
|
|
||
|
亚热带典型树种根毛特征及其与共生真菌的关系
植物生态学报
2023, 47 (1):
88-100.
DOI: 10.17521/cjpe.2022.0131
根毛和共生真菌增加了吸收面积, 提高了植物获取磷等土壤资源的能力。由于野外原位观测根表微观结构较为困难, 吸收细根、根毛、共生真菌如何相互作用并适应土壤资源供应, 缺乏相应的数据和理论。该研究以受磷限制的亚热带森林为对象, 选取了21种典型树种, 定量了根毛存在情况、属性变异, 分析了根毛形态特征与共生真菌侵染率、吸收细根功能属性之间的关系, 探讨了根表结构对低磷土壤的响应和适应格局。结果表明: 1)在亚热带森林根毛不是普遍存在的, 21个树种中仅发现7个树种存有根毛, 4个为丛枝菌根(AM)树种, 3个为外生菌根(ECM)树种。其中, 马尾松(Pinus massoniana)根毛出现率最高, 为86%; 2)菌根类型是理解根-根毛-共生真菌关系的关键, AM树种根毛密度与共生真菌侵染率正相关, 但ECM树种根毛直径与共生真菌侵染率负相关; 3) AM树种根毛长度和根毛直径、ECM树种根毛出现率与土壤有效磷含量呈负相关关系。该研究揭示了不同菌根类型树种根毛-共生真菌-根属性的格局及相互作用, 为精细理解养分获取策略奠定了基础。 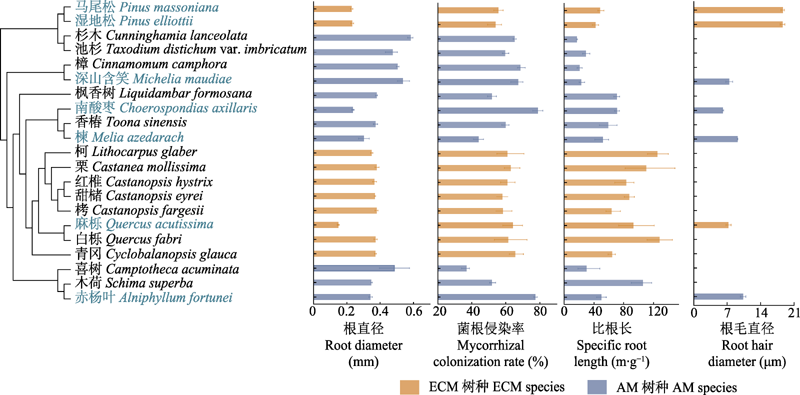 View image in article
图1
21个亚热带典型树种谱系发育关系及根属性和根毛属性的变异(平均值±标准误)。AM, 丛枝菌根; ECM, 外生菌根。观察到根毛的物种名用蓝色标注, 其中, 根毛直径仅在观察到根毛的7个物种中展示。
正文中引用本图/表的段落
综合考虑千烟洲站样地的树种分布和亚热带森林主要建群种等信息, 于千烟洲站选取21种亚热带典型树种作为研究对象(图1), 包含松科、柏科、漆树科、安息香科、山茶科、楝科、金缕梅科、蓝果树科、壳斗科、木兰科、樟科等11个科17个属; 包含11种AM树种, 10种ECM树种。采集样品时, 在目标树种的树干周围的不同方向, 沿树干向外寻找与主干相连的根系, 选择包含1-5级完整根级的侧根, 进行根样采集, 每个树种4个重复。为保证根毛结构的完整性, 从土壤中采集根样时需轻挖轻剪(Lynch et al., 2021), 将整个根样连根所附着的土壤一起取出, 带回实验室使用无菌刷子轻轻去除根部附着的土壤颗粒, 确保不损坏根毛。根据后续实验需要, 一部分根样储存在4 ℃冰箱, 一部分浸泡在FAA固定液里以备后续分析处理。取根样的同时, 在取样根系0.5 m范围内用土钻钻取0-20 cm的土壤样品, 每个树种4个重复。
首先遵循模拟河流分支溯源的根系分级方法(Pregitzer et al., 2002; McCormack et al., 2015), 将采集的完整根样分成不同根级, 前两级根为吸收细根(Guo et al., 2008a), 取根毛完整的1、2级根浸泡在FAA固定液中保存, 用于后续吸收根根毛属性的测定。根据以往关于根毛的研究(Gahoonia & Nielsen, 2004; Miguel et al., 2015), 通过光学体式显微镜观察根毛, 拍摄根毛图片并用ImageJ进行根毛数据提取, 是较为经济有效的方法(Zhu et al., 2010; Lynch et al., 2021)。本研究在搭载相机(Leica DFC450, Leica, Nussloch, Germany)的体视显微镜(Leica DM1000, Leica, Nussloch, Germany)下观察记录21个树种前两级根(吸收细根)的根毛存在情况并进行根毛属性的测定。具体操作为: 用镊子将每个树种的吸收细根从FAA固定液中取出置于培养皿中, 每个树种每个重复随机选取100-150个根段, 并记录所观察根段的总数, 显微镜观察后, 记录有根毛的根段个数; 通过观察有根毛的根段个数和观察的总根段个数之比计算根毛出现率。拍摄根毛照片时, 将显微镜镜头聚焦到有根毛的根段根表的两侧区域, 以最优视野对该根段各个区域进行图片拍摄。对每一张根毛照片采用Image J软件进行根毛密度、根毛长度、根毛直径数据的提取。根毛密度是指1 mm根段上的根毛个数; 根毛长度是指从根表面凸起的根毛细胞的长度; 根毛直径是指根毛细胞的宽度。来自同一根段多张照片的根毛密度、根毛长度、根毛直径的均值作为该根段根毛密度、根毛长度、根毛直径的值。
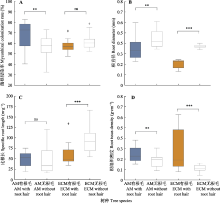
在11个AM树种中, 菌根真菌侵染率的种间差异显著(p < 0.05), 吸收根菌根真菌侵染率最高的是有根毛的南酸枣为78.88%, 最小的是没有观察到根毛的喜树(Camptotheca acuminata)为46.99%。而10个ECM树种中吸收根菌根真菌侵染率的种间差异不显著(p = 0.17), 侵染率最高的是没有观察到根毛的青冈(Cyclobalanopsis glauca)为65.54%, 最低的是有根毛的湿地松, 为54.06% (图1)。有根毛树种和无根毛树种的菌根真菌侵染率的差异与不同类型的菌根真菌有关。AM树种中有根毛树种的平均侵染率为66.93%, 显著高于无根毛的树种(平均菌根真菌侵染率为56.29%) (图3A)。而有根毛的ECM树种平均菌根真菌侵染率为57.99%, 无根毛的为61.17%, 二者差异不显著(图3A)。在7个有根毛的树种中, AM树种菌根真菌侵染率(66.93%)总体上比ECM树种(57.99%)高, 且AM树种中菌根侵染率排序为: 南酸枣>赤杨叶>深山含笑>楝; ECM树种中为: 麻栎>马尾松>湿地松(图1)。
无论是AM树种还是ECM树种, 吸收根直径、比根长及根组织密度均有种间差异(图1)。AM树种中吸收根直径最大(0.58 mm)、比根长最小(17.09 m·g-1)的是没有观察到根毛的杉木; 吸收根直径最小(0.25 mm)、根组织密度最高(0.33 g·cm-3)的是有根毛的南酸枣; 比根长最大(105.8 m·g-1)、根组织密度最低(0.1 g·cm-3)的是没有观察到根毛的木荷。ECM树种中, 没有观察到根毛的栗的吸收根直径最大(0.48 μm), 最小的是有根毛的麻栎(0.15 mm); 吸收根比根长最大的是无根毛的柯(Lithocarpus glaber, 124.97 m·g-1), 最小的是有根毛的湿地松(42.08 m·g-1)。而根组织密度最高的物种是有根毛的马尾松, 高达0.53 g·cm-3; 最低的是无根毛的白栎, 仅有0.07 g·cm-3 (图1)。无论是AM还是ECM树种, 有根毛的物种比没有观察到根毛的物种具有更小的吸收根直径(AM: 0.35 mm vs 0.45 mm; ECM: 0.20 mm vs 0.37 mm)和更高的根组织密度(AM: 0.24 g·cm-3 vs 0.18 g·cm-3; ECM: 0.28 g·cm-3 vs 0.11 g·cm-3) (图3B、3D)。有根毛和无根毛AM树种的比根长差异不显著(p = 0.51), 但ECM树种有根毛的物种的比根长显著小于没有观察到根毛的物种(61.10 m·g-1 vs 94.58 m·g-1) (图3C)。
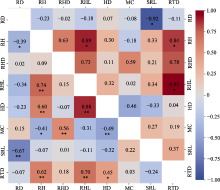
通过21个亚热带典型树种构建的进化树可以看出, 观察到有根毛的7个树种的系统发育关系区分明显(图1), 并且这7个物种的根及根毛属性种间差异显著(图1; 表1)。其中, 根毛出现率最大和最小值之间相差4.9倍, 种间变异系数为36.39%; 根毛密度的变异系数为52.08%, 除根毛密度最大的赤杨叶, 其余物种根毛密度主要集中在15-20 No.·mm-1之间; 根毛长度最大和最小值相差10.5倍, 变异系数达到了71.71%; 根毛直径也有46.98%的变异度。菌根真菌侵染率的种间变异最小为20.22%。根直径、比根长、根组织密度种间变异分别为43.10%、55.06%、53.26%。根据Blomberg’s K值结果分析这7个有根毛的物种的根毛和根属性的系统保守性, 发现根毛出现率、根毛密度、根毛长度的系统发育信号较弱且系统发育信号影响不显著(K < 1, p > 0.05), 而根毛直径受系统发育信号的影响较强(K > 1, p < 0.05) (表2)。而吸收根的比根长受系统发育信号的影响较小且不显著(K < 1, p > 0.05); 根直径的系统发育信号K值接近1且p < 0.01, 表现出很强的系统发育保守性; 根组织密度和菌根侵染率受系统发育信号影响不显著(K < 1, p > 0.05) (表2)。
本文的其它图/表
|



{kind=link}