不同水热梯度下冠层优势树种叶片热力性状及适应策略的变化趋势
周莹莹, 林华
植物生态学报
2023, 47 ( 5):
733-744.
DOI: 10.17521/cjpe.2022.0289
叶片温度是植物能量交换和生理过程发生的重要微环境参数。叶片热力性状能够在一定程度上调控叶片温度, 避免极端温度对叶片的伤害。但目前针对叶片热力性状的研究还很少。该研究选择云南省从热带到温带具有明显水热梯度的4种典型植被类型: 干热河谷植被、热带雨林、亚热带常绿阔叶林、温带针阔混交林, 对其冠层优势树种的叶片热力性状进行了系统地研究。这些性状包括了可能影响叶片温度的形态、光学、材料特性、解剖和生理的23个性状。研究结果表明: 干热河谷植被的植物主要依靠蒸腾降温, 叶片薄, 叶寿命短, 主要为“快速投资-收益”型植物; 热带雨林植物叶片大, 蒸腾速率不高, 不利于降温, 较厚的叶片、较高的含水量能在一定程度上缓解高温, 采取“慢速投资-收益”策略; 亚热带常绿阔叶林很少发生极端温度, 叶片没有明显的热适应性状, 叶片厚, 叶寿命长, 采取“慢速投资-收益”策略; 温带针阔混交林植被的叶片小而厚, 多成簇状生长, 有一定保温作用。温带针阔混交林的冠层常绿植物光合速率较低, 偏“慢速投资-收益”型; 而落叶植物的光合速率较高, 偏“快速投资-收益”型。该研究系统地研究了热力性状与植物适应策略沿水热梯度的变化, 为深入认识植物对环境适应策略提供了理论基础。
View image in article
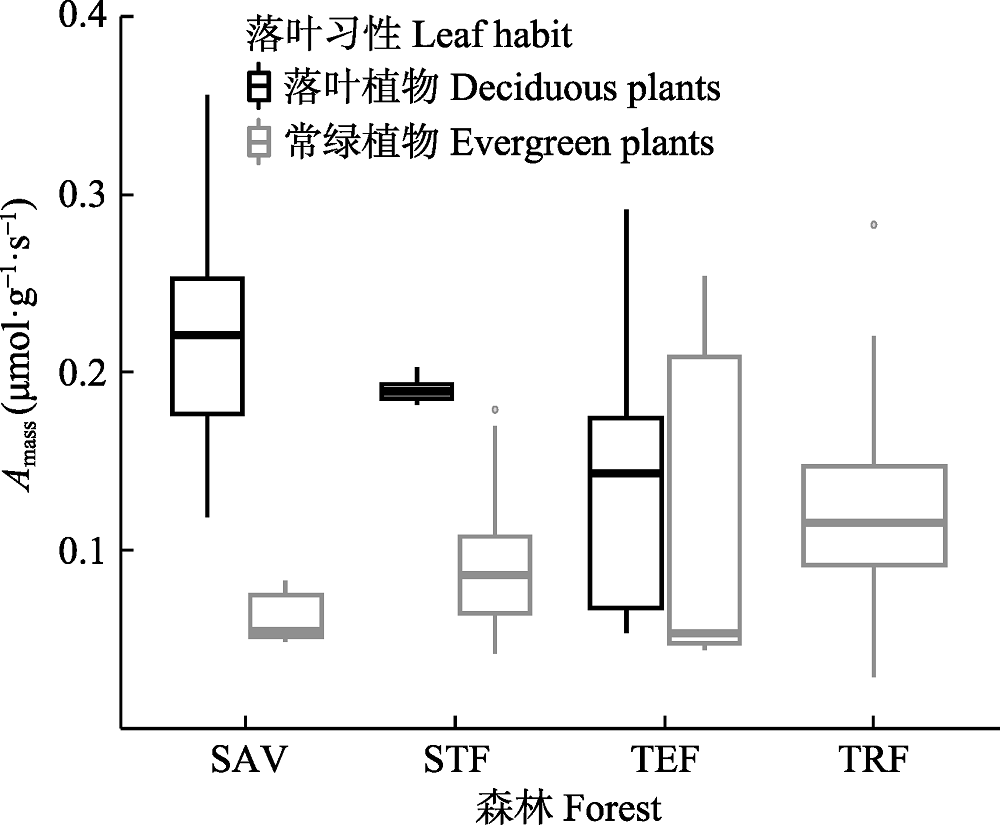
图4
云南省具有明显水热梯度的不同森林的单位质量最大净光合速率(Amass)。SAV, 元江干热河谷植被; TRF, 西双版纳热带雨林; STF, 哀牢山亚热带常绿阔叶林; TEF, 丽江温带针阔混交林。
正文中引用本图/表的段落
为适应不同的水热环境, 叶片性状表现出不同的特征。叶片形态性状方面, 热带雨林植物的叶片较大。主要因为热带雨林叶面积指数最高, 叶片间相互遮阴相对更严重, 大叶片有利于获得更多的光能。同时, 大叶片和高叶面积指数可以提高总的碳合成量, 从而保持较高生产力(Wright et al., 2017)。叶片材料特性方面, 比叶质量随着环境温度的降低而增加, 干物质含量也有相同的趋势, 但是差异不显著。冷的地方, 细胞生长速率慢, 单位面积的小细胞数量增多, 叶片小而厚, 密度大(Atkins, 2005; Poorter et al., 2009)。因此, 比叶质量越大的植物, 越偏向“慢速投资-收益”型。再者, 干物质含量增加有利于植物抵御霜冻和大风。因此, 寒冷和高海拔地区的植物一般具有更高的比叶质量(K?rner et al., 1986; Woodward, 1987; Atkins, 2005)。干热河谷植物细胞生长速率快, 因而叶片较薄, 比叶质量小。叶片生理性状方面, 从干热河谷植物到亚热带常绿阔叶林植物, 随着环境温度的降低, 光合速率降低。除了温度外, 生活型对光合速率的影响也很大(刘长成等, 2011)。本研究的结果也显示, 在每个样地内, 落叶植物的光合速率均显著高于常绿植物(图4)。干热河谷和温带针阔混交林由于落叶植物和常绿植物共存, 且落叶植物占优势, 所以光合速率在样地内变异很大, 而且平均值较高。
每株选取10-15片叶片, 使用精度为0.001 g的电子天平测量叶片鲜质量; 然后将叶片平放在扫描仪上进行扫描, 用ImageJ测定叶片的周长和面积; 用叶绿素测定仪SPAD-502 (Minolta, Osaka, Japan) 测定叶片绿度(避开主脉测量4次), 接着用光谱仪(USB4000, Ocean Optics, Delray Beach, USA)测量叶片的反射率和透射率, 根据反射率和透射率的值计算叶片吸收率; 最后将叶片装入信封袋中, 60 ℃烘48 h至恒质量后测量叶片干质量.计算单位面积叶片含水量(EWT, mg·cm-2)、叶干物质含量(LDMC, g·g-1)和比叶质量(LMA, mg·cm-2), 计算公式如下: ... 四种不同生活型植物幼苗对喀斯特生境干旱的生理生态适应性 1 2011 ... 为适应不同的水热环境, 叶片性状表现出不同的特征.叶片形态性状方面, 热带雨林植物的叶片较大.主要因为热带雨林叶面积指数最高, 叶片间相互遮阴相对更严重, 大叶片有利于获得更多的光能.同时, 大叶片和高叶面积指数可以提高总的碳合成量, 从而保持较高生产力(Wright et al., 2017).叶片材料特性方面, 比叶质量随着环境温度的降低而增加, 干物质含量也有相同的趋势, 但是差异不显著.冷的地方, 细胞生长速率慢, 单位面积的小细胞数量增多, 叶片小而厚, 密度大(Atkins, 2005; Poorter et al., 2009).因此, 比叶质量越大的植物, 越偏向“慢速投资-收益”型.再者, 干物质含量增加有利于植物抵御霜冻和大风.因此, 寒冷和高海拔地区的植物一般具有更高的比叶质量(K?rner et al., 1986; Woodward, 1987; Atkins, 2005).干热河谷植物细胞生长速率快, 因而叶片较薄, 比叶质量小.叶片生理性状方面, 从干热河谷植物到亚热带常绿阔叶林植物, 随着环境温度的降低, 光合速率降低.除了温度外, 生活型对光合速率的影响也很大(刘长成等, 2011).本研究的结果也显示, 在每个样地内, 落叶植物的光合速率均显著高于常绿植物(图4).干热河谷和温带针阔混交林由于落叶植物和常绿植物共存, 且落叶植物占优势, 所以光合速率在样地内变异很大, 而且平均值较高. ... 四种不同生活型植物幼苗对喀斯特生境干旱的生理生态适应性 1 2011 ... 为适应不同的水热环境, 叶片性状表现出不同的特征.叶片形态性状方面, 热带雨林植物的叶片较大.主要因为热带雨林叶面积指数最高, 叶片间相互遮阴相对更严重, 大叶片有利于获得更多的光能.同时, 大叶片和高叶面积指数可以提高总的碳合成量, 从而保持较高生产力(Wright et al., 2017).叶片材料特性方面, 比叶质量随着环境温度的降低而增加, 干物质含量也有相同的趋势, 但是差异不显著.冷的地方, 细胞生长速率慢, 单位面积的小细胞数量增多, 叶片小而厚, 密度大(Atkins, 2005; Poorter et al., 2009).因此, 比叶质量越大的植物, 越偏向“慢速投资-收益”型.再者, 干物质含量增加有利于植物抵御霜冻和大风.因此, 寒冷和高海拔地区的植物一般具有更高的比叶质量(K?rner et al., 1986; Woodward, 1987; Atkins, 2005).干热河谷植物细胞生长速率快, 因而叶片较薄, 比叶质量小.叶片生理性状方面, 从干热河谷植物到亚热带常绿阔叶林植物, 随着环境温度的降低, 光合速率降低.除了温度外, 生活型对光合速率的影响也很大(刘长成等, 2011).本研究的结果也显示, 在每个样地内, 落叶植物的光合速率均显著高于常绿植物(图4).干热河谷和温带针阔混交林由于落叶植物和常绿植物共存, 且落叶植物占优势, 所以光合速率在样地内变异很大, 而且平均值较高. ... Leaf-size divergence along rainfall and soil-nutrient gradients: Is the method of size reduction common among clades? 1 2003 ... 在物理性状方面, 叶片大小与叶片的边界层厚度正相关(Fetcher, 1981; Campbell & Norman, 2000).叶片与外界环境的热量和气体交换都要通过边界层, 边界层越厚, 交换效率越低.因此, 高温干旱地区的植物叶片面积较小, 可以更充分地与外界环境进行热量交换, 从而达到散热的目的(Geng et al., 2012; Pierce et al., 2012).但高温潮湿的地区, 叶片可以通过蒸腾散热, 叶片面积较大.低温地区的植物, 夜间叶片温度低于环境温度, 小叶片有利于从环境中吸收热量(McDonald et al., 2003; Peppe et al., 2011).研究表明, 夜间气温对叶片大小的影响贡献为51%, 昼间气温的贡献为38% (Wright et al., 2017).叶片颜色、反射率和吸收率影响叶片的辐射收支, 高温环境下, 减少叶片辐射吸收, 可以降低叶片温度; 而低温环境下, 增加叶片辐射吸收, 可以提高叶片温度.叶片材料特性包括含水量、密度和比叶质量等.叶片含水量高能够缓解叶片升温, 因此极端干热环境下, 肉质植物所占比例较高(Schymanski et al., 2013).植物叶片密度高, 非结构性碳以及半纤维素含量高, 有利于抵御寒冷和强风, 因此寒冷和高海拔地区植物的叶片密度较高(K?rner et al., 1986; Woodward, 1987; Atkins, 2005).叶片的解剖性状与叶片内的气体交换效率有关.叶片光合作用发生的场所——叶绿体主要是位于栅栏组织中, 栅栏组织有利于提高叶片的光合作用, 而海绵组织中有较大的细胞间隙便于气体运输(Kenzo et al., 2004).叶脉特征反映水分、养分和光合产物等物质运输的效率和安全性(Blonder et al., 2011; 李乐等, 2013).气孔是植物水分和CO2进出的门户, 决定了气体交换速率.气孔的结构特征可以反映植物的最大光合和蒸腾能力(Sack et al., 2003).在叶片生理性状方面, 叶片蒸腾是最有效的一种降温方式.由于水汽和CO2都从气孔进出, 因此它们之间是相互耦合的.目前, 主流的观点认为, 叶片的性状组合都是为最大化碳收益(Cordell et al., 2001).但也有学者认为, 极端高温环境下, 植物也可能优先蒸腾降温(Blonder & Michaletz, 2018).本研究假设干热和湿热环境下的植物会发育不同的热力性状来应对高温, 干热环境下的植物主要依靠物理降温, 湿热环境下的植物主要依靠蒸腾降温; 温湿度适宜地区的植物, 叶片性状优先满足生长, 热力性状不突出; 寒冷地区, 叶片热力性状有利于提高叶片温度. ... 大气水分亏缺对中国两种典型草地生态系统总初级生产力的影响 1 2020 ... 总体来说, 沿水热梯度, 叶片单位质量光合速率和叶片之间的权衡关系, 与全球尺度的规律(Wright et al., 2004)一致.叶脉密度与蒸腾速率的正相关关系, 反映了水分供需平衡(Sack et al., 2003; Blonder et al., 2011; Brodribb & Jordan, 2011; 张亚等, 2014).高的叶脉密度可以运输更多的水分, 从而支撑更高的蒸腾速率.蒸腾速率除了和SPI有关, 还受环境条件的制约.热带雨林和亚热带常绿阔叶林由于相对湿度较高, VPD较小, 不利于蒸腾(孟莹等, 2020), 因此叶脉密度和蒸腾速率之间的线性关系不强.叶片含水量与厚度正相关, 但相同厚度下热带雨林和干热河谷植物叶片的含水量更高, 这在一定程度上能够缓解高温的伤害. ... 大气水分亏缺对中国两种典型草地生态系统总初级生产力的影响 1 2020 ... 总体来说, 沿水热梯度, 叶片单位质量光合速率和叶片之间的权衡关系, 与全球尺度的规律(Wright et al., 2004)一致.叶脉密度与蒸腾速率的正相关关系, 反映了水分供需平衡(Sack et al., 2003; Blonder et al., 2011; Brodribb & Jordan, 2011; 张亚等, 2014).高的叶脉密度可以运输更多的水分, 从而支撑更高的蒸腾速率.蒸腾速率除了和SPI有关, 还受环境条件的制约.热带雨林和亚热带常绿阔叶林由于相对湿度较高, VPD较小, 不利于蒸腾(孟莹等, 2020), 因此叶脉密度和蒸腾速率之间的线性关系不强.叶片含水量与厚度正相关, 但相同厚度下热带雨林和干热河谷植物叶片的含水量更高, 这在一定程度上能够缓解高温的伤害. ... Plant thermoregulation: energetics, trait- environment interactions, and carbon economics 1 2015 ... 温度对植物的生长有着重要的影响.叶片温度决定了叶片气体交换和能量交换的微环境, 相对气温来说, 对植物叶片的生理过程和能量平衡更为重要(Campbell & Norman, 2000).即使在相同气温下, 叶片温度也具有很大的差异(Helliker & Richter, 2008; Michaletz et al., 2015; Fauset et al., 2018).叶片温度可以偏离气温10 ℃以上(Blonder & Michaletz, 2018).叶片温度与气温的差异, 主要由叶片性状的差异造成.根据前人的研究, 叶片物理和生理性状都可能对叶片温度造成影响.这些可能对叶片温度造成影响的性状统称为热力性状. ... The scaling of leaf area and mass: the cost of light interception increases with leaf size 2 2007 ... Leaf traits and ecological significance
每株选取10-15片叶片, 使用精度为0.001 g的电子天平测量叶片鲜质量; 然后将叶片平放在扫描仪上进行扫描, 用ImageJ测定叶片的周长和面积; 用叶绿素测定仪SPAD-502 (Minolta, Osaka, Japan) 测定叶片绿度(避开主脉测量4次), 接着用光谱仪(USB4000, Ocean Optics, Delray Beach, USA)测量叶片的反射率和透射率, 根据反射率和透射率的值计算叶片吸收率; 最后将叶片装入信封袋中, 60 ℃烘48 h至恒质量后测量叶片干质量.计算单位面积叶片含水量(EWT, mg·cm-2)、叶干物质含量(LDMC, g·g-1)和比叶质量(LMA, mg·cm-2), 计算公式如下: ... Global leaf trait relationships: mass, area, and the leaf economics spectrum 1 2013 ... 但植物除了适应环境温度, 还需要适应水分、养分等环境条件, 从而保证植株的存活, 最大化生长.因此叶片性状间既相互联系, 又相互制约.叶经济谱总结了全球尺度上叶性状的变化规律和相互关系, 定量化地描述了植物资源权衡策略的连续变化(Wright et al., 2004; Shipley et al., 2006; Lavorel, 2013), 是目前对植物性状间关系研究最为系统的理论.根据这个理论, 植物的生存策略在“快速投资-收益”到“缓慢投资-收益”间连续分布.“快速投资-收益”的植物代谢速率高, 叶氮含量高, 叶寿命短、比叶质量小; “缓慢投资-收益”的植物代谢速率慢、叶氮含量低、叶寿命长、比叶质量大(Wright et al., 2004, 2005; Osnas et al., 2013).在有限的资源环境中, 植物会在功能性状之间进行资源优化配置, 形成“此消彼长”的权衡关系.但是在叶片经济谱的研究中, 涉及叶片热力性状的研究很少, 因此, 热力性状之间是否也存在资源权衡? 不同水热梯度下, 叶片投资收益策略与热力适应策略之间有何关系还鲜有研究. ... Sensitivity of leaf size and shape to climate: global patterns and paleoclimatic applications 1 2011 ... 在物理性状方面, 叶片大小与叶片的边界层厚度正相关(Fetcher, 1981; Campbell & Norman, 2000).叶片与外界环境的热量和气体交换都要通过边界层, 边界层越厚, 交换效率越低.因此, 高温干旱地区的植物叶片面积较小, 可以更充分地与外界环境进行热量交换, 从而达到散热的目的(Geng et al., 2012; Pierce et al., 2012).但高温潮湿的地区, 叶片可以通过蒸腾散热, 叶片面积较大.低温地区的植物, 夜间叶片温度低于环境温度, 小叶片有利于从环境中吸收热量(McDonald et al., 2003; Peppe et al., 2011).研究表明, 夜间气温对叶片大小的影响贡献为51%, 昼间气温的贡献为38% (Wright et al., 2017).叶片颜色、反射率和吸收率影响叶片的辐射收支, 高温环境下, 减少叶片辐射吸收, 可以降低叶片温度; 而低温环境下, 增加叶片辐射吸收, 可以提高叶片温度.叶片材料特性包括含水量、密度和比叶质量等.叶片含水量高能够缓解叶片升温, 因此极端干热环境下, 肉质植物所占比例较高(Schymanski et al., 2013).植物叶片密度高, 非结构性碳以及半纤维素含量高, 有利于抵御寒冷和强风, 因此寒冷和高海拔地区植物的叶片密度较高(K?rner et al., 1986; Woodward, 1987; Atkins, 2005).叶片的解剖性状与叶片内的气体交换效率有关.叶片光合作用发生的场所——叶绿体主要是位于栅栏组织中, 栅栏组织有利于提高叶片的光合作用, 而海绵组织中有较大的细胞间隙便于气体运输(Kenzo et al., 2004).叶脉特征反映水分、养分和光合产物等物质运输的效率和安全性(Blonder et al., 2011; 李乐等, 2013).气孔是植物水分和CO2进出的门户, 决定了气体交换速率.气孔的结构特征可以反映植物的最大光合和蒸腾能力(Sack et al., 2003).在叶片生理性状方面, 叶片蒸腾是最有效的一种降温方式.由于水汽和CO2都从气孔进出, 因此它们之间是相互耦合的.目前, 主流的观点认为, 叶片的性状组合都是为最大化碳收益(Cordell et al., 2001).但也有学者认为, 极端高温环境下, 植物也可能优先蒸腾降温(Blonder & Michaletz, 2018).本研究假设干热和湿热环境下的植物会发育不同的热力性状来应对高温, 干热环境下的植物主要依靠物理降温, 湿热环境下的植物主要依靠蒸腾降温; 温湿度适宜地区的植物, 叶片性状优先满足生长, 热力性状不突出; 寒冷地区, 叶片热力性状有利于提高叶片温度. ... Combined use of leaf size and economics traits allows direct comparison of hydrophyte and terrestrial herbaceous adaptive strategies 1 2012 ... 在物理性状方面, 叶片大小与叶片的边界层厚度正相关(Fetcher, 1981; Campbell & Norman, 2000).叶片与外界环境的热量和气体交换都要通过边界层, 边界层越厚, 交换效率越低.因此, 高温干旱地区的植物叶片面积较小, 可以更充分地与外界环境进行热量交换, 从而达到散热的目的(Geng et al., 2012; Pierce et al., 2012).但高温潮湿的地区, 叶片可以通过蒸腾散热, 叶片面积较大.低温地区的植物, 夜间叶片温度低于环境温度, 小叶片有利于从环境中吸收热量(McDonald et al., 2003; Peppe et al., 2011).研究表明, 夜间气温对叶片大小的影响贡献为51%, 昼间气温的贡献为38% (Wright et al., 2017).叶片颜色、反射率和吸收率影响叶片的辐射收支, 高温环境下, 减少叶片辐射吸收, 可以降低叶片温度; 而低温环境下, 增加叶片辐射吸收, 可以提高叶片温度.叶片材料特性包括含水量、密度和比叶质量等.叶片含水量高能够缓解叶片升温, 因此极端干热环境下, 肉质植物所占比例较高(Schymanski et al., 2013).植物叶片密度高, 非结构性碳以及半纤维素含量高, 有利于抵御寒冷和强风, 因此寒冷和高海拔地区植物的叶片密度较高(K?rner et al., 1986; Woodward, 1987; Atkins, 2005).叶片的解剖性状与叶片内的气体交换效率有关.叶片光合作用发生的场所——叶绿体主要是位于栅栏组织中, 栅栏组织有利于提高叶片的光合作用, 而海绵组织中有较大的细胞间隙便于气体运输(Kenzo et al., 2004).叶脉特征反映水分、养分和光合产物等物质运输的效率和安全性(Blonder et al., 2011; 李乐等, 2013).气孔是植物水分和CO2进出的门户, 决定了气体交换速率.气孔的结构特征可以反映植物的最大光合和蒸腾能力(Sack et al., 2003).在叶片生理性状方面, 叶片蒸腾是最有效的一种降温方式.由于水汽和CO2都从气孔进出, 因此它们之间是相互耦合的.目前, 主流的观点认为, 叶片的性状组合都是为最大化碳收益(Cordell et al., 2001).但也有学者认为, 极端高温环境下, 植物也可能优先蒸腾降温(Blonder & Michaletz, 2018).本研究假设干热和湿热环境下的植物会发育不同的热力性状来应对高温, 干热环境下的植物主要依靠物理降温, 湿热环境下的植物主要依靠蒸腾降温; 温湿度适宜地区的植物, 叶片性状优先满足生长, 热力性状不突出; 寒冷地区, 叶片热力性状有利于提高叶片温度. ... Causes and consequences of variation in leaf mass per area (LMA): a meta-analysis 1 2009 ... 为适应不同的水热环境, 叶片性状表现出不同的特征.叶片形态性状方面, 热带雨林植物的叶片较大.主要因为热带雨林叶面积指数最高, 叶片间相互遮阴相对更严重, 大叶片有利于获得更多的光能.同时, 大叶片和高叶面积指数可以提高总的碳合成量, 从而保持较高生产力(Wright et al., 2017).叶片材料特性方面, 比叶质量随着环境温度的降低而增加, 干物质含量也有相同的趋势, 但是差异不显著.冷的地方, 细胞生长速率慢, 单位面积的小细胞数量增多, 叶片小而厚, 密度大(Atkins, 2005; Poorter et al., 2009).因此, 比叶质量越大的植物, 越偏向“慢速投资-收益”型.再者, 干物质含量增加有利于植物抵御霜冻和大风.因此, 寒冷和高海拔地区的植物一般具有更高的比叶质量(K?rner et al., 1986; Woodward, 1987; Atkins, 2005).干热河谷植物细胞生长速率快, 因而叶片较薄, 比叶质量小.叶片生理性状方面, 从干热河谷植物到亚热带常绿阔叶林植物, 随着环境温度的降低, 光合速率降低.除了温度外, 生活型对光合速率的影响也很大(刘长成等, 2011).本研究的结果也显示, 在每个样地内, 落叶植物的光合速率均显著高于常绿植物(图4).干热河谷和温带针阔混交林由于落叶植物和常绿植物共存, 且落叶植物占优势, 所以光合速率在样地内变异很大, 而且平均值较高. ... The “hydrology” of leaves: co-ordination of structure and function in temperate woody species 2 2003 ... 在物理性状方面, 叶片大小与叶片的边界层厚度正相关(Fetcher, 1981; Campbell & Norman, 2000).叶片与外界环境的热量和气体交换都要通过边界层, 边界层越厚, 交换效率越低.因此, 高温干旱地区的植物叶片面积较小, 可以更充分地与外界环境进行热量交换, 从而达到散热的目的(Geng et al., 2012; Pierce et al., 2012).但高温潮湿的地区, 叶片可以通过蒸腾散热, 叶片面积较大.低温地区的植物, 夜间叶片温度低于环境温度, 小叶片有利于从环境中吸收热量(McDonald et al., 2003; Peppe et al., 2011).研究表明, 夜间气温对叶片大小的影响贡献为51%, 昼间气温的贡献为38% (Wright et al., 2017).叶片颜色、反射率和吸收率影响叶片的辐射收支, 高温环境下, 减少叶片辐射吸收, 可以降低叶片温度; 而低温环境下, 增加叶片辐射吸收, 可以提高叶片温度.叶片材料特性包括含水量、密度和比叶质量等.叶片含水量高能够缓解叶片升温, 因此极端干热环境下, 肉质植物所占比例较高(Schymanski et al., 2013).植物叶片密度高, 非结构性碳以及半纤维素含量高, 有利于抵御寒冷和强风, 因此寒冷和高海拔地区植物的叶片密度较高(K?rner et al., 1986; Woodward, 1987; Atkins, 2005).叶片的解剖性状与叶片内的气体交换效率有关.叶片光合作用发生的场所——叶绿体主要是位于栅栏组织中, 栅栏组织有利于提高叶片的光合作用, 而海绵组织中有较大的细胞间隙便于气体运输(Kenzo et al., 2004).叶脉特征反映水分、养分和光合产物等物质运输的效率和安全性(Blonder et al., 2011; 李乐等, 2013).气孔是植物水分和CO2进出的门户, 决定了气体交换速率.气孔的结构特征可以反映植物的最大光合和蒸腾能力(Sack et al., 2003).在叶片生理性状方面, 叶片蒸腾是最有效的一种降温方式.由于水汽和CO2都从气孔进出, 因此它们之间是相互耦合的.目前, 主流的观点认为, 叶片的性状组合都是为最大化碳收益(Cordell et al., 2001).但也有学者认为, 极端高温环境下, 植物也可能优先蒸腾降温(Blonder & Michaletz, 2018).本研究假设干热和湿热环境下的植物会发育不同的热力性状来应对高温, 干热环境下的植物主要依靠物理降温, 湿热环境下的植物主要依靠蒸腾降温; 温湿度适宜地区的植物, 叶片性状优先满足生长, 热力性状不突出; 寒冷地区, 叶片热力性状有利于提高叶片温度. ...
本文的其它图/表
-
 表1
云南省具有明显水热梯度的样地基本信息
表1
云南省具有明显水热梯度的样地基本信息
-
表2
云南省具有明显水热梯度的4个样地的物种科属以及生活型信息
-
表3
叶片性状指标及其生态学意义
-
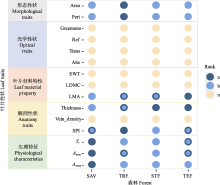 图1
云南省具有明显水热梯度的不同森林间叶片热力性状的差异。不同颜色代表差异显著, a、b, 代表从高到低排序; n, 代表4个森林间没有显著差异。SAV, 元江干热河谷植被; STF, 哀牢山亚热带常绿阔叶林; TEF, 丽江温带针阔混交林; TRF, 西双版纳热带雨林。Aarea, 单位面积最大净光合速率; Amass, 单位质量最大净光合速率; Abs, 叶片吸收率; Area, 单叶叶面积; EWT, 单位面积叶片含水量; Greenness, 绿度; LDMC, 叶干物质含量; LMA, 比叶质量; Peri, 单叶叶片周长; Ref, 叶片反射率; SPI, 气孔开口面积指数; Tr, 蒸腾速率; Thickness, 叶片厚度; Trans, 叶片透射率; Vein_density, 叶脉密度。
图1
云南省具有明显水热梯度的不同森林间叶片热力性状的差异。不同颜色代表差异显著, a、b, 代表从高到低排序; n, 代表4个森林间没有显著差异。SAV, 元江干热河谷植被; STF, 哀牢山亚热带常绿阔叶林; TEF, 丽江温带针阔混交林; TRF, 西双版纳热带雨林。Aarea, 单位面积最大净光合速率; Amass, 单位质量最大净光合速率; Abs, 叶片吸收率; Area, 单叶叶面积; EWT, 单位面积叶片含水量; Greenness, 绿度; LDMC, 叶干物质含量; LMA, 比叶质量; Peri, 单叶叶片周长; Ref, 叶片反射率; SPI, 气孔开口面积指数; Tr, 蒸腾速率; Thickness, 叶片厚度; Trans, 叶片透射率; Vein_density, 叶脉密度。
-
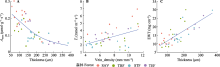 图2
云南省具有明显水热梯度的不同森林叶片性状间的相关关系。SAV, 元江干热河谷植被; STF, 哀牢山亚热带常绿阔叶林; TEF, 丽江温带针阔混交林; TRF, 西双版纳热带雨林。Amass, 单位质量最大净光合速率; EWT, 单位面积叶片含水量; Thickness, 叶片厚度; Tr, 蒸腾速率; Vein_density, 叶脉密度。
图2
云南省具有明显水热梯度的不同森林叶片性状间的相关关系。SAV, 元江干热河谷植被; STF, 哀牢山亚热带常绿阔叶林; TEF, 丽江温带针阔混交林; TRF, 西双版纳热带雨林。Amass, 单位质量最大净光合速率; EWT, 单位面积叶片含水量; Thickness, 叶片厚度; Tr, 蒸腾速率; Vein_density, 叶脉密度。
-
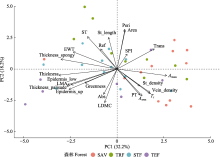 图3
云南省具有明显水热梯度的不同森林叶片性状的主成分(PC)分析。SAV, 元江干热河谷植被; STF, 哀牢山亚热带常绿阔叶林; TEF, 丽江温带针阔混交林; TRF, 西双版纳热带雨林。Aarea, 单位面积最大净光合速率; Amass, 单位质量最大净光合速率; Abs, 叶片吸收率; Area, 单叶叶面积; Epidermis_low, 下表皮厚度; Epidermis_up, 上表皮厚度; EWT, 单位面积叶片含水量; Greenness, 绿度; LDMC, 叶干物质含量; LMA, 比叶质量; Peri, 单叶叶片周长; PT, 栅栏组织与叶片厚度比值; Ref, 叶片反射率; SPI, 气孔开口面积指数; ST, 海绵组织与叶片厚度比值; St_density, 气孔密度; St_length, 气孔长度; Tr, 蒸腾速率; Thickness, 叶片厚度; Thickness_palisade, 栅栏组织厚度; Thickness_spongy, 海绵组织厚度; Trans, 叶片透射率; Vein_density, 叶脉密度。
图3
云南省具有明显水热梯度的不同森林叶片性状的主成分(PC)分析。SAV, 元江干热河谷植被; STF, 哀牢山亚热带常绿阔叶林; TEF, 丽江温带针阔混交林; TRF, 西双版纳热带雨林。Aarea, 单位面积最大净光合速率; Amass, 单位质量最大净光合速率; Abs, 叶片吸收率; Area, 单叶叶面积; Epidermis_low, 下表皮厚度; Epidermis_up, 上表皮厚度; EWT, 单位面积叶片含水量; Greenness, 绿度; LDMC, 叶干物质含量; LMA, 比叶质量; Peri, 单叶叶片周长; PT, 栅栏组织与叶片厚度比值; Ref, 叶片反射率; SPI, 气孔开口面积指数; ST, 海绵组织与叶片厚度比值; St_density, 气孔密度; St_length, 气孔长度; Tr, 蒸腾速率; Thickness, 叶片厚度; Thickness_palisade, 栅栏组织厚度; Thickness_spongy, 海绵组织厚度; Trans, 叶片透射率; Vein_density, 叶脉密度。
|

{kind=link}