
|
|
||
|
雷竹和青皮竹导管结构的轴向变化
植物生态学报
2024, 48 (7):
915-929.
DOI: 10.17521/cjpe.2023.0100
随着植株高度增加, 水分运输路径加长, 水分运输阻力增大。双子叶植物可以通过向下拓宽的木质部管道来补偿随着高度增加而增加的水分运输阻力。而单子叶植物无次生生长, 需终生使用同一套导管系统运输水分, 这会对其个体生长过程中维持木质部水力传输效率产生极大的限制。因此, 探明单子叶植物水力结构的轴向变化对于探讨该类物种的水分运输效率维持机制及其在自然界中能够广泛分布的原因显得尤为重要。该研究以树状单子叶植物雷竹(Phyllostachys violascens ‘Prevernalis’)和青皮竹(Bambusa textilis)为研究对象, 测定了植株茎干不同位置(即距茎尖不同距离)的导管大小、导管数量以及茎干外径等参数, 并进一步计算水力加权导管直径(Dh)、平均导管面积、导管密度、导管面积/导管密度等指标, 采用标准化主轴估计(SMA)的方法, 对各性状沿茎干轴向的变化规律及性状间的协变关系进行分析。研究结果显示, 雷竹和青皮竹从茎干顶端到基部, (1) Dh逐渐加宽、平均导管面积增大; (2)导管密度减小, 导管面积/导管密度增大; (3)导管密度与导管大小之间呈显著负相关关系。表明竹子从茎干顶部向基部, 导管大小逐渐拓宽, 单位木质部横截面积内的导管数量逐渐减少, 导管的大小与数量的变化是相互权衡的。 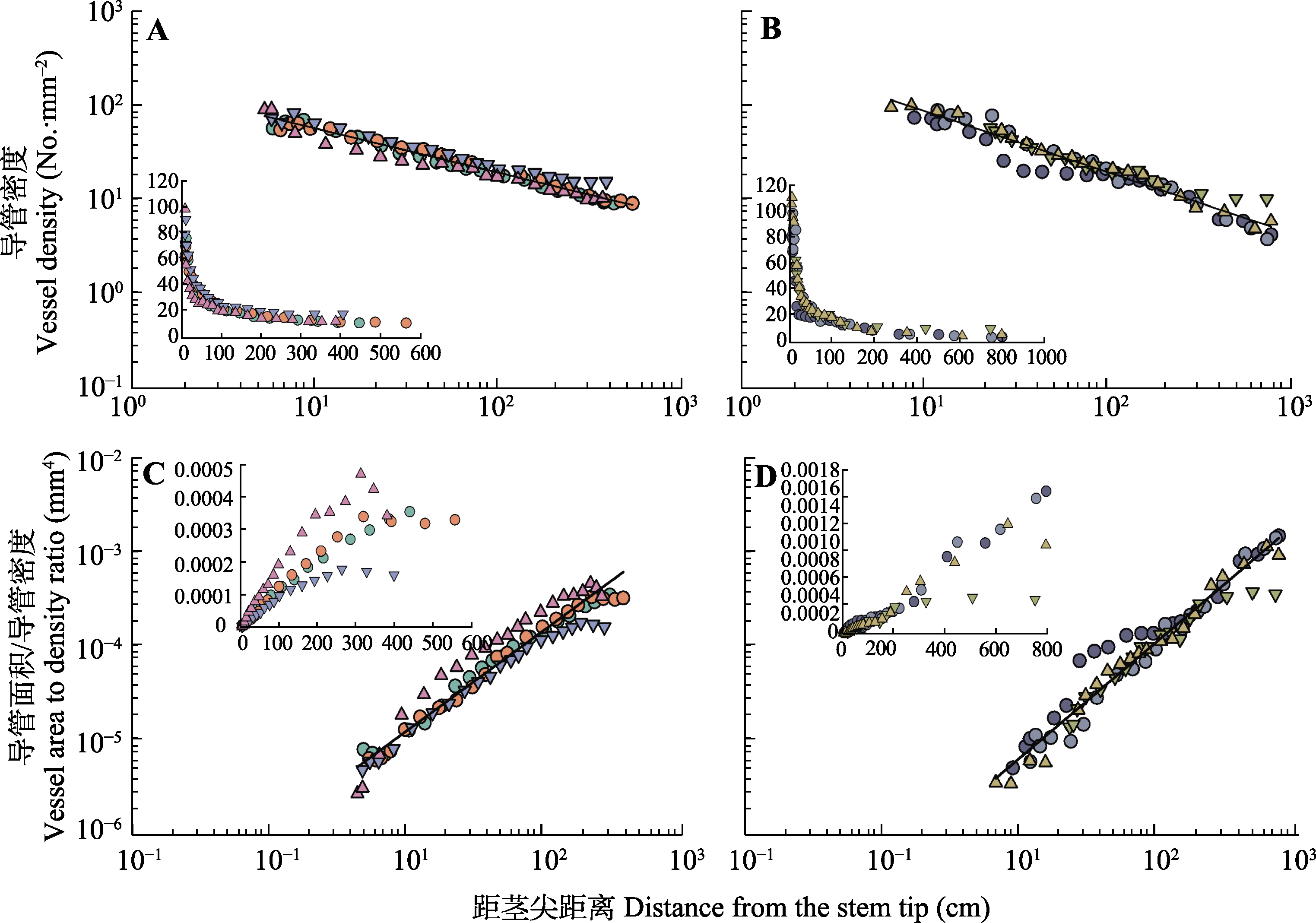 View image in article
图2
在雷竹(A、C)和青皮竹(B、D)的4个个体中, 导管密度及导管面积/导管密度随距茎尖距离(对数-对数尺度)的变化。内插图为线性比例关系。
正文中引用本图/表的段落
在雷竹和青皮竹每个物种4个个体内, 从茎尖向基部导管密度(即单位木质部面积内的导管数量)均呈减小的趋势。在物种水平上, 雷竹和青皮竹的导管密度与距茎尖距离均呈斜率显著大于-1的负异速生长关系, 其斜率分别为-0.471 (95% CI = -0.495- -0.448)和-0.669 (95% CI = -0.709- -0.632) (图2A、2B; 表2)。类似的结果在与茎干外径的关系中同样存在, 雷竹和青皮竹的导管密度与茎干外径的关系指数分别为-0.650 (95% CI = -0.688- -0.615)和-0.881 (95% CI = -0.924- -0.840) (表2)。这些结果表明从茎尖向基部, 随着距茎尖距离、茎干外径的增加, 导管密度逐渐减小, 但减小的速率小于距茎尖距离、茎干外径增加的速率。此外, 随着距茎尖距离的增加, 导管密度减小速度逐渐减小, 曲线变得平缓(图2)。同时, 雷竹和青皮竹的导管数量与距茎尖距离均呈斜率显著小于1的正异速生长关系, 其斜率分别为0.805 (95% CI = 0.764-0.867)和0.692 (95% CI = 0.654-0.732) (表2), 表明从茎尖向基部, 导管数量逐渐增加, 但增加的速率小于距茎尖距离增加的速率。雷竹和青皮竹的导管数量与茎干外径均呈正异速生长关系。其中, 雷竹的导管数量与茎干外径的关系斜率显著大于1, 为1.112 (95% CI = 1.087-1.138); 青皮竹的导管数量与茎干外径的关系斜率显著小于1, 为0.911 (95% CI = 0.883-0.940) (表2)。表明随着茎干外径的增加, 导管数量逐渐增加, 但相对变化速率在不同竹种间存在差异, 雷竹的增加速率大于茎干外径的增加速率, 青皮竹的增加速率小于茎干外径的增加速率。
导管面积/导管密度, 即平均导管面积与单位木质部面积导管数量的比值, 是一个用来反映木质部内导管组成的指标。较大的导管面积/导管密度表示木质部内的导管系统是由少数大导管组成, 较小的导管面积/导管密度表示木质部内的导管系统由较多小导管组成(Zanne et al., 2010)。导管面积/导管密度与距茎尖距离及茎干外径在两个物种中均呈显著正相关关系(图2; 表2)。在物种水平上, 雷竹和青皮竹的导管面积/导管密度与距茎尖距离之间的关系斜率分别为1.032 (95% CI = 0.979-1.089)和1.232 (95% CI = 1.168-1.298) (图2C、2D; 表2); 与茎干外径的关系斜率分别为1.427 (95% CI = 1.334-1.526)和1.622 (95% CI = 1.541-1.707) (表2)。表明随着距茎尖距离及茎干外径的增加, 导管面积/导管密度逐渐增大, 即从茎尖向基部, 木质部内的导管组成由较多小导管逐渐变为少数大导管。
植物组织尺度上的输水效率可以定义为单位横截面积上的水力传导率(体积流量/压力梯度)。某一给定木质部面积的导水率主要通过填充(即被侵填体堵塞)最少、最宽的管道来实现最大化, 这些管道也通常较长, 内部阻力较小。所以管道的最大直径、长度和每木质部横截面积的管道数量是导水效率的共同限制因子(Sperry et al., 2008)。以前大量研究主要集中在木质部输水管道直径由基部向顶部变细(呈现锥化)对消除导水力随路径长度增加而下降的影响(Enquist et al., 2000; Zaehle, 2005; Anfodillo et al., 2006; Mencuccini et al., 2007a), 忽略了另一个重要的事实, 即单位边材面积的管道数量(管道密度)随着管道变窄而增加(Sperry et al., 2008)。由于较宽的导管比较窄的导管占据更大的茎截面积, 导管直径沿茎的轴向变化必然会引起导管密度的变化(Olson et al., 2014)。在本研究中, 雷竹和青皮竹的导管密度由茎尖向基部逐渐减小, 其与距茎尖距离的异速生长指数分别是-0.471和-0.669 (图2A、2B; 表2)。该指数与Petit等(2010)研究的Eucalyptus regnans导管密度与距茎尖距离的异速生长指数(-0.49- -0.53)基本一致。同时导管密度也随着茎干外径由茎尖向基部的增加而减小(表2)。由于导管密度是指单位木质部面积的导管数量, 所以其变化主要由导管数量和木质部面积两个因素决定。雷竹和青皮竹导管数量与距茎尖距离均呈显著小于1的异速生长关系, 异速生长指数分别是0.805和0.692 (表2), 与Soriano等(2020)报道的辣木(Moringa oleifera) 0.656和香肉果(Casimiroa edulis) 0.731的异速生长指数结果相似。而雷竹和青皮竹木质部面积与距茎尖距离均呈显著大于1的异速生长关系, 其异速生长指数分别是1.257和1.343 (表1)。由此可见, 雷竹和青皮竹的导管数量随距茎尖距离的增加速度小于木质部面积随距茎尖距离的增加速度, 导致导管密度与距茎尖距离呈显著的负相关关系, 即导管密度的轴向变化是导管数量和木质部横截面积协同变化的结果。但也有报道认为, 与导管大小的向基式轴向增大相似, 导管密度的向基式轴向减小也可能是生长素浓度沿着轴向由基部向顶部递增的结果(Aloni, 1987; Anfodillo et al., 2012; Schuldt et al., 2013)。
木质部的负压系统本质上是危险的, 需要较多冗余的平行管道(Ewers et al., 2007)。位于植物远端的细枝面临着最大的水势负压, 同时暴露在最高水平的风引起的机械应力下(Schuldt et al., 2013), 茎干顶部的导管需要具有更强的抗气穴化栓塞或更容易修复栓塞的能力(Hacke et al., 2006), 也必须足够坚固以承受重力或环境对植物体施加的其他压力, 并避免因树液负压增大而爆裂(Sperry et al., 2008)。但另一方面, 在任何情况下, 为了提高水分运输的安全性而牺牲整体的水分运输效率又似乎会使植株处于竞争劣势。所以从基部到顶部的水分运输路径中, 当木质部面积逐渐减少, 木质部负压越来越大时, 为了达到必要的水分运输通道冗余并维持稳定的水分运输效率, 导管可能不得不变窄而数量增多, 以适应不断缩小的可用空间(McCulloh et al., 2010)。这也符合Tyree等(1994)提出的“考虑到碳投资的‘成本’, 自然选择的茎应该足够强壮, 能够提供静态支撑, 同时能够为植物所在的环境条件提供足够的水分输送”的观点。在本研究中, 雷竹和青皮竹的导管面积/导管密度与距茎尖距离呈显著正相关关系 (图2C、2D; 表2)也进一步表明, 从茎干顶部向基部, 木质部内的维管系统确实是由较多小导管逐渐变为少量大导管。茎干顶部更高的导管密度意味着单位面积木质部内具有更多的水分运输路径, 当其中部分导管因负压增大而栓塞导致水分运输功能丧失时, 还能有其他的水流路径来维持水分运输, 从而避免整个水分输导系统的崩溃, 一定程度上保证了水分输导的安全性(Zimmermann, 1982)。较低的导管面积/导管密度也反映在茎干顶部, 植株将采取更为保守的策略(Zanne et al., 2010)。而茎干基部木质部面积比顶部大, 导管面积/导管密度比较高, 表明木质部维管系统由少数大导管组成, 从生物物理学和生理学角度上看, 这既可以提高水分运输效率, 同时又使茎木质部横截面上具有更多的纤维和硬化细胞, 增强茎对冠层的机械支撑。对椰子和Iriartea deltoidea等单子叶植物的相关研究也表明, 随着植株高度的增加和茎干基部年龄的增长, 茎干基部的细胞硬度, 组织密度、硬度和强度逐渐增加, 并且在茎缘和茎基部最大, 以满足株高增长过程中的结构需求, 保证整个植株的机械稳定性(Rich, 1987; Aparecido et al., 2015)。
本文的其它图/表
|

{kind=link}