
|
|
||
|
全球变化对食物网结构影响机制的研究进展
植物生态学报
2021, 45 (10):
1064-1074.
DOI: 10.17521/cjpe.2020.0061
食物网主要依靠基于不同营养级间物种互作形成的上行与下行调控维持其结构。全球变化能够改变种间关系, 威胁生物多样性的维持, 然而目前对全球变化改变食物网结构的机制仍处于探索阶段。近年来通过大时空格局与多营养级食物网研究, 发现全球变化的作用机制主要可归结为3种: 物候错配、关键种丧失与生物入侵。该文聚焦于这3种机制, 综述各种机制造成的食物网结构变化并探讨相关的进化与生态驱动因素。三种干扰机制均通过改变原有种间关系, 影响食物网调控, 改变食物网结构。不同的是, 物候错配造成的种间关系变化是由于不同物种的物候对全球变化产生非同步响应所致; 关键种丧失则使营养级间取食/捕食关系发生变化甚至缺失; 而入侵物种通过竞争排除同营养级物种改变种间关系。最后, 该文提出食物网结构变化的实质是物种是否能够适应快速变化的生态环境, 并据此展望未来研究方向。随着全球变化影响日益加剧, 急需继续深入探索导致全球变化下食物网结构改变的机制, 为制定合理的生物多样性保护与生态修复规划提供重要理论支撑。 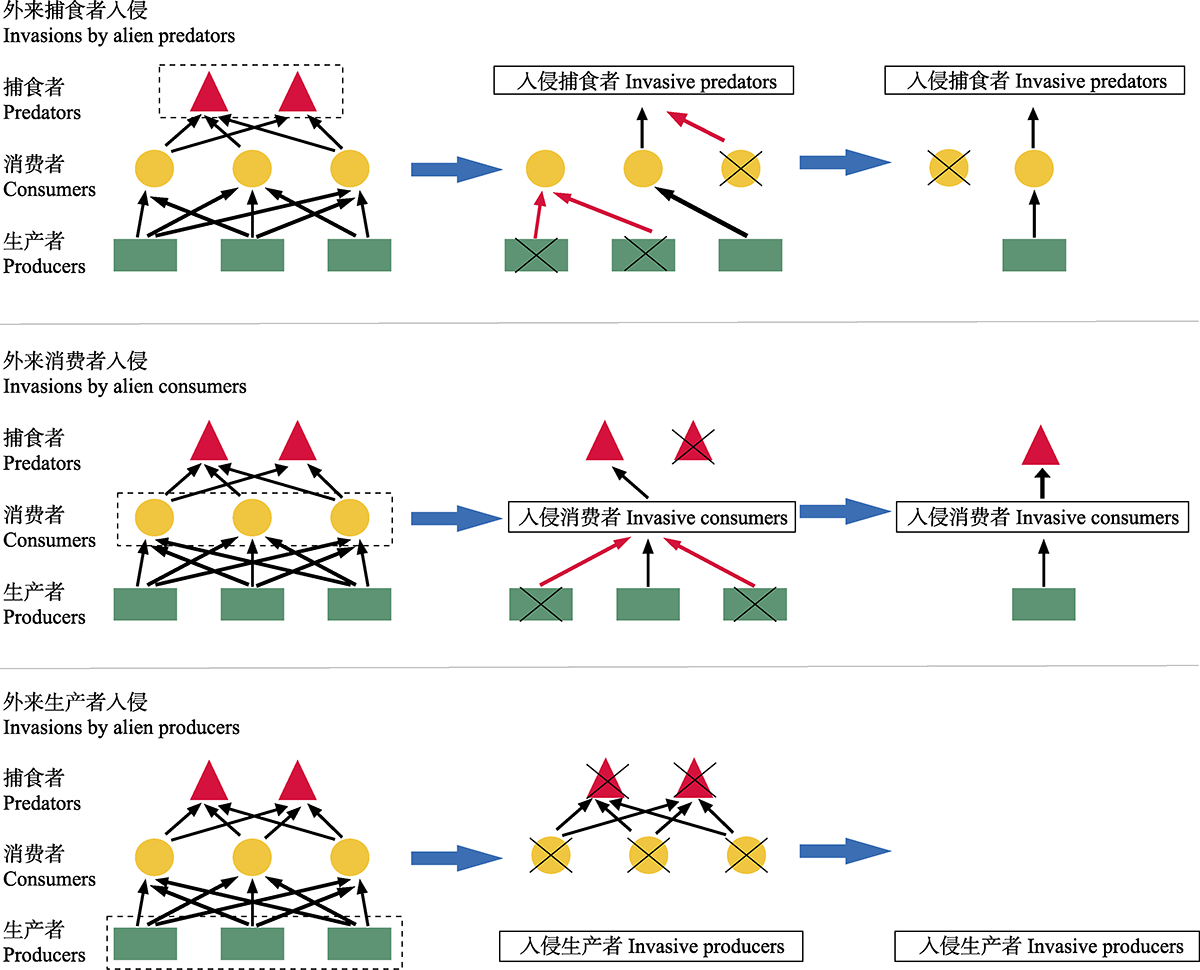 View image in article
图2
外来物种入侵影响食物网结构示意图。黑色箭头表示营养级关系; 红色箭头表示消费者取食力度或捕食者捕食强度增强。入侵捕食者在竞争排除同营养级中乡土种的同时由于其捕食偏好改变消费者表观竞争格局并影响低营养级; 入侵消费者通过竞争减少消费者物种数对生产者产生较大的取食压力, 同时阻断顶级捕食者的食物来源, 进而破坏食物网的上行和下行调控, 改变食物网结构; 在缺乏消费者取食的情况下, 入侵生产者通过竞争排除其他生产者并阻断能量流动, 导致消费者与捕食者灭绝, 引发整个食物网的崩溃。
正文中引用本图/表的段落
目前普遍认为入侵植物对食物网结构的破坏最为严重。许多入侵植物能在入侵地获得本地传粉与种子传播动物以及共生微生物(如根瘤菌), 在剥夺本地植物互惠网络的同时促进其种群扩张(Traveset & Richardson, 2014)。例如, 外来千屈菜科植物Lythrum salicaria的花序密度远高于本地植物种类, 因而成功吸引更多的传粉昆虫为其传粉, 使传粉者造访本地植物花序的频率至少下降了20% (Goodell & Parker, 2017)。许多豆科入侵植物就在入侵地获得了共生根瘤菌, 从而大大提高了其在新环境中的适合度(Traveset & Richardson, 2014)。在缺乏消费者取食的情况下, 入侵植物通过竞争排除其他植物, 最后甚至可能导致本地消费者与捕食者灭绝, 引发整个食物网的崩溃(Pyšek et al., 2012; Gallardo et al., 2016)(图2)。例如, 外来植物互花米草(Spartina alterniflora)入侵上海崇明东滩湿地后, 造成本地优势植物海三棱藨草(Scirpus mariqueter)分布区迅速缩小, 改变了地表植物碎屑理化性质与植物根系氧气含量, 进而影响地下微生物与底栖动物群落组成, 最终降低鸟类尤其是长距离迁徙候鸟的多样性(鞠瑞亭等, 2012; Tong et al., 2018)。在水生生态系统中, 大型入侵水生植物能够从根本上改变本地食物网结构, 大大减少处于最高营养级的鱼类多度(Gallardo et al., 2016)。外来植物入侵还可能协助更多外来物种的成功侵入, 比如入侵松科植物与其共生菌根真菌(Dickie et al., 2010), 加剧生物入侵对食物网的影响, 造成入侵崩溃(表1)。此外, 入侵植物降低植物多样性, 能够改变土壤微生物群落, 影响地下食物网结构(Kardol & Wardle, 2010)。
入侵捕食者或者病原体在竞争排除同营养级中乡土种的同时可能由于其捕食偏好改变消费者表观竞争格局, 由此产生的营养级联效应可能导致消费者物种组成与群落结构的改变并影响低营养级物种(图2)。外来鱼类物种入侵能够抑制底栖动物与浮游生物数量, 从而促进浮游植物的生长(Gallardo et al., 2016)。此外, 某些入侵捕食者通过捕食在不同生境间迁徙的消费者, 从而改变本地植被组成。例如, 北极狐(Alopex lagopus)入侵阿留申群岛的部分岛屿后大肆捕猎海鸟, 导致海鸟数量快速下降, 由此造成鸟粪匮乏, 使得土壤肥力下降, 从而导致当地原有的草原植被类型向适应贫瘠土壤的苔原植被类型转变(Croll et al., 2005)。然而某些广食性外来捕食者的入侵也可能替代原有捕食者的作用, 通过下行调控维持食物网稳定。例如, 在澳大利亚干旱地区, 移除外来顶级捕食者澳洲野狗(Canis lupus dingo)致使消费者与中型捕食者赤狐(Vulpes vulpes)的密度提高, 从而使草地盖度与小型哺乳动物种类减少(Letnic et al., 2009)。
入侵消费者与入侵杂食动物对食物网影响的研究案例较少, 其潜在威胁可能是通过竞争排除减少消费者物种数并对生产者产生较大的取食压力, 同时阻断部分顶级捕食者的食物来源, 进而破坏食物网的上行和下行调控, 改变食物网结构(图2)。例如, Gibson等(2013)发现在泰国水库岛屿森林中, 本地小型哺乳动物由于外来啮齿动物的入侵而加速灭绝。此外, 入侵土壤微生物可能影响地表植物获取根瘤菌, 从而影响植物多样性以及食物网结构(Ehrenfeld, 2010; Kardol &Wardle, 2010)。
种间关系的维持往往伴随着互作物种间长期的协同进化, 通常这种进化关系越紧密种间关系越牢固, 受外界干扰而发生快速变化的可能性就较低; 而协同进化关系不紧密的互作物种在全球变化背景下更容易发生种间关系变化(例如种间互作强度变化、互惠关系向拮抗关系转变等), 进而导致食物网结构迅速改变(Futuyma & Agrawal, 2009; Kiers et al., 2010)。然而, 具有紧密协同进化关系的物种一般为专性物种(specialist), 双方任何一方灭绝经常会导致另一方也灭绝, 因此在全球变化干扰强度大的情况下与泛性物种(generalist)相比具有较高的灭绝风险(Weiner et al., 2014)(图1, 图2)。此外, 在复杂食物网中并非所有物种均与其他任一物种间存在种间关系, 往往形成与部分物种间互作关系密切而与其他物种间关系较为疏远的情况, 即形成了嵌套结构(Dalsgaard et al., 2013)。目前认为嵌套结构的存在能够有效阻止种间关系变化产生的影响蔓延至整个食物网, 因而促进群落稳定性和物种共存(Neutel et al., 2007)。
本文的其它图/表
|
Copyright © 2026 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19
![]()
{kind=link}