|
|
||||||||||||||
|
全球变化对食物网结构影响机制的研究进展
植物生态学报
2021, 45 (10):
1064-1074.
DOI: 10.17521/cjpe.2020.0061
食物网主要依靠基于不同营养级间物种互作形成的上行与下行调控维持其结构。全球变化能够改变种间关系, 威胁生物多样性的维持, 然而目前对全球变化改变食物网结构的机制仍处于探索阶段。近年来通过大时空格局与多营养级食物网研究, 发现全球变化的作用机制主要可归结为3种: 物候错配、关键种丧失与生物入侵。该文聚焦于这3种机制, 综述各种机制造成的食物网结构变化并探讨相关的进化与生态驱动因素。三种干扰机制均通过改变原有种间关系, 影响食物网调控, 改变食物网结构。不同的是, 物候错配造成的种间关系变化是由于不同物种的物候对全球变化产生非同步响应所致; 关键种丧失则使营养级间取食/捕食关系发生变化甚至缺失; 而入侵物种通过竞争排除同营养级物种改变种间关系。最后, 该文提出食物网结构变化的实质是物种是否能够适应快速变化的生态环境, 并据此展望未来研究方向。随着全球变化影响日益加剧, 急需继续深入探索导致全球变化下食物网结构改变的机制, 为制定合理的生物多样性保护与生态修复规划提供重要理论支撑。
表1
文中相关专业术语解释
正文中引用本图/表的段落
除全球气候变化外, 许多其他类型的人类干扰也能够改变区域气候。例如, 生境片段化将大生境斑块分割为大量小生境片段, 导致边缘效应增强, 改变原有的林内小气候并改变物候特征, 可能导致物候错配进而影响种间关系的稳定(Haddad et al., 2015; Wilson et al., 2016)。物候特征异步变化是导致物候错配的主要因素。在受干扰环境中物候特征变化速率可能与物种的世代周期有关, 世代周期越短的生物适应环境变化的速度越快, 这与草本、灌木和小型消费者(例如植食性昆虫)物候提前幅度较大一致(Both et al., 2009; Thackeray et al., 2016)。目前尚不清楚不同生物类群适应气候变化的潜在机制, 这可能涉及由温度变化引发激素分泌变化调整生长节律以及表观遗传因子引发的可塑性变化。但是, 物种对气候变化的耐受能力是有极限的, Trisos等(2020)研究了30 000种陆地和海洋生物能够承受的温度阈值并根据模型预测发现大量物种将由于全球气温持续升高而在近期集体灭绝。然而, 在某些生态系统中气候变化却未导致物候错配的发生(Hansson et al., 2013)。因此, 需要更多案例研究以全面分析导致物候特征变化异步性的生物(例如种内遗传变异水平)与环境因素(气候变暖程度、昼夜温差范围、生境斑块大小等)并帮助预测未来不同生态系统中食物网结构演变的趋势。此外, 地下生物群落尤其是土壤微生物与小型土壤动物群落对生态系统服务功能贡献巨大且对食物网结构具有重要的支持作用, 但由于物种鉴定困难等原因, 目前关于此类群落中物候错配的报道较少。未来可结合DNA条形码(表1)技术深入了解这些群落中的食物网结构与动态及其对气候变化的响应。
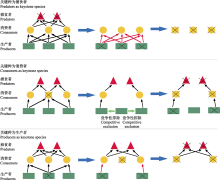
某些消费者也是关键种, 它们往往既以多种生产者为食, 也是众多捕食者的猎物。理论模型预测消费者关键种的丧失, 可能阻碍营养级间的正常能量流动, 抑制捕食者的能量来源, 影响捕食者多度与食物网稳定性(Kadoya et al., 2018)。同时, 消费者关键种丧失可能改变表观竞争格局, 导致植物间竞争加剧, 物种数减少, 降低食物网复杂性(图1)。传粉者、种子传播动物与根瘤菌是另一类消费者关键种, 它们与植物构成的互惠网络维持了植物种群的繁殖与更新。这一物种类群的衰退已对生物多样性与生态系统服务功能的维持构成严重威胁(Potts et al., 2010)。消费者关键种丧失与过度狩猎、生境片段化与气候变化等干扰因素有关(Potts et al., 2010; Ripple et al., 2015)。
关键种丧失对食物网结构与稳定性造成何种程度的影响与食物网复杂性有关。在复杂食物网中往往很少存在关键种, 因为在这样的食物网中很少存在不可替代的物种。因而即便某一营养级中物种的多样性发生变化, 其效应可能仅能传递至相邻营养级(Scherber et al., 2010)。据此推测在高纬度高海拔地区, 由于本地生态系统中食物网结构简单, 关键种丧失将造成较为严重的影响。此外, 食物网结构形成的生态与进化机制目前仍有待研究。目前认为不同营养级间的互作物种可能存在进化军备竞赛(表1), 例如菜粉蝶(Pieris rapae)与其宿主植物拟南芥(Arabidopsis thaliana)在细胞色素P450基因家族与谷胱甘肽转移酶等涉及植物防御与动物解毒功能的基因在进化上存在显著相关性, 这在一定程度上解释了消费者与生产者间相互适应的方式(Nallu et al., 2017)。了解这种进化方式的起源与历程, 有助于揭示关键种的形成机制并预测其丧失带来的长期效应。
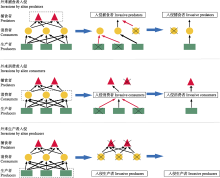
外来入侵物种对同营养级物种具有很强的竞争优势, 这可能是由于其具有内禀优势(表1), 资源利用效率高, 可塑性强等特点, 也可能是经历了快速适应性进化与天敌释放过程, 或者获得互利共生生物的协助(鞠瑞亭等, 2012)。因此在入侵地食物网中, 外来入侵物种通常没有强大的天敌且对低营养级物种的捕食强度很高, 排除同营养级物种后将大大降低食物网复杂性并阻碍营养与能量向高营养级流动(Ehrenfeld, 2010; Gallardo et al., 2016)。
目前普遍认为入侵植物对食物网结构的破坏最为严重。许多入侵植物能在入侵地获得本地传粉与种子传播动物以及共生微生物(如根瘤菌), 在剥夺本地植物互惠网络的同时促进其种群扩张(Traveset & Richardson, 2014)。例如, 外来千屈菜科植物Lythrum salicaria的花序密度远高于本地植物种类, 因而成功吸引更多的传粉昆虫为其传粉, 使传粉者造访本地植物花序的频率至少下降了20% (Goodell & Parker, 2017)。许多豆科入侵植物就在入侵地获得了共生根瘤菌, 从而大大提高了其在新环境中的适合度(Traveset & Richardson, 2014)。在缺乏消费者取食的情况下, 入侵植物通过竞争排除其他植物, 最后甚至可能导致本地消费者与捕食者灭绝, 引发整个食物网的崩溃(Pyšek et al., 2012; Gallardo et al., 2016)(图2)。例如, 外来植物互花米草(Spartina alterniflora)入侵上海崇明东滩湿地后, 造成本地优势植物海三棱藨草(Scirpus mariqueter)分布区迅速缩小, 改变了地表植物碎屑理化性质与植物根系氧气含量, 进而影响地下微生物与底栖动物群落组成, 最终降低鸟类尤其是长距离迁徙候鸟的多样性(鞠瑞亭等, 2012; Tong et al., 2018)。在水生生态系统中, 大型入侵水生植物能够从根本上改变本地食物网结构, 大大减少处于最高营养级的鱼类多度(Gallardo et al., 2016)。外来植物入侵还可能协助更多外来物种的成功侵入, 比如入侵松科植物与其共生菌根真菌(Dickie et al., 2010), 加剧生物入侵对食物网的影响, 造成入侵崩溃(表1)。此外, 入侵植物降低植物多样性, 能够改变土壤微生物群落, 影响地下食物网结构(Kardol & Wardle, 2010)。
本文的其它图/表
|