



草地占我国国土面积的40%以上, 在维持生物多样性、应对气候变化与发展畜牧业等方面发挥着重要作用(陈佐忠和汪诗平, 2000; Craine et al., 2013)。近年来, 受人类过度开发利用和气候变化影响, 我国90%的草地发生不同程度的退化(Han et al., 2008)。封育作为一种经济有效的恢复技术, 被广泛用于我国退化草地的植被恢复管理(闫玉春等, 2009)。大量研究表明, 短期封育可有效提高草地植被盖度、多样性和生物量(Qiu et al., 2013; Jing et al., 2014), 并可改善土壤水分和养分状况(Jeddi & Chaieb, 2010; Cheng et al., 2016), 但也有研究表明长期封育可能会降低生物多样性和生产力, 削弱草地固碳能力, 影响生态系统功能的恢复及可持续发展(Hu et al., 2016; Deng et al., 2017)。然而, 上述研究结果的差异多源于草地恢复实验研究地点或恢复时长的不同, 因此有必要对同一地点的草地恢复过程展开长期监测, 以更好地研究草地群落生态系统功能在封育恢复演替过程中的动态变化。
生态系统功能的发挥依赖于生物多样性, 更高的生物多样性可以维持更高的生态系统功能水平(Risser, 1995; Grime, 1997)。生产力是生态系统功能的最主要体现形式, 物种多样性和功能多样性是生物多样性的两个重要维度。物种多样性通常用生态系统中植物、动物和微生物种类的丰富性表示, 功能多样性是指影响生态系统功能的群落内物种功能性状的大小、范围和分布(Dı́az & Cabido, 2001; Petchey & Gaston, 2002)。虽然生物多样性对生态系统功能和生态系统过程的重要性已被广泛认可和接受, 但研究学者以往常采用物种丰富度或物种多样性指标表征生物多样性, 忽视了功能多样性的作用(Dı́az & Cabido, 2001; Wacker et al., 2009; 马建军等, 2012; 王海东等, 2013)。例如, 目前大量有关生态系统生产力与物种多样性关系的研究显示, 两者间关系可表现为正相关(Cardinale et al., 2012)、负相关(Suding et al., 2005; Grace et al., 2016)、非线性关系(Fraser et al., 2015)或无显著相关性(Adler et al., 2011)。这暗示生态系统生产力不仅受物种多样性的影响, 还可能受物种组成和物种所具有的功能性状的影响(Dı́az & Cabido, 2001; Flynn et al., 2011)。
植物功能性状是指影响植物生存、生长和繁殖的适应性与效应特征的性状, 其可反映植物应对环境变化的策略(Carmona et al., 2011), 并能够影响生态系统功能(Balvanera et al., 2006; Violle et al., 2007), 已经成为阐明植物群落和生态系统响应环境变化过程的关键途径(McGill et al., 2006)。例如, 植物功能性状可通过影响植物光合和养分分配过程, 影响生态系统的生产力。其中, 较高的植株高度、比叶面积和叶片氮含量通常有助于提高草地生态系统的生产力(Golodets et al., 2009; Grigulis et al., 2013)。同时, 群落水平的功能性状不仅可用于表征群落动态变化特征, 还可指示生态系统功能的变化, 比物种丰富度能更好地解释草地群落生产力变化(Mouillot et al., 2011; Roscher et al., 2012)。此外, 草地生产力还受植物功能性状多样性(如叶片氮含量、比叶面积和叶干物质含量)的强烈影响(Klumpp & Soussana, 2009; Zuo et al., 2016)。
种间功能性状差异、物种组成及其相对多度是功能多样性的重要组成部分(Dı́az & Cabido, 2001; Lepš et al., 2006)。目前, 有关功能多样性影响生态系统功能的作用机制存在两种假说: 一是“质量比假说” (Grime, 1998), 该假说认为优势物种的功能性状决定了生态系统功能, 并可用群落加权平均值来表征功能多样性(Garnier et al., 2004); 二是“生态位互补假说” (Tilman et al., 1997), 即群落内物种间功能性状的差异使得物种间形成资源利用最大化策略, 并可用多种功能多样性指数来表征功能多样性。然而, 两种假说对生态系统功能变异的解释能力可能受外界环境条件变化的影响, 尚未得到一致结论(Mokany et al., 2008; Wacker et al., 2009; 吕亭亭等, 2014)。
此外, 功能多样性与物种多样性具有显著内在联系, 两者是群落内物种共存的关键因素(董世魁等, 2017), 共同决定着生态系统功能变化(Flynn et al., 2011)。然而由于受环境条件、物种组成、土地利用等因素的影响(Flynn et al., 2009; Cadotte et al., 2011), 目前有关物种多样性与功能多样性关系的认识并不一致(Petchey & Gaston, 2002; Reich et al., 2004; Sasaki et al., 2009), 同时针对两者关系的研究多集中于森林生态系统(刘晓娟和马克平, 2015), 草地生态系统研究相对较少。因此, 通过研究群落演替进程中的物种多样性、功能多样性与群落生产力的变化格局及关系, 可深入理解生物多样性与生态系统功能的关系及其环境调控机制, 为生物多样性保护、植被恢复与生态系统经营实践提供科学基础。
基于此背景, 本研究以不同封育年限草地群落为研究对象, 对其群落生产力、物种多样性和功能多样性展开研究, 拟回答以下两个科学问题: (1)在长期封育演替进程中, 草地群落生产力、物种多样性和功能多样性如何变化? (2)功能多样性对封育演替草地群落生产力变异的解释度是否超过物种多样性? 本研究旨在加深理解草地恢复过程中生物多样性与生态系统功能的关系, 以期为草地封育演替过程中的生物多样性和生态系统功能恢复提供依据。
1 材料和方法
1.1 研究区概况
研究区位于宁夏固原云雾山国家级自然保护区(36.22°-36.32° N, 106.40°-106.47° E, 海拔1 800- 2 100 m)。该区域属中温带半干旱气候, 年平均气温5 ℃, 年降水量400-450 mm, 全年65%-75%的降水集中在7-9月份。土壤类型主要包括山地灰褐土和黑垆土两类。研究区植被为温带典型草原, 群落建群种和优势种主要包括长芒草(Stipa bungeana)、百里香(Thymus mongolicus)、白莲蒿(Artemisia stechmanniana)、大针茅(Stipa grandis), 伴生种以猪毛蒿(Artemisia scoparis)和星毛委陵菜(Potentilla acaulis)为主(Jing et al., 2014)。该保护区始建于1982年, 自成立之初陆续实施了一系列草地生态恢复措施(如封育、补播等), 以恢复该区域内的退化草地群落。
1.2 研究方法
1.2.1 实验设计
为探究草地群落生产力和植物多样性在长期封育演替中的变化特征及生产力的驱动因子, 本研究采用“空间替代时间法”, 于2016年8月在该区域内选择土壤类型一致, 地形相似的连续放牧(封育0年)、封育9年、封育26年和封育34年的草地群落作为研究样地。因该区域地形以丘陵为主, 为减少各样地立地条件本身差异的干扰, 统一在各研究样地的阴坡、中上坡位置随机设置12个10 m × 10 m的小区, 小区间隔超过2 m。在每个小区内随机选取一个1 m × 1 m的小样方, 用于植物群落调查与植物功能性状取样测定。
1.2.2 指标测定
在2016年8月中下旬(植物生长旺盛时期), 对各小区1 m × 1 m样方内的物种组成、物种高度、多度和盖度进行测定。同时针对样方内出现的所有物种, 随机选取相应物种的10-20片健康成熟叶片, 放入自封袋并置于存有冰袋的储藏箱内带回实验室进行相关植物功能性状测定。本研究对7种植物功能性状展开测定: 植株高度(野外测定)、叶片厚度(LT)、比叶面积(SLA)、叶干物质含量(LDMC)、叶片碳含量(LCC)、叶片氮含量(LNC)和叶片磷含量(LPC)。在室内实验中, 首先对各样方内各物种的叶片进行扫描并用ImageJ软件获得叶片面积; 之后将叶片浸水至饱和后擦干称质量获得饱和鲜质量, 最后将叶片置于65 ℃烘箱烘干称质量获得叶片干质量。比叶面积指单位干质量的叶面积, 为叶片面积与叶片烘干质量的比值。叶干物质含量是指叶片的干质量与饱和鲜质量之比。将烘干的叶片粉碎后, 采用浓硫酸-重铬酸钾外加热法测定LCC, 采用半微量凯氏定氮法测定LNC, 采用酸溶-钼锑抗比色法测定LPC。
1.3 数据计算和统计分析
1.3.1 物种多样性指数
本研究选取4种常见物种多样性指数, 物种丰富度(S)、Shannon-Wiener多样性指数(H′)、Pielou均匀度指数(E)和Simpson优势度指数(D), 计算公式如下:
式中, pi为物种i的相对多度, N为物种数目。
1.3.2 植物功能性状群落加权平均值
本研究以各物种的相对多度为权重(Garnier et al., 2004), 计算7种植物功能性状的群落加权平均值(CWM), 计算公式如下:
式中, Traiti为物种i的性状值。
1.3.3 功能多样性指数
本研究选取FRic, FEve和FDis分别表征功能丰富度、均匀度和离散度。
FRic计算公式如下
式中, SFci为群落i内物种所占据的生态位空间, Rc为性状c的绝对值范围(Mason et al., 2005)。
FEve计算公式如下:
式中, bl为分支, PEWi为局部加权平均的均匀度, EWbl为加权平均的均匀度, pj为物种j的相对多度, dij为物种i与物种j间的欧式距离(Mouchet et al., 2010)。
FDis计算公式如下:
式中, g为加权重心, xik为物种i性状k的值, zi为物种i到g的加权距离(Laliberté & Legendre, 2010)。
1.3.4 统计分析
在统计分析前, 对数据进行正态性和方差齐次性检验, 对非正态性数据进行对数转换。借助F-diversity软件计算物种多样性和功能多样性指数。采用单因素方差分析和Duncan新复极差法检验地上生产力、物种多样性、植物功能性状及功能多样性在不同封育年限草地群落间的差异。采用回归分析和Pearson相关系数分析群落生产力、植物功能性状、物种多样性和功能多样性之间的关系。采用随机森林模型分析群落加权平均功能性状、4种物种多样性指数和3种功能多样性指数对群落地上生产力影响的大小, 并进一步采用方差分解方法分析3种指标体系(植物功能性状、物种多样性和功能多样性)对群落地上生产力变异的解释度。随机森林模型借助R语言“randomForest”包实现, 并利用其“rfPermute”程序包对各因子重要性的显著水平进行评估。方差分解基于“vegan”包的“varpart”功能来实现。上述统计检验的显著水平均设为p < 0.05。
2 结果和分析
2.1 封育草地群落地上生产力和物种多样性
封育显著提高草地群落的植物地上生产力和凋落物生物量, 但对物种多样性影响较小(表1)。具体而言, 不同封育年限草地群落地上生产力为: 封育34年≈封育26年>封育9年>连续放牧草地; 凋落物生物量为: 封育34年>封育26年>封育9年≈连续放牧草地。此外, 虽然封育未显著改变草地群落的Shannon-Wiener指数, 但封育34年的值显著低于封育26年。类似地, 草地群落的Pielou均匀度指数和Simpson优势度指数在连续放牧草地、封育9年和26年草地间无显著差异, 但其均在封育34年后显著下降。相比之下, 不同封育年限草地群落的物种丰富度无显著差异。
表1 封育草地群落植物地上生产力与物种多样性(平均值±标准误)
Table 1
| 封育年限 GED (a) | 纬度 Latitude (° N) | 经度 Longitude (° E) | 海拔 Altitude (m) | 地上生产力 ANPP (g·m-2) | 凋落物生物量 Litter biomass (g·m-2) | 丰富度 Richness | Shannon- Wiener 指数 H′ | Pielou均匀度指数 E | Simpson优势度指数 D |
|---|---|---|---|---|---|---|---|---|---|
| 0 | 36.22 | 106.38 | 1 942 | 115.60 ± 13.13c | 30.78 ± 8.02c | 13.33 ± 0.74a | 2.21 ± 0.05ab | 0.86 ± 0.01a | 0.85 ± 0.01a |
| 9 | 36.22 | 106.39 | 1 928 | 165.14 ± 10.10b | 61.66 ± 7.38bc | 12.64 ± 0.62a | 2.17 ± 0.07ab | 0.86 ± 0.01a | 0.85 ± 0.01a |
| 26 | 36.20 | 106.41 | 1 885 | 253.38 ± 15.32a | 160.06 ± 34.83b | 14.17 ± 0.67a | 2.30 ± 0.05a | 0.87 ± 0.01a | 0.87 ± 0.01a |
| 34 | 36.24 | 106.38 | 2 098 | 261.79 ± 19.87a | 346.74 ± 57.52a | 14.42 ± 0.96a | 2.11 ± 0.06b | 0.80 ± 0.01b | 0.81 ± 0.01b |
同列不同小写字母表示差异显著(p < 0.05)。
Different lowercase letters indicate significant differences in the same column (p < 0.05). ANPP, aboveground net primary productivity; D, Simpson dominance index; E, Pielou evenness index; GED, grazing exclusion duration; H′, Shannon-Wiener index.
2.2 封育草地植物群落加权平均功能性状和功能多样性
在草地长期封育演替进程中, 不同群落加权平均功能性状的变化特征存在差异(图1)。具体而言, 群落加权平均植株高度、叶片厚度和叶片碳含量均随封育年限增加而增加, 群落加权平均叶片氮和磷含量无显著变化。此外, 与持续放牧草地相比, 封育9年草地的群落加权平均叶干物质含量显著增加, 但封育9年和34年草地的群落加权平均比叶面积显著减小。另一方面, 长期封育(封育26年和34年)显著增加草地群落的功能丰富度和离散度, 但封育未显著影响草地群落的功能均匀度。
图1
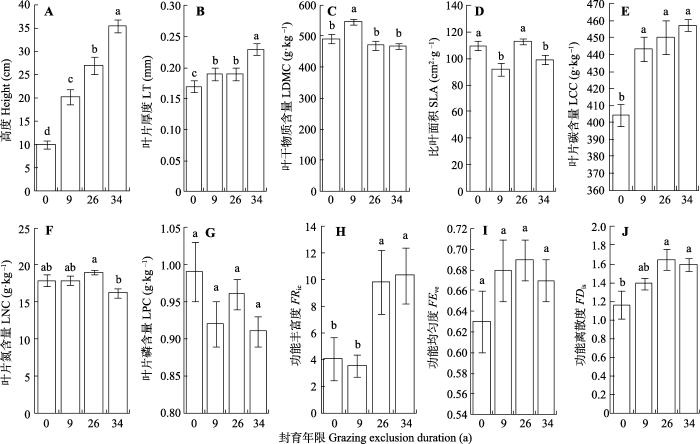
图1
封育草地植物群落加权平均功能性状和功能多样性(平均值±标准误)。不同小写字母表示差异显著(p < 0.05)。
Fig. 1
Community weighted mean functional traits and functional diversity of grassland communities after grazing exclusion (mean ± SE). Different lowercase letters indicate significant differences (p < 0.05). FDis, functional dispersion; FEve, functional evenness; FRic, functional richness; LCC, leaf carbon concentration; LDMC, leaf dry matter content; LNC, leaf nitrogen concentration; LPC, leaf phosphorus concentration; LT, leaf thickness; SLA, specific leaf area.
2.3 封育草地植物群落生产力与物种多样性、功能多样性关系
图2
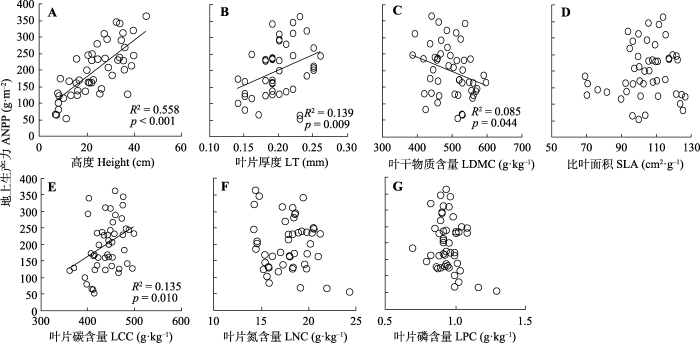
图2
封育草地群落地上生产力与植物群落加权平均功能性状的关系。
Fig. 2
Relationships between aboveground net primary productivity (ANPP) and community-weighted mean functional traits in grasslands after grazing exclusion. LCC, leaf carbon concentration; LDMC, leaf dry matter content; LNC, leaf nitrogen concentration; LPC, leaf phosphorus concentration; LT, leaf thickness; SLA, specific leaf area.
图3
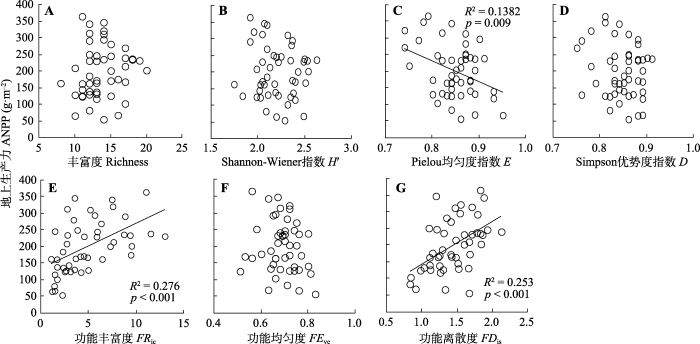
图3
封育草地群落地上生产力与物种多样性(A-D)和功能多样性(E-G)的关系。
Fig. 3
Relationship between aboveground net primary productivity (ANPP) and plant species diversity (A-D) and functional diversity (E-G) in grassland communities after grazing exclusion. D, Simpson dominance index; E, Pielou evenness index; FDis, functional dispersion; FEve, functional evenness; FRic, functional richness; H′, Shannon-Wiener index.
图4
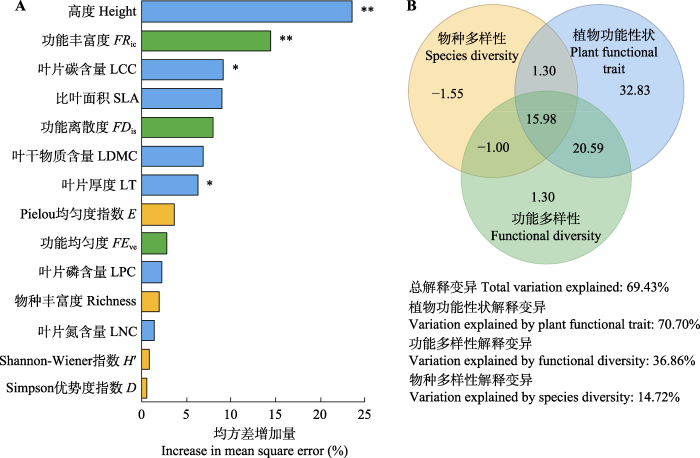
图4
封育草地群落地上生产力影响因子的随机森林模型(A)和方差分解(B)结果。*, p < 0.05; **, p < 0.01。
Fig. 4
Results of random forest model (A) and variance partition analysis (B) for factors influencing aboveground net primary productivity of grassland communities after grazing exclusion. *, p < 0.05; **, p < 0.0l. D, Simpson dominance index; E, Pielou evenness index; FDis, functional dispersion; FEve, functional evenness; FRic, functional richness; H′, Shannon-Wiener index; LCC, leaf carbon concentration; LDMC, leaf dry matter content; LNC, leaf nitrogen concentration; LPC, leaf phosphorus concentration; LT, leaf thickness; SLA, specific leaf area.
Pearson相关分析结果显示, 不同群落加权平均功能性状、功能多样性与物种多样性之间的相关关系存在差异(表2)。其中, 群落加权平均高度与Pielou均匀度指数、Simpson优势度指数均呈显著负相关关系, 同时群落加权平均叶干物质含量与物种丰富度呈显著负相关关系, 群落加权平均比叶面积与物种丰富度、Shannon-Wiener指数均存在显著正相关关系, 群落加权平均叶片氮和磷含量则均与Shannon-Wiener指数、Pielou均匀度指数和Simpson优势度指数存在显著正相关关系。植物群落功能丰富度与物种丰富度呈显著正相关关系, 同时群落功能均匀度与Shannon-Wiener指数呈显著正相关关系。相比之下, 群落加权平均叶片厚度和C含量以及功能离散度均与物种多样性指数无显著相关关系。
表2 封育草地植物群落加权平均功能性状、功能多样性与物种多样性的Pearson相关系数
Table 2
| 高度 Height | 叶片厚度 LT | 叶干物质 含量 LDMC | 比叶面积 SLA | 叶片碳含量 LCC | 叶片氮含量 LNC | 叶片磷含量 LPC | 功能丰富度 FRic | 功能均匀度 FEve | 功能离散度 FDis | |
|---|---|---|---|---|---|---|---|---|---|---|
| 物种丰富度 Richness | 0.106 | 0.014 | -0.347* | 0.301* | -0.077 | 0.133 | 0.145 | 0.551** | 0.199 | 0.206 |
| Shannon-Wiener指数 H′ | -0.232 | -0.188 | -0.233 | 0.327* | -0.145 | 0.369** | 0.347* | 0.284 | 0.294* | 0.100 |
| Pielou均匀度指数 E | -0.502** | -0.280 | 0.108 | 0.082 | -0.120 | 0.416** | 0.364* | -0.243 | 0.225 | -0.094 |
| Simpson优势度指数 D | -0.376** | -0.263 | -0.043 | 0.266 | -0.104 | 0.414** | 0.360* | 0.037 | 0.181 | 0.048 |
*, p < 0.05; **, p < 0.01. D, Simpson dominance index; E, Pielou evenness index; FDis, functional dispersion; FEve, functional evenness; FRic, functional richness; H′, Shannon-Wiener index; LCC, leaf carbon concentration; LDMC, leaf dry matter content; LNC, leaf nitrogen concentration; LPC, leaf phosphorus concentration; LT, leaf thickness; SLA, specific leaf area.
3 讨论
与前人研究(Jing et al., 2013; Alberti et al., 2017)类似, 本研究发现草地群落地上生产力和凋落物生物量随着封育年限增加而增加, 主要是因为围封直接减少了牛、羊等食草动物的采食。最新研究表明, 封育后草地生产力和凋落物生物量的提高主要源于禾草类植物生物量的增加(Su & Xu, 2021), 因为食草动物可能更偏爱取食禾草类植物(Dı́az et al., 2007), 同时禾草凋落物分解过程较为缓慢, 进一步促使地表凋落物累积。封育通常可以提高植物多样性, 尤其是对于水分受限的生态系统而言(Fernández-Lugo et al., 2013; Cheng et al., 2016)。然而本研究发现物种丰富度在长期封育过程中无明显变化, 推测是本研究区草地群落内的杂类草功能群的生物量相对稳定所致, 因为杂类草植物是构成草地群落物种库的主体(Li et al., 2017)。类似地, 针对我国草地封育研究的整合分析结果表明, 封育整体上未显著改变草地群落的物种多样性(Xiong et al., 2016)。此外, 本研究发现, 相比于封育26年的草地, 封育34年的草地群落的Shannon-Wiener指数和Pielou均匀度指数显著降低, 凋落物生物量显著增加, 高寒草甸也有类似发现(刘晓琴等, 2016)。这暗示长期封育后地表过量累积的凋落物可能已开始对草地植物群落演替产生不利影响。因此, 建议对长期封育草地群落进行适度利用(如轻中度放牧、刈割等), 以刺激草地群落更新, 维持和提高生物多样性和生态系统功能。
在草地封育演替进程中, 土壤水分、温度和养分等外界环境条件会同步发生变化(Qiu et al., 2013; Jing et al., 2014)。环境变化不仅可直接影响个体物种功能性状, 还可通过影响群落物种组成或多度, 进而引起群落水平功能性状的变化(Walker et al., 1999)。本研究发现不同群落加权平均功能性状在封育演替过程中的变化特征存在差异。例如, 群落加权平均高度和叶片碳含量沿封育演替进程不断增加, 表明草地群落的光获取能力与光合能力不断增强, 这可能与土壤水分和养分的增加密不可分。然而, 在封育演替中后期, 群落加权平均叶片厚度增加, 比叶面积减小, 暗示植物群落开始趋于资源保守利用策略。由此可见, 草地群落在封育演替早期处于相对开放的环境, 植物功能性状多表现为资源获取型策略, 但随着演替进行, 凋落物的不断累积导致环境更加郁闭, 植物功能性状开始趋向资源保守型策略(Zhang et al., 2018)。功能丰富度和功能离散度表征群落中植物功能性状的类型和分布范围(Villéger et al., 2008), 并反映群落内物种生态位分化程度(Mason et al., 2005)。本研究发现, 封育中后期草地群落功能丰富度和离散度显著增加, 表明此时群落内物种占据的生态位空间更大, 各物种生态位分化程度更高, 对资源利用更加充分, 种间资源竞争强度较弱(Petchey & Gaston, 2006), 这也为该阶段保守型的植物功能性状提供了支持。
类似前人研究(Lanta & Lepš, 2006; Adler et al., 2011; Roscher et al., 2012), 我们发现封育草地群落的地上生产力与物种多样性之间无明显关系, 功能多样性比物种多样性更能决定群落生产力。这表明该区域草地群落生产力主要受物种属性而非物种丰富度的影响, 因为功能多样性能够综合不同物种的属性, 可更好表征生物从环境中利用资源的能力(Petchey & Gaston, 2006)。相比之下, 物种数量和物种多样性包含较少的物种特征信息, 忽略了物种间功能性状的相似性和差异性。群落加权平均值可用于评估优势性状对生产力的影响, 而功能多样性指数则可量化该性状的变异对生产力的影响(Villéger et al., 2008)。方差分解结果表明, 植物功能性状群落加权平均值对草地群落生产力变异的解释程度高于功能多样性指数和物种多样性指数。这说明该群落内优势物种对生物量的影响作用更大, 生态系统功能可能主要取决于群落中优势物种的功能性状。具体而言, 植株高度是影响草地群落生产力的最主要因子, 这点在前人研究中亦有所体现(Golodets et al., 2009; Grigulis et al., 2013)。由此可见, 在本研究中, “质量比假说”相比于“生态位互补假说”, 能更好地解释功能多样性对该区域封育演替草地群落生产力变化的贡献。本研究还发现功能多样性与物种多样性之间存在紧密联系(Dı́az & Cabido, 2001; Naeem, 2002; 董世魁等, 2017)。这是由于群落内各物种具有特定的功能特征, 物种多样性的增加会加大群落内植物功能性状的变化范围, 使得功能多样性与物种多样性呈现正相关关系(Dı́az & Cabido, 2001; Sasaki et al., 2009)。反之, 物种多样性下降或物种丧失会引起功能性状丢失或性状空间维度减小, 进而导致功能多样性的下降。
通常而言, 一种植物功能性状可能影响多种生态系统功能, 同一种生态系统功能可能同时受到多种植物功能性状的共同作用(潘权等, 2021), 此外不同功能性状之间存在内在联系, 因此未来研究应关注多种功能性状对生态系统服务的影响。值得一提的是, 最新提出的“植物功能性状网络”概念, 可为探究植物功能性状相关性对生态系统功能的影响提供重要思路(He et al., 2020)。由于生态系统同时提供多种功能与服务(如生产力、养分循环、资源获取等), 即生态系统多功能性, 而且不同功能间可能存在协同、权衡与兼容等关系。只分析生物多样性与单一生态系统功能(生产力)之间的关系, 可能会低估生物多样性的重要性及其丧失对生态系统带来的不利影响(Wagg et al., 2014)。此外, 全球变化(如升温、CO2浓度升高和氮沉降等)与土地利用方式变化同步发生, 其可能通过改变植物功能性状和群落组成(Suding et al., 2005), 影响生态系统过程和功能, 进而调节土地利用方式变化背景下生物多样性与生态系统功能的关系(刘晓娟和马克平, 2015)。
综上可知, 在长达34年的封育演替过程中, 本研究区域典型草原的地上生产力、功能丰富度和功能离散度显著增加, 物种多样性和功能均匀度无明显变化, 群落水平不同植物功能性状的变化特征存在差异。功能多样性相比物种多样性能够更好地解释封育演替草地群落生产力的变化, 且“质量比假说”是功能多样性影响生产力的主要作用机制。由于物种多样性和功能多样性紧密相关, 二者不能相互替代, 未来研究应在全球变化背景下, 从功能多样性和物种多样性两个维度研究其与生态系统多功能性的关系, 来更加准确和全面地研究植物群落在恢复演替进程中的生态功能动态, 以实现更好的生态恢复效果(Cadotte et al., 2011)。
特邀编委: 汪诗平 编辑: 乔鲜果
致谢
感谢西北农林科技大学草业与草原学院赵婷、赵伟和程帅在野外采样工作和室内分析中给予的帮助。
参考文献
Productivity is a poor predictor of plant species richness
DOI:10.1126/science.1204498 URL [本文引用: 2]
Herbivore exclusion promotes a more stochastic plant community assembly in a natural grassland
DOI:10.1002/ecy.1741
PMID:28112395
[本文引用: 1]

Both bottom-up (e.g., nutrients) and top-down (e.g., herbivory) forces structure plant communities, but it remains unclear how they affect the relative importance of stochastic and deterministic processes in plant community assembly. Moreover, different-sized herbivores have been shown to have contrasting effects on community structure and function, but their effects on the processes governing community assembly (i.e., how they generate the impacts on structure) remain largely unknown. We evaluated the influence of bottom-up and top-down forces on the relative importance of deterministic and stochastic processes during plant community assembly. We used the data of a 7-yr factorial experiment manipulating nutrient availability (ambient and increased) and the presence of vertebrate herbivores (>1 kg) of different body size in a floodplain grassland in The Netherlands. We used a null model that describes a community composition expected by chance (i.e., stochastic assembly) and compared the plant community composition in the different treatments with this null model (the larger the difference, the more deterministically assembled). Our results showed that herbivore exclusion promoted a more stochastic plant community assembly, whereas increased nutrients played a relatively minor role in determining the relative importance of stochasticity in community assembly. Large herbivores facilitated intermediate-sized mammal herbivores, resulting in synergistic effects of enhanced grazing pressure and a more deterministic and convergent plant community assembly. We conclude that herbivores can act as strong deterministic forces during community assembly in natural systems. Our results also reveal that although large- and intermediate-sized mammal herbivores often have contrasting effects on many community and ecosystem properties, they can also synergistically homogenize plant communities.© 2017 by the Ecological Society of America.
Quantifying the evidence for biodiversity effects on ecosystem functioning and services
Concern is growing about the consequences of biodiversity loss for ecosystem functioning, for the provision of ecosystem services, and for human well being. Experimental evidence for a relationship between biodiversity and ecosystem process rates is compelling, but the issue remains contentious. Here, we present the first rigorous quantitative assessment of this relationship through meta-analysis of experimental work spanning 50 years to June 2004. We analysed 446 measures of biodiversity effects (252 in grasslands), 319 of which involved primary producer manipulations or measurements. Our analyses show that: biodiversity effects are weaker if biodiversity manipulations are less well controlled; effects of biodiversity change on processes are weaker at the ecosystem compared with the community level and are negative at the population level; productivity-related effects decline with increasing number of trophic links between those elements manipulated and those measured; biodiversity effects on stability measures ('insurance' effects) are not stronger than biodiversity effects on performance measures. For those ecosystem services which could be assessed here, there is clear evidence that biodiversity has positive effects on most. Whilst such patterns should be further confirmed, a precautionary approach to biodiversity management would seem prudent in the meantime.
Beyond species: functional diversity and the maintenance of ecological processes and services
DOI:10.1111/j.1365-2664.2011.02048.x URL [本文引用: 2]
Biodiversity loss and its impact on humanity
Plant traits that predict resistance to herbivores
DOI:10.1111/j.1365-2435.2010.01794.x URL [本文引用: 1]
Long-term grazing exclusion effects on vegetation characteristics, soil properties and bacterial communities in the semi-arid grasslands of China
DOI:10.1016/j.ecoleng.2016.09.003 URL [本文引用: 2]
Global diversity of drought tolerance and grassland climate-change resilience
DOI:10.1038/nclimate1634 URL [本文引用: 1]
Effects of grazing exclusion on carbon sequestration in China’s grassland
DOI:10.1016/j.earscirev.2017.08.008 URL [本文引用: 1]
Vive la différence: plant functional diversity matters to ecosystem processes
DOI:10.1016/S0169-5347(01)02283-2 URL [本文引用: 6]
Plant trait responses to grazing-A global synthesis
DOI:10.1111/j.1365-2486.2006.01288.x URL [本文引用: 1]
Relationship between plant species diversity and functional diversity in alpine grasslands
高寒草地植物物种多样性与功能多样性的关系
Productivity: key factor affecting grazing exclusion effects on vegetation and soil
DOI:10.1007/s11258-013-0196-8 URL [本文引用: 1]
Loss of functional diversity under land use intensification across multiple taxa
DOI:10.1111/j.1461-0248.2008.01255.x
PMID:19087109
[本文引用: 1]
Land use intensification can greatly reduce species richness and ecosystem functioning. However, species richness determines ecosystem functioning through the diversity and values of traits of species present. Here, we analyze changes in species richness and functional diversity (FD) at varying agricultural land use intensity levels. We test hypotheses of FD responses to land use intensification in plant, bird, and mammal communities using trait data compiled for 1600+ species. To isolate changes in FD from changes in species richness we compare the FD of communities to the null expectations of FD values. In over one-quarter of the bird and mammal communities impacted by agriculture, declines in FD were steeper than predicted by species number. In plant communities, changes in FD were indistinguishable from changes in species richness. Land use intensification can reduce the functional diversity of animal communities beyond changes in species richness alone, potentially imperiling provisioning of ecosystem services.
Functional and phylogenetic diversity as predictors of biodiversity-ecosystem-function relationships
How closely does variability in ecologically important traits reflect evolutionary divergence? The use of phylogenetic diversity (PD) to predict biodiversity effects on ecosystem functioning, and more generally the use of phylogenetic information in community ecology, depends in part on the answer to this question. However, comparisons of the predictive power of phylogenetic diversity and functional diversity (FD) have not been conducted across a range of experiments. To address how phylogenetic diversity and functional trait variation control biodiversity effects on biomass production, we summarized the results of 29 grassland plant experiments where both the phylogeny of plant species used in the experiments is well described and where extensive trait data are available. Functional trait variation was only partially related to phylogenetic distances between species, and the resulting FD values therefore correlate only partially with PD. Despite these differences, FD and PD predicted biodiversity effects across all experiments with similar strength, including in subsets that excluded plots with legumes and that focused on fertilization experiments. Two- and three-trait combinations of the five traits used here (leaf nitrogen percentage, height, specific root length, leaf mass per unit area, and nitrogen fixation) resulted in the FD values with the greatest predictive power. Both PD and FD can be valuable predictors of the effect of biodiversity on ecosystem functioning, which suggests that a focus on both community trait diversity and evolutionary history can improve understanding of the consequences of biodiversity loss.
Worldwide evidence of a unimodal relationship between productivity and plant species richness
DOI:10.1126/science.aab3916 URL [本文引用: 1]
Plant functional markers capture ecosystem properties during secondary succession
DOI:10.1890/03-0799 URL [本文引用: 2]
A community-level test of the leaf-height-seed ecology strategy scheme in relation to grazing conditions
DOI:10.1111/j.1654-1103.2009.01071.x URL [本文引用: 2]
Integrative modelling reveals mechanisms linking productivity and plant species richness
DOI:10.1038/nature16524 URL [本文引用: 1]
Relative contributions of plant traits and soil microbial properties to mountain grassland ecosystem services
DOI:10.1111/1365-2745.12014 URL [本文引用: 2]
Biodiversity and ecosystem function: the debate deepens
DOI:10.1126/science.277.5330.1260 URL [本文引用: 1]
Benefits of plant diversity to ecosystems: immediate, filter and founder effects
DOI:10.1046/j.1365-2745.1998.00306.x URL [本文引用: 1]
Rangeland degradation and restoration management in China
DOI:10.1071/RJ08009 URL [本文引用: 1]
Plant trait networks: improved resolution of the dimensionality of adaptation
DOI:10.1016/j.tree.2020.06.003 URL [本文引用: 1]
A synthesis of the effect of grazing exclusion on carbon dynamics in grasslands in China
DOI:10.1111/gcb.13133 URL [本文引用: 1]
Changes in soil properties and vegetation following livestock grazing exclusion in degraded arid environments of South Tunisia
DOI:10.1016/j.flora.2009.03.002 URL [本文引用: 1]
Assessment of vegetative ecological characteristics and the succession process during three decades of grazing exclusion in a continental steppe grassland
DOI:10.1016/j.ecoleng.2013.04.035 URL [本文引用: 1]
Changes in plant community composition and soil properties under 3-decade grazing exclusion in semiarid grassland
DOI:10.1016/j.ecoleng.2013.12.023 URL [本文引用: 3]
Using functional traits to predict grassland ecosystem change: a mathematical test of the response-and-effect trait approach
DOI:10.1111/j.1365-2486.2009.01905.x URL [本文引用: 1]
A distance-based framework for measuring functional diversity from multiple traits
A new framework for measuring functional diversity (FD) from multiple traits has recently been proposed. This framework was mostly limited to quantitative traits without missing values and to situations in which there are more species than traits, although the authors had suggested a way to extend their framework to other trait types. The main purpose of this note is to further develop this suggestion. We describe a highly flexible distance-based framework to measure different facets of FD in multidimensional trait space from any distance or dissimilarity measure, any number of traits, and from different trait types (i.e., quantitative, semi-quantitative, and qualitative). This new approach allows for missing trait values and the weighting of individual traits. We also present a new multidimensional FD index, called functional dispersion (FDis), which is closely related to Rao's quadratic entropy. FDis is the multivariate analogue of the weighted mean absolute deviation (MAD), in which the weights are species relative abundances. For unweighted presence-absence data, FDis can be used for a formal statistical test of differences in FD. We provide the "FD" R language package to easily implement our distance-based FD framework.
Effect of functional group richness and species richness in manipulated productivity-diversity studies: a glasshouse pot experiment
DOI:10.1016/j.actao.2005.08.003 URL [本文引用: 1]
Quantifying and interpreting functional diversity of natural communities: practical considerations matter
Patterns and thresholds of grazing-induced changes in community structure and ecosystem functioning: species-level responses and the critical role of species traits
DOI:10.1111/1365-2664.12806 URL [本文引用: 1]
Plant functional traits-Concepts, applications and future directions
植物功能性状研究进展
Effects of exclosure duration on the community structure and species diversity of an alpine meadow in the Qinghai-Tibet Plateau
封育年限对高寒草甸群落组分和物种多样性的影响
Relationship between functional diversity and productivity in meadow and marsh plant communities
DOI:10.3724/SP.J.1258.2014.00037 URL [本文引用: 1]
草甸和沼泽植物群落功能多样性与生产力的关系
DOI:10.3724/SP.J.1258.2014.00037
[本文引用: 1]
功能多样性-生产力关系研究结果支持质量比假说和多样性假说, 但对于这两种假说的适用条件尚有争议。通过对吉林省西部草甸和沼泽植物群落的地上生物量、2个物种多样性指标(物种丰富度和Shannon-Weaver指数)、7种植物性状的两类功能多样性指标(群落权重均值和Rao二次熵), 以及土壤环境因子进行调查测量, 研究了群落功能多样性与生产力的关系。结果表明: 1)功能多样性与生产力的关系比物种多样性与生产力的关系更为密切; 2)功能群落权重均值解释生产力变异的能力好于Rao二次熵, 即优势物种对群落生产力的影响作用更大; 3)水淹条件影响着功能多样性与生产力的关系, 以群落权重均值为基础的质量比假说适于解释草甸群落功能多样性与生产力的关系, 而以Rao二次熵为基础的多样性假说适于解释有强烈环境筛(水淹)的沼泽群落功能多样性与生产力的关系。
Changes in plant functional groups and species diversity under three grassland using modes in typical grassland area of Inner Mongolia, China
DOI:10.3724/SP.J.1258.2012.00001 URL [本文引用: 1]
内蒙古典型草原区3种不同草地利用模式下植物功能群及其多样性的变化
DOI:10.3724/SP.J.1258.2012.00001
[本文引用: 1]
以呼伦贝尔草原辉河国家级自然保护区草原群落为研究对象, 选取3块生境条件基本相同的样地(250 m × 250 m) ——放牧场、割草场及公共草场, 每块样地设置3条样带, 每条样带上设置8个样方(1 m × 1 m), 研究了不同草地利用模式下草原群落的物种组成、生物多样性及群落的功能群特征。结果表明: 在放牧场、割草场和公共草场3种不同利用模式下, 植物功能群结构及其多样性发生了变化。灌木、半灌木的累积优势度在各利用模式间差异显著(p p p p 增加, 使得草地植物群落结构简单、脆弱。丰富度指数和多样性指数在放牧场最低, 在割草场最高; 均匀度变化并不明显。割草场利用模式有助于改善草原的健康状况, 而放牧场及公共草场利用模式均存在不同程度的过度放牧现象, 尤其是公共草场利用模式由于所有权不明确而缺乏有效的管理, 导致草原退化明显。
Functional richness, functional evenness and functional divergence: the primary components of functional diversity
DOI:10.1111/j.0030-1299.2005.13886.x URL [本文引用: 2]
Rebuilding community ecology from functional traits
DOI:10.1016/j.tree.2006.02.002 URL [本文引用: 1]
Functional identity is more important than diversity in influencing ecosystem processes in a temperate native grassland
DOI:10.1111/j.1365-2745.2008.01395.x URL [本文引用: 1]
Functional diversity measures: an overview of their redundancy and their ability to discriminate community assembly rules
DOI:10.1111/j.1365-2435.2010.01695.x URL [本文引用: 1]
Functional structure of biological communities predicts ecosystem multifunctionality
DOI:10.1371/journal.pone.0017476 URL [本文引用: 1]
Disentangling the impacts of diversity on ecosystem functioning in combinatorial experiments
DOI:10.1890/0012-9658(2002)083[2925:DTIODO]2.0.CO;2 URL [本文引用: 1]
Effects of plant functional traits on ecosystem services: a review
DOI:10.17521/cjpe.2020.0142 URL [本文引用: 1]
植物功能性状对生态系统服务影响研究进展
DOI:10.17521/cjpe.2020.0142
[本文引用: 1]
全面认识和理解生态系统服务的形成机制是维持其持续供给的前提。植物功能性状直接参与多种生态系统过程, 影响生态系统服务供给, 探讨植物功能性状与生态系统服务的关系是揭示生态系统服务形成机制的重要途径。该文采用系统的文献综述方法, 分析了植物功能性状与生态系统服务关系的研究特点, 总结了影响不同生态系统服务的主要植物功能性状, 阐述了可能的影响途径。结果表明: 植物功能性状与生态系统服务关系研究以草地和森林等自然生态系统为主; 大部分研究集中在生态系统供给服务和支持服务, 包括生物量、净初级生产力、土壤肥力等; 根据植物功能性状对不同生态系统服务的影响程度, 植物功能性状可以聚类为土壤保持服务相关性状、水分循环相关性状、多功能相关性状、产品提供服务与养分循环相关性状以及授粉与生物控制服务相关性状; 并阐述了植物功能性状指标影响不同的生态系统服务途径。围绕植物功能性状对生态系统服务的影响, 今后尚需进一步探讨生态系统多功能性、植物功能性状相关性、气候变化和人类活动不确定性、时空尺度差异等因素对二者关系的影响。
Functional diversity (FD), species richness and community composition
DOI:10.1046/j.1461-0248.2002.00339.x URL [本文引用: 2]
Functional diversity: back to basics and looking forward
Functional diversity is a component of biodiversity that generally concerns the range of things that organisms do in communities and ecosystems. Here, we review how functional diversity can explain and predict the impact of organisms on ecosystems and thereby provide a mechanistic link between the two. Critical points in developing predictive measures of functional diversity are the choice of functional traits with which organisms are distinguished, how the diversity of that trait information is summarized into a measure of functional diversity, and that the measures of functional diversity are validated through quantitative analyses and experimental tests. There is a vast amount of trait information available for plant species and a substantial amount for animals. Choosing which traits to include in a particular measure of functional diversity will depend on the specific aims of a particular study. Quantitative methods for choosing traits and for assigning weighting to traits are being developed, but need much more work before we can be confident about trait choice. The number of ways of measuring functional diversity is growing rapidly. We divide them into four main groups. The first, the number of functional groups or types, has significant problems and researchers are more frequently using measures that do not require species to be grouped. Of these, some measure diversity by summarizing distances between species in trait space, some by estimating the size of the dendrogram required to describe the difference, and some include information about species' abundances. We show some new and important differences between these, as well as what they indicate about the responses of assemblages to loss of individuals. There is good experimental and analytical evidence that functional diversity can provide a link between organisms and ecosystems but greater validation of measures is required. We suggest that non-significant results have a range of alternate explanations that do not necessarily contradict positive effects of functional diversity. Finally, we suggest areas for development of techniques used to measure functional diversity, highlight some exciting questions that are being addressed using ideas about functional diversity, and suggest some directions for novel research.
Ecosystem carbon and nitrogen accumulation after grazing exclusion in semiarid grassland
DOI:10.1371/journal.pone.0055433 URL [本文引用: 2]
Species and functional group diversity independently influence biomass accumulation and its response to CO2 and N.
Biodiversity and ecosystem function
DOI:10.1046/j.1523-1739.1995.09040742.x URL [本文引用: 1]
Using plant functional traits to explain diversity-productivity relationships
DOI:10.1371/journal.pone.0036760 URL [本文引用: 2]
Two-phase functional redundancy in plant communities along a grazing gradient in Mongolian rangelands
DOI:10.1890/08-1850.1 URL [本文引用: 2]
Root, not aboveground litter, controls soil carbon storage under grazing exclusion across grasslands worldwide
DOI:10.1002/ldr.4008 URL [本文引用: 1]
Functional- and abundance-based mechanisms explain diversity loss due to N fertilization
Plant diversity and ecosystem productivity: theoretical considerations
Ecosystem processes are thought to depend on both the number and identity of the species present in an ecosystem, but mathematical theory predicting this has been lacking. Here we present three simple models of interspecific competitive interactions in communities containing various numbers of randomly chosen species. All three models predict that, on average, productivity increases asymptotically with the original biodiversity of a community. The two models that address plant nutrient competition also predict that ecosystem nutrient retention increases with biodiversity and that the effects of biodiversity on productivity and nutrient retention increase with interspecific differences in resource requirements. All three models show that both species identity and biodiversity simultaneously influence ecosystem functioning, but their relative importance varies greatly among the models. This theory reinforces recent experimental results and shows that effects of biodiversity on ecosystem functioning are predicted by well-known ecological processes.
New multidimensional functional diversity indices for a multifaceted framework in functional ecology
DOI:10.1890/07-1206.1 URL [本文引用: 2]
Let the concept of trait be functional
DOI:10.1111/j.0030-1299.2007.15559.x URL [本文引用: 1]
Diversity effects in early- and mid-successional species pools along a nitrogen gradient
Biodiversity experiments with grassland species have shown that plant productivity commonly increases with species richness and that this increase can be stronger in nutrient-rich than in nutrient-poor environments. It has been suggested that these effects are due to functional diversity among species. To investigate this, we established five early- and five mid-successional pools of six species each, expecting stronger effects in the latter. The 10 six-species mixtures and the 60 corresponding species monocultures were grown at four levels of soil nitrogen. We used five species traits (plant length, leaf length, leaf tissue density, specific leaf area, leaf nitrogen concentration) to calculate a functional diversity index weighted by relative species densities in mixture (Qd/m). This functional diversity index and aboveground biomass production (standing crop at end of experiment) in mixture were similar between early- and mid-successional communities but varied among communities within successional status. Mixture biomass was between the mean and maximum monoculture biomass, indicating that the latter species did not dominate mixtures and complementarity was not strong enough to cause transgressive overyielding. As expected, high mixture biomass was positively correlated with Qd/m. Also as expected, net biodiversity effects (mixture-mean monoculture biomass) were stronger in mid- than in early-successional pools. Net biodiversity effects increased with nitrogen level in some pools but decreased in others. The increase was consistent with a marginally positive effect of nitrogen level on functional diversity and a positive relation between functional diversity and net biodiversity effect.
Soil biodiversity and soil community composition determine ecosystem multifunctionality
Plant attribute diversity, resilience, and ecosystem function: the nature and significance of dominant and minor species
DOI:10.1007/s100219900062 URL [本文引用: 1]
Effects of clipping and fertilizing on the relationships between species diversity and ecosystem functioning and mechanisms of community stability in alpine meadow
DOI:10.3724/SP.J.1258.2013.00028 URL [本文引用: 1]
刈割、施肥对高寒草甸物种多样性与生态系统功能关系的影响及群落稳定性机制
DOI:10.3724/SP.J.1258.2013.00028
[本文引用: 1]
植物群落中不同“功能身份”物种的多样性与特定生态系统功能之间具有何种关系及其作用机制尚不明确。通过在高寒矮嵩草(Kobresia humilis)草甸为期5年的刈割(不刈割、留茬3 cm、留茬1 cm)、施肥(施肥、不施肥)和浇水(浇水、不浇水)控制实验, 研究了刈割与土壤资源获得性梯度上不同“功能身份”物种(群落中所有物种、响应物种、作用物种和共有物种)的多样性变化与群落地上净初级生产力和稳定性的关系以及稳定性机制。研究结果显示: 群落中响应物种、作用物种和共有物种数分别占全部物种数的36.6%、18.3%和64.8%, 物种多样性对生态系统功能具有不同的效应, 净初级生产力主要受响应物种和作用物种的多样性变化影响, 而稳定性则主要由共有物种的多样性变化决定; 群落稳定性的维持主要依赖于共有物种的多样性增加, 其作用机制是投资组合效应, 而超产效应和异步性效应对稳定性并无作用; 刈割和施肥对物种多样性、稳定性和净初级生产力具有相反的影响, 前者能增加物种多样性和稳定性, 并降低净初级生产力, 而后者的作用正相反。这与群落中全部物种的多样性变化受刈割影响较大, 而作用物种的多样性变化受资源获得性影响较大有关。上述结果表明高寒草甸生态系统地上净初级生产力主要由少数影响生产力的作用物种的多样性决定, 而稳定性则由大量共有物种的多样性所掌控。投资组合效应是物种多样性导致稳定性的机制。由于群落中不同物种的多样性效应具有分异性, 对于特定的生态系统功能而言, 物种的“功能身份”可能比物种多样性本身更重要, 不加区别地笼统定义物种多样性与生态系统功能的关系可能欠妥。
Effects of grazing exclusion on carbon sequestration and plant diversity in grasslands of China-A meta-analysis
DOI:10.1016/j.ecoleng.2016.06.124 URL [本文引用: 1]
Advances in research on the effects of exclosure on grasslands
围封对草地的影响研究进展
Response of plant functional traits at species and community levels to grazing exclusion on Inner Mongolian steppe, China
DOI:10.1071/RJ16086 URL [本文引用: 1]
Testing associations of plant functional diversity with carbon and nitrogen storage along a restoration gradient of sandy grassland

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}