|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
亚热带典型树种根毛特征及其与共生真菌的关系
植物生态学报
2023, 47 (1):
88-100.
DOI: 10.17521/cjpe.2022.0131
根毛和共生真菌增加了吸收面积, 提高了植物获取磷等土壤资源的能力。由于野外原位观测根表微观结构较为困难, 吸收细根、根毛、共生真菌如何相互作用并适应土壤资源供应, 缺乏相应的数据和理论。该研究以受磷限制的亚热带森林为对象, 选取了21种典型树种, 定量了根毛存在情况、属性变异, 分析了根毛形态特征与共生真菌侵染率、吸收细根功能属性之间的关系, 探讨了根表结构对低磷土壤的响应和适应格局。结果表明: 1)在亚热带森林根毛不是普遍存在的, 21个树种中仅发现7个树种存有根毛, 4个为丛枝菌根(AM)树种, 3个为外生菌根(ECM)树种。其中, 马尾松(Pinus massoniana)根毛出现率最高, 为86%; 2)菌根类型是理解根-根毛-共生真菌关系的关键, AM树种根毛密度与共生真菌侵染率正相关, 但ECM树种根毛直径与共生真菌侵染率负相关; 3) AM树种根毛长度和根毛直径、ECM树种根毛出现率与土壤有效磷含量呈负相关关系。该研究揭示了不同菌根类型树种根毛-共生真菌-根属性的格局及相互作用, 为精细理解养分获取策略奠定了基础。
表2
7个亚热带典型树种根毛与根属性变异与系统发育信号(Blomberg’s K)
正文中引用本图/表的段落
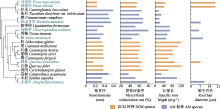
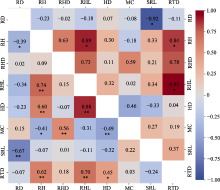
通过21个亚热带典型树种构建的进化树可以看出, 观察到有根毛的7个树种的系统发育关系区分明显(图1), 并且这7个物种的根及根毛属性种间差异显著(图1; 表1)。其中, 根毛出现率最大和最小值之间相差4.9倍, 种间变异系数为36.39%; 根毛密度的变异系数为52.08%, 除根毛密度最大的赤杨叶, 其余物种根毛密度主要集中在15-20 No.·mm-1之间; 根毛长度最大和最小值相差10.5倍, 变异系数达到了71.71%; 根毛直径也有46.98%的变异度。菌根真菌侵染率的种间变异最小为20.22%。根直径、比根长、根组织密度种间变异分别为43.10%、55.06%、53.26%。根据Blomberg’s K值结果分析这7个有根毛的物种的根毛和根属性的系统保守性, 发现根毛出现率、根毛密度、根毛长度的系统发育信号较弱且系统发育信号影响不显著(K < 1, p > 0.05), 而根毛直径受系统发育信号的影响较强(K > 1, p < 0.05) (表2)。而吸收根的比根长受系统发育信号的影响较小且不显著(K < 1, p > 0.05); 根直径的系统发育信号K值接近1且p < 0.01, 表现出很强的系统发育保守性; 根组织密度和菌根侵染率受系统发育信号影响不显著(K < 1, p > 0.05) (表2)。
亚热带森林生态系统中树种丰富共存, 具有古老的系统发育过程(Ma et al., 2018), 根毛性状作为重要的原始发育特征在物种间具有显著差异(Datta et al., 2011)。本研究观察到在21个树种中, 7个树种存在根毛。广布种马尾松的根毛出现频率最高、根毛最长且最粗, 而演替早期、生长迅速的赤杨叶则根毛密度最高(表1)。大多数植物在演化过程采取了更加高效的水和矿质资源方式, 逐渐减弱了对根毛的依赖。本研究表明大部分亚热带树种并不发育根毛, 而是以其他方式获取土壤资源。根毛的发育与吸收根系的皮层细胞及植物分配的碳及相关的基因信号有关(Salazar-Henao et al., 2016; Zhang et al., 2018)。本研究中根毛直径和根直径具有较强的系统发育信号(表2), 说明根内细胞组织模式具有保守性。根毛密度和根毛长度具有较强的可塑性, 使根系觅食方式更加灵活(Rondina et al., 2019; Ruiz et al., 2020), 有利于植物响应土壤高资源异质性。例如, 桉属(Eucalyptus) 5种树木的根毛长度变异可达到3倍(Bichara et al., 2021); 热带森林中生长较快的树种发育更多的根毛(da Silva et al., 2021)。因此, 根毛的出现是植物应对环境资源变化的重要性状, 且不同物种间具有较大差异。本研究表明木本植物的根毛不是普遍存在的, 而且亚热带典型树种的根毛变异具有显著的种间格局, 这可能与土壤资源供应和演化历史有关。需要注意的是, 本研究中未观察到根毛的物种, 并不意味着该物种不存在根毛, 可能是单次采样的局限性所致。
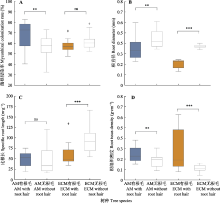
值得一提的是, AM树种根毛密度随着共生真菌侵染率的增加而增加(图5B), 表明在根毛存在的情况下, AM真菌更倾向于侵染根毛更密的树种。这种情况是可能存在的, Wu等(2016)发现枳橘被AM真菌侵染的吸收根的根毛密度显著高于没有被侵染的根; 或许AM真菌通过调节植物生理活动来影响根毛性状。ECM树种根毛直径与共生真菌侵染率呈负相关关系(图5D); 表明ECM真菌倾向于侵染根毛更细的树种。ECM树种真菌对根毛的影响较为复杂, 一方面根毛为ECM真菌提供侵染位点, 而另一方面菌丝产生的哈氏网会覆盖在根毛上从而影响根毛生长, 而且ECM真菌分泌物能抑制根毛的发育(Peterson & Farquhar, 1996; Novero et al., 2008; Rigas et al., 2013)。本研究中发现有根毛的ECM树种包括了被子植物麻栎、裸子植物马尾松和湿地松, 而根毛直径受系统发育信号影响大(表2); 因此, 菌根真菌侵染率和根毛直径的负相关关系很可能是由于马尾松和湿地松的根毛直径显著大, 而麻栎的根毛直径最小所造成的。总之, 根毛形态属性与菌根侵染率的关系因不同菌根类型而异, 而其中具体的相互作用机制需要进一步的研究。
本文的其它图/表
|