
植物生态学报 ›› 2018, Vol. 42 ›› Issue (2): 185-194.DOI: 10.17521/cjpe.2017.0118
王树林,鲁为华( ),陈乙实,景鹏成
),陈乙实,景鹏成
出版日期:2018-02-20
发布日期:2018-04-16
通讯作者:
鲁为华 ORCID:0000-0003-0776-8912
基金资助:
WANG Shu-Lin,LU Wei-Hua(),CHEN Yi-Shi,JING Peng-Cheng
Online:2018-02-20
Published:2018-04-16
Contact:
Wei-Hua LU ORCID:0000-0003-0776-8912
Supported by:摘要:
为揭示北疆地区豆科植物种子对绵羊消化道作用的响应, 采集了北疆地区天然草地中常见的15种豆科植物种子。首先测定种子的长、宽、厚和质量, 并计算种子的形状指数。其次, 用种子饲喂绵羊, 收粪试验在种子摄食后的第6、12、24、36、48和72 h进行, 测定种子经绵羊消化道作用后的回收率、种子在绵羊消化道内的平均滞留时间以及消化道作用前后种子的萌发行为, 并研究了种子大小及形状指数对平均滞留时间和种子回收率的影响。结果如下: 15种植物种子质量在1.50-37.68 mg之间, 形状指数在0.001-0.12之间, 均为中等或较大类型的球(圆)形种子; 种子被绵羊采食后的排放动态符合高斯模型: Y = 0.02 + 0.74e -0.5(( X - 29.61)/9.41)2, 种子排放高峰期集中在采食后的24-36 h时间段内; 种子回收率最大的是洋甘草(Glycyrrhiza glabra)(39.25%), 最小的是草木犀(Melilotus officinalis)(4.28%); 平均滞留时间最长的是草木犀(37.19 h), 最短的是新疆棘豆(Oxytropis sinkiangensis)(22.33 h); 种子回收率与种子大小和形状指数之间分别具有符合形如Y = 6.45 + 2.05X - 0.04X 2和Y = 2.59 + 36.97e -24.47 X的函数关系模型; 平均滞留时间与种子大小和形状指数之间分别具有符合形如Y = 12.48 + 37.44e -0.07 X和Y = 3.93 + 2055.33X - 21757.99X 2的函数关系模型, 此结果表明, 较大、较小和形状不规则的豆科种子较易被绵羊消化和排泄。经绵羊消化道作用后, 多叶锦鸡儿(Caragana pleiophylla)种子的萌发率由消化前的96.22%显著降低至35.17%, 野火球(Trifolium lupinaster)和狐尾黄耆(Astragalus alopecurus)种子萌发率和消化前相比差异不显著, 其余12种种子的萌发率均显著提高(p < 0.05)。
王树林, 鲁为华, 陈乙实, 景鹏成. 北疆地区15种豆科植物种子对绵羊消化道作用的响应. 植物生态学报, 2018, 42(2): 185-194. DOI: 10.17521/cjpe.2017.0118
WANG Shu-Lin, LU Wei-Hua, CHEN Yi-Shi, JING Peng-Cheng. Effect of sheep digestive tract on the recovery and germination of seeds of fifteen leguminous plants in the northern Xinjiang region, China. Chinese Journal of Plant Ecology, 2018, 42(2): 185-194. DOI: 10.17521/cjpe.2017.0118
| 种名 Species | 单粒质量 Single seed mass (mg) | 形状指数 Shape index | 种名 Species | 单粒质量 Single seed mass (mg) | 形状指数 Shape index |
|---|---|---|---|---|---|
| 洋甘草 Glycyrrhiza glabra | 6.69 ± 0.03 | 0.001 ± 0.000 | 骆驼刺 Alhagi sparsifolia | 4.47 ± 0.03 | 0.070 ± 0.007 |
| 草木犀 Melilotus officinalis | 2.17 ± 0.02 | 0.050 ± 0.010 | 山羊豆 Galega officinalis | 6.22 ± 0.03 | 0.070 ± 0.006 |
| 苦马豆 Sphaerophysa salsula | 7.28 ± 0.02 | 0.070 ± 0.010 | 铃铛刺 Halimodendron halodendron | 15.73 ± 0.05 | 0.070 ± 0.005 |
| 蒙古黄耆 Astragalus mongholicus | 10.59 ± 0.02 | 0.100 ± 0.003 | 野苜蓿 Medicago falcata | 1.98 ± 0.01 | 0.070 ± 0.010 |
| 木黄耆 Astragalus arbuscula | 3.72 ± 0.03 | 0.120 ± 0.004 | 细叶野豌豆 Vicia tenuifolia | 14.84 ± 0.06 | 0.001 ± 0.001 |
| 新疆棘豆 Oxytropis sinkiangensis | 2.63 ± 0.03 | 0.070 ± 0.001 | 狐尾黄耆 Astragalus alopecurus | 34.42 ± 0.12 | 0.070 ± 0.001 |
| 多叶锦鸡儿 Caragana pleiophylla | 37.68 ± 0.07 | 0.070 ± 0.010 | 弯果胡卢巴 Trigonella arcuata | 1.50 ± 0.03 | 0.100 ± 0.008 |
| 野火球 Trifolium lupinaster | 1.67 ± 0.01 | 0.070 ± 0.010 |
表1 15种豆科种子的单粒质量及种子形状指数(平均值±标准误差)
Table 1 Single seed mass and seed shape index of seeds of fifteen leguminous species (mean ± SE)
| 种名 Species | 单粒质量 Single seed mass (mg) | 形状指数 Shape index | 种名 Species | 单粒质量 Single seed mass (mg) | 形状指数 Shape index |
|---|---|---|---|---|---|
| 洋甘草 Glycyrrhiza glabra | 6.69 ± 0.03 | 0.001 ± 0.000 | 骆驼刺 Alhagi sparsifolia | 4.47 ± 0.03 | 0.070 ± 0.007 |
| 草木犀 Melilotus officinalis | 2.17 ± 0.02 | 0.050 ± 0.010 | 山羊豆 Galega officinalis | 6.22 ± 0.03 | 0.070 ± 0.006 |
| 苦马豆 Sphaerophysa salsula | 7.28 ± 0.02 | 0.070 ± 0.010 | 铃铛刺 Halimodendron halodendron | 15.73 ± 0.05 | 0.070 ± 0.005 |
| 蒙古黄耆 Astragalus mongholicus | 10.59 ± 0.02 | 0.100 ± 0.003 | 野苜蓿 Medicago falcata | 1.98 ± 0.01 | 0.070 ± 0.010 |
| 木黄耆 Astragalus arbuscula | 3.72 ± 0.03 | 0.120 ± 0.004 | 细叶野豌豆 Vicia tenuifolia | 14.84 ± 0.06 | 0.001 ± 0.001 |
| 新疆棘豆 Oxytropis sinkiangensis | 2.63 ± 0.03 | 0.070 ± 0.001 | 狐尾黄耆 Astragalus alopecurus | 34.42 ± 0.12 | 0.070 ± 0.001 |
| 多叶锦鸡儿 Caragana pleiophylla | 37.68 ± 0.07 | 0.070 ± 0.010 | 弯果胡卢巴 Trigonella arcuata | 1.50 ± 0.03 | 0.100 ± 0.008 |
| 野火球 Trifolium lupinaster | 1.67 ± 0.01 | 0.070 ± 0.010 |
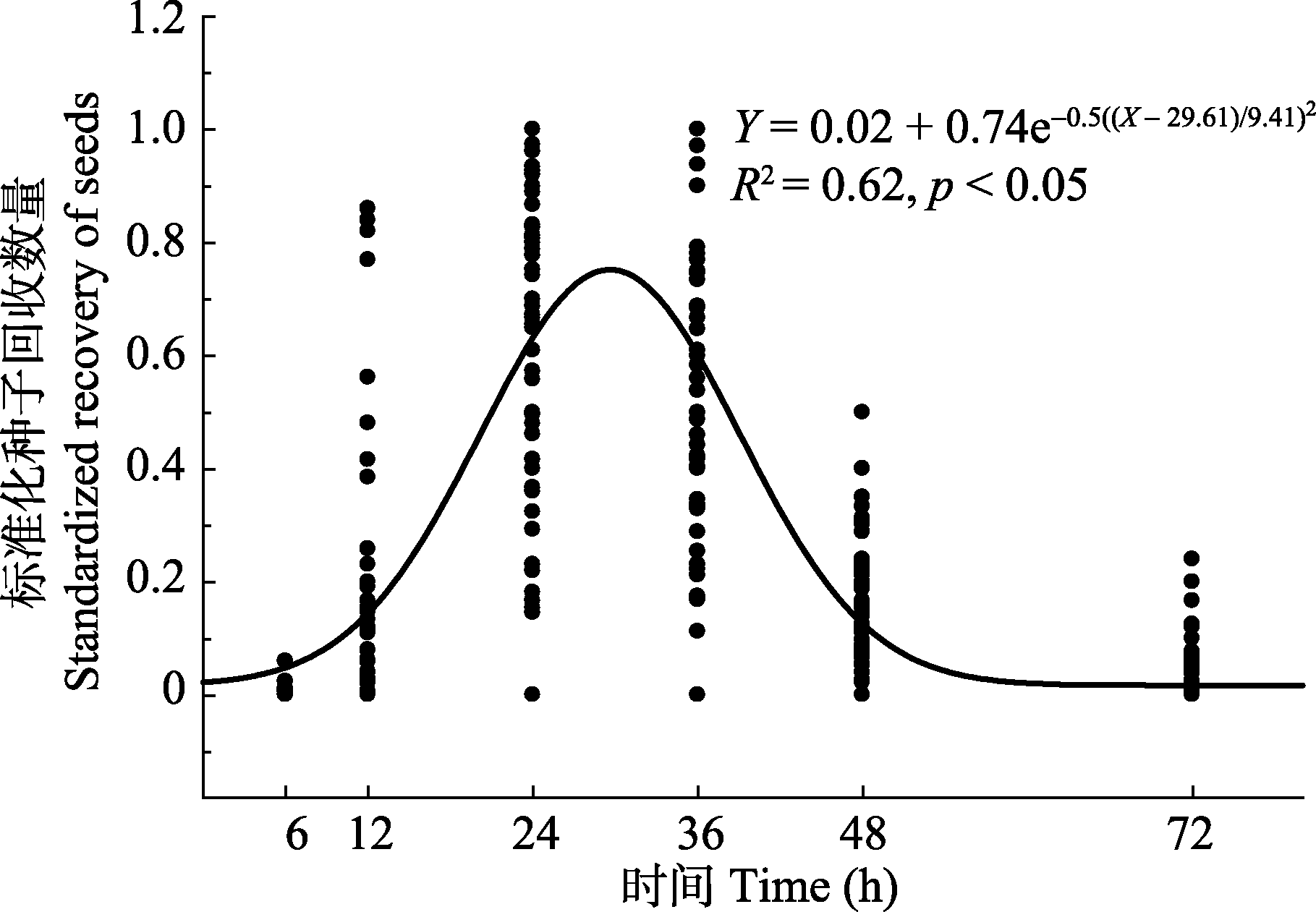
图1 种子饲喂给绵羊后的排放动态。
Fig. 1 Temporal changes in seed recovery after ingestion by sheep.
| 种名 Species | 种子回收率 SRP (%) | 平均滞留时间 MRT (h) | 种名 Species | 种子回收率 SRP (%) | 平均滞留时间 MRT (h) |
|---|---|---|---|---|---|
| 洋甘草 Glycyrrhiza glabra | 39.25 ± 20.40a | 31.62 ± 0.73bcd | 骆驼刺 Alhagi sparsifolia | 25.06 ± 9.25ab | 28.27 ± 0.27bcd |
| 草木犀 Melilotus officinalis | 4.28 ± 1.76c | 37.19 ± 2.12ab | 山羊豆 Galega officinalis | 38.02 ± 9.60a | 27.55 ± 1.75bcd |
| 苦马豆 Sphaerophysa salsula | 38.60 ± 19.77a | 32.48 ± 0.39abcd | 铃铛刺 Halimodendron halodendron | 34.56 ± 15.89a | 36.51 ± 5.93ab |
| 蒙古黄耆 Astragalus mongholicus | 4.88 ± 1.02c | 36.62 ± 0.54ab | 野苜蓿 Medicago falcata | 9.41 ± 1.26bc | 24.58 ± 0.84cd |
| 木黄耆 Astragalus arbuscula | 5.02 ± 1.25c | 32.51 ± 3.93abcd | 细叶野豌豆 Vicia tenuifolia | 34.90 ± 2.27a | 32.78 ± 2.94abcd |
| 新疆棘豆 Oxytropis sinkiangensis | 16.04 ± 4.11bc | 22.33 ± 0.77d | 狐尾黄耆 Astragalus alopecurus | 25.61 ± 11.75ab | 34.60 ± 0.84abc |
| 多叶锦鸡儿 Caragana pleiophylla | 8.93 ± 4.24bc | 33.76 ± 1.19abc | 弯果胡卢巴 Trigonella arcuata | 6.65 ± 1.44c | 24.95 ± 1.34cd |
| 野火球 Trifolium lupinaster | 14.01 ± 7.18bc | 30.04 ± 1.49bcd |
表2 15种豆科种子经绵羊采食后的种子回收率及种子在绵羊体内的平均滞留时间(平均值±标准误差)
Table 2 Seed recovery percentages and mean retention time of 15 leguminous species after sheep ingestion (mean ± SE)
| 种名 Species | 种子回收率 SRP (%) | 平均滞留时间 MRT (h) | 种名 Species | 种子回收率 SRP (%) | 平均滞留时间 MRT (h) |
|---|---|---|---|---|---|
| 洋甘草 Glycyrrhiza glabra | 39.25 ± 20.40a | 31.62 ± 0.73bcd | 骆驼刺 Alhagi sparsifolia | 25.06 ± 9.25ab | 28.27 ± 0.27bcd |
| 草木犀 Melilotus officinalis | 4.28 ± 1.76c | 37.19 ± 2.12ab | 山羊豆 Galega officinalis | 38.02 ± 9.60a | 27.55 ± 1.75bcd |
| 苦马豆 Sphaerophysa salsula | 38.60 ± 19.77a | 32.48 ± 0.39abcd | 铃铛刺 Halimodendron halodendron | 34.56 ± 15.89a | 36.51 ± 5.93ab |
| 蒙古黄耆 Astragalus mongholicus | 4.88 ± 1.02c | 36.62 ± 0.54ab | 野苜蓿 Medicago falcata | 9.41 ± 1.26bc | 24.58 ± 0.84cd |
| 木黄耆 Astragalus arbuscula | 5.02 ± 1.25c | 32.51 ± 3.93abcd | 细叶野豌豆 Vicia tenuifolia | 34.90 ± 2.27a | 32.78 ± 2.94abcd |
| 新疆棘豆 Oxytropis sinkiangensis | 16.04 ± 4.11bc | 22.33 ± 0.77d | 狐尾黄耆 Astragalus alopecurus | 25.61 ± 11.75ab | 34.60 ± 0.84abc |
| 多叶锦鸡儿 Caragana pleiophylla | 8.93 ± 4.24bc | 33.76 ± 1.19abc | 弯果胡卢巴 Trigonella arcuata | 6.65 ± 1.44c | 24.95 ± 1.34cd |
| 野火球 Trifolium lupinaster | 14.01 ± 7.18bc | 30.04 ± 1.49bcd |
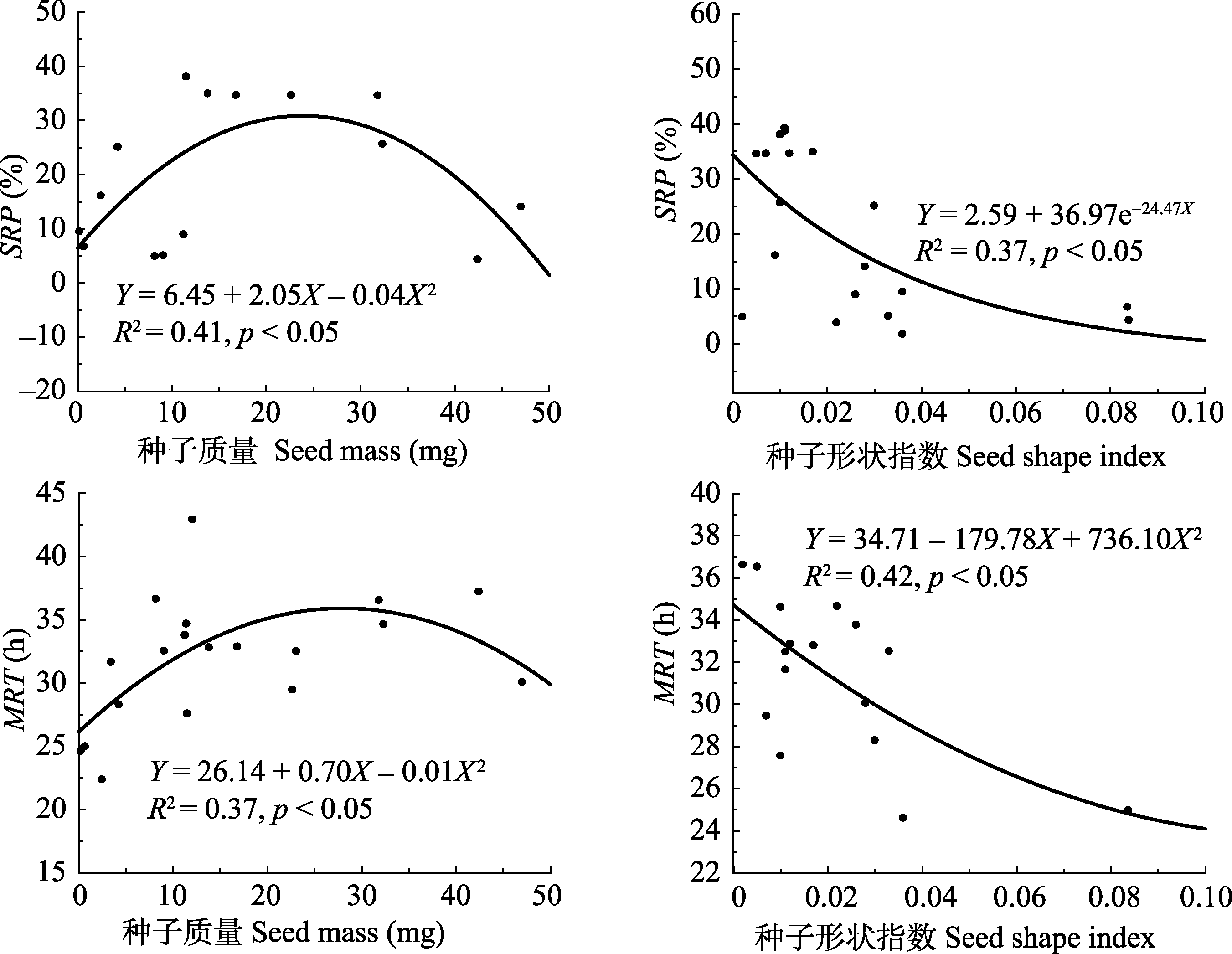
图2 种子质量和形状指数与种子回收率和平均滞留时间的关系。
Fig. 2 The relationship between seed mass, seed shape index and seed recovery percentage and mean retention time.
| 种名 Species | 萌发率 Germination rates (%) | 萌发率增值 Appreciation of germination rates (%) | |
|---|---|---|---|
| 对照 Control | 消化后 After sheep ingested | ||
| 洋甘草 Glycyrrhiza glabra | 15.36 ± 5.55b | 68.41 ± 13.86a | 53.05 |
| 草木犀 Melilotus officinalis | 6.67 ± 1.16b | 23.68 ± 1.67a | 17.01 |
| 苦马豆 Sphaerophysa salsula | 15.00 ± 7.00b | 71.01 ± 3.62a | 56.01 |
| 蒙古黄耆 Astragalus mongholicus | 14.33 ± 3.22b | 25.00 ± 3.61a | 10.67 |
| 木黄耆 Astragalus arbuscula | 45.00 ± 2.00b | 53.70 ± 1.57a | 8.70 |
| 新疆棘豆 Oxytropis sinkiangensis | 33.33 ± 1.54a | 35.39 ± 3.08a | 2.06 |
| 多叶锦鸡儿 Caragana pleiophylla | 96.22 ± 6.33a | 35.17 ± 3.32b | -61.05 |
| 野火球 Trifolium lupinaster | 28.33 ± 3.06a | 21.43 ± 3.11a | -6.90 |
| 骆驼刺 Alhagi sparsifolia | 8.33 ± 3.51b | 42.25 ± 2.63a | 33.92 |
| 山羊豆 Galega officinalis | 13.33 ± 1.53a | 14.45 ± 1.27a | 1.12 |
| 铃铛刺 Halimodendron halodendron | 1.67 ± 0.58b | 11.96 ± 2.00a | 10.29 |
| 野苜蓿 Medicago falcata | 7.82 ± 1.05b | 62.36 ± 3.01a | 54.54 |
| 细叶野豌豆 Vicia tenuifolia | 8.33 ± 0.58b | 28.52 ± 6.73a | 20.19 |
| 狐尾黄耆 Astragalus alopecurus | 6.30 ± 2.14a | 5.83 ± 2.56a | -0.47 |
| 弯果胡卢巴 Trigonella arcuata | 5.20 ± 1.31b | 20.32 ± 5.51a | 15.12 |
表3 绵羊消化前后15种豆科种子的萌发率(平均值±标准误差)
Table 3 Germination percentages of seeds of 15 leguminous species before and after sheep ingestion (mean ± SE)
| 种名 Species | 萌发率 Germination rates (%) | 萌发率增值 Appreciation of germination rates (%) | |
|---|---|---|---|
| 对照 Control | 消化后 After sheep ingested | ||
| 洋甘草 Glycyrrhiza glabra | 15.36 ± 5.55b | 68.41 ± 13.86a | 53.05 |
| 草木犀 Melilotus officinalis | 6.67 ± 1.16b | 23.68 ± 1.67a | 17.01 |
| 苦马豆 Sphaerophysa salsula | 15.00 ± 7.00b | 71.01 ± 3.62a | 56.01 |
| 蒙古黄耆 Astragalus mongholicus | 14.33 ± 3.22b | 25.00 ± 3.61a | 10.67 |
| 木黄耆 Astragalus arbuscula | 45.00 ± 2.00b | 53.70 ± 1.57a | 8.70 |
| 新疆棘豆 Oxytropis sinkiangensis | 33.33 ± 1.54a | 35.39 ± 3.08a | 2.06 |
| 多叶锦鸡儿 Caragana pleiophylla | 96.22 ± 6.33a | 35.17 ± 3.32b | -61.05 |
| 野火球 Trifolium lupinaster | 28.33 ± 3.06a | 21.43 ± 3.11a | -6.90 |
| 骆驼刺 Alhagi sparsifolia | 8.33 ± 3.51b | 42.25 ± 2.63a | 33.92 |
| 山羊豆 Galega officinalis | 13.33 ± 1.53a | 14.45 ± 1.27a | 1.12 |
| 铃铛刺 Halimodendron halodendron | 1.67 ± 0.58b | 11.96 ± 2.00a | 10.29 |
| 野苜蓿 Medicago falcata | 7.82 ± 1.05b | 62.36 ± 3.01a | 54.54 |
| 细叶野豌豆 Vicia tenuifolia | 8.33 ± 0.58b | 28.52 ± 6.73a | 20.19 |
| 狐尾黄耆 Astragalus alopecurus | 6.30 ± 2.14a | 5.83 ± 2.56a | -0.47 |
| 弯果胡卢巴 Trigonella arcuata | 5.20 ± 1.31b | 20.32 ± 5.51a | 15.12 |
| [1] | Baskin CC, Baskin JM ( 2014). Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination. 2nd edn. Academic Press, New York. |
| [2] |
Blackshaw RE, Rode LM ( 1991). Effect of ensiling and rumen digestion by cattle on weed seed viability. Weed Science, 39, 104-108.
DOI URL |
| [3] |
Brochet AL, Guillemain M, Gauthier-Clerc M, Fritz H, Green AJ ( 2010). Endozoochory of Mediterranean aquatic plant seeds by teal after a period of desiccation: Determinants of seed survival and influence of retention time on germinability and viability. Aquatic Botany, 93, 99-106.
DOI URL |
| [4] |
Calvi?o-Cancela M, Martín-Herrero J ( 2009). Effectiveness of a varied assemblage of seed dispersers of a fleshy-fruited plant. Ecology, 90, 3503-3515.
DOI URL PMID |
| [5] | Chen BJ ( 2008). Legume forage resources and floristic characteristics in Gansu Province. Pratacultural Science, 25(4), 42-45. |
| [ 陈本建 ( 2008). 甘肃省豆科牧草资源及植物区系分析. 草业科学, 25(4), 42-45.] | |
| [6] |
Cosyns E, Delporte A, Lens L, Hoffmann M ( 2005). Germination success of temperate grassland species after passage through ungulate and rabbit guts. Journal of Ecology, 93, 353-361.
DOI URL |
| [7] |
Couvreur M, Cosyns E, Hermy M, Hoffmann M ( 2005). Complementarity of epi- and endozoochory of plant seeds by free ranging donkeys. Ecography, 28, 37-48.
DOI URL |
| [8] |
D’Hondt B, Hoffmann M ( 2011). A reassessment of the role of simple seed traits in mortality following herbivore ingestion. Plant Biology, 13, 118-124.
DOI URL PMID |
| [9] | G?kbulak F ( 2002). Effect of American bison ( Bison bison L.) on the recovery and germinability of seeds of range forage species. Grass & Forage Science, 57, 395-400. |
| [10] |
G?kbulak F ( 2006). Recovery and germination of grass seeds ingested by cattle. The Journal of Biological Sciences, 6, 23-27.
DOI URL |
| [11] |
Grande D, Martinvicente A ( 2013). Endozoochorus seed dispersal by goats: Recovery, germinability and emergence of five Mediterranean shrub species. Spanish Journal of Agricultural Research, 11, 347-355.
DOI URL |
| [12] | Han JG ( 2007). Grassland Science. 3rd edn. China Agriculture Press, Beijing. |
| [ 韩建国 ( 2007). 草地学. 第三版. 中国农业出版社, 北京.] | |
| [13] | Heidrun W, Oliver T ( 2008). A mechanistic simulation model of seed dispersal by animals. Journal of Ecology, 96, 1011-1022. |
| [14] | Janzen DH ( 1977). Variation in seed size within a crop of a costa rican Mucuna andreana( Leguminosae). American Journal of Botany, 64, 347-349. |
| [15] | Janzen DH ( 1984). Dispersal of small seeds by big herbivores: Foliage is the fruit. The American Naturalist, 123, 338-353. |
| [16] | Jaroszewicz B ( 2013). Endozoochory by European bison influences the build-up of the soil seed bank in subcontinental coniferous forest. European Journal of Forest Research, 132, 445-452. |
| [17] | Jurado E, Estrada E, Moles A ( 2001). Characterizing plant attributes with particular emphasis on seeds in Tamaulipan thornscrub in semi-arid Mexico. Journal of Arid Environments, 48, 309-321. |
| [18] | Klein J ( 1981). La Mesta. 2nd edn. Alianza Editorial, Madrid, Spain. |
| [19] | Lu WH, Wan JJ, Yang JJ, Ren AT, Yu L ( 2013). Review of endozoochory of plant seeds by herbivores. Acta Prataculturae Sinica, 22, 306-313. |
| [ 鲁为华, 万娟娟, 杨洁晶, 任爱天, 于磊 ( 2013). 草食动物对植物种子的消化道传播研究进展. 草业学报, 22, 306-313.] | |
| [20] | Ma SB, Jiang HQ ( 1999). Study on the seed weight and seed seize variation pattern and their biological significance in Podophylloideae (Berberjdaceae). Acta Botanica Boreali-?Occidentalia Sinica, 19, 715-724. |
| [ 马绍宾, 姜汉侨 ( 1999). 小檗科鬼臼亚科种子大小变异式样及其生物学意义. 西北植物学报, 19, 715-724.] | |
| [21] | Mancilla-Leytón JM, Fernández-Alés R, Vicente AM ( 2012). Plant-ungulate interaction: Goat gut passage effect on survival and germination of Mediterranean shrub seeds. Journal of Vegetation Science, 22, 1031-1037. |
| [22] | Manzano P, Malo JE, Peco B ( 2005). Sheep gut passage and survival of Mediterranean shrub seeds. Seed Science Research, 15, 21-28. |
| [23] | Miloti? T, Hoffmann M ( 2016). How does gut passage impact endozoochorous seed dispersal success? Evidence from a gut environment simulation experiment. Basic and Applied Ecology, 17, 165-176. |
| [24] | Mou XM, Yu YW, Zhang HM, Sun H, Wang HC, Xu CL, Hua LM ( 2013). Effects of yakdung deposition on community characteristics and niche parameters in alpine meadow. Pratacultural Science, 30, 1594-1601. |
| [ 牟晓明, 于应文, 张红梅, 孙红, 王虎成, 徐长林, 花力民 ( 2013). 牦牛粪对高寒草甸植被群落特征和生态位参数的影响. 草业科学, 30, 1594-1601.] | |
| [25] | Mouissie AM, Veen CEJVD, Veen GF, Diggelen RV ( 2005 a). Ecological correlates of seed survival after ingestion by fallow deer. Functional Ecology, 19, 284-290. |
| [26] | Mouissie AM, Vos P, Verhagen HMC, Bakker JP ( 2005 b). Endozoochory by free-ranging, large herbivores: Ecological correlates and perspectives for restoration. Basic and Applied Ecology, 6, 547-558. |
| [27] | Myers JA, Vellend M, Gardescu S, Marks PL ( 2004). Seed dispersal by white-tailed deer: Implications for long-????distance dispersal, invasion, and migration of plants in eastern North America. Oecologia, 139, 35-44. |
| [28] | Oudtshoorn KVRV, Rooyen MWV ( 1998). Dispersal Biology of Desert Plants. Springer, Berlin. |
| [29] | Pan L, Niu JL, Bu PD, Du H, Cheng JB, Sun XZ, Wang XM, Qin JJ, Yuan YM, Zhang XK ( 2015). Effects of saikosaponin on in vitro fermentation parameters and bacterial quantity. Acta Prataculturae Sinica, 24(6), 85-91. |
| [ 潘龙, 牛俊丽, 卜登攀, 杜洪, 程建波, 孙先枝, 王秀敏, 秦俊杰, 袁耀明, 张幸开 ( 2015). 柴胡皂苷对体外发酵参数及细菌数量变化的影响. 草业学报, 24(6), 85-91.] | |
| [30] | Plumptre AJ ( 2003). Seed germination and early seedling establishment of some elephant-dispersed species in Banyang-?mbo wildlife sanctuary, South-western Cameroon. Journal of Tropical Ecology, 19, 229-237. |
| [31] | Ramos ME, Robles AB, Castro J ( 2006). Efficiency of endozoochorous seed dispersal in six dry-fruited species (Cistaceae): From seed ingestion to early seedling establishment. Plant Ecology, 185, 97-106. |
| [32] | Russi L, Cocks PS, Roberts EH ( 1992). The fate of Legume seeds eaten by sheep from a Mediterranean grassland. Journal of Applied Ecology, 29, 772-778. |
| [33] | Soons MB, van der Vlugt C, Van Lith B, Heil GW, Klaassen M ( 2008). Small seed size increases the potential for dispersal of wetland plants by ducks. Journal of Ecology, 96, 619-627. |
| [34] | Spiegel O, Nathan R ( 2007). Incorporating dispersal distance into the disperser effectiveness framework: Frugivorous birds provide complementary dispersal to plants in a patchy environment. Ecology Letters, 10, 718-728. |
| [35] | Thompson K, Band SR, Hodgson JG ( 1993). Seed size and shape predict persistance in soil. Functional Ecology, 7, 236-241. |
| [36] | Thomson FJ, Moles AT, Auld TD, Kingsford RT ( 2011). Seed dispersal distance is more strongly correlated with plant height than with seed mass. Journal of Ecology, 99, 1299-1307. |
| [37] | Wali N, Wang SL, Zhao G, Yang JJ, Lü YP, Lu WH ( 2016). Effect of sheep digestion on morphology, recovery and germinability of four Leguminous plants seed. Pratacultural Science, 33, 1566-1573. |
| [ 娜丽克斯·外里, 王树林, 赵刚, 杨洁晶, 吕艳萍, 鲁为华 ( 2016). 4种豆科牧草种子对绵羊消化道作用的反应. 草业科学, 33, 1566-1573.] | |
| [38] | Wan JJ, Yang JJ, Ren AT, Ma CH, Yu L, Lu WH ( 2013). Effects of herbivores on seed germination by meta analysis. Pratacultural Science, 30, 1852-1861. |
| [ 万娟娟, 杨洁晶, 任爱天, 马春晖, 于磊, 鲁为华 ( 2013). 草食动物影响植物种子萌发的整合分析. 草业科学, 30, 1852-1861.] | |
| [39] | Wang BC, Smith TB ( 2002). Closing the seed dispersal loop. Trends in Ecology and Evolution, 17, 379-386. |
| [40] | Wang CJ, Wang WQ, Lu WH, Wen CL, Yin XJ, Zhao QZ ( 2016). Feed intake distribution model for herd based on grazing spatio-temporal trajectory data. Transactions of the Chinese Society of Agricultural Engineering, 32(13), 125-130. |
| [ 汪传建, 王伟强, 鲁为华, 文春雷, 尹小君, 赵庆展 ( 2016). 基于放牧时空轨迹数据的牧群采食量分布模型. 农业工程学报, 32(13), 125-130.] | |
| [41] | Wang SL, Lu WH, Wali N, Ma CH, Zhang QB, Wang CJ ( 2017). Recovery and germination of seeds after passage through the gut of Kazakh sheep on the north slope of the Tianshan Mountains. Seed Science Research, 27, 43-49. |
| [42] | Wang SL, Wali N, Lu WH, Wang WQ, Wang CJ ( 2017). Morphological characteristics of six kinds of mucilaginous seeds and their response to digestion in rumen of sheep. Acta Prataculturae Sinica, 26(4), 89-98. |
| [ 王树林, 娜丽克斯·外里, 鲁为华, 王伟强, 汪传建 ( 2017). 六种粘液种子性状多样性及其对绵羊瘤胃消化的反应. 草业学报, 26(4), 89-98.] | |
| [43] | Willms WD, Acharya SN, Rode LM ( 1995). Feasibility of using cattle to disperse cicer milkvetch ( Astragalus cicer L.) seed in pastures. Canadian Veterinary Journal, 75, 173-175. |
| [44] | Yan XH, An SZ, Yu X, Jia SB, Pan XD, Cheng FX, Li H ( 2009). Observation on grazing behavior of Kazak sheep in summer. Journal of Xinjiang Agricultural University, 32(4), 38-40. |
| [ 闫学慧, 安沙舟, 余雄, 贾帅兵, 潘晓东, 程发祥, 李海 ( 2009). 哈萨克羊夏季牧食行为观测. 新疆农业大学学报, 32(4), 38-40.] | |
| [45] | Yang JJ, Wan JJ, Wali N, Ren AT, Lu WH ( 2015). Seed morphology and effect of sheep rumen digestion on germinability of 28 grass of Tianshan. Acta Prataculturae Sinica, 24(2), 104-115. |
| [ 杨洁晶, 万娟娟, 娜丽克斯·外里, 任爱天, 鲁为华 ( 2015). 28种植物种子形态学性状及其萌发对绵羊瘤胃消化的反应. 草业学报, 24(2), 104-115.] | |
| [46] | Yu L, Lu WH, Yan P, Luo LJ, Zhang QB ( 2014). Resources and assessment of natural grassland in Shaertao Mountains, Zhaosu County in Xinjiang. Chinese Journal of Grassland, 36(6), 4-11. |
| [ 于磊, 鲁为华, 阎平, 罗良俊, 张前兵 ( 2014). 新疆昭苏县境沙尔套山天然草地植物资源与评价. 中国草地学报, 36(6), 4-11.] | |
| [47] | Yu XJ, Xu CL, Wang F, Shang ZH, Long RJ ( 2012). Recovery and germinability of seeds ingested by yaks and Tibetan sheep could have important effects on the population dynamics of alpine meadow plants on the Qinghai-Tibetan Plateau. Rangeland Journal, 34, 249-255. |
| [48] | Zhao LL, Wang ZL, Du JC, Li QF, Song CL ( 2007). The effect of sulfuric acid treatment on hard seeds of Melimotoides ruthenica strains. Chinese Journal of Grassland, 29(3), 73-77. |
| [ 赵丽丽, 王照兰, 杜建材, 李青丰, 宋彩玲 ( 2007). 硫酸处理打破扁蓿豆不同品系种子硬实的效果研究. 中国草地学报, 29(3), 73-77.] |
| [1] | 苏华, 许宏, 苏本营, 李永庚. 养分添加对退化草地豆科植物草木犀功能性状的影响[J]. 植物生态学报, 2020, 44(9): 926-938. |
| [2] | 王祖幸, 何维明. 土埋深度影响物种水平的种子质量-种子出苗关系[J]. 植物生态学报, 2019, 43(10): 899-908. |
| [3] | 李道新, 李果, 沈泽昊, 徐慎东, 韩庆瑜, 王功芳, 田风雷. 植物生长型显著影响三峡大老岭地区木本植物种子质量的海拔格局[J]. 植物生态学报, 2017, 41(5): 539-548. |
| [4] | 刘双娥, 李义勇, 方熊, 黄文娟, 龙凤玲, 刘菊秀. 不同氮添加量和添加方式对南亚热带4个主要树种幼苗生长的影响[J]. 植物生态学报, 2015, 39(10): 950-961. |
| [5] | 朱军涛, 李向义, 张希明, 林丽莎, 杨尚功. 塔克拉玛干沙漠南缘豆科与非豆科植物的氮分配[J]. 植物生态学报, 2010, 34(9): 1025-1032. |
| [6] | 丑敏霞, 魏新元. 豆科植物共生结瘤的分子基础和调控研究进展[J]. 植物生态学报, 2010, 34(7): 876-888. |
| [7] | 张黎, 于贵瑞, 何洪林, 张雷明. 基于模型数据融合的长白山阔叶红松林碳循环模拟[J]. 植物生态学报, 2009, 33(6): 1044-1055. |
| [8] | 滕年军, 陈彤, 林金星. 植物有性生殖对大气CO2浓度变化响应的研究进展[J]. 植物生态学报, 2006, 30(6): 1054-1063. |
| [9] | 全占军, 程宏, 于云江, 邹学勇. 煤矿井田区地表沉陷对植被景观的影响——以山西省晋城市东大煤矿为例[J]. 植物生态学报, 2006, 30(3): 414-420. |
| [10] | 袁志友, 李凌浩, 韩兴国. 藜个体在高密度种群中的氮素利用效率[J]. 植物生态学报, 2004, 28(3): 294-299. |
| [11] | 牛书丽, 蒋高明. 豆科固氮植物对CO2加富的生理响应[J]. 植物生态学报, 2003, 27(6): 844-851. |
| [12] | 葛滢, 常杰, 岳春雷, 陆大根. 杭州石荠苧种子萌发的生理生态学研究[J]. 植物生态学报, 1998, 22(2): 171-177. |
| [13] | 曾小平, 赵平, 彭少麟, 余作岳. 5种木本豆科植物的光合特性研究[J]. 植物生态学报, 1997, 21(6): 539-544. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
Copyright © 2022 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19