




在全球环境变化、人类经济活动和栖息地改变等因素驱动下, 全球物种灭绝不断加剧, 生态系统功能受到严重威胁。这些全球变化因子既可直接影响生态系统功能, 又可通过改变生物多样性而对生态系统功能产生间接作用。在这一背景下, 生态系统能否长期可持续地维持功能和服务, 有赖于生态系统自身的稳定性。因此, 理解生态系统稳定性及其维持机制, 对生态系统可持续管理与保护政策的制定具有重要科学参考意义。
早在20世纪50年代, 生态学家就开始关注生态系统的稳定性问题, 提出了一些重要认识和假说。Odum (1953)和Elton (1958)根据野外观测数据, 认为简单生态系统具有较低的时间稳定性和抵抗入侵的能力。同期, MacArthur (1955)根据食物网理论, 提出稳定性随着能量通路复杂性的增加而增强的观点。这一多样性促进稳定性的观点, 在之后很长时间内被人们广泛接受, 即认为物种多样性高的生态系统中不同物种相互补充抵御自然环境中的扰动, 从而增加生态系统的稳定性。然而, 1973年May发表了一项研究结果, 基于随机矩阵理论从数学上推导得出: 生态系统复杂性越高(即物种多样性越高, 种间连接越多、越强), 则物种实现稳定共存的概率越低。这一复杂性削弱稳定性的结论, 挑战了早期关于稳定性的直观认识, 开启了生态学界近半个世纪的“多样性-稳定性”之争(McCann, 2000)。
关于稳定性议题, 国内学者已发表了多篇综述(具体见下文), 本文在前期工作基础上, 重点阐述近年来关于稳定性内涵的最新理论认识, 以及多样性-稳定性关系的多维度和多尺度理论扩展。首先, 稳定性是一个多维概念, 有多种不同定义方法, 对此国内已有学者做过综述, 涉及稳定性的定义、机制和测度方法, 以及对应的干扰类型(马风云, 2002; 柳新伟等, 2004)。近期研究在稳定性内涵、不同指标间内在关系方面取得了新的重要理论进展, 本文将作具体介绍。此外, 关于多样性-稳定性关系及其机制, 国内外开展了大量实验和理论研究, 在20世纪90年代已经引起国内研究者的关注和讨论, 对问题发展、有关概念的内涵和外延进行了比较全面的回顾和辨析(黄建辉和韩兴国, 1995; 刘增文和李雅素, 1997), 国内学者对相关理论假说、经典实验有多篇综述文章(王国宏, 2002; 高东和何霞红, 2010; 张景慧和黄永梅, 2016)。本文梳理了最近关于生物多样性实验开展的多项整合分析研究, 及在多维度框架下开展的多样性-稳定性关系研究。以往关于稳定性的研究主要关注局域尺度, 而近期研究发展了新的多尺度理论框架, 最后本文详细探讨了稳定性的尺度依赖、多样性-稳定性的多尺度关系等新议题。基于对这些最新进展的系统梳理, 本文提出了几个有待进一步研究的问题和方向, 以期为后续相关研究提供参考。
1 生态系统稳定性的内涵与度量指标
1.1 稳定性的内涵
从系统外部讲, 扰动(perturbation)或干扰(disturbance)是指影响生态系统结构和动态的生物和非生物环境改变(Donohue et al., 2016)。不同扰动方式可从强度、时序特征(频率和持续时间等)、空间分布、作用方式、来源等方面来理解和区分。传统上可以将扰动分为脉冲扰动(pulse perturbation)、胁迫扰动(press perturbation)和环境随机扰动(environmental stochasticity)或称噪音扰动(noise)。脉冲扰动指持续时间短、一次性的剧烈环境变化, 如病虫害爆发、台风、火烧等自然灾害; 胁迫指持续时间长但相对缓和的环境变化, 如持续几十年的捕捞压力等; 环境随机扰动指生物和非生物环境中的随机因素导致的持续环境波动(Kéfi et al., 2019)。自然界中发生的干扰往往是这3种典型干扰模式的组合, 呈现出更加多样的特征, 例如全球气候变化造成的干扰就同时包含脉冲(极端降水事件)、胁迫(气温上升)与噪声(降水持续波动)。在不同干扰模式下生态系统的响应模式也不同, 因此要理解某一稳定性指标需要理解其背后的干扰模式。文献计量分析表明, 多数关于稳定性的研究考虑脉冲扰动(22%)和胁迫扰动(32%), 环境随机扰动占比只有约5%, 其余为移除扰动对照的研究(Kéfi et al., 2019)(图1); 大多数研究只考虑一种干扰因素, 大量的研究从单一维度观点出发, 理论研究中占比约83%, 实证研究中占比约80% (Donohue et al., 2016)。
图1
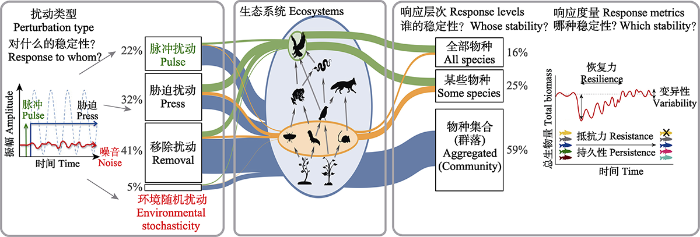
图1
评价生态系统稳定性需要确定扰动类型、生态系统、响应层次和响应度量。图上的百分比和不同颜色连接曲线的粗细表示在所引文献数据库中研究的数量比例(修改自Donohue et al., 2016; Kéfi et al., 2019)。
Fig. 1
Assessing stability in ecological systems requires identifying the perturbation type, ecosystem component, response level, and response metrics. The percentages and the width of the colored linking lines indicate the percentage of studies included in the references database (adapted from Donohue et al., 2016; Kéfi et al., 2019).
从系统内部讲, 生态系统对干扰的响应是一个多层次、多尺度的过程, 度量生态系统稳定性在实际操作中反映为量化特定层次和尺度的响应方式。具体地, 度量生态系统稳定性的指标既可以是生态系统总体的特征, 例如物种丰富度、生产力等群落层面的指标, 也可以是更精细的系统结构和组成特征, 例如均匀度、种群大小等物种层面的指标。对于不同响应层次, 群落层面的研究最多, 占比约达59%, 其余为物种层面的响应(Kéfi et al., 2019)(图1)。最近理论研究揭示了物种多度组成对稳定性指标的影响, 指出群落整体层面的变异性等指标主要由常见种决定, 而物种层面的恢复力主要由稀有种决定(Haegeman et al., 2016; Arnoldi et al., 2019)。考虑生态系统的空间属性, 稳定性指标可以分为区域(regional)或集合群落水平的指标和局部(local)群落水平的指标。研究生态系统稳定性问题的时候, 需要分别明确扰动类型以及生态系统的响应层次, 并且区分稳定性类型(图1)。
1.2 稳定性的多维度量
生态系统稳定性囊括了生态系统各个方面的动力学特征, 是一个多角度、多维度的概念, 在实际研究中具体化为多样化的定义和度量指标。根据Grimm和Wissel (1997)统计, 生态学文献中使用过超过160种关于稳定性的定义。Donohue等(2016)对稳定性相关文献做了计量分析, 发现不同稳定性定义的使用情况差异很大。大部分定义只在个别文献中提及, 且并未给出明确的度量方法。大多数关于稳定性的研究都集中于少量几个稳定性指标, 其中使用最多的有4种: 抵抗力(resistance)、恢复力(resilience)、持久性(persistence)和变异性(variability)(图1, 图2; 表1)。抵抗力指生态系统抵抗外界干扰并使自身的结构和功能保持原状的能力, 一般对应于脉冲或胁迫扰动, 即既包括一次性施加的扰动也包括长期的外部压力。恢复力指生态系统在遭到外界干扰因素的破坏以后恢复到原状的能力, 一般对应于脉冲扰动, 如病虫害爆发、人为活动、极端气候等非正常外力的短期内一次性施加。持久性指生态系统在一定边界范围内, 维持某一特定状态的持续时间, 一般对应于环境随机或胁迫扰动。变异性是指生态系统或种群的某一属性(如生物量)随时间波动的大小, 变异性越小, 稳定性越高, 对应于环境随机扰动。生态学研究中, 常用变异性的倒数表示稳定性, 称为时间稳定性(temporal stability)或不变性(invariability) (Tilman et al., 2006; Wang et al., 2017)。
图2
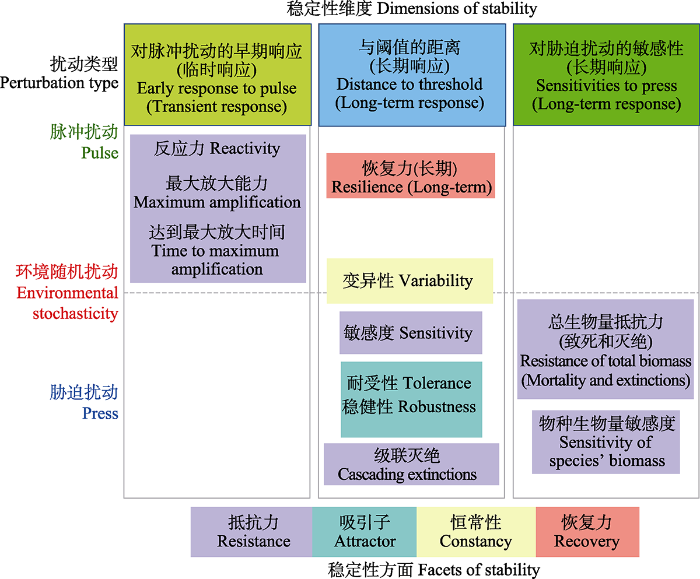
表1
稳定性主要的测度指标及其定义、对应扰动形式、响应层次和维度分类(
Table 1
| 稳定性指标 Stability metrics | 定义 Definition | 对应扰动形式 Perturbation type | 响应层次 Response level | 维度分类 Dimensionality |
|---|---|---|---|---|
| 抵抗力 Resistance | 系统抵抗扰动并使自身的结构和功能保持原状的能力。 The ability of the system to resist perturbations and keep its structure and function intact. | 胁迫扰动 Press | 物种、群落 Species, community | 对胁迫扰动的敏感性 Sensitivities to press |
| 恢复力 Resilience | 系统在遭到扰动因素的破坏以后恢复到原状的能力。 The ability of the system to return to its original state after being destroyed by perturbations. | 脉冲扰动 Pulse | 物种、群落 Species, community | 与阈值的距离 Distance to threshold |
| 持久性 Persistence | 在一定边界范围内, 系统保持恒定或维持某一特定状态的持续时间。 The duration of the system remains constant or maintains a certain state for the duration within a certain boundary. | 环境随机扰动 Environmental stochasticity | 群落 Community | 对胁迫扰动的敏感性 Sensitivities to press |
| 变异性 Variability | 系统随时间受扰动作用下产生的变化幅度。 The amplitude of change caused by perturbation in the system over time. | 环境随机扰动 Environmental stochasticity | 物种、群落 Species, community | 与阈值的距离 Distance to threshold |
| 反应力 Reactivity | 系统受外部扰动状态放大时的最大瞬时速率。 Maximum instantaneous rate at which perturbations can be amplified. | 脉冲扰动 Pulse | 群落 Community | 对脉冲扰动的早期响应 Early response to pulse |
| 最大放大能力 Maximum amplification | 系统受脉冲扰动后状态放大到最大幅度的能力。 The ability by which the perturbation that grows the largest is amplified after a pulse perturbation. | 脉冲扰动 Pulse | 群落 Community | 对脉冲扰动的早期响应 Early response to pulse |
| 达到最大放大的时间 Time to maximum amplification | 系统受脉冲扰动后状态达到最大放幅度的时间。 Time to achieve the maximum amplification after a pulse perturbation. | 脉冲扰动 Pulse | 群落 Community | 对脉冲扰动的早期响应 Early response to pulse |
| 敏感度 Sensitivity | 系统受胁迫扰动下物种生物量的平均变化范围。 Average change in the biomass of species after a press perturbation. | 胁迫扰动 Press | 物种、群落 Species, community | 与阈值的距离 Distance to threshold |
| 耐受性 Tolerance | 系统所有物种收到胁迫扰动下导致出现至少一个物种灭绝的全局或局部的最小致死增长量。 Minimum global increase in mortality that leads to at least one extinction under press perturbation. | 胁迫扰动 Press | 物种 Species | 与阈值的距离 Distance to threshold |
| 稳健性 Robustness | 将存活物种数量减少到原始数量的一半以下所需的主动灭绝的数量。 Number of actively performed extinctions required to reduce the number of surviving species to 50% of the original number. | 胁迫扰动 Press | 物种 Species | 与阈值的距离 Distance to threshold |
| 级联灭绝 Cascading extinctions | 首次灭绝后次生灭绝发生的平均数量。 Average number of secondary extinctions following first extinction. | 胁迫扰动 Press | 物种 Species | 与阈值的距离 Distance to threshold |
鉴于稳定性指标的多维性, 近期研究试图揭示不同稳定性指标间的关系, 并取得了一系列重要进展。Donohue等(2013)基于对海岸带生态系统的野外控制实验研究, 发展了多维度稳定性的概念框架。通过模拟移除捕食者和初级消费者, 探究了持久力(灭绝或入侵物种数)、抵抗力、时间变异性、空间变异性等多种稳定性的测度指标, 并利用相关性和主成分分析揭示不同指标间的关系。该研究发现, 虽然稳定性具有多维定义和指标, 但不同指标间具有一定的相关性, 因此该生态系统的稳定性本质上并不具有很高的维度。换句话说, 由于稳定性指标间具有强相关性, 因此当我们理解了一个稳定性指标, 也就理解了其他指标。但这项研究同时发现, 在外界环境干扰下, 不同稳定性指标之间的相关性大大减弱, 意味着稳定性的维度增加, 然而关于相关性丧失的原因, 文章作者并未给出具体解释, 有待进一步探究。这项研究开创性地提出了多维稳定性框架, 开启了关于生态系统稳定性的新研究方向。为了阐明不同维度的稳定性指标之间的关系, 以及干扰方式如何影响这些相关性, Radchuk等(2019)利用多种生态系统模型研究了不同干扰强度和类型对种群和群落水平稳定性维度的影响。结果表明, 干扰类型而非干扰强度对稳定性的维度有重要影响, 且这种影响主要发生在种群水平。该研究表明, 对稳定性的研究仍需要在多维框架下开展。
由于大多数稳定性指标都是从群落组成数据的时间序列计算得来的, 本质上是由生态系统内在的动力学模式决定的, 所以不同指标之间必然存在某种程度上的关联。例如Arnoldi等(2016)从数学上分析了理论研究中常用的几个稳定性指标, 揭示了其内在关联。特别地, 对单种群的简单生态系统来说, 脉冲干扰下的恢复力稳定性和环境噪声下的变异性呈正相关关系。Domínguez-García等(2019)利用计算机模拟研究了32种稳定性指标, 并基于相关性分析和聚类分析尝试将不同指标进行归类, 使得类内紧密相关, 类间相互独立。其研究聚类归纳得到, 不同稳定性指标对应的3个维度(图2; 表1): 对脉冲扰动的早期响应, 即脉冲扰动后群落在短期内与系统初始状态的差异; 与阈值的距离, 即系统越过参照阈值达到新的状态的可能性, 或表述为发生系统稳态转换的难易程度; 对胁迫扰动的敏感性, 即量化扰动后和扰动前群落之间总生物量和单个物种生物量变化。
2 多样性-稳定性关系研究进展
2.1 基于生物多样性实验的最新整合分析
20世纪90年代以来, 全球各地开展了一系列生物多样性控制实验和观测研究, 包括陆地生态系统、水生生态系统、微宇宙实验(microcosm)、中宇宙实验(mesocosm), 以及宏观的多地点联网的野外观测和控制实验。总体上, 这些研究表明生物多样性可以促进生态系统功能的稳定性, 但其对种群或功能群层面的稳定性可能有不同作用(Tilman et al., 2014)。其中, 稳定性往往被定义为较小的时间变异性, 对应的扰动方式为环境随机性。比如, Bai等(2004)对内蒙古自然草地生态系统生物量时间波动的长期观测研究表明, 物种和功能群在面对降水波动时呈现出非同步的互补动态, 使得生产力的稳定性从物种到功能群, 再到整个群落水平逐渐增加。基于Cedar Creek草地多样性实验, Tilman (1999)和Tilman等(2006)证实了物种多样性可增加草地群落层面生产力的时间稳定性, 但同时会降低群落内种群层面生产力的稳定性。基于欧洲多国联网多样性实验(BIODEPTH)的结果也表明, 物种多样性可增加群落层面生产力的稳定性, 但降低种群水平的稳定性; 而对于介于种群和群落之间的功能群水平, 物种多样性对其稳定性无显著作用(Hector et al., 2010)。然而, 基于野外观测数据的整合分析表明, 物种多样性对种群和群落水平的稳定性都有较弱的促进作用(Houlahan et al., 2018)。
此外, 有学者探讨了极端气候事件下的物种多样性与稳定性的关系。这里, 稳定性主要通过抵抗力和恢复力度量, 对应的干扰是极端气候事件带来的脉冲扰动。Isbell等(2015)基于46个草地植物多样性控制实验数据开展了整合分析, 考察生物多样性是否影响生态系统对极端干旱的抵抗力和恢复力。结果表明, 物种多样性可以增强生态系统对极端干旱发生时的抵抗力, 但对干旱后的恢复力没有影响。因此, 人为活动带来环境变化造成的生物多样性丧失可能会削减生态系统的稳定性, 而这种作用主要通过改变生态系统对极端环境事件的抵抗力实现。然而, 另一针对极端降水和干旱事件对地上生物量影响的整合分析却发现, 虽然物种丰富度可以增加群落生产力, 但并未增强群落生产力对极端事件的抵抗力和恢复力(de Boeck et al., 2018)。对此, 作者研究认为在极端气候变化下调节或紊乱多样性-稳定性关系的可能是生物和非生物因素的综合作用, 如群落聚合、多样性对群落性状的影响, 气候事件的重复周期等。
总而言之, 过去30年来关于多样性-稳定性的研究给出了一个重要预测, 即物种多样性丧失可能削弱生态系统的稳定性。这一结论在全球变化背景下具有重要意义。全球变化因子(比如温度和降水变化、氮沉降、CO2浓度、火干扰等)对生态系统结构和过程可产生重要影响, 进而改变了生态系统功能与稳定性。其中, 群落结构和物种组成的变化可能是导致生态系统功能和稳定性变化的关键因素(Hooper et al., 2012; Tilman et al., 2012)。近期研究基于12个多年控制实验数据, 就多个重要的环境变化因子如何影响生态系统生产力的时间稳定性做了整合分析(Hautier et al., 2015)。研究发现, 氮添加、水、CO2浓度、食草动物和火等环境变化因子引起的生物多样性变化与生产力稳定性的变化高度相关, 因此环境变化改变生态系统稳定性可能主要是通过改变物种多样性实现的。此外, 环境变化可能改变生物多样性的稳定化作用。基于全球草地营养添加实验网络(NutNet)的研究发现, 物种多样性可通过增加种群波动的异步性增强生态系统稳定性, 但这一作用在氮、磷、钾添加处理下大大减弱甚至消失(Hautier et al., 2014)。
2.2 多样性-稳定性关系机制: 理论模型
利用群落和生态系统模型, 理论生态学家对多样性-稳定性关系做了深入探讨, 针对多样性的稳定化机制提出了多种假说。为了解释经验分析结果, 这些理论研究大多针对时间稳定性。早期研究基于唯象模型(phenomenological model, 即不考虑种群动态过程), 发现不同物种间种群波动的非同步性可产生缓冲作用, 从而增强群落或生态系统水平的稳定性。也即, 生物多样性通过引入多样化的种群动态(作为对环境波动的响应), 可增强群落或生态系统面对环境波动的稳定性。借鉴于金融学中的名词, 这一作用机制被称为“保险效应” (insurance effect)(Yachi & Loreau, 1999)或“投资组合效应” (portfolio effect)(Tilman, 1999)。此外, 生物多样性可增强生态系统功能(如群落总生物量和生产力), 从而降低统计随机性对整体群落的影响, 对生态系统产生稳定化作用。这一作用被称为“超产效应” (overyielding effect)(Tilman, 1999)。对这些机制的 详细解读和数学原理, 可见张景慧和黄永梅(2016)一文。
为此, 生态学家进一步利用群落动态模型研究多样性-稳定性关系, 特别是种群动态和种间关系的作用(Ives & Hughes, 2002; Loreau & de Mazancourt, 2013)。比如在竞争群落中, 种间竞争可导致单个物种的种群波动增加, 但同时竞争使得不同物种间的时间波动有更大的异步性, 因此在群落水平上, 竞争对稳定性的作用由以上两种因素的相对重要性决定。Ives等(1999)基于一个种间对称的竞争模型(即不同物种具有相同的种群增长速率和种间竞争强度)发现, 竞争对种群稳定性的负作用与其对种间异步性的正作用相互抵消, 从而群落稳定性不受竞争强度的影响。Loreau和de Mazancourt (2013)利用一个多物种竞争模型, 发现在以下3种情况下生物多样性可通过增强保险效应和超产效应促进稳定性: (1)物种对环境波动的内在响应存在异步性; (2)物种对扰动响应速度存在种间差异; (3)种间竞争强度降低。这些动态模型为理解多样性的稳定化机制提供了动力学上的解释, 并为生物多样性实验研究提供了一种新方法, 用以量化多样性的不同稳定化机制的作用大小, 及其在不同生态系统间的差异(de Mazancourt et al., 2013)。此外, 不少学者利用复杂食物网模型研究多样性与稳定性关系, 徐光华等(2019)对此做了详细综述和分类归纳, 指出未来的模型将更能反映自然群落的复杂作用, 对稳定性多维度的刻画也会进一步拓展。
总体而言, 相比于唯象模型, 群落动态模型揭示了类似机制, 即生态系统稳定性取决于物种水平的稳定性、种间异步性引起的保险效应、多样性的超产效应引起的统计随机性下降等。虽然群落动态模型并未提出新的多样性作用机制, 但这类模型为理解多样性-稳定性关系提供了更深层次的解释, 避免了现象学模型中假设不相容的问题, 并为定 量预测不同生态学过程的稳定化作用提供了理论 框架。
2.3 多样性-稳定性关系的多维度
生物多样性和稳定性都具有多维定义和度量指标, 这是导致二者复杂关系的主要因素。近几十年来生物多样性的概念被扩展至多维度的测量指标, 主要包括: 分类学多样性、功能多样性和系统发育多样性。其中, 分类学多样性是最基本和最传统的多样性测度指标, 通过物种的分类信息和计数直接量化群落组成, 目前大多数多样性-稳定性研究中采用的都是分类学多样性。功能多样性刻画群落中物种功能性状的差异分化情况, 即影响物种功能的形态、生理或物候等性状的组成与变化(Dı́az & Cabido, 2001; Mason et al., 2005)。有研究者认为, 功能多样性对稳定性的影响比分类学多样性更直接。功能多样性或冗余表征了种间功能性状的相异性和相似性, 可分别通过生态位互补和保险效应影响生态系统稳定性。近年来, 关于功能多样性内涵及其与生态系统稳定性关系的研究日益增多, 深入揭示了生物群落性状分布与生态系统稳定性之间的内在联系(Zhao et al., 2016; Hallett et al., 2017; Yuan et al., 2019)。系统发育多样性刻画了群落中物种在系统发育树上的亲缘关系。在生态位保守性的假定下, 亲缘关系越近的种具有越相似的环境适应性。因此, 系统发育多样性越高(即种间亲缘关系越远), 群落中不同物种对环境波动的响应差异越大, 且由于相互竞争弱而对环境波动产生相对独立的响应, 从而可在一定程度上抵消种间波动, 增强群落整体的稳定性(Cadotte et al., 2012)。基于微宇宙实验的研究也表明, 食细菌性纤毛原生生物群落的遗传多样性可促进群落生物量的稳定性(Pu et al., 2014)。近期研究利用野外样地群落观测数据积累, 综合考察了3类多样性指标对草地生态系统稳定性的影响, 发现物种多样性和谱系多样性可增强种间异步性和群落的时间稳定性, 而功能多样性对稳定性作用较弱(Craven et al., 2018)。在大尺度上, Mazzochini等(2019)利用物种分布模型模拟南美洲巴西旱热落叶矮灌木林(Caatinga)木本植物的分布和多样性组成, 结合气温、降水、云量等气候变量, 发现区域尺度的植物群落遗传多样性与植被生产力稳定性呈正相关关系, 因此将遗传多样性加入现有区域和全球气候模式将有助于提高对未来生态系统功能预测的准确性。
以往多样性-稳定性关系研究大多关注某一稳定性指标, 近期研究则试图在多维度框架下探讨多样性-稳定性关系, 即基于同一生态系统考察不同稳定性指标与不同多样性之间的关系有何差异。这些研究既包括理论分析, 也包括实验研究。Ives和Carpenter (2007)利用种群动态模型研究了多种稳定性指标与物种多样性的关系。该研究模拟了脉冲扰动和胁迫扰动两种干扰方式, 考察了变异性、抵抗力、恢复力、多稳态系统的稳态数目、稳态类型(如点平衡、振荡或混沌)、物种维持等, 分析了这些稳定性指标与物种多样性的关系。该研究发现, 在不同稳定性定义下, 多样性-稳定性关系可呈现不同形式。比如, 多样性越高, 脉冲扰动后种群的渐进恢复速率越低, 但重复脉冲扰动下群落总生物量的不变性增强。虽然由于模型的简化性, 其结论在真实生态系统中不一定适用, 但该研究阐明了不同的稳定性指标可能得出不同结论的事实。近期两项实验研究都发现了类似结果, 即不同多样性指标与多样性的关系不同。Isbell等(2015)基于全球草地生物多样性实验数据, 考察了极端气候事件下生物多样性对生态系统生产力的时间稳定性、抵抗力和恢复力的影响。研究发现, 物种数为1-2种的低多样性群落中生产力在极端气候事件期间改变了约50%, 而物种数为16-32的高多样性群落仅变化约25%。因此, 生物多样性增加了生态系统的抵抗力, 但对恢复力无显著作用。由于多样性更高的生态系统具有较高的抵抗力, 因此最终体现出更高的时间稳定性。Pennekamp等(2018)以水生纤毛虫群落为研究对象进行了一项大型的双因子(温度×物种丰富度)控制实验, 发现物种多样性提高了生态系统功能的时间稳定性(即降低了时间变异性), 但降低了其对增温的抵抗力。这一控制实验研究揭示了生物多样性对生态系统稳定性不同指标存在相反的作用, 但是否可以推广到其他生态系统类型, 仍有待进一步的 验证。
2.4 多营养级生态系统的多样性-稳定性关系研究
在自然界中, 生态系统过程及其稳定性并不是由某个单一营养级的多样性决定的, 而是由不同营养级之间通过物质和能量传输的相互作用决定的(Brose, 2008)。根据May (1973)的预测, 物种之间的能量传递路径越复杂、传递效率越高, 生态系统的持久力和恢复力越弱。由于May的结论是基于随机网络结构给出的, 因此很多生态学家试图利用经验食物网数据探索自然生态系统稳定性的维持机制。这些研究发现自然界食物网中多样、非随机化的网络结构都对维持稳定性有重要作用, 包括模块化的网络结构(Thébault & Fontaine, 2010), 弱种间交互作用(McCann et al., 1998), 两两物种间作用的相关性(Jacquet et al., 2016), 物种个体大小分布(Brose et al., 2006)等。
实验研究方面, 早先多样性-稳定性研究多关注仅含初级生产者的生态系统, 但随着理论认识和实践手段的增强, 多营养级的多样性-稳定性实验和观测研究积累日渐丰富。其中, 由于水生原生动物和微生物群落种群动态周期短、易观测, 开展了许多有关的控制试验研究, 其结果为理论验证和发展提供了很多证据, 比如捕食过程如何改变多样性-稳定性关系(Jiang et al., 2009), 以及集合群落动态和稀有种对系统稳定性的作用等(Downing et al., 2014)。Jiang和Pu (2009)通过基于29项包含陆地和水生不同生态系统类型实验研究的整合分析发现, 单营养级和多营养级群落中的多样性-稳定性关系存在差异, 在多营养级群落中多样性具有更强的稳定化作用(即降低时间变异性)。最近基于多营养级浮游生物的实验还表明, 营养级内的水平多样性可促进生态系统的恢复力和抵抗力稳定性, 但营养级间的垂直多样性却有相反的作用(Zhao et al., 2019)。近期在脊椎动物的群落稳定性方面的研究也取得了新进展。Catano等(2020)在大陆尺度上对北美鸟类多样性观测数据的研究表明, 空间群落异质性与空间异步性和区域群落稳定性成正相关关系, 证实了鸟类在群落内部和群落之间的多样性直接影响鸟类群落稳定性。Yeager等(2020)利用沿海养殖池塘的水生生物和鱼类观测数据, 发现无脊椎动物群落和鱼类群落的稳定性均与池塘面积大小呈正相关关系, 但与近海距离呈负相关关系。其结果还表明, 池塘面积大小及与外部海域环境距离表征了水生物种扩散限制条件, 二者对多营养级水生生物群落稳定性的作用要强于物种丰度对群落稳定性的作用。
3 多样性-稳定性关系的多尺度拓展
多样性-稳定性研究为理解和预测生物多样性丧失的生态效应提供了重要科学基础。然而, 这些研究主要关注局域尺度, 空间范围量级为1-100 m2, 时间量级为1-10代种群世代(Gonzalez et al., 2020)。而自然界中的物种丧失发生在不同空间尺度上, 除了局域尺度的物种丰富度下降, 还包括区域尺度的物种灭绝和生境同质化(habitat homogenization)。为了应对栖息地破坏和气候变化等环境因素改变导致的区域生物多样性变化, 一个重要问题是如何将以往局域尺度的研究结论推广至大尺度, 这对理解大尺度物种丧失的生态效应、制定生态保护和管理策略具有重要意义。为此, 我们需要发展多尺度的稳定性研究框架, 并在此基础上探讨多样性的稳定化作用。
3.1 多尺度稳定性理论框架
基于生态等级理论框架, Jørgensen和Nielsen (2013)从一般意义上探讨了不同等级的生物和生态系统的稳定性之间的联系。该框架广泛地涉及了细胞、器官、个体、种群、群落、景观、生物圈等不同等级, 但由于假定同一等级内的不同组分是完全独立的, 因此在实际应用中具有一定的局限性。Wang和Loreau (2014, 2016)基于集合群落理论, 发展了一个时间稳定性的空间分解框架。类似于生物多样性的空间分解, 该框架提出了α、β和γ稳定性的概念。数学上, 区域或集合群落的γ稳定性可分解为局部α稳定性和β稳定性。具体地, α和γ稳定性分别指代了局域和区域尺度生态系统的时间稳定性, β稳定性指示了不同局域生态系统的时间波动的非同步性。从局部尺度到区域尺度, 生态系统稳定性逐渐增加, 从而产生正的稳定性-面积关系(Wang et al., 2017)。进一步地, 可类似地将局域生态系统稳定性分解为物种稳定性和种间异步性(Thibaut & Connolly, 2013)。因此, 最终区域生态系统稳定性可分解为物种稳定性、种间异步性和空间异步性的乘积(Wang et al., 2019)。这一分解框架表明, 通过研究不同生态因素对区域生态系统稳定性的3个分量的影响, 可以阐明区域尺度生态系统稳定性的决定机制(图3)。
图3
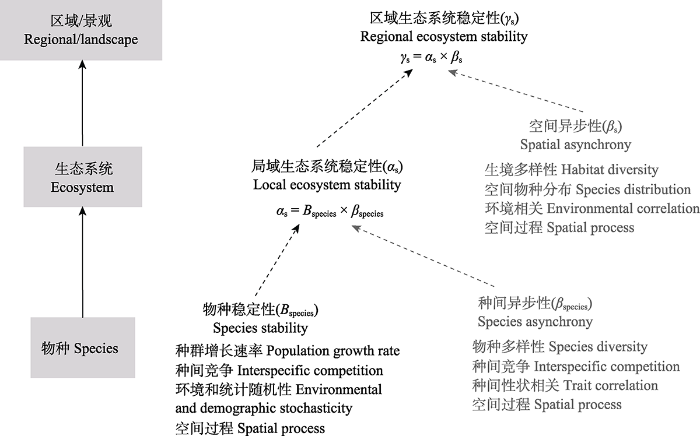
图3
不同尺度稳定性之间的内在关联。区域生态系统稳定性可以分解为局域稳定性和空间异步性的乘积, 局域稳定性可进一步分解为物种稳定性和种间异步性的乘积。不同生态学过程通过影响物种稳定性、种间异步性和空间异步性, 最终决定区域生态系统的稳定性(修改自Wang & Loreau, 2014)。
Fig. 3
Intrinsic links between stability at different scales. Regional ecosystem stability can be partitioned into the product of local ecosystem stability and spatial asynchrony, and local ecosystem stability can be further decomposed into the product of species stability and species asynchrony. Different ecological processes can influence the stability of regional ecosystems by affecting species stability, inter-species asynchrony, and spatial asynchrony (adapted from Wang & Loreau, 2014).
基于这一框架, Wang和Loreau (2016)使用竞争性集合群落模型研究了不同尺度的物种多样性与生态系统变异之间的关系。该模型表明, α多样性可增强局域生态系统的稳定性, 而β多样性可增强不同生态系统之间的空间异步性, 即β稳定性。因此, α和β多样性分别通过局域和区域尺度的保险效应为区域生态系统提供稳定化作用。该研究首次预测多样性-稳定性关系可发生在α、β和γ等不同尺度, 因此生物多样性和栖息地的丧失会破坏区域范围内的生态系统稳定性。该研究有助于理解全球环境变化(如环境同质化)和生物多样性丧失对生态系统稳定性的交互影响, 并对开展景观生态系统管理与保护具有实际意义。
3.2 经验研究
基于以上框架, 生态学家研究了自然和实验植物群落中不同尺度稳定性及其与多样性的关系。Wilcox等(2017)通过收集来自全球五大洲62个植物群落的数据, 定量评估了物种和局部群落间的异步性对集合群落功能稳定性的作用。该研究指出, 局域群落之间的异步性与β多样性无显著关系, 而是与物种在不同斑块间的异步波动有关, 这可能源于斑块之间的环境差异。Wang等(2019)利用美国新墨西哥州沙漠植物群落数据, 考察了从物种到群落、再到集合群落尺度的生产力稳定性的变化, 以及多样性如何塑造不同尺度的同步性和稳定性。结果显示, 植被生产力稳定性主要来自种间波动的异步性, 而群落之间的空间异步性作用相对较小。在局域和区域尺度, α和β多样性分别是维持种间和群落间异步性的关键因素。此外, 该研究还揭示了异步性的尺度推移规律, 即物种异步性从局部到区域尺度增大、空间异步性从物种到群落水平也增大。Zhang等(2019)在温带草原开展实验研究了氮添加对不同尺度的植物物种丰富度和群落生物量稳定的影响。研究发现, 局域尺度上, 氮添加会减少物种丰富度和种间异步性, 从而降低群落稳定性。但在区域尺度上, 氮添加没有改变局部群落之间的空间异步性, 表明局部群落间的动态差异可提供空间保险效应而维持区域水平的生态系统稳定性。
近期研究还将多尺度稳定性理论应用于全球粮食生产和海洋生态系统研究。Mehrabi和Ramankutty (2019)分析了全球粮食生产的稳定性, 将4种主要粮食作物(即水稻(Oryza sativa)、小麦(Triticum aestivum)、大豆(Glycine max)、玉米(Zea mays)), 在全球尺度的时间稳定性分解为局域稳定性(α稳定性)和不同地区之间的异步性(β稳定性)。研究发现, 近几十年来全球粮食作物产量的空间同步性下降, 从而增强了全球尺度作物产量的稳定性。该研究评估了不同情景下的全球粮食产量的稳定性, 指出在气候变化、人口增长和消费趋势变化下, 保持区域之间粮食生产的异步性是维持全球粮食安全的关键。Lamy等(2019)对在美国加利福尼亚南部深海海藻森林环境中的鱼类群落稳定性做了研究, 量化了不同系统组织层面(种群和群落)和空间尺度(局域和区域)的鱼类生物量稳定性和同步性。研究发现, 物种间的异步性或空间保险效应较强, 而不同群落之间的异步性或空间保险效应较弱, 这些效应可能与洋流和太平洋年际振荡有关系。不同物种对暖化水温和低营养条件的耐受性差异及其互补性可能是物种发挥物种空间保险效应的基础, 而由于洋流混合过程而引起的空间同步性导致了较低的空间保险效应。
4 结论和展望
自20世纪50年代以来, 多样性-稳定性关系一直是生态学研究的一个核心议题。Ives和Carpenter (2007)在其综述中指出, 多样性-稳定性之争持续了半个世纪却仍无定论, 表明该问题本身有很大的“稳定性”。近年来的多维度多尺度多样化的研究极大增进了对稳定性的内涵、其多维度和多尺度定义,以及多样性-稳定性关系的认识, 取得了一系列重要进展。特别地, 近期研究阐明了不同稳定性指标之间的内在关联, 揭示了不同稳定性指标与多样性的关系存在差异, 建立了多尺度的稳定性理论框架。这些结果为化解多样性-稳定性之争提供了新认识, 并对今后稳定性研究的开展有指导意义。
基于当前研究中的不足, 我们提出以下可能的研究方向和建议。第一, 目前关于稳定性的文献往往对其内涵缺乏明确界定, 笼统地用稳定性指代生态系统对某一特定扰动的特定响应, 这不利于阐明其作用机制以及不同研究之间的比较和整合, 因此建议后续研究对稳定性概念给出明确定义, 进一步探究多样性对不同维度的稳定性指标的作用机制(Kéfi et al., 2019)。第二, 理论研究需进一步整合稳定性与群落生态学过程, 建议加强物种共存机制如何影响多样性-稳定性关系(Tredennick et al., 2017; Gonzalez et al., 2020)、生物多样性影响生态系统功能与稳定性的关联机制(Cardinale et al., 2013)等方面的研究, 阐明食物网中跨营养级的垂直多样性如何与营养级内部的水平多样性交互作用并影响稳定性(Zhao et al., 2019), 发展新的基于功能性状的理论模型、整合物种共存、生态网络、生态系统功能、稳定性等研究领域(Zhang et al., 2013; Krause et al., 2014)。第三, 目前关于多样性-稳定性关系的研究主要在自然环境下开展, 而理解环境因子(如温度、降水、营养资源、火烧等)的变化对多样性-稳定性关系的影响, 对预测生态系统对全球变化的响应有重要意义(de Laender et al., 2016)。因此, 建议加强环境因子与生物多样性的交互实验研究, 阐明环境变化对多样性-稳定性关系的影响。第四, 如何将生态学实验和理论研究应用于生态系统管理与保护是一个关键问题(Manning et al., 2019)。近期发展的多尺度稳定性理论构建了不同尺度稳定性之间的关联, 为联系小尺度生态学研究与大尺度生态管理提供了一个可能框架, 但景观尺度的经验研究仍缺乏。随着生态系统监测手段的扩展和长时间数据的系统积累, 建议加强大尺度的多样性-稳定性关系的经验研究, 这将为揭示和量化自然生态系统中生物多样性的生态功能及其服务价值提供依据(Yu et al., 2010; He et al., 2019)。
参考文献
Resilience, reactivity and variability: a mathematical comparison of ecological stability measures
DOI:10.1016/j.jtbi.2015.10.012
PMID:26542944
[本文引用: 1]

In theoretical studies, the most commonly used measure of ecological stability is resilience: ecosystems asymptotic rate of return to equilibrium after a pulse-perturbation -or shock. A complementary notion of growing popularity is reactivity: the strongest initial response to shocks. On the other hand, empirical stability is often quantified as the inverse of temporal variability, directly estimated on data, and reflecting ecosystems response to persistent and erratic environmental disturbances. It is unclear whether and how this empirical measure is related to resilience and reactivity. Here, we establish a connection by introducing two variability-based stability measures belonging to the theoretical realm of resilience and reactivity. We call them intrinsic, stochastic and deterministic invariability; respectively defined as the inverse of the strongest stationary response to white-noise and to single-frequency perturbations. We prove that they predict ecosystems worst response to broad classes of disturbances, including realistic models of environmental fluctuations. We show that they are intermediate measures between resilience and reactivity and that, although defined with respect to persistent perturbations, they can be related to the whole transient regime following a shock, making them more integrative notions than reactivity and resilience. We argue that invariability measures constitute a stepping stone, and discuss the challenges ahead to further unify theoretical and empirical approaches to stability. Copyright © 2015 Elsevier Ltd. All rights reserved.
The inherent multidimensionality of temporal variability: How common and rare species shape stability patterns
DOI:10.1111/ele.v22.10 URL [本文引用: 1]
Ecosystem stability and compensatory effects in the Inner Mongolia grassland
DOI:10.1038/nature02850 URL [本文引用: 1]
Complex food webs prevent competitive exclusion among producer species
DOI:10.1098/rspb.2008.0718 URL [本文引用: 1]
Allometric scaling enhances stability in complex food webs
DOI:10.1111/ele.2006.9.issue-11 URL [本文引用: 1]
Phylogenetic diversity promotes ecosystem stability
Biodiversity simultaneously enhances the production and stability of community biomass, but the effects are independent
To predict the ecological consequences of biodiversity loss, researchers have spent much time and effort quantifying how biological variation affects the magnitude and stability of ecological processes that underlie the functioning of ecosystems. Here we add to this work by looking at how biodiversity jointly impacts two aspects of ecosystem functioning at once: (1) the production of biomass at any single point in time (biomass/area or biomass/ volume), and (2) the stability of biomass production through time (the CV of changes in total community biomass through time). While it is often assumed that biodiversity simultaneously enhances both of these aspects of ecosystem functioning, the joint distribution of data describing how species richness regulates productivity and stability has yet to be quantified. Furthermore, analyses have yet to examine how diversity effects on production covary with diversity effects on stability. To overcome these two gaps, we reanalyzed the data from 34 experiments that have manipulated the richness of terrestrial plants or aquatic algae and measured how this aspect of biodiversity affects community biomass at multiple time points. Our reanalysis confirms that biodiversity does indeed simultaneously enhance both the production and stability of biomass in experimental systems, and this is broadly true for terrestrial and aquatic primary producers. However, the strength of diversity effects on biomass production is independent of diversity effects on temporal stability. The independence of effect sizes leads to two important conclusions. First, while it may be generally true that biodiversity enhances both productivity and stability, it is also true that the highest levels of productivity in a diverse community are not associated with the highest levels of stability. Thus, on average, diversity does not maximize the various aspects of ecosystem functioning we might wish to achieve in conservation and management. Second, knowing how biodiversity affects productivity gives no information about how diversity affects stability (or vice versa). Therefore, to predict the ecological changes that occur in ecosystems after extinction, we will need to develop separate mechanistic models for each independent aspect of ecosystem functioning.
Local species diversity, β-diversity and climate influence the regional stability of bird biomass across North America
DOI:10.1098/rspb.2019.2520 URL [本文引用: 1]
Multiple facets of biodiversity drive the diversity- stability relationship
Patterns and drivers of biodiversity-stability relationships under climate extremes
DOI:10.1111/jec.2018.106.issue-3 URL [本文引用: 1]
Reintroducing environmental change drivers in biodiversity-Ecosystem functioning research
DOI:10.1016/j.tree.2016.09.007 URL [本文引用: 1]
Predicting ecosystem stability from community composition and biodiversity
DOI:10.1111/ele.12088
PMID:23438189
[本文引用: 1]
As biodiversity is declining at an unprecedented rate, an important current scientific challenge is to understand and predict the consequences of biodiversity loss. Here, we develop a theory that predicts the temporal variability of community biomass from the properties of individual component species in monoculture. Our theory shows that biodiversity stabilises ecosystems through three main mechanisms: (1) asynchrony in species' responses to environmental fluctuations, (2) reduced demographic stochasticity due to overyielding in species mixtures and (3) reduced observation error (including spatial and sampling variability). Parameterised with empirical data from four long-term grassland biodiversity experiments, our prediction explained 22-75% of the observed variability, and captured much of the effect of species richness. Richness stabilised communities mainly by increasing community biomass and reducing the strength of demographic stochasticity. Our approach calls for a re-evaluation of the mechanisms explaining the effects of biodiversity on ecosystem stability.© 2013 Blackwell Publishing Ltd/CNRS.
Vive la différence: plant functional diversity matters to ecosystem processes
DOI:10.1016/S0169-5347(01)02283-2 URL [本文引用: 1]
Unveiling dimensions of stability in complex ecological networks
DOI:10.1073/pnas.1904470116
PMID:31801881
[本文引用: 5]
Understanding the stability of ecological communities is a matter of increasing importance in the context of global environmental change. Yet it has proved to be a challenging task. Different metrics are used to assess the stability of ecological systems, and the choice of one metric over another may result in conflicting conclusions. Although each of the multitude of metrics is useful for answering a specific question about stability, the relationship among metrics is poorly understood. Such lack of understanding prevents scientists from developing a unified concept of stability. Instead, by investigating these relationships we can unveil how many dimensions of stability there are (i.e., in how many independent components stability metrics can be grouped), which should help build a more comprehensive concept of stability. Here we simultaneously measured 27 stability metrics frequently used in ecological studies. Our approach is based on dynamical simulations of multispecies trophic communities under different perturbation scenarios. Mapping the relationships between the metrics revealed that they can be lumped into 3 main groups of relatively independent stability components: early response to pulse, sensitivities to press, and distance to threshold. Selecting metrics from each of these groups allows a more accurate and comprehensive quantification of the overall stability of ecological communities. These results contribute to improving our understanding and assessment of stability in ecological communities.
Navigating the complexity of ecological stability
DOI:10.1111/ele.12648
PMID:27432641
[本文引用: 6]
Human actions challenge nature in many ways. Ecological responses are ineluctably complex, demanding measures that describe them succinctly. Collectively, these measures encapsulate the overall 'stability' of the system. Many international bodies, including the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services, broadly aspire to maintain or enhance ecological stability. Such bodies frequently use terms pertaining to stability that lack clear definition. Consequently, we cannot measure them and so they disconnect from a large body of theoretical and empirical understanding. We assess the scientific and policy literature and show that this disconnect is one consequence of an inconsistent and one-dimensional approach that ecologists have taken to both disturbances and stability. This has led to confused communication of the nature of stability and the level of our insight into it. Disturbances and stability are multidimensional. Our understanding of them is not. We have a remarkably poor understanding of the impacts on stability of the characteristics that define many, perhaps all, of the most important elements of global change. We provide recommendations for theoreticians, empiricists and policymakers on how to better integrate the multidimensional nature of ecological stability into their research, policies and actions.© 2016 John Wiley & Sons Ltd/CNRS.
On the dimensionality of ecological stability
DOI:10.1111/ele.12086
PMID:23419041
[本文引用: 1]
Ecological stability is touted as a complex and multifaceted concept, including components such as variability, resistance, resilience, persistence and robustness. Even though a complete appreciation of the effects of perturbations on ecosystems requires the simultaneous measurement of these multiple components of stability, most ecological research has focused on one or a few of those components analysed in isolation. Here, we present a new view of ecological stability that recognises explicitly the non-independence of components of stability. This provides an approach for simplifying the concept of stability. We illustrate the concept and approach using results from a field experiment, and show that the effective dimensionality of ecological stability is considerably lower than if the various components of stability were unrelated. However, strong perturbations can modify, and even decouple, relationships among individual components of stability. Thus, perturbations not only increase the dimensionality of stability but they can also alter the relationships among components of stability in different ways. Studies that focus on single forms of stability in isolation therefore risk underestimating significantly the potential of perturbations to destabilise ecosystems. In contrast, application of the multidimensional stability framework that we propose gives a far richer understanding of how communities respond to perturbations.© 2013 Blackwell Publishing Ltd/CNRS.
Multiple diversity- stability mechanisms enhance population and community stability in aquatic food webs
Biodiversity has been shown to increase the temporal stability of community and ecosystem attributes through multiple mechanisms, but these same mechanisms make less consistent predictions about the effects of richness on population stability. The overall effects of biodiversity on population and community stability will therefore depend on the dominant mechanisms that are likely to vary with the nature of biodiversity loss and the degree of environmental variability. We conducted a mesocosm experiment in which we generated a gradient in zooplankton species richness by directly manipulating dominant species and by allowing/preventing immigration from a metacommunity. The mesocosms were maintained under either constant or variable nutrient environments. Population, community, and ecosystem data were collected for five months. We found that zooplankton population and community stability is enhanced in species-rich communities in both constant and variable environments. Species richness increased primarily through the addition of species with low abundance. The communities that were connected to a metacommunity via immigration were the most diverse and the most stable, indicating the importance of both metacommunity dynamics and rare species for stability. We found little evidence for selection effects or overyielding as stabilizing forces. We did find support for asynchronous dynamics and statistical averaging, both of which predict destabilizing effects at the population level. We also found support for weak interactions, which predicts that both populations and communities will become more stable as richness increases. In order to understand the effects of biodiversity loss on stability, we will need to understand when different stabilizing mechanisms tend to operate but also how multiple mechanisms interact.
Research advances on biodiversity and ecosystem stability
生物多样性与生态系统稳定性研究进展
Scaling-up biodiversity-ecosystem functioning research
DOI:10.1111/ele.13456
PMID:31997566
[本文引用: 2]
A rich body of knowledge links biodiversity to ecosystem functioning (BEF), but it is primarily focused on small scales. We review the current theory and identify six expectations for scale dependence in the BEF relationship: (1) a nonlinear change in the slope of the BEF relationship with spatial scale; (2) a scale-dependent relationship between ecosystem stability and spatial extent; (3) coexistence within and among sites will result in a positive BEF relationship at larger scales; (4) temporal autocorrelation in environmental variability affects species turnover and thus the change in BEF slope with scale; (5) connectivity in metacommunities generates nonlinear BEF and stability relationships by affecting population synchrony at local and regional scales; (6) spatial scaling in food web structure and diversity will generate scale dependence in ecosystem functioning. We suggest directions for synthesis that combine approaches in metaecosystem and metacommunity ecology and integrate cross-scale feedbacks. Tests of this theory may combine remote sensing with a generation of networked experiments that assess effects at multiple scales. We also show how anthropogenic land cover change may alter the scaling of the BEF relationship. New research on the role of scale in BEF will guide policy linking the goals of managing biodiversity and ecosystems.© 2020 The Authors. Ecology Letters published by CNRS and John Wiley & Sons Ltd.
Babel, or the ecological stability discussions: an inventory and analysis of terminology and a guide for avoiding confusion
DOI:10.1007/s004420050090
PMID:28307528
[本文引用: 1]
We present an inventory and analysis of discussions of ecological stability, considering 163 definitions of 70 different stability concepts. Our aim is to derive a strategy that can help to dispel the existing "confusion of tongues" on the subject of "stability" and prevent its future recurrence. The strategy consists of three questions that should be kept in mind when communicating about stability properties. These three questions should overcome the three main sources of confusion in terminology. Firstly, which stability properties are being addressed in the stability statement? Our analysis shows that the general term "stability" is so ambiguous as to be useless.It can be replaced by the stability properties "staying essentially unchanged" (constancy), "returning to the reference state (or dynamic) after a temporary disturbance" (resilience), and "persistence through time of an ecological system" (persistence). Second, to what ecological situation does the statement refer? An ecological situation is defined by a set of features that, taken as a whole, determine the domain of validity of a stability statement. The six most important features form the "ecological checklist", which serves to classify ecological situations and thereby provides a system of coordinates for communication. The six points are: variable of interest, level of description, reference state, disturbance, spatial scale and temporal scale. Thirdly, is the statement anchored in the situation in question, or is there unacceptable generalisation by inferring "stability" of the whole system from a certain stability property in a certain ecological ecological situation? This question separates the scientifically valuable content of a statement from the desire for general statements which is often projected through stability statements.
Resilience, invariability, and ecological stability across levels of organization
Functional diversity increases ecological stability in a grazed grassland
DOI:10.1007/s00442-016-3802-3
PMID:28097426
[本文引用: 1]
Understanding the factors governing ecological stability in variable environments is a central focus of ecology. Functional diversity can stabilize ecosystem function over time if one group of species compensates for an environmentally driven decline in another. Although intuitively appealing, evidence for this pattern is mixed. We hypothesized that diverse functional responses to rainfall will increase the stability of vegetation cover and biomass across rainfall conditions, but that this effect depends on land-use legacies that maintain functional diversity. We experimentally manipulated grazing in a California grassland to create land-use legacies of low and moderate grazing, across which we implemented rainout shelters and irrigation to create dry and wet conditions over 3 years. We found that the stability of the vegetation cover was greatly elevated and the stability of the biomass was slightly elevated across rainfall conditions in areas with histories of moderate grazing. Initial functional diversity-both in the seed bank and aboveground-was also greater in areas that had been moderately grazed. Rainfall conditions in conjunction with this grazing legacy led to different functional diversity patterns over time. Wet conditions led to rapid declines in functional diversity and a convergence on resource-acquisitive traits. In contrast, consecutively dry conditions maintained but did not increase functional diversity over time. As a result, grazing practices and environmental conditions that decrease functional diversity may be associated with lasting effects on the response of ecosystem functions to drought. Our results demonstrate that theorized relationships between diversity and stability are applicable and important in the context of working grazed landscapes.
Eutrophication weakens stabilizing effects of diversity in natural grasslands
DOI:10.1038/nature13014 URL [本文引用: 1]
Plant ecology. Anthropogenic environmental changes affect ecosystem stability via biodiversity
DOI:10.1126/science.aaa1788
PMID:25883357
[本文引用: 1]
Human-driven environmental changes may simultaneously affect the biodiversity, productivity, and stability of Earth's ecosystems, but there is no consensus on the causal relationships linking these variables. Data from 12 multiyear experiments that manipulate important anthropogenic drivers, including plant diversity, nitrogen, carbon dioxide, fire, herbivory, and water, show that each driver influences ecosystem productivity. However, the stability of ecosystem productivity is only changed by those drivers that alter biodiversity, with a given decrease in plant species numbers leading to a quantitatively similar decrease in ecosystem stability regardless of which driver caused the biodiversity loss. These results suggest that changes in biodiversity caused by drivers of environmental change may be a major factor determining how global environmental changes affect ecosystem stability. Copyright © 2015, American Association for the Advancement of Science.
Ecosystem traits linking functional traits to macroecology
DOI:10.1016/j.tree.2018.11.004 URL [本文引用: 1]
General stabilizing effects of plant diversity on grassland productivity through population asynchrony and overyielding
Insurance effects of biodiversity can stabilize the functioning of multispecies ecosystems against environmental variability when differential species' responses lead to asynchronous population dynamics. When responses are not perfectly positively correlated, declines in some populations are compensated by increases in others, smoothing variability in ecosystem productivity. This variance reduction effect of biodiversity is analogous to the risk-spreading benefits of diverse investment portfolios in financial markets. We use data from the BIODEPTH network of grassland biodiversity experiments to perform a general test for stabilizing effects of plant diversity on the temporal variability of individual species, functional groups, and aggregate communities. We tested three potential mechanisms: reduction of temporal variability through population asynchrony; enhancement of long-term average performance through positive selection effects; and increases in the temporal mean due to overyielding. Our results support a stabilizing effect of diversity on the temporal variability of grassland aboveground annual net primary production through two mechanisms. Two-species communities with greater population asynchrony were more stable in their average production over time due to compensatory fluctuations. Overyielding also stabilized productivity by increasing levels of average biomass production relative to temporal variability. However, there was no evidence for a performance-enhancing effect on the temporal mean through positive selection effects. In combination with previous work, our results suggest that stabilizing effects of diversity on community productivity through population asynchrony and overyielding appear to be general in grassland ecosystems.
A global synthesis reveals biodiversity loss as a major driver of ecosystem change
DOI:10.1038/nature11118 URL [本文引用: 1]
Negative relationships between species richness and temporal variability are common but weak in natural systems
DOI:10.1002/ecy.2514
PMID:30198573
[本文引用: 1]
Effects of species diversity on population and community stability (or more precisely, the effects of species richness on temporal variability) have been studied for several decades, but there have been no large-scale tests in natural communities of predictions from theory. We used 91 data sets including plants, fish, small mammals, zooplankton, birds, and insects, to examine the relationship between species richness and temporal variability in populations and communities. Seventy-eight of 91 data sets showed a negative relationship between species richness and population variability; 46 of these relationships were statistically significant. Only five of the 13 positive richness-population variability relationships were statistically significant. Similarly, 51 of 91 data sets showed a negative relationship between species richness and community variability; of these, 26 were statistically significant. Seven of the 40 positive richness-community-variability relationships were statistically significant. We were able to test transferability (i.e., the predictive ability of models for sites that are spatially distinct from sites that were used to build the models) for 69 of 91 data sets; 35 and 31 data sets were transferable at the population and community levels, respectively. Only four were positive at the population level, and two at the community level. We conclude that there is compelling evidence of a negative relationship between species richness and temporal variability for about one-half of the ecological communities we examined. However, species richness explained relatively little of the variability in population or community abundances and resulted in small improvements in predictive ability.© 2018 by the Ecological Society of America.
Biodiversity and ecosystem stability
DOI:10.17520/biods.1995006 URL [本文引用: 1]
生物多样性和生态系统稳定性
Biodiversity increases the resistance of ecosystem productivity to climate extremes
DOI:10.1038/nature15374 URL [本文引用: 2]
Stability and diversity of ecosystems
DOI:10.1126/science.1133258 URL [本文引用: 2]
Stability and variability in competitive communities
Long-term variability in the abundance of populations depends on the sensitivity of species to environmental fluctuations and the amplification of environmental fluctuations by interactions among species. Although competitive interactions and species number may have diverse effects on variability measured at the individual species level, a combination of theoretical analyses shows that these factors have no effect on variability measured at the community level. Therefore, biodiversity may increase community stability by promoting diversity among species in their responses to environmental fluctuations, but increasing the number and strength of competitive interactions has little effect.
General relationships between species diversity and stability in competitive systems
DOI:10.1086/338994 URL [本文引用: 1]
No complexity-stability relationship in empirical ecosystems
DOI:10.1038/ncomms12573
PMID:27553393
[本文引用: 1]
Understanding the mechanisms responsible for stability and persistence of ecosystems is one of the greatest challenges in ecology. Robert May showed that, contrary to intuition, complex randomly built ecosystems are less likely to be stable than simpler ones. Few attempts have been tried to test May's prediction empirically, and we still ignore what is the actual complexity-stability relationship in natural ecosystems. Here we perform a stability analysis of 116 quantitative food webs sampled worldwide. We find that classic descriptors of complexity (species richness, connectance and interaction strength) are not associated with stability in empirical food webs. Further analysis reveals that a correlation between the effects of predators on prey and those of prey on predators, combined with a high frequency of weak interactions, stabilize food web dynamics relative to the random expectation. We conclude that empirical food webs have several non-random properties contributing to the absence of a complexity-stability relationship.
Predation alters relationships between biodiversity and temporal stability
DOI:10.1086/596540
PMID:19199526
[本文引用: 1]
Ecologists disagree on how diversity affects stability. At the heart of the controversy is the relationship between diversity and population stability, with conflicting findings from both theoretical and empirical studies. To help reconcile these results, we propose that this relationship may depend on trophic complexity, such that positive relations tend to emerge in multitrophic but not single-trophic communities. This hypothesis is based on the premise that stabilizing weak trophic interactions restrain population oscillations associated with strong trophic interactions in diverse multitrophic communities. We tested this hypothesis using simple freshwater bacterivorous protist communities differing in diversity with and without a predatory protist species. Coupling weak and strong trophic interactions reduced population temporal variability of the strong-interacting species, supporting the stabilizing role of weak interactions. In keeping with our hypothesis, predation altered the overall effect of diversity on population temporal stability and, in particular, caused a reversal of the diversity-stability relationship (negative without predators and positive with predators) for the strong-interacting species. A similar role of predation was also observed when examining the relationship between diversity and temporal stability of community biomass. Together, these findings demonstrated strong interactive effects of trophic interactions and diversity on temporal stability of population and community properties.
Different effects of species diversity on temporal stability in single-trophic and multitrophic communities
DOI:10.1086/605961
PMID:19775227
[本文引用: 1]
The question of how species diversity affects ecological stability has long interested ecologists and yet remains largely unresolved. Historically, attempts to answer this question have been hampered by the presence of multiple potentially confounding stability concepts, confusion over responses at different levels of ecological organization, discrepancy between theoretical predictions, and, particularly, the paucity of empirical studies. Here we used meta-analyses to synthesize results of empirical studies published primarily in the past 2 decades on the relationship between species diversity and temporal stability. We show that the overall effect of increasing diversity was positive for community-level temporal stability but neutral for population-level temporal stability. There were, however, striking differences in the diversity-stability relationship between single- and multitrophic systems, with diversity stabilizing both population and community dynamics in multitrophic but not single-trophic communities. These patterns were broadly equivalent across experimental and observational studies as well as across terrestrial and aquatic studies. We discuss possible mechanisms for population stability to increase with diversity in multitrophic systems and for diversity to influence community-level stability in general. Overall, our results indicate that diversity can affect temporal stability, but the effects may critically depend on trophic complexity.
The properties of the ecological hierarchy and their application as ecological indicators
DOI:10.1016/j.ecolind.2012.04.010 URL [本文引用: 1]
Advancing our understanding of ecological stability
DOI:10.1111/ele.v22.9 URL [本文引用: 7]
Trait-based approaches for understanding microbial biodiversity and ecosystem functioning
Species insurance trumps spatial insurance in stabilizing biomass of a marine macroalgal metacommunity
DOI:10.1002/bes2.1557 [本文引用: 1]
A conceptual analysis of ecosystem stability
生态系统稳定性定义剖析
History and status of research of ecosystem stability
生态系统稳定性研究的历史与现状
Linking biodiversity and ecosystems: towards a unifying ecological theory
DOI:10.1098/rstb.2009.0155 URL [本文引用: 1]
Biodiversity and ecosystem stability: a synthesis of underlying mechanisms
DOI:10.1111/ele.2013.16.issue-s1 URL [本文引用: 2]
Research advances on ecosystem stability
生态系统稳定性若干问题研究评述
Fluctuations of animal populations and a measure of community stability
DOI:10.2307/1929601 URL [本文引用: 1]
Functional richness, functional evenness and functional divergence: the primary components of functional diversity
DOI:10.1111/oik.2005.111.issue-1 URL [本文引用: 1]
Plant phylogenetic diversity stabilizes large-scale ecosystem productivity
DOI:10.1111/geb.12963
[本文引用: 1]
Aim Global carbon cycle models do not incorporate the stabilizing effect of biodiversity on productivity despite this phenomenon has been widely described in several local scale manipulative experiments. The reason is a lack of evidence supporting the importance of biodiversity on spatial scales at which climate models are built. Here, we test the hypothesis that diversity enhances productivity stability at a large scale. Location South American dryland known as Caatinga (similar to 830,000 km(2)). Time period 2001-2010. Major taxa studied Woody plants. Methods We used the enhanced vegetation index of Caatinga vegetation remnants, from 2001 to 2010, to calculate vegetation productivity stability across years. We used occurrence records of 606 woody species from floristic surveys to derive species richness and phylogenetic diversity at similar to 5 km and similar to 55 km (0.5 degrees) resolution. Climate data were obtained from global databases. Results Plant phylogenetic diversity has a strong positive correlation with productivity stability even after controlling for several climatic variables, such as rainfall, temperature and cloudiness, at both resolutions. Species richness was not significant when climatic variables were included. Main conclusions This result expands by several orders of magnitude the spatial scale of the evidence that biodiversity strengths the resilience of key ecosystem functions. We highlight that, by incorporating plant phylogenetic diversity, regional and global climate models can generate more accurate predictions about future ecosystem functioning and services that are critical to humankind.
Weak trophic interactions and the balance of nature
DOI:10.1038/27427 URL [本文引用: 1]
The diversity-stability debate
DOI:10.1038/35012234 URL [本文引用: 1]
Synchronized failure of global crop production
Biodiversity increases and decreases ecosystem stability
DOI:10.1038/s41586-018-0627-8 URL [本文引用: 1]
The complexity and stability of ecosystems
DOI:10.1038/307321a0 URL [本文引用: 2]
Phylogenetic diversity stabilizes community biomass
DOI:10.1093/jpe/rtt071 URL [本文引用: 1]
The dimensionality of stability depends on disturbance type
DOI:10.1111/ele.13226 URL [本文引用: 1]
Stability of ecological communities and the architecture of mutualistic and trophic networks
DOI:10.1126/science.1188321
PMID:20705861
[本文引用: 1]
Research on the relationship between the architecture of ecological networks and community stability has mainly focused on one type of interaction at a time, making difficult any comparison between different network types. We used a theoretical approach to show that the network architecture favoring stability fundamentally differs between trophic and mutualistic networks. A highly connected and nested architecture promotes community stability in mutualistic networks, whereas the stability of trophic networks is enhanced in compartmented and weakly connected architectures. These theoretical predictions are supported by a meta-analysis on the architecture of a large series of real pollination (mutualistic) and herbivory (trophic) networks. We conclude that strong variations in the stability of architectural patterns constrain ecological networks toward different architectures, depending on the type of interaction.
Understanding diversity- stability relationships: towards a unified model of portfolio effects
DOI:10.1111/ele.12019
PMID:23095077
[本文引用: 2]
A major ecosystem effect of biodiversity is to stabilise assemblages that perform particular functions. However, diversity-stability relationships (DSRs) are analysed using a variety of different population and community properties, most of which are adopted from theory that makes several restrictive assumptions that are unlikely to be reflected in nature. Here, we construct a simple synthesis and generalisation of previous theory for the DSR. We show that community stability is a product of two quantities: the synchrony of population fluctuations, and an average species-level population stability that is weighted by relative abundance. Weighted average population stability can be decomposed to consider effects of the mean-variance scaling of abundance, changes in mean abundance with diversity and differences in species' mean abundance in monoculture. Our framework makes explicit how unevenness in the abundances of species in real communities influences the DSR, which occurs both through effects on community synchrony, and effects on weighted average population variability. This theory provides a more robust framework for analysing the results of empirical studies of the DSR, and facilitates the integration of findings from real and model communities.© 2012 Blackwell Publishing Ltd/CNRS.
The ecological consequences of changes in biodiversity: a search for general principles
Biodiversity and ecosystem functioning
DOI:10.1146/ecolsys.2014.45.issue-1 URL [本文引用: 1]
Biodiversity impacts ecosystem productivity as much as resources, disturbance, or herbivory
DOI:10.1073/pnas.1208240109
PMID:22689971
[本文引用: 1]
Although the impacts of the loss of biodiversity on ecosystem functioning are well established, the importance of the loss of biodiversity relative to other human-caused drivers of environmental change remains uncertain. Results of 11 experiments show that ecologically relevant decreases in grassland plant diversity influenced productivity at least as much as ecologically relevant changes in nitrogen, water, CO(2), herbivores, drought, or fire. Moreover, biodiversity became an increasingly dominant driver of ecosystem productivity through time, whereas effects of other factors either declined (nitrogen addition) or remained unchanged (all others). In particular, a change in plant diversity from four to 16 species caused as large an increase in productivity as addition of 54 kg · ha(-1) · y(-1) of fertilizer N, and was as influential as removing a dominant herbivore, a major natural drought, water addition, and fire suppression. A change in diversity from one to 16 species caused a greater biomass increase than 95 kg · ha(-1) · y(-1) of N or any other treatment. Our conclusions are based on >7,000 productivity measurements from 11 long-term experiments (mean length, ~ 13 y) conducted at a single site with species from a single regional species pool, thus controlling for many potentially confounding factors. Our results suggest that the loss of biodiversity may have at least as great an impact on ecosystem functioning as other anthropogenic drivers of environmental change, and that use of diverse mixtures of species may be as effective in increasing productivity of some biomass crops as fertilization and may better provide ecosystem services.
Biodiversity and ecosystem stability in a decade-long grassland experiment
DOI:10.1038/nature04742 URL [本文引用: 2]
The relationship between species richness and ecosystem variability is shaped by the mechanism of coexistence
DOI:10.1111/ele.12793
PMID:28598032
[本文引用: 1]
Theory relating species richness to ecosystem variability typically ignores the potential for environmental variability to promote species coexistence. Failure to account for fluctuation-dependent coexistence may explain deviations from the expected negative diversity-ecosystem variability relationship, and limits our ability to predict the consequences of increases in environmental variability. We use a consumer-resource model to explore how coexistence via the temporal storage effect and relative nonlinearity affects ecosystem variability. We show that a positive, rather than negative, diversity-ecosystem variability relationship is possible when ecosystem function is sampled across a natural gradient in environmental variability and diversity. We also show how fluctuation-dependent coexistence can buffer ecosystem functioning against increasing environmental variability by promoting species richness and portfolio effects. Our work provides a general explanation for variation in observed diversity-ecosystem variability relationships and highlights the importance of conserving regional species pools to help buffer ecosystems against predicted increases in environmental variability.© 2017 John Wiley & Sons Ltd/CNRS.
Further thoughts on diversity and stability in ecosystems
DOI:10.17520/biods.2002015 URL [本文引用: 1]
再论生物多样性与生态系统的稳定性
DOI:10.17520/biods.2002015
[本文引用: 1]
本文在简述生物多样性与生态系统稳定性研究动态的基础上,从生物多样性和稳定性的概念出发,指出忽视多样性和稳定性的生物组织层次可能是造成观点纷争的根源之一。特定生物组织层次的稳定性可能更多地与该层次的多样性特征相关。探讨多样性和稳定性的关系应从不同的生物组织层次上进行。扰动是生态系统多样性与稳定性关系悖论中的重要因子,如果根据扰动的性质,把生态系统(或其他组织层次)区分为受非正常外力干扰和受环境因子时间异质性波动干扰2类系统,稳定性的4个内涵可以理解为:对于受非正常外力干扰的系统而言,抵抗力和恢复力是稳定性适宜的测度指标;对于受环境因子时间异质性波动干扰的系统而言,利用持久性和变异性衡量系统的稳定性则更具实际意义。结合对群落和种群层次多样性与稳定性相关机制的初步讨论,本文认为: 在特定的前提下,多样性可以导致稳定性。
Stability and synchrony across ecological hierarchies in heterogeneous metacommunities: linking theory to data
DOI:10.1111/ecog.2019.v42.i6 URL [本文引用: 2]
Ecosystem stability in space: α, β and γ variability
DOI:10.1111/ele.12292 URL [本文引用: 3]
Biodiversity and ecosystem stability across scales in metacommunities
DOI:10.1111/ele.12582 URL [本文引用: 3]
An invariability-area relationship sheds new light on the spatial scaling of ecological stability
DOI:10.1038/ ncomms15211 URL [本文引用: 2]
Asynchrony among local communities stabilises ecosystem function of metacommunities
DOI:10.1111/ele.12861
PMID:29067791
[本文引用: 1]
Temporal stability of ecosystem functioning increases the predictability and reliability of ecosystem services, and understanding the drivers of stability across spatial scales is important for land management and policy decisions. We used species-level abundance data from 62 plant communities across five continents to assess mechanisms of temporal stability across spatial scales. We assessed how asynchrony (i.e. different units responding dissimilarly through time) of species and local communities stabilised metacommunity ecosystem function. Asynchrony of species increased stability of local communities, and asynchrony among local communities enhanced metacommunity stability by a wide range of magnitudes (1-315%); this range was positively correlated with the size of the metacommunity. Additionally, asynchronous responses among local communities were linked with species' populations fluctuating asynchronously across space, perhaps stemming from physical and/or competitive differences among local communities. Accordingly, we suggest spatial heterogeneity should be a major focus for maintaining the stability of ecosystem services at larger spatial scales.© 2017 The Authors. Ecology Letters published by CNRS and John Wiley & Sons Ltd.
The complexity-stability relationship: progress in mathematical models
DOI:10.17520/biods.2019138 URL [本文引用: 1]
复杂性-稳定性研究: 数学模型的进展
DOI:10.17520/biods.2019138
[本文引用: 1]
对自然生态系统的观察给人们以复杂的群落更稳定的直观印象, 但数学模型却得出了截然相反的结论。这一“悖论”使得复杂性-稳定性研究自20世纪70年代以来成为长期的热点。本文对这一领域的数学模型研究进行简要综述。首先对这一论题进行概念剖析, 然后将各类模型分为线性和非线性两大类, 前者即群落矩阵法, 后者包括相互作用矩阵法、复杂网络数值模拟法和食物网构件动力学法。它们分别基于不同的群落构建方法和稳定性判断标准, 探求各物种是如何相互作用并实现共存的。总体而言, 在随机构建的群落模型中, 多样性和连接度的增长不利于系统稳定; 而在更接近真实自然群落的模型中, 相互作用方式、网络拓扑结构、相互作用强度分布等方面的机制提供了稳定效应, 按此组织的生态网络可达到很高的复杂度。然而, 复杂性-稳定性的研究还远未结束, 当前的模型仍不足以反映自然群落中的复杂相互作用, 稳定性的概念也有待拓展。对这一议题的深入研究在生态学理论和生态系统管理实践方面都具有重大价值。
Biodiversity and ecosystem productivity in a fluctuating environment: the insurance hypothesis
Although the effect of biodiversity on ecosystem functioning has become a major focus in ecology, its significance in a fluctuating environment is still poorly understood. According to the insurance hypothesis, biodiversity insures ecosystems against declines in their functioning because many species provide greater guarantees that some will maintain functioning even if others fail. Here we examine this hypothesis theoretically. We develop a general stochastic dynamic model to assess the effects of species richness on the expected temporal mean and variance of ecosystem processes such as productivity, based on individual species' productivity responses to environmental fluctuations. Our model shows two major insurance effects of species richness on ecosystem productivity: (i) a buffering effect, i.e., a reduction in the temporal variance of productivity, and (ii) a performance-enhancing effect, i.e., an increase in the temporal mean of productivity. The strength of these insurance effects is determined by three factors: (i) the way ecosystem productivity is determined by individual species responses to environmental fluctuations, (ii) the degree of asynchronicity of these responses, and (iii) the detailed form of these responses. In particular, the greater the variance of the species responses, the lower the species richness at which the temporal mean of the ecosystem process saturates and the ecosystem becomes redundant. These results provide a strong theoretical foundation for the insurance hypothesis, which proves to be a fundamental principle for understanding the long-term effects of biodiversity on ecosystem processes.
Predicting the stability of multitrophic communities in a variable world
DOI:10.1002/ecy.2992 [本文引用: 1]
Linking stoichiometric homoeostasis with ecosystem structure, functioning and stability
DOI:10.1111/j.1461-0248.2010.01532.x URL [本文引用: 1]
Temporal stability of aboveground biomass is governed by species asynchrony in temperate forests
DOI:10.1016/j.ecolind.2019.105661 URL [本文引用: 1]
Biodiversity and stability mechanisms: understanding and future research
生物多样性与稳定性机制研究进展
Trait diversity promotes stability of community dynamics
DOI:10.1007/s12080-012-0160-6 URL [本文引用: 1]
Nitrogen addition does not reduce the role of spatial asynchrony in stabilising grassland communities
DOI:10.1111/ele.2019.22.issue-4 URL [本文引用: 1]
Diversity components and assembly patterns of plant functional traits determine community spatial stability under resource gradients in a desert steppe
DOI:10.1071/RJ15094 URL [本文引用: 1]
Horizontal and vertical diversity jointly shape food web stability against small and large perturbations
DOI:10.1111/ele.2019.22.issue-7 URL [本文引用: 2]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}