|
|
||||||||||||||||||||||||||||
|
雷竹和青皮竹导管结构的轴向变化
植物生态学报
2024, 48 (7):
915-929.
DOI: 10.17521/cjpe.2023.0100
随着植株高度增加, 水分运输路径加长, 水分运输阻力增大。双子叶植物可以通过向下拓宽的木质部管道来补偿随着高度增加而增加的水分运输阻力。而单子叶植物无次生生长, 需终生使用同一套导管系统运输水分, 这会对其个体生长过程中维持木质部水力传输效率产生极大的限制。因此, 探明单子叶植物水力结构的轴向变化对于探讨该类物种的水分运输效率维持机制及其在自然界中能够广泛分布的原因显得尤为重要。该研究以树状单子叶植物雷竹(Phyllostachys violascens ‘Prevernalis’)和青皮竹(Bambusa textilis)为研究对象, 测定了植株茎干不同位置(即距茎尖不同距离)的导管大小、导管数量以及茎干外径等参数, 并进一步计算水力加权导管直径(Dh)、平均导管面积、导管密度、导管面积/导管密度等指标, 采用标准化主轴估计(SMA)的方法, 对各性状沿茎干轴向的变化规律及性状间的协变关系进行分析。研究结果显示, 雷竹和青皮竹从茎干顶端到基部, (1) Dh逐渐加宽、平均导管面积增大; (2)导管密度减小, 导管面积/导管密度增大; (3)导管密度与导管大小之间呈显著负相关关系。表明竹子从茎干顶部向基部, 导管大小逐渐拓宽, 单位木质部横截面积内的导管数量逐渐减少, 导管的大小与数量的变化是相互权衡的。
表3
雷竹和青皮竹导管密度与导管大小的标准化主轴估计回归关系
正文中引用本图/表的段落
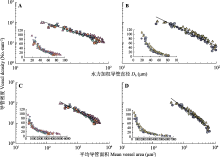
在雷竹与青皮竹中, 导管密度与Dh、平均导管面积均呈显著的负的异速生长关系。其中, 导管密度与Dh的关系斜率在两个竹种中均显著小于-1,分别是-1.541 (95% CI = -1.653- -1.437)和-2.241 (95% CI = -2.400- -2.091) (图3A、3B; 表3)。而导管密度与平均导管面积的关系斜率在雷竹中显著大于-1 (-0.819, 95% CI = -0.877- -0.765), 在青皮竹中显著小于-1 (-1.168, 95% CI = -1.24- -1.100) (图3C、3D; 表3), 表明在竹子茎干内导管的大小和数量之间存在权衡关系, 随着水力加权导管直径或平均导管面积的增大, 单位茎干质部面积上的导管数量逐渐减小, 但导管大小与数量的相对变化速率在不同竹种间存在差异。
进一步分析导管大小与导管密度的轴向变化关系, 结果显示雷竹和青皮竹植株茎干轴向的导管大小(水力加权导管直径或平均导管面积)与导管密度均呈显著的负相关关系, 与多数木本植物在种间水平上的研究结果(Carlquist & Hoekman, 1985; Wiemann et al., 1998; Baas et al., 2004; Preston et al., 2006; Wheeler et al., 2007; Sperry et al., 2008; Fan et al., 2009; Martínez-Cabrera et al., 2009; Lintunen et al., 2010; McCulloh et al., 2010; Poorter et al., 2010; Schuldt et al., 2013; Olson et al., 2014; Aparecido et al., 2015)一致。并且, 本研究中雷竹和青皮竹的水力加权导管直径与导管密度之间的异速生长指数为-1.541和-2.240 (图3A, 3B; 表3), 与棕榈种(Iriartea deltoidea)内水平(Renninger et al., 2013)以及针叶植物、热带散孔材树种和温带树种种间水平的异速生长指数变化范围-2.9- -1.4 (McCulloh et al., 2010; Aparecido et al., 2015)完全重合。这些结果表明无论是树状单子叶植物还是木本双子叶植物, 茎干内的导管大小和数量都遵循相似的变化规律, 两者沿茎干的轴向变化是相互权衡的。这种权衡关系可能反映了植物在最大化水力传导面积与机械支撑和存储之间的权衡, 即水分运输有效性和机械安全性之间的权衡。因为, 一方面导管大小和密度的变化会直接影响到茎木质部水分运输的效率和安全性。大导管虽然具有较高的水分运输效率, 但也存在更大的栓塞风险, 特别是在有冻融循环或干旱的生境中, 任何一个导管的栓塞都会导致冗余水分运输路径的减少(Tyree et al., 1994; Tyree & Zimmermann, 2002; Schenk et al., 2008)。相反, 在导管直径一定时, 导管密度即单位木质部面积上的导管数量越大, 意味着茎内维管系统的互联性越好(Preston et al., 2006), 当导管内充满水时植株的水分运输效率较高, 但当有部分导管发生气穴化栓塞时栓塞扩散的风险也会增加(Carlquist & Hoekman, 1985; Hacke & Sperry, 2001)。另一方面, 从生物物理学和机械支撑功能的角度看, 导管大小和密度的变化会通过影响木材内空腔空间的总量来影响木材密度, 进而影响到茎干的机械支撑强度。在茎干纤维基质中紧密排布较多的大导管, 虽然导水率较高, 但木材密度低, 木材太过脆弱, 在极端情况(如负压过大)下容易内裂(Niklas, 1994; Jacobsen et al., 2005)。而如果在纤维基质中稀疏排布一些小导管, 木材会非常致密, 机械支撑功能强, 但导水率很低, 植株需要将大量碳分配到边材中以满足其水力需求(Taneda & Tateno, 2004)。所以, 导管又大又多或者又少又细这两种极端情况对植物体都不利, 茎木质部的维管系统结构要么是少量的大管道, 要么是大量的小管道。从茎干基部到顶部导管大小与密度的相互权衡变化的潜在优势在于, 在兼顾植物水分运输效率和安全性以及机械支撑稳定性最优化的基础上, 还可以使单位生长投资的水力导度最大化, 并最大限度地减小基部与顶部小枝之间的导水力差异, 使水分分配更均匀, 将水分运输路径因植株长高而延长引起的对植株生产力和组织生长的水力限制最小化(Sperry et al., 2008)。
值得注意的是, 在本研究中尽管雷竹和青皮竹的导管大小与导管密度之间都是负的异速生长关系, 但异速生长指数在这两个竹种中相差较大。导管大小和导管密度之间的关系要受到导管腔面积在木质部面积中的占比(即导管腔面积/木质部面积)的影响, 这个比值的最大值肯定是略小于1 (即整个木质部横截面不可能都被导管腔占据) (Sperry et al., 2008; Zanne et al., 2010)。根据“填充限制(packing limit)”假说, 假设木质部导水面积沿茎干轴向是不变的, 并且边材内的管腔是由平均直径的导管所构成, 那么导管密度与导管直径之间的幂函数关系指数为–2, 即VD
本文的其它图/表
|