
|
|
||
|
雷竹和青皮竹导管结构的轴向变化
植物生态学报
2024, 48 (7):
915-929.
DOI: 10.17521/cjpe.2023.0100
随着植株高度增加, 水分运输路径加长, 水分运输阻力增大。双子叶植物可以通过向下拓宽的木质部管道来补偿随着高度增加而增加的水分运输阻力。而单子叶植物无次生生长, 需终生使用同一套导管系统运输水分, 这会对其个体生长过程中维持木质部水力传输效率产生极大的限制。因此, 探明单子叶植物水力结构的轴向变化对于探讨该类物种的水分运输效率维持机制及其在自然界中能够广泛分布的原因显得尤为重要。该研究以树状单子叶植物雷竹(Phyllostachys violascens ‘Prevernalis’)和青皮竹(Bambusa textilis)为研究对象, 测定了植株茎干不同位置(即距茎尖不同距离)的导管大小、导管数量以及茎干外径等参数, 并进一步计算水力加权导管直径(Dh)、平均导管面积、导管密度、导管面积/导管密度等指标, 采用标准化主轴估计(SMA)的方法, 对各性状沿茎干轴向的变化规律及性状间的协变关系进行分析。研究结果显示, 雷竹和青皮竹从茎干顶端到基部, (1) Dh逐渐加宽、平均导管面积增大; (2)导管密度减小, 导管面积/导管密度增大; (3)导管密度与导管大小之间呈显著负相关关系。表明竹子从茎干顶部向基部, 导管大小逐渐拓宽, 单位木质部横截面积内的导管数量逐渐减少, 导管的大小与数量的变化是相互权衡的。 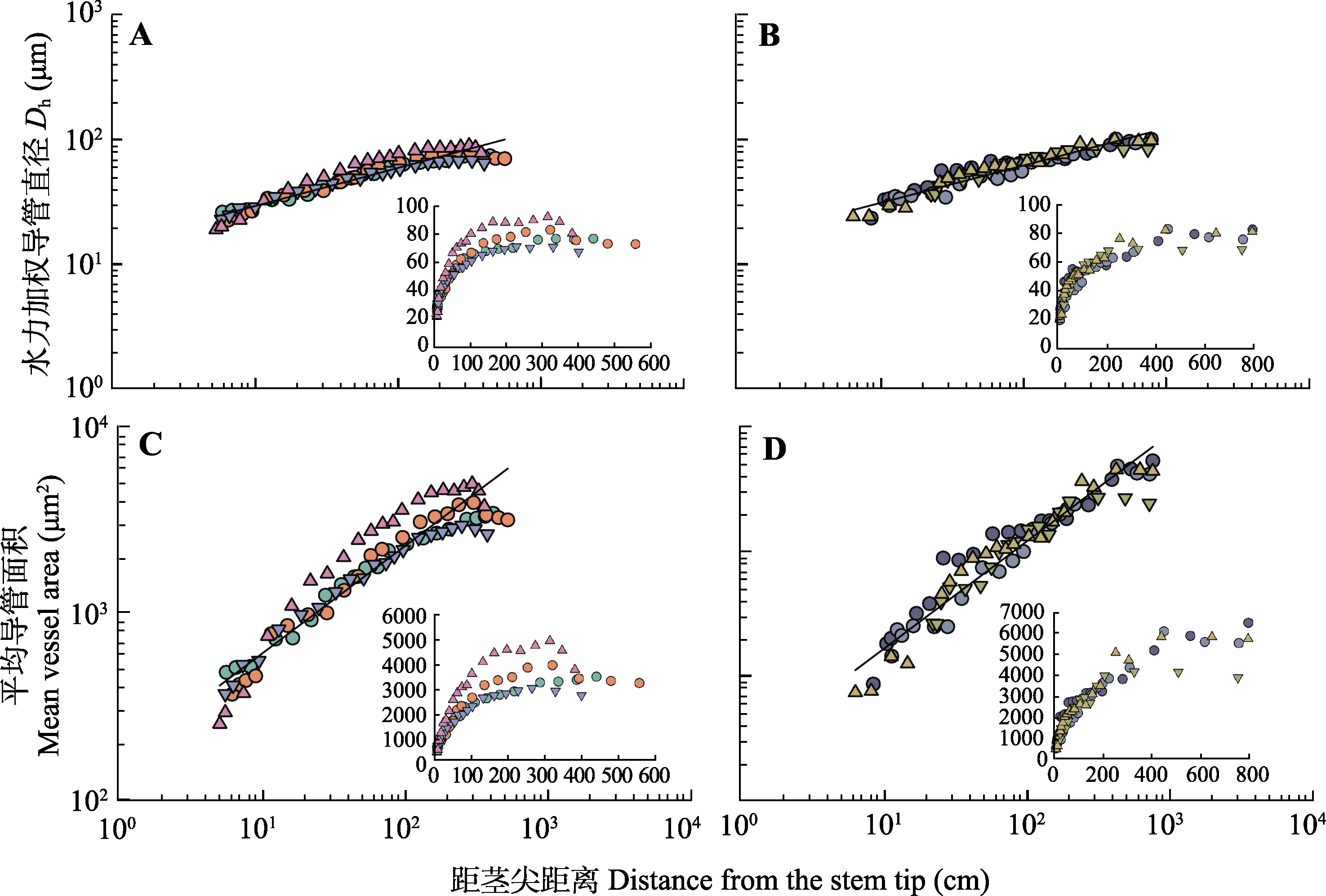 View image in article
图1
在雷竹(A、C)和青皮竹(B、D)的4个个体中, 水力加权导管直径及平均导管面积随距茎尖距离(对数-对数尺度)的变化关系。内插图为线性比例关系。
正文中引用本图/表的段落
雷竹和青皮竹各选4个个体, 测定茎干不同位置木质部导管的大小, 分析导管大小随距茎尖距离的变化规律。结果显示, Dh、平均导管面积与距茎尖距离均呈异速生长指数显著小于1的正异速生长关系, 即导管大小与距茎尖距离存在显著的幂函数关系, 这个幂函数关系解释了总变异的89%-92% (表1)。在物种水平上, 雷竹和青皮竹的Dh与距茎尖距离的关系斜率分别为0.305 (95% CI = 0.284-0.328) 和0.299 (95% CI = 0.279-0.320) (图1A、1B; 表1); 平均导管面积与距茎尖距离的关系斜率分别为0.574 (95% CI = 0.535-0.617)和0.573 (95% CI = 0.538-0.611) (表1), 表明从茎尖向基部, 导管大小逐渐变大, 但增大的速率小于距茎尖距离增加的速率。此外, 随着距茎尖距离的增加, 导管拓宽的速度逐渐减小, 曲线变得平缓(图1)。
植物木质部解剖特征影响其体内水分长距离运输, 从而影响整个植株的形态和功能。雷竹和青皮竹木质部导管的轴向解剖结果显示, 茎干木质部水力加权导管直径从茎干顶部向基部逐渐增宽, 异速生长指数约为0.3, 与双子叶木本植物的指数变化范围0.15-0.33基本一致(Anfodillo et al., 2006, 2012; Coomes et al., 2007; Petit et al., 2008, 2009, 2011; Lintunen et al., 2010; Bettiati et al., 2012; Lazzarin et al., 2016; Echeverría et al., 2019; Williams et al., 2019; Fajardo et al., 2020); 也与南美瓜多竹(Guadua angustifolia)的研究结果在95%置信区间内无显著差异(Petit et al., 2014), 均大于基于WBE的水力优化模型的预测值0.2 (West et al., 1999; Anfodillo et al., 2006)。与此同时, 平均导管面积与距茎尖距离也呈正的异速生长关系(图1C、1D; 表1), 与上述水力加权导管直径-距茎尖距离之间的关系类似。这些结果证明即便是在没有形成层、木质部维管系统都是一次性形成的树状单子叶植物中, 木质部管道的大小同样不符合管道模型理论所预测的管道大小在每级分枝都维持不变(Shinozaki et al., 1964a, 1964b; 范泽鑫等, 2005), 说明对于高大的树状植物, 无论是单子叶植物还是双子叶植物, 茎干内的木质部维管系统从顶部向基部按比例逐渐变大是一种普遍规律, 这是植物在自然选择作用下进化的结果, 在植物发育调控过程中具有重要作用。
虽然雷竹和青皮竹的导管大小(包括水力加权导管直径和平均导管面积)与距茎尖距离之间总体上呈一定的指数异速生长关系, 但导管大小随距茎尖距离的变化速度出现明显的拐点(距茎尖大约100-200 cm), 导管大小在茎干上部的变化速率显著快于茎干下部, 距茎尖200 cm以下导管大小变化幅度较小, 速率较慢, 而从距茎尖200 cm往上直到茎干顶端导管直径显著减小(图1)。该结果与Bactris gasipaes和瓜多竹这两种单子叶植物的变化趋势(Petit et al., 2014)一致, 也与木本双子叶植物研究中的导管快速拓宽通常发生在顶端几米处, 基部的导管直径变化可以忽略不计的结果(Anfodillo et al., 2006; Petit et al., 2010)类似。根据水力分割假说, 植物远端器官具有更大的水分运输阻力, 特别是叶片及承载叶片的枝条。在植物体总水力阻力中的大部分都局限在距顶端的前20-40 cm处或1 m内(Yang & Tyree, 1993; Becker et al., 2000; Petit et al., 2008, 2010; Petit & Anfodillo, 2009), 而茎干下部无侧枝, 对总水分运输阻力的影响与顶部相比相对较小(Lintunen et al., 2010); 茎干下部的导管较大, 水势差足以克服重力作用以维持正常的水分运输, 满足植株地上部分的水分运输需求, 导管继续增大成为多余, 反而会导致导管构建的碳投资增大。因此, 导管大小向基部方向的轴向拓宽可能是为了实现导管大小成本-收益最优化的结果(范泽鑫等, 2005; Anfodillo et al., 2006), 同时又可以克服因水分运输路径的延长而快速累积的阻力, 并降低导管空穴化栓塞的风险。
本文的其它图/表
|

{kind=link}