalobos-Vega R, Bustamante M, Miralles- Wilhelm F (2006). Nutrient availability constrains the hydraulic architecture and water relations of savannah trees.Plant, Cell & Environment, 29, 2153-2167. |
| 6 | Bucci SJ, Scholz FG, Goldstein G, Meinzer FC, Sternberg LDASL (2003). Dynamic changes in hydraulic conductivity in petioles of two savanna tree species: Factors and mechanisms contributing to the refilling of embolized vessels.Plant, Cell & Environment, 26, 1633-1645. |
| 7 | Carnicer J, Barbeta A, Sperlich D, Coll M, Penuelas J (2013). Contrasting trait syndromes in angiosperms and conifers are associated with different responses of tree growth to temperature on a large scale.Front Plant Science, 4, 409. |
| 8 | Coll M, Penuelas J, Ninyerola M, Pons X, Carnicer J (2013). Multivariate effect gradients driving forest demographic responses in the Iberian Peninsula.Forest Ecology & Management, 303, 195-209. |
| 9 | Feild TS, Brodribb TJ (2001). Stem water transport and freeze-thaw xylem embolism in conifers and angiosperms in a Tasmanian treeline heath.Oecologia, 127, 314-320. |
| 10 | Giordano R, Salleo A, Salleo S, Wanderlingh F (1978). Flow in xylem vessels and Poiseuille’s law.Canadian Journal of Botany, 56, 333-338. |
| 11 | Gomez-Aparicio L, Garcia VR, Ruiz-Benito P, Zavala MA (2011). Disentangling the relative importance of climate, size and competition on tree growth in Iberian forests: Implications for forest management under global change.Globle Change Biology, 17, 2400-2414. |
| 12 | Gong R, Gao Q (2015). Research progress in the effects of leaf hydraulic characteristics on plant physiological functions.Chinese Journal of Plant Ecology, 39, 300-308. (in Chinese with English abstract)[龚容, 高琼 (2015). 叶片结构的水力学特性对植物生理功能影响的研究进展. 植物生态学报, 39, 300-308.] |
| 13 | Hao GY, Hoffmann WA, Scholz FG, Bucci SJ, Meinzer FC, Franco AC, Cao KF, Goldstein G (2008). Stem and leaf hydraulics of congeneric tree species from adjacent tropical savanna and forest ecosystems.Oecologia, 155, 405-415. |
| 14 | Hoffmann WA, Marchin RM, Abit P, Lau OL (2011). Hydraulic failure and tree dieback are associated with high wood density in a temperate forest under extreme drought.Global Change Biology, 17, 2731-2742. |
| 15 | Jacobsen AL, Agenbag L, Esler KJ, Pratt RB, Ewers FW, Davis SD (2007). Xylem density, biomechanics and anatomical traits correlate with water stress in 17 evergreen shrub species of the Mediterranean-type climate region of South Africa.Journal of Ecology, 95, 171-183. |
| 16 | Jin Y, Wang CK (2015). Trade-offs between plant leaf hydraulic and economic traits.Chinese Journal of Plant Ecology, 39, 1021-1032. (in Chinese with English abstract)[金鹰, 王传宽 (2015). 植物叶片水力与经济性状权衡关系的研究进展. 植物生态学报, 39, 1021-1032.] |
| 17 | Johnson DM, Mcculloh KA, Woodruff DR, Meinzer FC (2012). Evidence for xylem embolism as a primary factor in dehydration-induced declines in leaf hydraulic conductance.Plant, Cell & Environment, 35, 760-769. |
| 18 | Kim YX, Steudle E (2007). Light and turgor affect the water permeability (aquaporins) of parenchyma cells in the midrib ofZea mays. Journal of Experimental Botany, 58, 4119-4129. |
| 19 | Li JY, Zhai HB (2000). Hydraulic architecture and drought resistance of woody plants.Chinese Journal of Applied Ecology, 11, 301-305. (in Chinese with English abstract)[李吉跃, 翟洪波 (2000). 木本植物水力结构与抗旱性. 应用生态学报, 11, 301-305.] |
| 20 | McCulloh K, Sperry JS, Lachenbruch B, Meinzer FC, Reich PB (2010). Moving water well: Comparing hydraulic efficiency in twigs and trunks of coniferous, ring-porous, and diffuse-porous saplings from temperate and tropical forests.New Phytologist, 186, 439-450. |
| 21 | Meinzer FC, Johnson DM, Lachenbruch B, McCulloh KA, Woodruff DR (2009). Xylem hydraulic safety margins in woody plants: Coordination of stomatal control of xylem tension with hydraulic capacitance.Functional Ecology, 23, 922-930. |
| 22 | Nardini A, Battistuzzo M, Savi T (2013). Shoot desiccation and hydraulic failure in temperate woody angiosperms during an extreme summer drought.New Phytologist, 200, 322-329. |
| 23 | Nardini A, Luglio J (2014). Leaf hydraulic capacity and drought vulnerability: Possible trade-offs and correlations with climate across three major biomes.Functional Ecology, 28, 810-818. |
| 24 | Nardini A, Pedà G, Rocca NL (2012a). Trade-offs between leaf hydraulic capacity and drought vulnerability: Morpho- anatomical bases, carbon costs and ecological cones- quences.New Phytologist, 196, 788-798. |
| 25 | Nardini A, Pedà G, Salleo S (2012b). Alternative methods for scaling leaf hydraulic conductance offer new insights into the structure-function relationships of sun and shade leaves.Functional Plant Biology, 39, 394-401. |
| 26 | Nardini A, Salleo S, Raimondo F (2003). Changes in leaf hydraulic conductance correlate with leaf vein embolism in Cercis siliquastrum L.Trees, 17, 529-534. |
| 27 | Pan X, Qiu Q, Li JY, Wang JH, He Q, Su Y, Ma JW, Du K (2015). Drought resistance evaluation based on leaf anatomical structures of 25 shrubs on the Tibetan Plateau.Journal of South China Agricultural University, 36, 61-68. (in Chinese with English abstract)[潘听, 邱权, 李吉跃, 王军辉, 何茜, 苏艳, 马建伟, 杜坤 (2015). 基于叶片解剖结构对青藏高原25种灌木的抗旱性评价. 华南农业大学学报, 36, 61-68.] |
| 28 | Pan YP, Chen YP (2014). Recent advances in leaf hydraulic traits.Chinese Journal of Ecology, 33, 2834-2841. (in Chinese with English abstract)[潘莹萍, 陈亚鹏 (2014). 叶片水力性状研究进展. 生态学杂志, 33, 2834-2841.] |
| 29 | Sack L, Holbrook NM (2006). Leaf hydraulics.Annual Review of Plant Biology, 57, 361-381. |
| 30 | Scholz FG, Bucci SJ, Goldstein G (2014). Strong hydraulic segmentation and leaf senescence due to dehydration may trigger die-back in Nothofagus dombeyi under severe droughts: A comparison with the co-occurring Austrocedrus chilensis.Trees, 28, 1475-1487. |
| 31 | Scoffoni C, McKown AD, Rawls M, Sack L (2012). Dynamics of leaf hydraulic conductance with water status: Quantification and analysis of species differences under steady state. Journal of Experimental Botany, 63, 643-658. |
| 32 | Scoffoni C, Rawls M, McKown A, Cochard H, Sack L (2011). Decline of leaf hydraulic conductance with dehydration: Relationship to leaf size and venation architecture.Plant Physiology, 156, 832-843. |
| 33 | Simonin KA, Limm EB, Dawson TE (2012). Hydraulic conductance of leaves correlates with leaf lifespan: Implications for lifetime carbon gain.New Phytologist, 193, 939-947. |
| 34 | Sperry JS, Hacke UG, Pittermann J (2006). Size and function in conifer tracheids and angiosperm vessels.American Journal of Botany, 93, 1490-1500. |
| 35 | Sperry JS, Meinzer FC, McCulloh KA (2008). Safety and efficiency conflicts in hydraulic architecture: Scaling from tissues to trees.Plant, Cell & Environment, 31, 632-645. |
| 36 | Tyree MT, Hammel HT (1972). The measurement of the turgor pressure and the water relations of plants by the pressure- bomb technique.Journal of Experimental Botany, 23, 267-282. |
| 37 | Vilagrosaa A, Morales F, Abadía A, Bellot J, Cochardd H, Gil-Pelegrine E (2010). Are symplast tolerance to intense drought conditions and xylem vulnerability to cavitation coordinated? An integrated analysis of photosynthetic, hydraulic and leaf level processes in two Mediterranean drought-resistant species.Environmental & Experimental Botany, 69, 233-242. |
| 38 | Villagra M, Campanello PI, Bucci SJ, Goldstein G (2013). Functional relationships between leaf hydraulics and leaf economic traits in response to nutrient addition in subtropical tree species.Tree Physiology, 33, 1308-1318. |
| 39 | Wang CK, Han Yi, Chen JQ, Wang XC, Zhang QZ, Bond- Lamberty B (2013). Seasonality of soil CO2 efflux in a temperate forest: Biophysical effects of snowpack and spring freeze-thaw cycles.Agricultural & Forest Meteorology, 177, 83-92. |
| 40 | Wikberg J, Ögren E (2004). Interrelationships between water use and growth traits in biomass-producing willows.Trees, 18, 70-76. |
| 41 | Zhang HY, Wang CK, Wang XC (2014). Spatial variations in non-structural carbohydrates in stems of twelve temperate tree species.Trees, 28, 77-89. |
| 42 | Zhang SB, Zhang JL, Cao KF (2016). Effects of seasonal drought on water status, leaf spectral traits and fluorescence parameters in Tarenna depauperata Hutchins, a Chinese savanna evergreen species.Plant Science Journal, 34, 117-126. (in Chinese with English abstract)[张树斌, 张教林, 曹坤芳 (2016). 季节性干旱对白皮乌口树(Tarenna depauperata Hutchins)水分状况、叶片光谱特征和荧光参数的影响. 植物科学学报, 34, 117-126.] |
| 43 | Zhang YJ, Cao KF, Sack L, Li N, Wei XM, Goldstein G (2015). Extending the generality of leaf economic design principles in the cycads, an ancient lineage.New Phytologist, 206, 817-829. |
| 44 | Zhang ZL, Liu GD, Zhang FC, Zheng CX, Kang YH (2014). Research progress of plant leaf hydraulic conductivity.Chinese Journal of Ecology, 33, 1663-1670. (in Chinese with English abstract)[张志亮, 刘国东, 张富仓, 郑彩霞, 康银红 (2014). 植物叶片导水率的研究进展. 生态学杂志, 33, 1663-1670.] |
| 45 | Zhuo LX, Li JH, Li YY, Zhao LM (2012). Comparison of hydraulic traits in branches and leaves of diffuse- and ring- porous species.Acta Ecological Sinica, 32, 5087-5094. (in Chinese with English Abstract)[左力翔, 李俊辉, 李秧秧, 赵丽敏 (2012). 散孔材与环孔材树种枝干, 叶水力学特性的比较研究. 生态学报, 32, 5087-5094.] |
| 46 | Zhu SD, Chen YJ, Cao KF, Ye Q (2015). Interspecific variation in branch and leaf traits among three Syzygium tree species from different successional tropical forests.Functional Plant Biology, 42, 423-432. |
| 47 | Zwieniecki MA, Brodribb TJ, Holbrook NM (2007). Hydraulic design of leaves: Insights from rehydration kinetics.Plant, Cell & Environment, 30, 910-921. |
| [1] | 和璐璐, 张萱, 章毓文, 王晓霞, 刘亚栋, 刘岩, 范子莹, 何远洋, 席本野, 段劼. 辽东山区不同坡向长白落叶松人工林树冠特征与林木生长关系[J]. 植物生态学报, 2023, 47(11): 1523-1539. |
| [2] | 任金培, 李俊鹏, 王卫锋, 代永欣, 王林. 八个树种叶水力性状对水分条件的响应及其驱动因素[J]. 植物生态学报, 2021, 45(9): 942-951. |
| [3] | 李豪, 马如玉, 强波, 贺聪, 韩路, 王海珍. 胡杨当年生小枝茎构型对展叶效率的影响[J]. 植物生态学报, 2021, 45(11): 1251-1262. |
| [4] | 宋慧清, 倪鸣源, 朱师丹. 乔木与木质藤本的水力与光合性状的差异: 以热带森林崖豆藤属和买麻藤属为例[J]. 植物生态学报, 2020, 44(3): 192-204. |
| [5] | 宋清华, 赵成章, 史元春, 杜晶, 王继伟, 陈静. 不同坡向甘肃臭草根系分叉数和连接长度的权衡关系[J]. 植物生态学报, 2015, 39(6): 577-585. |
| [6] | 张茜,赵成章,董小刚,马小丽,侯兆疆,李钰. 高寒退化草地狼毒种群不同海拔花大小-数量的权衡关系[J]. 植物生态学报, 2014, 38(5): 452-459. |
| [7] | 杨冬梅,占峰,张宏伟. 清凉峰不同海拔木本植物小枝内叶大小-数量权衡关系[J]. 植物生态学报, 2012, 36(4): 281-291. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
Copyright © 2022 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19