
|
|
||
|
被子植物蜜距的多样性及进化生态学研究
植物生态学报
2023, 47 (9):
1193-1210.
DOI: 10.17521/cjpe.2022.0445
蜜距将花蜜隐藏在距的深处, 多样的蜜距形态和长度吸引各种喙长的传粉动物访花, 促进传粉生态位形成, 这对造就物种多样性起到了重要作用。因此蜜距被认为是促进物种形成的关键创新性状, 是研究被子植物花部结构演化、植物-传粉者协同进化的理想材料。系统地总结蜜距植物及其形态特征和传粉/盗蜜者, 有助于全面了解这一性状在被子植物中的系统发育分布和进化生态学意义。该文对国内外植物分类数据库(检索词“距”或“spur”)和已发表文章(检索词“有距植物属中文名+传粉”或“属拉丁名+ pollinator/nectar robber”)进行系统的梳理和总结, 收集蜜距植物、距长、蜜距形态、传粉者、盗蜜者等信息。整理出具有蜜距的植物分布在被子植物13个目23个科271个属, 共计3 427种。其中蜜距植物种数最多的科为兰科(1 536种)、罂粟科(487种)、毛茛科(351种)、凤仙花科(284种)、堇菜科(197种), 它们占所有蜜距植物种数的83.3%。根据蜜距的发育来源, 蜜距可分为6种类型: 花被片蜜距、花瓣蜜距、萼片蜜距、被丝托蜜距、花萼蜜距和花冠蜜距。花瓣蜜距分布于212个属, 显著高于其他类型。对2 546种植物的蜜距长度分析发现: 凤仙花科的蜜距平均长度最长, 达23.8 mm; 千屈菜科的蜜距最短, 平均仅1.6 mm。形态上, 大部分蜜距具有不同程度的弯曲, 这可能有助于筛选合适的传粉者, 提高访花者的传粉效率; 单花蜜距数量有1-6个, 大部分为1个, 多个蜜距可能增加传粉者的访花时间; 部分兰科和堇菜科植物蜜距内部具有可能分泌花蜜的附属物。蜜距植物的传粉者主要为膜翅目、鳞翅目、部分长喙双翅目昆虫及鸟类; 盗蜜者主要为膜翅目昆虫。蜜距的长度和传粉者的喙长普遍存在地理镶嵌的协同进化关系, 进化生态学家提出了“达尔文机械模型”的假说来解释这种现象; 而当传粉者喙长不变, 距长随着传粉者种类改变时则用“传粉者转变模型”来解释蜜距的伸长。未来在蜜距的进化生态学研究中, 不仅要关注蜜距植物的距长和传粉者喙长的形态学特征及传粉生态学研究, 还应借助发育解剖学和基因组学, 以及新的技术手段, 从细胞的分裂、伸长, 功能基因, 非生物因子等各个方面, 探索蜜距演化的驱动因素。 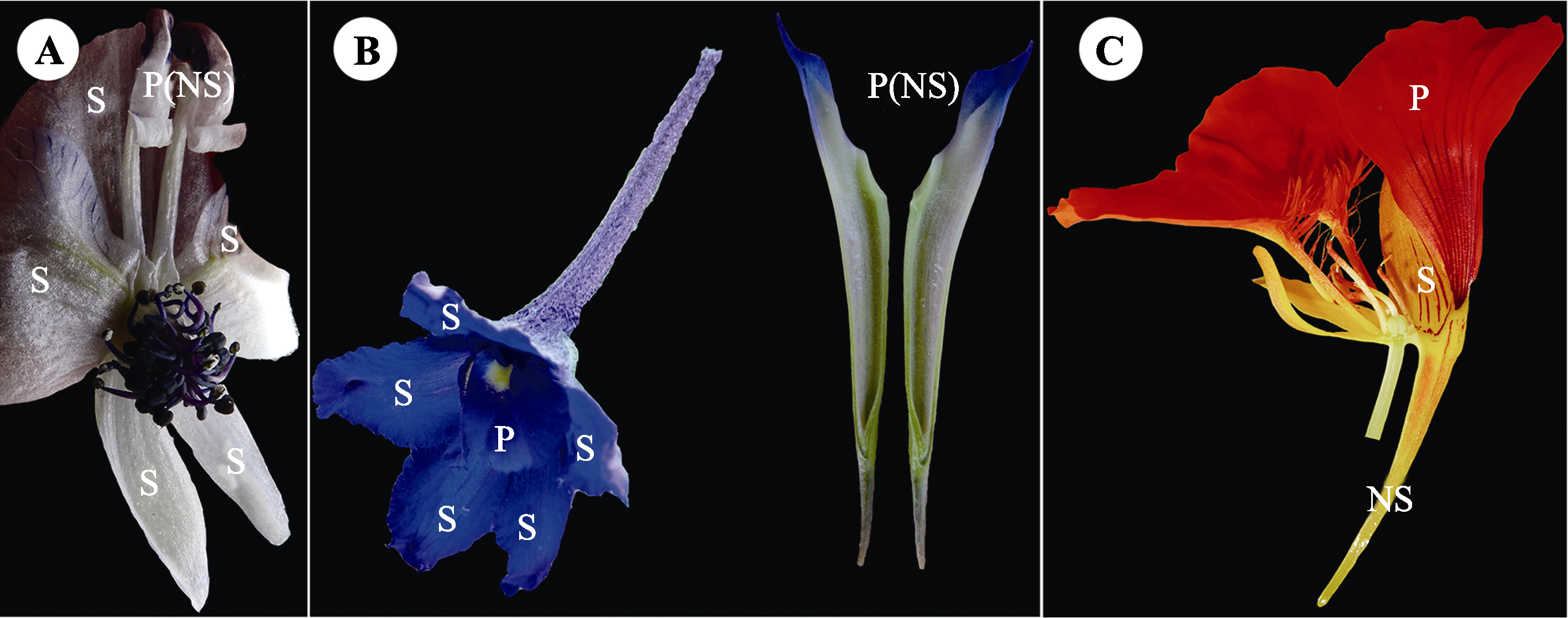 View image in article
图2
花瓣卷缩形成的蜜距(翠雀属、乌头属)和被丝托蜜距的结构解剖。A, 中甸乌头的花瓣蜜距。B, 中甸翠雀花的花瓣蜜距(被包裹在萼距里面)。C, 旱金莲的被丝托蜜距。NS, 蜜距; P, 花瓣; S, 萼片。拍摄者: 金晓芳。
正文中引用本图/表的段落
毛茛科中, 耧斗菜属(图1D)花瓣下方常向下延长形成蜜距, 极少数为囊状或不明显(Hodges, 1997), 如无距耧斗菜(Aquilegia ecalcarata); 乌头属(Aconitum) (图1C)、翠雀属与飞燕草属有2枚花瓣卷缩形成管状的蜜距(蜜腺位于蜜距末端), 并由1枚萼片包裹(图2A、2B) (Jabbour & Renner, 2012; Antoń & Kamińska, 2015), 乌头属少数种无距, 如空茎乌头(A. apetalum)等; 尾囊草属(Urophysa)的距瓣尾囊草(U. rockii)花瓣有短距, 而尾囊草(U. henryi)花瓣无距(Zhao et al., 2016)。
被丝托蜜距是指来源于被丝托的蜜距, 包括旱金莲科(图1L, 图2C)和千屈菜科, 一共2个属(表1)。旱金莲科仅旱金莲属(Tropaeolum)有蜜距, 在发育过程中, 被丝托的近轴区域发育最快, 在此处形成蜜距(Louis et al., 2001)。千屈菜科仅萼距花属(Cuphea)有蜜距, 该属植物被丝托呈管状, 部分种类具蜜距(Graham, 1990)。
本文的其它图/表
|



{kind=link}