


土壤水分有效性对土壤碳矿化速率及微生物活性至关重要。大量研究表明, 土壤CO2排放量对土壤含水量的变化存在阈值响应, 即土壤CO2排放量随着土壤含水量的增加呈先升高后降低的变化趋势(Schjønning et al., 2003)。例如, Das等(2019)发现美国东部滨海地区土壤含水量为60%时需氧微生物活性和CO2排放量达到最大, 低于40%或高于70%均会产生显著抑制作用。然而, 土壤含水量对土壤CH4排放量的影响与CO2排放量不同; 随着土壤含水量的升高, 土壤O2含量减少, 需氧微生物活性降低, 但这却有利于厌氧微生物活性的增强, 进而提高土壤CH4排放(Vizza et al., 2017)。土壤含盐量高是滨海湿地的重要环境特征, 也是影响生态系统碳循环过程的重要因素(Xi et al., 2014)。例如, 贺强等(2009)分析发现黄河三角洲滨海湿地土壤0-20 cm盐分变化范围为1.25-17.77 g·kg-1, 属于重盐土类型。土壤高盐分首先会提高土壤溶液渗透压, 并对微生物产生离子毒害作用, 显著降低微生物活性甚至改变其群落结构; 其次会造成土壤有机碳的溶解、迁移速率以及可被微生物利用性降低, 这两个方面都将显著影响土壤碳矿化能力(Rath et al., 2016)。在滨海湿地生态系统中, 目前研究主要关注水分或盐分单一因子对土壤碳矿化的影响, 而水盐协同作用的影响以及机理尚不明确(Jones et al., 2017)。此外, 过去针对土壤碳矿化的研究主要聚焦于土壤CO2的排放, 由于CH4排放量相对较低, 其在土壤碳矿化的研究中常常被忽略。然而, CH4增温潜势是CO2的28倍(IPCC, 2013), 湿地作为大气中CH4最大的天然来源, CH4排放特征应需要加强研究(Tollefson, 2022)。
黄河三角洲滨海湿地是我国重要的新生湿地类型, 其成土年龄短, 土壤有机碳储量低并且主要聚集于土壤表层, 因此具有非常大的碳汇功能提升潜力(于君宝等, 2013; Bai et al., 2016)。同时, 黄河入海泥沙淤积在三角洲使得大面积的盐沼湿地不受潮汐作用影响, 而主要受地下水(咸水)和降雨(淡水)交互作用的影响(Han et al., 2018)。因此, 研究土壤水盐变化对土壤碳矿化的影响对于理解该盐沼湿地的碳汇功能至关重要。本研究选取黄河三角洲典型盐沼湿地为研究对象, 通过水盐梯度模拟实验, 对土壤理化性质、微生物群落结构及碳矿化速率进行分析, 探讨: 1)土壤水盐变化对土壤理化性质及微生物的影响特征; 2)土壤水盐变化对土壤碳矿化的影响特征及其机制。本研究假设土壤水盐变化通过改变土壤环境及微生物结构等因素对土壤CO2和CH4排放产生不同的影响。同时, 期望能更深入了解滨海盐沼湿地碳矿化过程及其调控机制, 并可为模拟和预测滨海盐沼湿地“碳汇”功能提供重要科学数据。
1 材料和方法
1.1 研究区概况
本研究区处于中国科学院黄河三角洲滨海湿地生态系统野外科学观测研究站(37.76° N, 118.99° E), 地处山东省东营市垦利区境内。该区属于暖温带大陆性季风气候, 夏季盛行东南风, 气候炎热多雨; 冬季盛行西北风, 气候寒冷干燥。该研究区年平均气温12.9 ℃, 最低月(1月)平均气温-2.8 ℃, 最高月(7月)平均气温26.7 ℃; 年降水量556.1 mm, 其中约70%的降水集中于生长季; 平均年日照时间约2 750.9 h, 年蒸发量1 962 mm, 蒸降比约为3.2:1。该地区地势平坦, 土壤类型主要以潮土和滨海盐土为主, 土壤质地以壤土为主。该区植被的分布主要受土壤水分和盐分两个环境因子的共同影响(贺强等, 2009), 主要的优势种为芦苇(Phragmites australis)、盐地碱蓬(Suaeda salsa)、白茅(Imperata cylindrica)、荻(Miscanthus sacchariflora)和柽柳(Tamarix chinensis)。
根据降水量和土壤含水量波动情况, 可将黄河三角洲典型非潮汐芦苇湿地生长季分为4个阶段, 即干旱阶段、前湿润阶段、淹水阶段和后湿润阶段, 即形成了以“春季干旱盐分浓度高、夏秋多雨水涝时间长”为典型区域特征的水盐时空分布格局。在非潮汐芦苇湿地的研究表明, 土壤饱和含水量变化范围可以从干旱期的最低值26.4%到淹水期的最大值100% (李新鸽等, 2019)。同时, 土壤水分是盐分迁移运动的载体, “盐随水走”原理使土壤水分在蒸发过程中将深层土壤中的溶解性盐分及地下水通过土壤毛细管上升作用聚集于表层, 造成了土壤盐分的“表聚”现象, 土壤0-20 cm盐分变化范围为1.25- 17.77 g·kg-1 (贺强等, 2009)。
1.2 实验设计
本实验选择远离海岸且不受潮汐影响的芦苇盐沼湿地为研究对象。2019年6月中旬采用不锈钢铲采取芦苇群落0-10 cm的表层土壤, 然后进行风干处理并剔除植物碎屑、根及杂质, 最后通过2 mm不锈钢筛, 使土壤均质化。研究样地表层土壤理化性质见表1。
表1 黄河三角洲滨海盐沼湿地0-10 cm表层土壤基本特征(平均值±标准误)
Table 1
| TC (g·kg-1) | TN (g·kg-1) | NH4+ -N (mg·kg-1) | NO3- -N (mg·kg-1) | pH | Soil salinity (%) |
|---|---|---|---|---|---|
| 12.76 ± 0.07 | 0.21 ± 0.01 | 6.56 ± 0.06 | 1.49 ± 0.03 | 7.64 ± 0.04 | 0.90 ± 0.04 |
NH4+-N, 铵态氮含量; NO3--N, 硝态氮含量; Soil salinity, 土壤盐分含量; TC, 土壤总碳含量; TN, 土壤总氮含量.
NH4+-N, ammonium nitrogen content; NO3--N, nitrate nitrogen content; TC, soil total carbon content; TN, soil total nitrogen content.
实验采用土壤水分和盐分双因子设计, 根据我们测定的该芦苇湿地土壤含水量和土壤盐分的动态变化范围, 设置土壤水分含量为4个水平: 25%、50%、75%和100%土壤饱和含水量, 分别用W1、W2、W3和W4表示; 土壤盐分含量为2个水平: 低盐(9 g·kg-1)和高盐(18 g·kg-1), 分别用S1和S2表示。实验中4个水分水平和2个盐分水平两两交叉, 共有8个处理, 每个处理4组重复, 其中S1W3处理接近6月中旬的土壤原生生境。
实验开始时, 为更接近野外土壤环境, 将100 g风干土放入1 000 mL棕色遮光的广口瓶中培养, 并根据黄河三角洲滨海盐沼湿地盐土的主要组成成分, 采用NaCl溶液进行土壤盐浓度的调节, 采用去离子水对土壤水分进行调节。当所有处理中的土壤水盐含量达到所要求的水平时, 开始进行实验。实验中通过瓶子质量的记录, 每天对土壤样品表面均匀添加去离子水以保持土壤水分的恒定, 整个实验过程室温保持在25 ℃下。实验结束后利用无菌铲进行土壤样品的采集, 每个土壤样品分为3份, 一份自然风干后用于测定土壤pH、总碳、总氮、铵态氮和硝态氮含量等指标; 一份鲜样装到无菌的密封袋(在-80 ℃的条件下储存)用于土壤微生物群落结构多样性分析; 一份鲜样装到无菌的密封袋(在4 ℃的条件下储存)用于土壤微生物生物量碳及溶解性有机碳含量的测定。
1.3 土壤碳矿化速率测定
本实验在培养后第1、3、5、7、9、15、21和40天后收集和测量气体样本。广口瓶用装有三通阀且气密性好的盖子密封, 用连有三通阀的注射器进行气体样本的收集, 并注射到预先抽成真空的20 mL气体采样瓶中。随后土壤培养24 h, 收集第二份气体样本。为降低瓶内气体浓度升高造成的处理间压力差异, 在采集完培养24 h的样品后, 就打开瓶盖进行通风, 一直到下一个取样日期。气体样品在24 h内使用配备FID和ECD的气相色谱仪(Agilent 7890A, Agilent, Palo Alto, USA)分析CO2和CH4气体浓度。
式中, ρ是CO2或CH4在标准温度和压力下的密度(1.977和0.716 g·L-1), V是广口瓶的体积(L), m是土壤的质量(g), dc/dt是气体浓度梯度随时间变化的线性回归斜率, T是培养温度(K)。通过使用梯形法则, 将CO2或CH4累积排放量计算为速率所限定的面积之和。
1.4 土壤理化性质测定
采用环刀法测定了表层土壤饱和含水量, 采用水土质量比5:1的传统方法测定土壤含盐量和土壤pH; 采用元素分析仪(vario MACRO cube, Elementar, Hanau, Germany)测定培养前后土壤样品的总碳(TC)和总氮(TN)含量; 采用0.5 mol·L-1 K2SO4浸提法提取土壤可溶性有机碳(DOC), 通过总有机碳分析仪(TOC-VCPH, Shimadzu, Kyoto, Japan)测定其含量; 采用氯仿熏蒸-0.5 mol·L-1 K2SO4浸提法提取土壤微生物生物量碳(MBC), 并使用总有机碳分析仪测定其含量(Joergensen, 1996); 采用连续流动分析仪(Auto Analyzer III, Seal, Norderstedt, Germany)测定土壤铵态氮和硝态氮含量。
1.5 全球增温潜势(GWP)计算
GWP是某一给定物质在一定时间范围内与CO2相比得到的相对辐射影响值, 是评估不同温室气体之间温室效应的重要衡量标准。CH4的全球增温潜势在100年尺度上是CO2的28倍(IPCC, 2013)。土壤CO2和CH4排放的总GWP计算公式如下:
1.6 土壤微生物多样性分析
土壤样品利用MoBio PowerSoil DNA Isolation Kit (MoBio Laboratories, San Diego, USA)试剂盒提取土壤微生物DNA, 采用Nanodrop紫外分光光度计(Thermo Scientific, Wilmington, USA)检测DNA浓度和纯度, 利用1%琼脂糖凝胶电泳检测蛋白污染及DNA的完整度。后续以提取的土壤DNA为模板进行PCR扩增实验, 使用细菌引物组515F (5′-GTGCCAGCMGCCGCGGTAA-3′)和806R (5′-GGACTACHVGGGTWTCTAAT-3′)扩增16S rRNA基因的V3-V4可变区。测序工作委托广东美格基因科技有限公司完成。
1.7 数据处理
采用双因素方差分析法分析土壤盐分、水分以及它们潜在的交互作用对各土壤指标的影响。采用单因素方差分析和最小显著差异(LSD)法比较不同水盐处理下土壤环境因子及土壤CO2和CH4排放量差异(α = 0.05)。在进行方差分析之前, 对原始数据进行正态性检验和等方差检验。采用Pearson相关分析方法, 确定土壤理化性质、微生物多样性与土壤碳矿化速率的相关性。所有数据分析均基于统计分析软件SPSS 26.0完成, 相关的图形均基于SigmaPlot 14.0与R 3.6.3软件完成, 图表中数据为平均值±标准误。
2 结果
2.1 土壤水盐变化对土壤理化性质和微生物生物量的影响
统计分析表明, 土壤水分和盐分对TC和TN含量均没有交互作用(p > 0.05) (表2)。土壤水分含量的增加显著提高了TC含量, 但盐分对其没有显著影响(图1A)。土壤水分对TN含量影响不显著, 但除W4处理外, 盐分增加显著提高了TN含量(图1B)。水盐变化对土壤DOC含量具有弱交互作用(p = 0.06); 在低水分处理下, DOC随着土壤盐分的增加呈减少趋势, 但在高水分处理下呈增加趋势(图1C)。土壤水分和盐分处理对MBC含量存在显著交互作用(p < 0.05), 在低盐处理下MBC含量随着水分含量的增加呈先增加后降低的单峰型变化趋势, 在W3处理中达到峰值, 在高盐处理下, MBC随水分变化出现与低盐处理相同的变化趋势; 同时统计分析表明, 除W4处理外, 高盐处理的MBC显著低于低盐处理(图1D)。
表2 土壤水分和盐分对土壤理化性质和碳矿化速率影响的方差分析
Table 2
| 因子 Factor | TC | TN | DOC | MBC | Cumulative CO2 | Cumulative CH4 | GWP | CH4:CO2 | |
|---|---|---|---|---|---|---|---|---|---|
| 土壤水分 Soil moisture | p | <0.01 | 0.65 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | 0.09 |
| F | 33.78 | 0.56 | 21.08 | 8.33 | 72.17 | 6.38 | 14.21 | 2.42 | |
| 土壤盐分 Soil salinity | p | 0.22 | <0.01 | 0.15 | 0.02 | <0.01 | <0.01 | <0.01 | 0.01 |
| F | 1.59 | 9.06 | 2.22 | 6.07 | 47.68 | 14.73 | 22.99 | 7.71 | |
| 土壤水分×盐分 Soil moisture × salinity | p | 0.79 | 0.92 | 0.06 | 0.04 | 0.29 | 0.96 | 0.95 | 0.88 |
| F | 0.35 | 0.17 | 2.81 | 3.33 | 1.33 | 0.10 | 0.12 | 0.23 |
CH4:CO2, 土壤CH4与CO2通量比率; Cumulative CH4, 土壤CH4累积排放量; Cumulative CO2, 土壤CO2累积排放量; DOC, 土壤可溶性有机碳含量; GWP, 全球增温潜势; MBC, 土壤微生物生物量碳含量; TC, 土壤总碳含量; TN, 土壤总氮含量。
CH4:CO2, soil CH4 to CO2 ratio; Cumulative CH4, soil cumulative CH4 emissions; Cumulative CO2, soil cumulative CO2 emissions; DOC, soil dissolved organic carbon content; GWP, global warming potential; MBC, soil microbial biomass carbon content; TC, soil total carbon content; TN, soil total nitrogen content.
图1
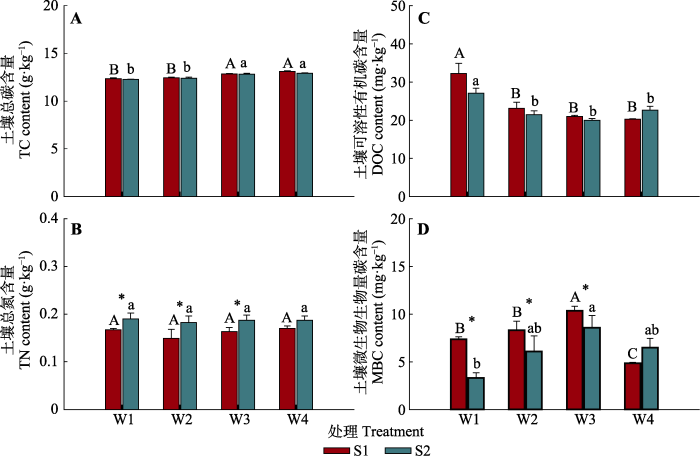
图1
不同水盐处理下土壤总碳(TC) (A)、总氮(TN) (B)、可溶性有机碳(DOC) (C)和微生物生物量碳(MBC) (D)含量的均值变化(标准值±标准误)。不同大写字母表示在低盐分(S1)下不同水分处理间的差异显著(p < 0.05), 不同小写字母表示在高盐分(S2)下不同水分处理间的差异显著(p < 0.05), *表示相同水分下不同盐分处理间的差异显著(p < 0.05)。W1, 25%土壤饱和含水量; W2, 50%土壤饱和含水量; W3, 75%土壤饱和含水量; W4, 100%土壤饱和含水量。
Fig. 1
Effects of soil moisture and salinity on mean values of soil total carbon (TC) content (A), soil total nitrogen (TN) content (B), soil dissolved organic carbon (DOC) content (C) and soil microbial biomass carbon (MBC) content (D) (mean ± SE). Different uppercase letters indicate significant difference between soil moisture treatments under low salinity level (S1) (p < 0.05), different lowercase letters indicate significant difference between soil moisture treatments under high salinity level (S2) (p < 0.05), and * indicates the significant difference between salinity treatments under the same moisture condition (p < 0.05). W1, 25% soil saturated moisture content; W2, 50% soil saturated moisture content; W3, 75% soil saturated moisture content; W4, 100% soil saturated moisture content.
2.2 土壤水盐变化对土壤微生物群落结构的影响
本实验采用物种数量(observed species指数)和Chao1指数反映细菌群落丰富度, 数值越大, 细菌物种越丰富; 采用Shannon-Wiener多样性指数和PDwhole tree指数反映细菌群落的多样性, 数值越大,细菌群落多样性越高。土壤α多样性分析表明, 土壤水分含量的增加显著增加了物种数量、Chao1指数、Shannon-Wiener多样性指数以及PD whole tree指数, 说明土壤水分含量显著提高了细菌群落物种丰富度及群落多样性。同时分析结果表明, 低盐处理中土壤α多样性指数(物种数量、Chao1指数、Shannon-Wiener多样性指数以及PD whole tree指数)均高于高盐处理, 这表明土壤盐分升高对土壤细菌群落物种丰富度及群落多样性具有显著的抑制作用(表3, 表4)。
表3 土壤水分和盐分含量对土壤细菌α多样性指数的双因素方差分析结果
Table 3
| 因子 Factor | 物种数量 Observed species | PD whole tree指数 PD whole tree index | Shannon-Wiener多样性指数 Shannon-Wiener diversity index | Chao1指数 Chao1 index | |
|---|---|---|---|---|---|
| 土壤水分 Soil moisture | p | <0.01 | <0.01 | <0.01 | <0.01 |
| F | 31.96 | 29.35 | 40.49 | 6.46 | |
| 土壤盐分 Soil salinity | p | <0.01 | <0.01 | <0.01 | 0.30 |
| F | 29.51 | 22.25 | 39.07 | 1.11 | |
土壤水分  ×盐分 ×盐分Soil moisture  × salinity × salinity | p | <0.01 | <0.01 | 0.01 | 0.82 |
| F | 9.02 | 6.93 | 5.25 | 0.31 |
表4 不同水盐处理下土壤细菌α多样性指数(平均值±标准误)
Table 4
| 盐分处理 Salinity treatment | 水分处理 Moisture treatment | 物种数量 Observed species | PD whole tree指数 PD whole tree index | Shannon-Wiener多样性指数 Shannon-Wiener diversity index | Chao1指数 Chao1 index |
|---|---|---|---|---|---|
| S1 | W1 | 2 603.26 ± 41.57Ba | 213.04 ± 10.30Ba | 6.59 ± 0.41Ba | 3 845.65 ± 165.76Ba |
| W2 | 3 068.08 ± 36.67Aa | 252.76 ± 3.12Aa | 8.29 ± 0.12Ba | 4 383.72 ± 70.05Aa | |
| W3 | 3 227.84 ± 34.21Aa | 265.12 ± 3.10Aa | 8.81 ± 0.05Aa | 4 577.47 ± 47.85Aa | |
| W4 | 3 241.94 ± 56.62Aa | 268.94 ± 5.06Aa | 8.44 ± 0.13Aa | 4 558.03 ± 63.28Aa | |
| S2 | W1 | 2 629.74 ± 18.92Ba | 223.32 ± 2.96Ba | 6.46 ± 0.08Ca | 3 881.19 ± 89.97Ba |
| W2 | 2 541.14 ± 64.56Bb | 219.07 ± 4.55Bb | 6.95 ± 0.19Bb | 3 836.97 ± 84.56ABb | |
| W3 | 2 903.29 ± 39.22Ab | 241.88 ± 2.68Ab | 7.72 ± 0.10Ab | 4 290.19 ± 56.94Ab | |
| W4 | 3 002.14 ± 29.46Ab | 251.94 ± 2.18Ab | 8.00 ± 0.02Ab | 4 395.78 ± 60.89Aa |
不同大写字母表示相同盐分下不同水分处理之间的差异显著(p < 0.05), 不同小写字母表示相同水分下不同盐分处理之间的差异显著(p < 0.05)。S1, 9 g·kg-1的土壤盐分; S2, 18 g·kg-1的土壤盐分; W1, 25%土壤饱和含水量; W2, 50%土壤饱和含水量; W3, 75%土壤饱和含水量; W4, 100%土壤饱和含水量。
Different uppercase letters indicate significant difference among different moisture treatments at the same salinity level (p < 0.05), and different lowercase letters indicate significant difference between salinity treatments at the same moisture condition (p < 0.05). S1, 9 g·kg-1 soil salinity; S2, 18 g·kg-1 soil salinity; W1, 25% soil saturated moisture content; W2, 50% soil saturated moisture content; W3, 75% soil saturated moisture content; W4, 100% soil saturated moisture content.
图2
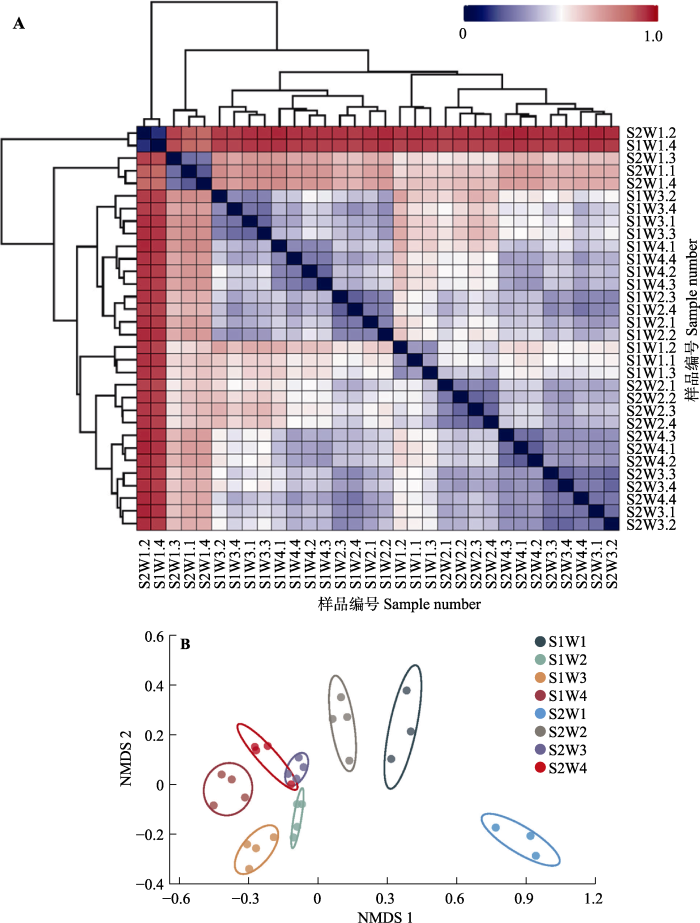
图2
土壤细菌群落样本距离热图(A)及非度量多维尺度分析(NMDS)分析图(B)。A中大写字母和数字代表土壤样品编号, B中大写字母和数字代表实验处理编号。S1, 9 g·kg-1的土壤盐分; S2, 18 g·kg-1的土壤盐分; W1, 25%的土壤饱和含水量; W2, 50%的土壤饱和含水量; W3, 75%的土壤饱和含水量; W4, 100%的土壤饱和含水量。
Fig. 2
Heat map of soil bacterial community sample distance (A) and non-metric multidimensional scaling (NMDS) analysis (B). In A, uppercase letters and numbers represent soil sample numbers; in B, uppercase letters and numbers represent the experimental treatment number. S1, 9 g·kg-1 soil salinity; S2, 18 g·kg-1 soil salinity; W1, 25% soil saturation moisture content; W2, 50% soil saturation moisture content; W3, 75% soil saturation moisture content; W4, 100% soil saturation moisture content.
2.3 土壤CO2和CH4排放对土壤水盐变化的响应特征
在不同土壤水盐条件下, 土壤CO2排放速率均具有显著的时间动态变化特征。在培养初期, 不同处理下的CO2排放速率迅速上升, 随着培养时间的推移在第3-5天达到最大值, 随后迅速下降并于培养后期逐渐趋于平缓(图3A)。CO2累积排放量随时间变化呈现初期迅速增加, 后期趋于缓慢上升的趋势(图3C)。CH4排放速率在培养初期迅速上升, 在第3天达到最大值后出现下降趋势, 但在培养后期随时间推移表现出略增的平稳趋势(图3B); CH4累积排放量随时间推移总体表现为稳步增加的趋势(图3D)。统计分析表明, 土壤水分和盐分对土壤CO2和CH4累积排放量、GWP、CH4:CO2的影响均不存在交互作用(p > 0.05) (表2)。土壤水分对土壤CO2和CH4累积排放量、GWP具有显著影响, 对CH4:CO2没有影响; 而土壤盐分对4个指标都具有显著影响(p < 0.05)。
图3
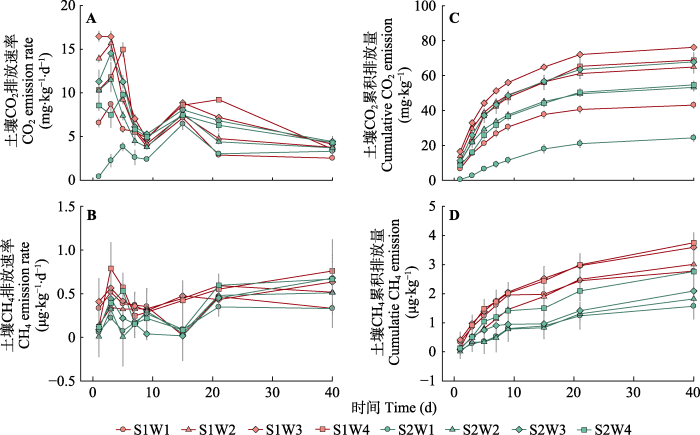
图3
不同水盐处理下土壤CO2排放速率(A)、土壤CH4排放速率(B)、土壤CO2累积排放量(C)和土壤CH4累积排放量(D)的动态变化(平均值±标准误)。S1, 9 g·kg-1的土壤盐分; S2, 18 g·kg-1的土壤盐分; W1, 25%的土壤饱和含水量; W2, 50%的土壤饱和含水量; W3, 75%的土壤饱和含水量; W4, 100%的土壤饱和含水量。
Fig. 3
Dynamic changes of soil CO2 emission rate (A), soil CH4 emission rate (B), soil cumulative CO2 emission rate (C) and soil cumulative CH4 emission rate (D) under different soil moisture and salinity treatments (mean ± SE). S1, 9 g·kg-1 soil salinity; S2, 18 g·kg-1 soil salinity; W1, 25% soil saturation moisture content; W2, 50% soil saturation moisture content; W3, 75% soil saturation moisture content; W4, 100% soil saturation moisture content.
在两个盐度水平下, 土壤CO2累积排放量均随水分含量的增加呈先增加后降低的趋势, 在W1处理中排放量最小, 而在W3处理中排放量最大; 同时本研究发现在不同水分条件下, 盐分升高则显著抑制了土壤CO2累积排放量(图4A), 在低盐(S1)处理中, 不同水分处理下的土壤CO2累积排放量最小值为215.46 mg·kg-1, 最大值为380.84 mg·kg-1; 而在高盐(S2)处理中土壤CO2累积排放量最小值为121.55 mg·kg-1, 最大值为339.27 mg·kg-1。水分含量的增加显著增加了土壤CH4累积排放量; 除W1处理外, 盐分增加则对CH4累积排放量具有显著的抑制作用(图4B)。同时, 和CO2累积排放量相一致, GWP随水分含量的增加呈先增加后降低的趋势, 在W3处理下达到最大值; 盐分对GWP具有显著抑制作用(图4C)。CH4:CO2在不同盐分下对水分含量变化的响应不一致, 在低盐中各处理之间差异不显著; 在高盐处理中则表现为随水分增加而显著增加的变化趋势。同时盐分升高显著降低了W3处理中的CH4:CO2 (图4D)。
图4
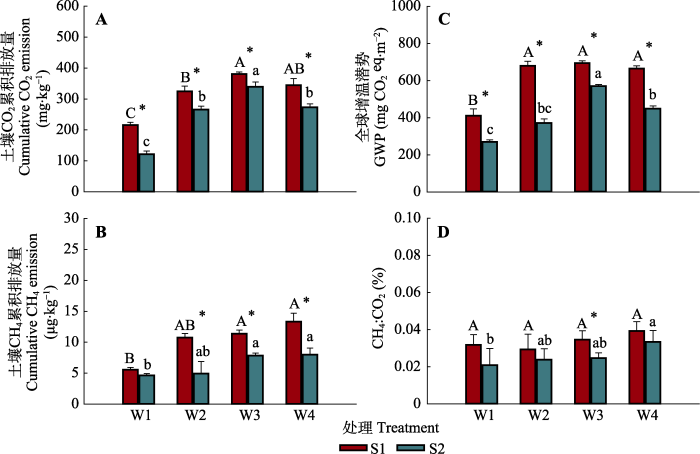
图4
不同水盐处理下土壤CO2累积排放量(A)、CH4累积排放量(B)、全球增温潜势(GWP) (C)及CH4:CO2通量比率(D)的变化(平均值±标准误)。不同大写字母表示在低盐分(S1)下水分处理间的差异显著(p < 0.05), 不同小写字母表示在高盐分(S2)下水分处理间的差异显著(p < 0.05), *表示相同水分下盐分处理间的差异显著(p < 0.05)。S1, 9 g·kg-1的土壤盐分; S2, 18 g·kg-1的土壤盐分; W1, 25%的土壤饱和含水量; W2, 50%的土壤饱和含水量; W3, 75%的土壤饱和含水量; W4, 100%的土壤饱和含水量。eq, 当量。
Fig. 4
Effects of soil moisture and salinity on values of soil cumulative CO2 emission (A) and soil cumulative CH4 emission (B), global warming potential (GWP) (C) and CH4:CO2 (D) (mean ± SE). Different uppercase letters indicate significant difference between soil moisture treatments under low salinity level (S1) (p < 0.05), different lowercase letters indicate significant difference between soil moisture treatments under high salinity level (S2) (p < 0.05), and * indicates significant difference between salinity treatments under the same moisture condition (p < 0.05). S1, 9 g·kg-1 soil salinity; S2, 18 g·kg-1 soil salinity; W1, 25% soil saturation moisture content; W2, 50% soil saturation moisture content; W3, 75% soil saturation moisture content; W4, 100% soil saturation moisture content. eq, equivalent.
2.4 水盐变化背景下影响土壤CO2和CH4排放的因子及相关关系
Pearson相关分析表明, 土壤CO2排放量与CH4排放量、TC含量、MBC含量、MBN含量、GWP及CH4:CO2均存在显著正相关关系, 与TN和DOC含量呈显著负相关关系; 土壤CH4排放与TC含量、MBC含量、GWP及CH4:CO2呈显著正相关关系, 与TN含量呈显著负相关关系; MBC含量与GWP呈显著正相关关系; GWP与CH4:CO2存在显著正相关关系; CO2和CH4排放量均与土壤细菌α多样性指数(物种数量、Chao1指数、Shannon-Wiener多样性指数以及PD whole tree指数)均呈显著正相关关系(图5)。
图5
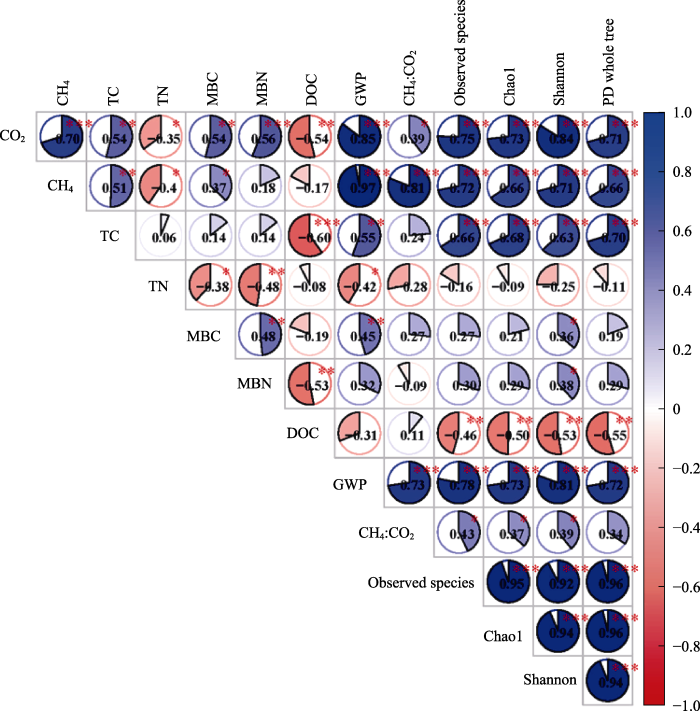
图5
土壤碳矿化速率与土壤理化性质、微生物特征之间的Pearson相关关系。*, p < 0.05; **, p < 0.01; ***, p < 0.001。CH4, 土壤CH4排放量; CH4:CO2, 土壤CH4与CO2通量比率; Chao1, Chao1指数; CO2, 土壤CO2排放量; DOC, 土壤可溶性有机碳含量; GWP, 全球增温潜势; MBC, 土壤微生物生物量碳含量; MBN, 土壤微生物生物量氮含量; Observed species, 物种数量; PD whole tree, PD whole tree指数; Shannon, Shannon-Wiener多样性指数; TC, 土壤总碳含量; TN, 土壤总氮含量。
Fig. 5
Pearsonʼs correlation between soil carbon mineralization rate and physical and chemical properties as well as microbial characteristics. *, p < 0.05; **, p < 0.01; ***, p < 0.001. CH4, soil CH4 emission; CH4:CO2, soil CH4 to CO2 ratio; Chao1, Chao1 index; CO2, soil CO2 emission; DOC, soil dissolved organic carbon content; GWP, global warming potential; MBC, soil microbial biomass carbon content; MBN, soil microbial biomass nitrogen content; PD whole tree, PD whole tree index; Shannon, Shannon-Wiener diversity index; TC, soil total carbon content; TN, soil total nitrogen content.
3 讨论
持续的黄河泥沙入海及淤积造陆使得大面积的黄河三角洲滨海盐沼湿地不受潮汐作用影响, 而主要受地下水(咸水)和降雨(淡水)交互作用的影响(Han et al., 2018)。土壤水分和盐分是影响该区域盐沼湿地植被群落结构和生态系统功能的重要因子(贺强等, 2009)。土壤水盐条件的变化通过改变植被群落组成、生产力和土壤微生物特征, 显著影响该生态系统的碳积累和碳循环过程。土壤碳矿化向大气释放CO2和CH4的过程是滨海湿地碳输出的重要过程, 研究土壤水盐变化对其影响特征及背后的机制对于准确理解滨海湿地碳积累有着重要的意义。通过水盐梯度模拟实验, 本研究发现土壤水分和盐分对黄河三角洲芦苇盐沼湿地土壤碳矿化速率没有交互作用, 这说明土壤水分和盐分通过各自的调控机制影响着该盐沼湿地土壤碳矿化过程。
3.1 土壤水分对土壤碳矿化的影响
土壤水分是调控土壤碳矿化的重要因素。大量研究已经表明, 土壤碳矿化速率随土壤水分增加表现出先增加后降低的单峰型变化趋势(Ilstedt et al., 2000)。例如, Das等(2019)在美国东部滨海地区研究发现土壤含水量为60%时需氧微生物活性和CO2排放量达到最大, 土壤含水量低于40%或高于70%均会产生显著抑制作用; Lewis等(2014)在美国佛罗里达中西部亚热带滨海盐沼湿地的研究发现, 土壤水分从75%增加到85%时, 土壤CO2排放量显著被抑制, 降低了65%。在本研究中, 土壤CO2累积排放量随土壤水分增加表现出先增加后降低的单峰型变化趋势, 其中75%土壤饱和含水量处理下的CO2累积排放量最高, 25%土壤饱和含水量处理下最低。在低水分条件下, 土壤水分增加刺激土壤微生物活性(包括胞外酶活性), 微生物数量增加, 加速DOC的分解, 从而促进CO2排放; 但土壤含水量过高则会减缓土壤中O2的扩散, 进而抑制有氧微生物活性和土壤CO2排放速率(Davidson & Janssens, 2006)。土壤水分不但影响微生物生长和繁殖, 也是影响土壤微生物群落结构的主要因素(Brockett et al., 2012)。Dinter等(2019)在德国瓦登海盐沼湿地研究发现土壤水分条件通过影响土壤微生物群落结构进而调控土壤有机碳矿化过程。本研究发现土壤微生物量(用微生物生物量碳含量代表)随着土壤水分的增加也呈现出先增加后降低的单峰型变化趋势; 同时发现土壤水分的增加显著提高了土壤细菌α多样性指数。这与Li等(2022)在崇明岛东滩滨海湿地中研究发现土壤水分与土壤微生物多样性指数呈显著正相关关系结果相一致, 表明土壤水分是影响滨海湿地土壤微生物群落组成和结构的重要因素。聚类分析结果显示, 不同水分处理间的土壤细菌聚类远, 说明土壤水分升高导致了土壤微生物群落差异增大。相关性分析表明土壤CO2排放量与土壤微生物生物量、细菌群落丰富度及多样性指数具有显著正相关关系。因此, 通过调控微生物生物量和群落结构等指标, 土壤水分变化显著影响微生物主导的土壤CO2排放特征。
作为滨海盐沼湿地土壤碳矿化排放的重要过程, CH4排放是土壤中CH4产生、消耗和传输等诸多过程的综合结果(王洁等, 2016)。前人研究已经证明土壤水分和土壤CH4通量之间存在显著正相关关系, 即土壤水分越高, 越有利于厌氧微生物的活性以及CH4排放(Vizza et al., 2017)。Christiansen等(2016)在加拿大温哥华岛滨海地区研究发现, 土壤水分含量升高显著抑制CH4氧化过程, 进而促进CH4排放, 并认为CH4产生过程是控制CH4通量的主要因素。同时, 土壤水分对CH4产生和消耗过程中产甲烷菌和甲烷氧化菌的数量及活性也具有重要的影响(Wagner, 2017)。例如, Chi等(2021)在黄河三角洲滨海湿地研究发现, 土壤水分增加显著提高了古菌多样性指数并改变了群落结构。土壤水分含量的提高会降低O2含量, 进而为激发产甲烷菌活性提供了有利的厌氧环境, 产甲烷菌丰富度和产甲烷菌群落比率逐渐升高, 从而促进CH4的产生及排放(Le Mer & Roger, 2001)。本研究聚类分析表明不同土壤水分处理间土壤细菌群落差异较大; 土壤水分含量增加显著提高了土壤CH4排放量、土壤微生物群落丰富度及多样性指数; 同时, 相关性分析也表明土壤CH4排放量与土壤微生物群落丰富度和多样性指数呈显著正相关关系, 这说明土壤水分条件变化通过改变土壤厌氧微生物群落结构及其介导的分解过程, 进而促进CH4的产生及排放。
此外, 本研究发现GWP随土壤水分含量增加呈先增加后降低的趋势, 这表明土壤水分变化对GWP具有重要的影响, 适度土壤水分含量提升了该湿地的GWP, 而土壤水分含量过低或过高都会降低GWP。而在高盐分下CH4:CO2随土壤水分含量的增加而增加, 而在低盐分下不同水分处理之间差异不显著, 这表明在盐分含量高的滨海盐沼湿地土壤水分变化对CH4:CO2具有显著影响, 较低的土壤水分含量可以降低CH4:CO2, 而水分增加将会使CH4:CO2升高。此外, 本研究还发现土壤CH4排放与GWP以及CH4:CO2呈显著正相关关系, 且系数比土壤CO2排放大, 这表明CH4排放作为该滨海湿地碳库矿化输出的重要部分, 不应该被忽视, 并应在未来研究中加强该过程的研究。
3.2 土壤盐分对土壤碳矿化的影响
土壤盐分高是滨海盐沼湿地的重要环境特征。当外部环境盐浓度上升时, 微生物通常需要在细胞质中积累并产生大量渗透液, 以防止细胞脱水, 并维持水分平衡; 盐分过高则会不断提高土壤微生物细胞外的渗透压, 引起土壤微生物出现脱水、休眠甚至死亡; 还可以直接对微生物产生离子(如SO42-、Na+、Cl-)毒性, 导致微生物生物量和活性降低, 进而影响土壤碳矿化过程(Empadinhas & da Costa, 2008; Rath & Rousk, 2015)。例如, Mavi等(2012)通过室内培养实验研究表明, 高土壤盐分提高了微生物的胞外渗透势, 降低了土壤微生物活性, 进而显著降低了土壤CO2排放量。Setia等(2011)在澳大利亚南部沿海地区研究发现盐分增加均显著抑制了不同质地的土壤碳矿化。本研究发现土壤盐分升高对微生物数量和细菌群落α多样性指数均产生了显著抑制作用, 且不同盐分处理间的土壤细菌聚类远, 土壤群落结构差异大, 这表明土壤盐分是影响该滨海湿地土壤微生物群落活性和结构的重要因素。统计分析表明土壤盐分升高降低了CO2排放量, 因此本研究推测土壤盐度升高引起的微生物生物量的降低和群落结构的变化对土壤CO2排放产生了积极影响。本研究发现, CO2排放量与土壤DOC含量呈显著正相关关系, 表明土壤DOC有效性对碳矿化至关重要。一般而言, 土壤高盐分含量易导致土壤颗粒絮凝, 进而降低土壤有机质(SOM)的溶解度、土壤DOC的迁移和可利用性(Ury et al., 2022)。例如, Zhao等(2018)在黄河三角洲盐沼湿地研究发现盐分与DOC含量存在显著负相关关系。然而, Wong等(2010)发现, 在高土壤水分含量条件下, 土壤盐离子增加却有助于打破土壤团聚体, 提高SOM溶解度, 从而增加DOC含量, 这也是影响滨海盐沼湿地土壤碳矿化功能的一个重要因素。本研究发现水盐变化对土壤DOC含量具有弱交互作用, 低土壤水分条件下盐分增加引起DOC含量降低, 而在高土壤水分下盐分增加却提高了DOC含量。
CH4产生和硫酸盐还原是滨海盐沼湿地有机底物厌氧矿化的两个重要过程, 产甲烷菌可利用底物醋酸产生CH4, 而硫酸盐还原菌可利用醋酸产生CO2和H2S, 并抑制CH4产生。滨海盐沼湿地土壤中含有大量的SO42-, 可使土壤产CH4过程向SO42-还原过程转变, 从而降低土壤CH4排放量(张子川等, 2015)。Baldwin等(2006)采用小型生态系研究方法, 向淡水湿地添加不同浓度NaCl溶液, 结果发现盐度升高显著降低CH4产生率。Poffenbarger等(2011)分析了来自多个滨海湿地的CH4和盐分数据, 发现由于具有更高盐度的滨海湿地SO42-浓度高以及硫酸盐还原菌活性高, 释放的CH4显著低于低盐湿地。在黄河三角洲滨海盐沼湿地, 土壤盐分与土壤微生物量、活性及多样性呈显著负相关关系, 这主要与盐分升高导致的土壤渗透压的增加引起了某些微生物类群的死亡消失, 进而导致土壤微生物群落多样性下降有关(王震宇等, 2009)。本研究发现土壤盐分含量的增加对土壤CH4累积排放量、微生物生物量、细菌群落丰富度及多样性具有显著的抑制作用, 且不同盐分处理中土壤细菌群落结构差异大。因此我们推测, 类似于土壤盐度对CO2排放的影响, 盐分升高通过降低厌氧微生物生物量、细菌群落丰富度及多样性指数, 进而抑制滨海湿地土壤CH4产生和排放。同时本研究发现, GWP和CH4:CO2均随土壤盐分含量的增加而显著降低, 这表明土壤盐分变化通过影响土壤条件以及微生物特征对滨海盐沼湿地土壤有机碳分解方式、气体排放比例及总量产生了积极影响。
4 结论
研究水盐浓度变化对土壤碳矿化(CO2和CH4排放)的影响, 将对滨海盐沼湿地生态系统土壤碳循环及积累机制具有重要的意义。本研究结果表明, 水盐变化对土壤CO2和CH4排放量没有显著交互作用。土壤CO2排放量随水分含量的增加呈先增后减的单峰型变化趋势, CH4排放量则是随着含水量的增加而升高。然而, 土壤盐分的升高对土壤CO2和CH4排放均具有显著抑制作用。在气候变化背景下, 黄河三角洲气候暖干化趋势明显, 这将会进一步影响土壤水盐环境及植物、微生物特征, 从而对该区域滨海湿地碳循环过程和碳汇功能产生深远影响。因此, 未来应加强对土壤水盐运移如何影响滨海湿地碳循环过程和碳汇形成机制的研究, 以期为预测和模拟气候变化背景下滨海盐沼湿地碳汇功能的潜在变化趋势提供理论支持。
参考文献
Depth-distribution patterns and control of soil organic carbon in coastal salt marshes with different plant covers
DOI:10.1038/srep34835 [本文引用: 1]
The short-term effects of salinization on anaerobic nutrient cycling and microbial community structure in sediment from a freshwater wetland
DOI:10.1672/0277-5212(2006)26[455:TSEOSO]2.0.CO;2 URL [本文引用: 1]
Soil moisture is the major factor influencing microbial community structure and enzyme activities across seven biogeoclimatic zones in western Canada
DOI:10.1016/j.soilbio.2011.09.003 URL [本文引用: 1]
Elucidating the archaeal community and functional potential in two typical coastal wetlands with different stress patterns
DOI:10.1016/j.jclepro.2020.124894 [本文引用: 1]
Microbial and environmental controls of methane fluxes along a soil moisture gradient in a Pacific coastal temperate rainforest
DOI:10.1007/s10021-016-0003-1 URL [本文引用: 1]
Lower mineralizability of soil carbon with higher legacy soil moisture
DOI:10.1016/j.soilbio.2018.12.006 URL [本文引用: 2]
Temperature sensitivity of soil carbon decomposition and feedbacks to climate change
DOI:10.1038/nature04514 [本文引用: 1]
Impact of sea level change on coastal soil organic matter, priming effects and prokaryotic community assembly
DOI:10.1093/femsec/fiz129 [本文引用: 1]
Osmoadaptation mechanisms in prokaryotes: distribution of compatible solutes
PMID:18843593

Microorganisms respond to osmotic stress mostly by accumulating compatible solutes, either by uptake from the medium or by de novo synthesis. These osmotically active molecules preserve the positive turgor pressure required for cell division. The diversity of compatible solutes is large but falls into a few major chemical categories; they are usually small organic molecules such as amino acids or their derivatives, and carbohydrates or their derivatives. Some are widely distributed in nature while others seem to be exclusively present in specific groups of organisms. This review discusses the diversity and distribution of known classes of compatible solutes found in prokaryotes as well as the increasing knowledge of the genes and pathways involved in their synthesis. The alternative roles of some archetypal compatible solutes not subject to osmoregulatory constraints are also discussed.
Global warming, carbon emissions and uncertainty
全球变暖、碳排放及不确定性
The priming effect of organic matter: a question of microbial competition
DOI:10.1016/S0038-0717(03)00123-8 URL [本文引用: 1]
Precipitation events reduce soil respiration in a coastal wetland based on four-year continuous field measurements
Relationships between salt marsh vegetation distribution/diversity and soil chemical factors in the Yellow River Estuary, China
黄河河口盐沼植被分布、多样性与土壤化学因子的相关关系
Sea-level rise and coastal groundwater inundation and shoaling at select sites in California, USA
DOI:10.1016/j.ejrh.2015.12.055 URL [本文引用: 1]
Optimum soil water for soil respiration before and after amendment with glucose in humid tropical acrisols and a boreal mor layer
DOI:10.1016/S0038-0717(00)00073-0 URL [本文引用: 1]
The fumigation-extraction method to estimate soil microbial biomass: calibration of the kEC value
DOI:10.1016/0038-0717(95)00102-6 URL [本文引用: 1]
Selenium and salt mobilization in wetland and arid upland soils of Pariette Draw, Utah (USA)
DOI:10.1016/j.geoderma.2017.06.028 URL [本文引用: 1]
Climate-driven thresholds in reactive mineral retention of soil carbon at the global scale
DOI:10.1038/s41558-018-0341-4
[本文引用: 1]
Soil organic matter can release carbon dioxide to the atmosphere as the climate warms. Organic matter sorbed to reactive (iron- and aluminium-bearing) soil minerals is an important mechanism for long-term carbon storage. However, the global distribution of mineral-stored carbon across climate zones and consequently its overall contribution to the global soil carbon pool is poorly known. We measured carbon held by reactive minerals across a broad range of climates. Carbon retained by reactive minerals was found to contribute between 3 and 72% of organic carbon found in mineral soil, depending on mean annual precipitation and potential evapotranspiration. Globally, we estimate similar to 600 Gt of soil carbon is retained by reactive minerals, with most occurring in wet forested biomes. For many biomes, the fraction of organic carbon retained by reactive minerals is responsive to slight shifts in effective moisture, suggesting high sensitivity to future changes in climate.
Production, oxidation, emission and consumption of methane by soils: a review
DOI:10.1016/S1164-5563(01)01067-6 URL [本文引用: 1]
Effects of flooding and warming on soil organic matter mineralization in Avicennia germinans mangrove forests and Juncus roemerianus salt marshes
Fungi drive soil multifunctionality in the coastal salt marsh ecosystem
DOI:10.1016/j.scitotenv.2021.151673 [本文引用: 1]
Effects of changes in precipitation on soil respiration in coastal wetlands of the Yellow River Delta
降雨量改变对黄河三角洲滨海湿地土壤呼吸的影响
Salinity and sodicity affect soil respiration and dissolved organic matter dynamics differentially in soils varying in texture
DOI:10.1016/j.soilbio.2011.10.003 URL [本文引用: 1]
Crop rotation complexity regulates the decomposition of high and low quality residues
DOI:10.1016/j.soilbio.2014.07.027 URL [本文引用: 1]
A blueprint for blue carbon: toward an improved understanding of the role of vegetated coastal habitats in sequestering CO2
DOI:10.1890/110004 URL [本文引用: 1]
Salinity influence on methane emissions from tidal marshes
DOI:10.1007/s13157-011-0197-0 URL [本文引用: 1]
Comparative toxicities of salts on microbial processes in soil
DOI:10.1128/AEM.04052-15
PMID:26801570
[本文引用: 1]
Soil salinization is a growing threat to global agriculture and carbon sequestration, but to date it remains unclear how microbial processes will respond. We studied the acute response to salt exposure of a range of anabolic and catabolic microbial processes, including bacterial (leucine incorporation) and fungal (acetate incorporation into ergosterol) growth rates, respiration, and gross N mineralization and nitrification rates. To distinguish effects of specific ions from those of overall ionic strength, we compared the addition of four salts frequently associated with soil salinization (NaCl, KCl, Na2SO4, and K2SO4) to a nonsaline soil. To compare the tolerance of different microbial processes to salt and to interrelate the toxicity of different salts, concentration-response relationships were established. Growth-based measurements revealed that fungi were more resistant to salt exposure than bacteria. Effects by salt on C and N mineralization were indistinguishable, and in contrast to previous studies, nitrification was not found to be more sensitive to salt exposure than other microbial processes. The ion-specific toxicity of certain salts could be observed only for respiration, which was less inhibited by salts containing SO4(2-) than Cl(-) salts, in contrast to the microbial growth assessments. This suggested that the inhibition of microbial growth was explained solely by total ionic strength, while ion-specific toxicity also should be considered for effects on microbial decomposition. This difference resulted in an apparent reduction of microbial growth efficiency in response to exposure to SO4(2-) salts but not to Cl(-) salts; no evidence was found to distinguish K(+) and Na(+) salts.Copyright © 2016, American Society for Microbiology. All Rights Reserved.
Salt effects on the soil microbial decomposer community and their role in organic carbon cycling: a review
DOI:10.1016/j.soilbio.2014.11.001 URL [本文引用: 1]
Linking soil microbial activity to water- and air-phase contents and diffusivities
DOI:10.2136/sssaj2003.1560 URL [本文引用: 1]
Salinity effects on carbon mineralization in soils of varying texture
DOI:10.1016/j.soilbio.2011.05.013 URL [本文引用: 1]
Scientists raise alarm over “dangerously fast” growth in atmospheric methane
DOI:10.1038/d41586-022-00312-2 [本文引用: 1]
Saltwater intrusion in context: soil factors regulate impacts of salinity on soil carbon cycling
DOI:10.1007/s10533-021-00869-6 [本文引用: 1]
Regulators of coastal wetland methane production and responses to simulated global change
DOI:10.5194/bg-14-431-2017
URL
[本文引用: 2]
. Wetlands are the largest natural source of methane (CH4) emissions to the atmosphere, which vary along salinity and productivity gradients. Global change has the potential to reshape these gradients and therefore alter future contributions of wetlands to the global CH4 budget. Our study examined CH4 production along a natural salinity gradient in fully inundated coastal Alaska wetlands. In the laboratory, we incubated natural sediments to compare CH4 production rates between non-tidal freshwater and tidal brackish wetlands, and quantified the abundances of methanogens and sulfate-reducing bacteria in these ecosystems. We also simulated seawater intrusion and enhanced organic matter availability, which we predicted would have contrasting effects on coastal wetland CH4 production. Tidal brackish wetlands produced less CH4 than non-tidal freshwater wetlands probably due to high sulfate availability and generally higher abundances of sulfate-reducing bacteria, whereas non-tidal freshwater wetlands had significantly greater methanogen abundances. Seawater addition experiments with freshwater sediments, however, did not reduce CH4 production, perhaps because the 14-day incubation period was too short to elicit a shift in microbial communities. In contrast, increased organic matter enhanced CH4 production in 75 % of the incubations, but this response depended on the macrophyte species added, with half of the species treatments having no significant effect. Our study suggests that CH4 production in coastal wetlands, and therefore their overall contribution to the global CH4 cycle, will be sensitive to increased organic matter availability and potentially seawater intrusion. To better predict future wetland contributions to the global CH4 budget, future studies and modeling efforts should investigate how multiple global change mechanisms will interact to impact CH4 dynamics.\n
Effect of varying soil water potentials on methanogenesis in aerated marshland soils
DOI:10.1038/s41598-017-14980-y [本文引用: 1]
Research progresses on methanogenesis pathway and methanogens in coastal wetlands
滨海湿地甲烷产生途径和产甲烷菌研究进展
DOI:10.13287/j.1001-9332.201603.014
[本文引用: 1]
滨海湿地在全球碳循环中起着重要的作用,其甲烷排放量占全球海洋甲烷排放的75%.本文综述了滨海湿地主要甲烷产生途径、产甲烷菌种类及其影响因子.滨海湿地SO<sub>4</sub><sup>2-</sup>含量丰富,乙酸发酵和H<sub>2</sub>/CO<sub>2</sub>途径产甲烷受抑制,乙酸营养型和氢营养型产甲烷菌丰度较低;而利用甲胺类等“非竞争性”底物的C1甲基化合物歧化途径不受硫酸还原菌竞争底物的限制,兼性营养型产甲烷菌成为产甲烷优势菌.盐度与SO<sub>4</sub><sup>2-</sup>含量和植被类型密切相关,影响竞争性电子和产甲烷底物的种类和含量,对甲烷产生途径和产甲烷菌群落结构有重要影响.目前,滨海湿地产甲烷菌群落结构、甲烷产生途径的关键控制因素尚需明确,其对甲烷排放的影响有待进一步研究.
Microbial community characteristics in a degraded wetland of the Yellow River Delta
黄河三角洲退化湿地微生物特性的研究
Soil carbon dynamics in saline and sodic soils: a review
DOI:10.1111/j.1475-2743.2009.00251.x URL [本文引用: 1]
Salinity influence on soil microbial respiration rate of wetland in the Yangtze River estuary through changing microbial community
DOI:10.1016/j.jes.2014.07.016 URL [本文引用: 1]
Estimation of soil organic carbon storage in coastal wetlands of modern Yellow River Delta based on landscape pattern
基于景观格局的现代黄河三角洲滨海湿地土壤有机碳储量估算
Co-effects of salinity and moisture on CO2and N2O emissions of laboratory-incubated salt-affected soils from different vegetation types
DOI:10.1016/j.geoderma.2018.06.025 URL [本文引用: 1]
Effects of seawater and NaCl solution pulses on methane production potential from laboratory-incubated tidal freshwater and brackish marsh soil
盐分对河口淡水、微咸水沼泽湿地土壤甲烷产生潜力的影响
Effects of water and salinity regulation measures on soil carbon sequestration in coastal wetlands of the Yellow River Delta
DOI:10.1016/j.geoderma.2017.10.058 URL [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}