
植物生态学报 ›› 2025, Vol. 49 ›› Issue (9): 1410-1423.DOI: 10.17521/cjpe.2024.0467 cstr: 32100.14.cjpe.2024.0467
郭志红1( ), 杨妮4,5, 张涛4,5, 李海波4,5, 田太安4,5, 黄小波1,2,3, 李聪1, 马驷驹1, 苏建荣1,2,3, 李帅锋1,2,3,*()(
), 杨妮4,5, 张涛4,5, 李海波4,5, 田太安4,5, 黄小波1,2,3, 李聪1, 马驷驹1, 苏建荣1,2,3, 李帅锋1,2,3,*()( )
)
收稿日期:2024-12-24
接受日期:2025-04-08
出版日期:2025-09-20
发布日期:2025-04-09
通讯作者:
*李帅锋: ORCID: 0000-0002-2555-1808 (shuaifengli@163.com)基金资助:
GUO Zhi-Hong1(), YANG Ni4,5, ZHANG Tao4,5, LI Hai-Bo4,5, TIAN Tai-An4,5, HUANG Xiao-Bo1,2,3, LI Cong1, MA Si-Ju1, SU Jian-Rong1,2,3, LI Shuai-Feng1,2,3,*()()
Received:2024-12-24
Accepted:2025-04-08
Online:2025-09-20
Published:2025-04-09
Supported by:摘要: 生态位理论和中性理论所阐释的群落构建过程对生物多样性维持机制至关重要, 是森林生态学研究热点, 山地具有丰富的生物多样性, 而沿海拔梯度上菌根植物功能多样性与群落构建过程变化格局的研究仍相对较少。该研究基于梵净山国家级自然保护区海拔600-2 100 m建立的8个1 hm2动态监测样地, 将胸径≥1 cm的261种木本植物分为3个功能群: 丛枝菌根(AM)植物、外生菌根(EcM)植物和杜鹃花类菌根(ErM)植物, 分析其群落构建过程与功能多样性沿海拔梯度的变化格局, 揭示构建过程在维持功能多样性中的潜在作用。结果表明: 不同菌根植物功能多样性随海拔变化差异显著, AM和EcM植物功能丰富度与功能分散度随海拔升高呈现显著下降趋势, AM和EcM植物的叶面积和比叶面积功能性状加权平均值随海拔升高呈减小趋势, EcM植物叶干物质含量、叶片氮含量和磷含量随海拔升高呈增加趋势, ErM植物的叶面积、叶干物质含量、叶片磷含量随海拔升高呈增加趋势。3种菌根植物群落构建均由随机性过程主导, 其中ErM植物的漂变过程对群落构建的贡献均比AM和EcM植物大。β最近亲缘指数(βNTI)对AM植物的功能多样性没有显著影响, 而对EcM和ErM植物的功能性状加权平均值影响显著, βNTI对EcM和ErM植物功能多样性具有显著正效应, 维持了其功能多样性。此外, 土壤养分(土壤有机碳、全氮和水解性氮)含量对AM和EcM植物的功能多样性具有显著正效应, 而对ErM植物功能多样性具有显著负效应; 海拔对AM和ErM植物具有显著负效应, 而对EcM植物具有显著正效应。研究结果为揭示西南山地生物多样性维持机制提供科学依据, 对于中亚热带天然林保护与修复具有重要意义。
郭志红, 杨妮, 张涛, 李海波, 田太安, 黄小波, 李聪, 马驷驹, 苏建荣, 李帅锋. 梵净山天然林菌根植物功能多样性与群落构建沿海拔梯度的变化. 植物生态学报, 2025, 49(9): 1410-1423. DOI: 10.17521/cjpe.2024.0467
GUO Zhi-Hong, YANG Ni, ZHANG Tao, LI Hai-Bo, TIAN Tai-An, HUANG Xiao-Bo, LI Cong, MA Si-Ju, SU Jian-Rong, LI Shuai-Feng. Change of functional diversity and community assembly of mycorrhizal plant along an altitudinal gradient in primary forests of Fanjing Mountain, Guizhou, China. Chinese Journal of Plant Ecology, 2025, 49(9): 1410-1423. DOI: 10.17521/cjpe.2024.0467
| 菌根类型 Mycorrhizal type | 海拔 Altitude (m) | 平均胸径 Average diameter (cm) | 平均高度 Average height (m) | 多度 Abundance | 香农-威纳指数 Shannon-Wiener index |
|---|---|---|---|---|---|
| 丛枝菌根植物 Arbuscular mycorrhizal plant | 600 | 4.47 ± 5.46d | 6.16 ± 6.09d | 3 424 | 0.24 |
| 700 | 5.17 ± 6.87c | 7.23 ± 6.19b | 3 767 | 0.25 | |
| 850 | 5.65 ± 5.79b | 7.54 ± 6.36a | 4 195 | 0.28 | |
| 1 000 | 3.93 ± 4.09e | 6.13 ± 3.85d | 5 746 | 0.32 | |
| 1 050 | 5.25 ± 5.57c | 7.17 ± 6.07b | 3 455 | 0.24 | |
| 1 300 | 4.67 ± 5.85d | 6.27 ± 6.5cd | 4 002 | 0.27 | |
| 1 600 | 5.62 ± 7.39b | 5.24 ± 3.51e | 2 376 | 0.2 | |
| 2 100 | 8.91 ± 3.70a | 6.43 ± 3.00c | 3 249 | 0.24 | |
| 合计 Total | 5.46 ± 5.59 | 6.52 ± 5.19 | 30 214 | - | |
| 外生菌根植物 Ectomycorrhizal plant | 600 | 12.68 ± 20.72c | 9.43 ± 9.40d | 404 | 0.22 |
| 700 | 8.20 ± 12.34e | 8.19 ± 7.40e | 664 | 0.28 | |
| 850 | 8.44 ± 8.89e | 9.08 ± 7.27de | 422 | 0.22 | |
| 1 000 | 11.17 ± 11.01d | 11.56 ± 7.15c | 937 | 0.32 | |
| 1 050 | 22.90 ± 11.95a | 21.46 ± 10.42a | 481 | 0.23 | |
| 1 300 | 14.32 ± 10.95b | 13.48 ± 10.36b | 778 | 0.3 | |
| 1 600 | 13.65 ± 12.91bc | 9.04 ± 7.97de | 564 | 0.25 | |
| 2 100 | 13.25 ± 8.21 bc | 8.04 ± 4.71e | 395 | 0.22 | |
| 合计 Total | 13.08 ± 12.12 | 11.28 ± 8.09 | 4 645 | - | |
| 杜鹃花类菌根植物 Ericoid mycorrhizal plant | 600 | 4.61 ± 4.81c | 4.59 ± 2.49cd | 62 | 0.05 |
| 700 | 7.07 ± 5.68b | 6.85 ± 3.59b | 26 | 0.05 | |
| 850 | 4.92 ± 2.68c | 5.05 ± 2.19c | 153 | 0.11 | |
| 1 000 | 2.35 ± 0.86e | 4.09 ± 1.36e | 641 | 0.28 | |
| 1 050 | 2.64 ± 1.00d | 4.47 ± 1.19d | 2 972 | 0.29 | |
| 1 300 | 2.54 ± 1.13d | 3.80 ± 1.58f | 735 | 0.29 | |
| 1 600 | 4.17 ± 7.41c | 4.60 ±1.71d | 41 | 0.05 | |
| 2 100 | 19.75 ± 1.51a | 9.80 ± 1.25a | 83 | 0.08 | |
| 合计 Total | 6.00 ± 3.14 | 5.40 ± 1.92 | 4 713 | - |
表1 丛枝菌根、外生菌根和杜鹃花类菌根植物多样性及群落结构特征(平均值±标准差)
Table 1 Plant diversity and community characteristics of arbuscular mycorrhizal (AM), ectomycorrhizal (EcM) and ericoid mycorrhizal (ErM) plant species (mean ± SD)
| 菌根类型 Mycorrhizal type | 海拔 Altitude (m) | 平均胸径 Average diameter (cm) | 平均高度 Average height (m) | 多度 Abundance | 香农-威纳指数 Shannon-Wiener index |
|---|---|---|---|---|---|
| 丛枝菌根植物 Arbuscular mycorrhizal plant | 600 | 4.47 ± 5.46d | 6.16 ± 6.09d | 3 424 | 0.24 |
| 700 | 5.17 ± 6.87c | 7.23 ± 6.19b | 3 767 | 0.25 | |
| 850 | 5.65 ± 5.79b | 7.54 ± 6.36a | 4 195 | 0.28 | |
| 1 000 | 3.93 ± 4.09e | 6.13 ± 3.85d | 5 746 | 0.32 | |
| 1 050 | 5.25 ± 5.57c | 7.17 ± 6.07b | 3 455 | 0.24 | |
| 1 300 | 4.67 ± 5.85d | 6.27 ± 6.5cd | 4 002 | 0.27 | |
| 1 600 | 5.62 ± 7.39b | 5.24 ± 3.51e | 2 376 | 0.2 | |
| 2 100 | 8.91 ± 3.70a | 6.43 ± 3.00c | 3 249 | 0.24 | |
| 合计 Total | 5.46 ± 5.59 | 6.52 ± 5.19 | 30 214 | - | |
| 外生菌根植物 Ectomycorrhizal plant | 600 | 12.68 ± 20.72c | 9.43 ± 9.40d | 404 | 0.22 |
| 700 | 8.20 ± 12.34e | 8.19 ± 7.40e | 664 | 0.28 | |
| 850 | 8.44 ± 8.89e | 9.08 ± 7.27de | 422 | 0.22 | |
| 1 000 | 11.17 ± 11.01d | 11.56 ± 7.15c | 937 | 0.32 | |
| 1 050 | 22.90 ± 11.95a | 21.46 ± 10.42a | 481 | 0.23 | |
| 1 300 | 14.32 ± 10.95b | 13.48 ± 10.36b | 778 | 0.3 | |
| 1 600 | 13.65 ± 12.91bc | 9.04 ± 7.97de | 564 | 0.25 | |
| 2 100 | 13.25 ± 8.21 bc | 8.04 ± 4.71e | 395 | 0.22 | |
| 合计 Total | 13.08 ± 12.12 | 11.28 ± 8.09 | 4 645 | - | |
| 杜鹃花类菌根植物 Ericoid mycorrhizal plant | 600 | 4.61 ± 4.81c | 4.59 ± 2.49cd | 62 | 0.05 |
| 700 | 7.07 ± 5.68b | 6.85 ± 3.59b | 26 | 0.05 | |
| 850 | 4.92 ± 2.68c | 5.05 ± 2.19c | 153 | 0.11 | |
| 1 000 | 2.35 ± 0.86e | 4.09 ± 1.36e | 641 | 0.28 | |
| 1 050 | 2.64 ± 1.00d | 4.47 ± 1.19d | 2 972 | 0.29 | |
| 1 300 | 2.54 ± 1.13d | 3.80 ± 1.58f | 735 | 0.29 | |
| 1 600 | 4.17 ± 7.41c | 4.60 ±1.71d | 41 | 0.05 | |
| 2 100 | 19.75 ± 1.51a | 9.80 ± 1.25a | 83 | 0.08 | |
| 合计 Total | 6.00 ± 3.14 | 5.40 ± 1.92 | 4 713 | - |
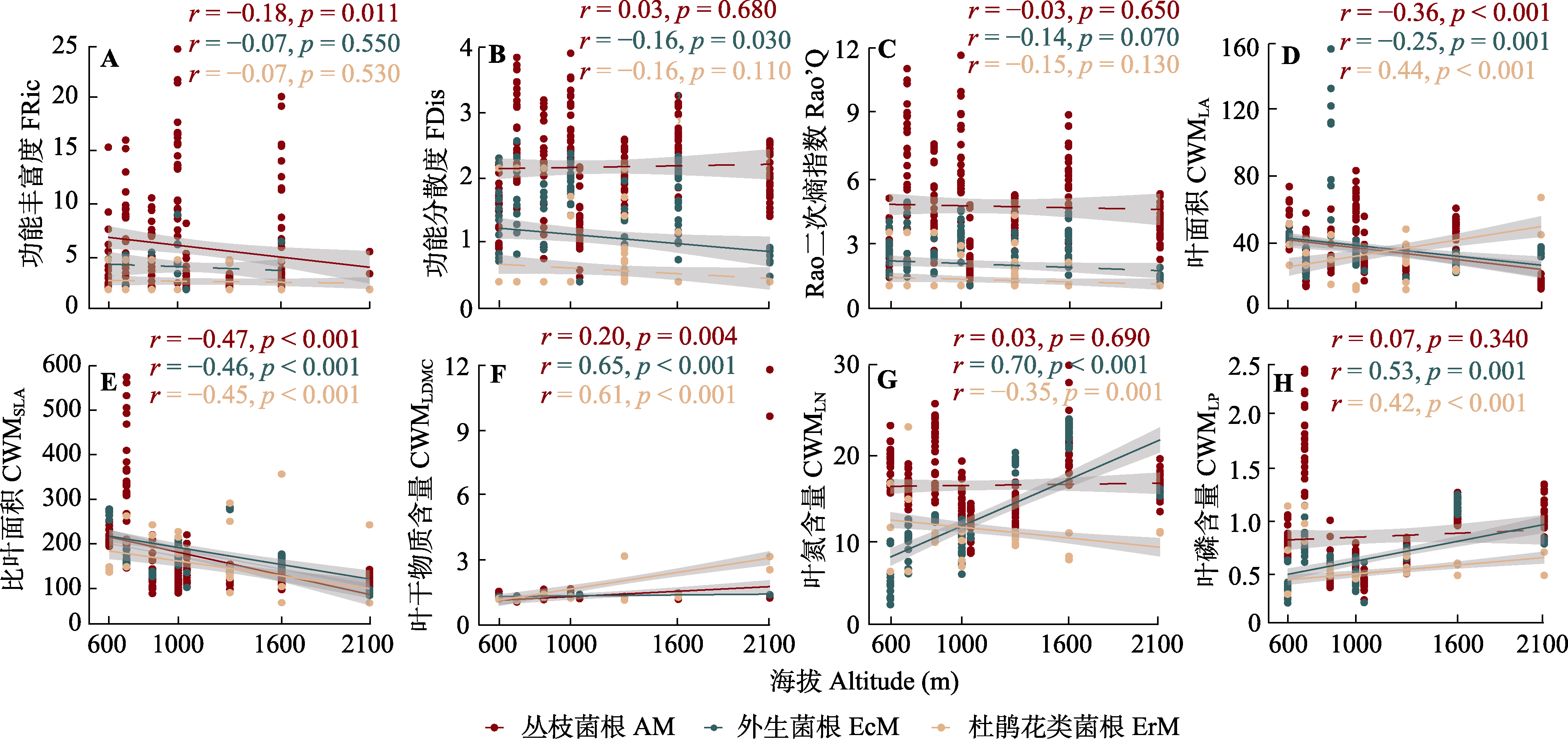
图1 丛枝菌根、外生菌根和杜鹃花类菌根植物的功能多样性随海拔高度变化的关系。CWM, 群落加权平均值。p < 0.05表示差异显著; p > 0.05表示没有显著差异。
Fig. 1 Functional diversity of arbuscular mycorrhizal (AM), ectomycorrhizal (EcM), and ericoid mycorrhizal (ErM) plant species in relation to the altitudinal gradient. CWM, community weighted mean; FRic, functional richness index; FDis, functional dispersion index; LA, leaf area; LDMC, leaf dry matter content; LN, leaf nitrogen content; LP, leaf phosphorus content; Rao’Q, Rao’s quadratic entropy index; SLA, specific leaf area. p < 0.05 indicates significant difference; p > 0.05 indicates no significant difference.
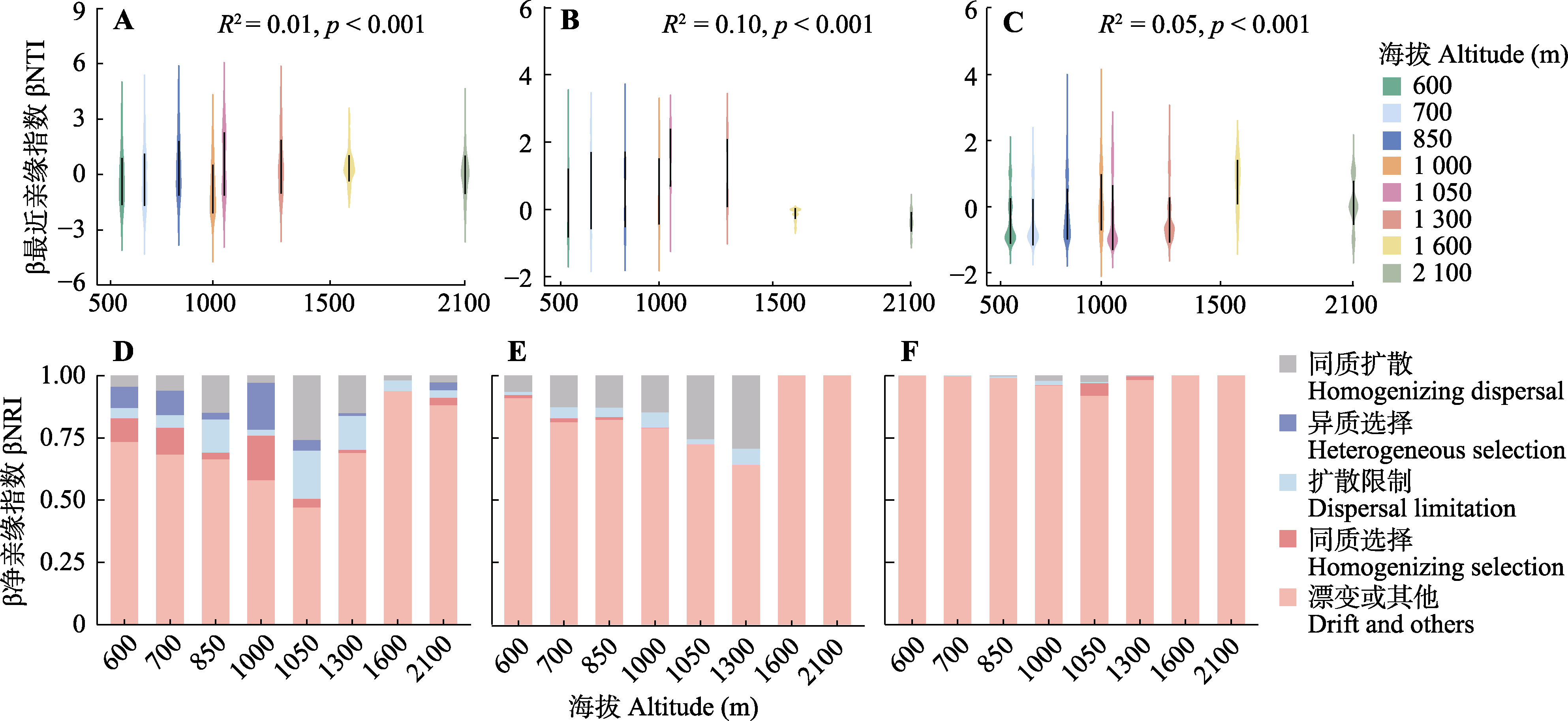
图2 丛枝菌根(A、D)、外生菌根(B、E)、杜鹃花类菌根(C、F)植物的群落构建过程(β最近亲缘指数和β净亲缘指数)以及与海拔高度的关系。p < 0.001表示差异显著。
Fig. 2 Community assembly processes (Beta Nearest Taxon Index (βNTI) and Beta Net Relatedness Index (βNRI)) and relationships with altitudinal gradients for arbuscular mycorrhizal (AM) (A, D), ectomycorrhizal (EcM) (B, E), and ericoid mycorrhizas (ErM) (C, F) plant species. p < 0.001 indicates significant difference.
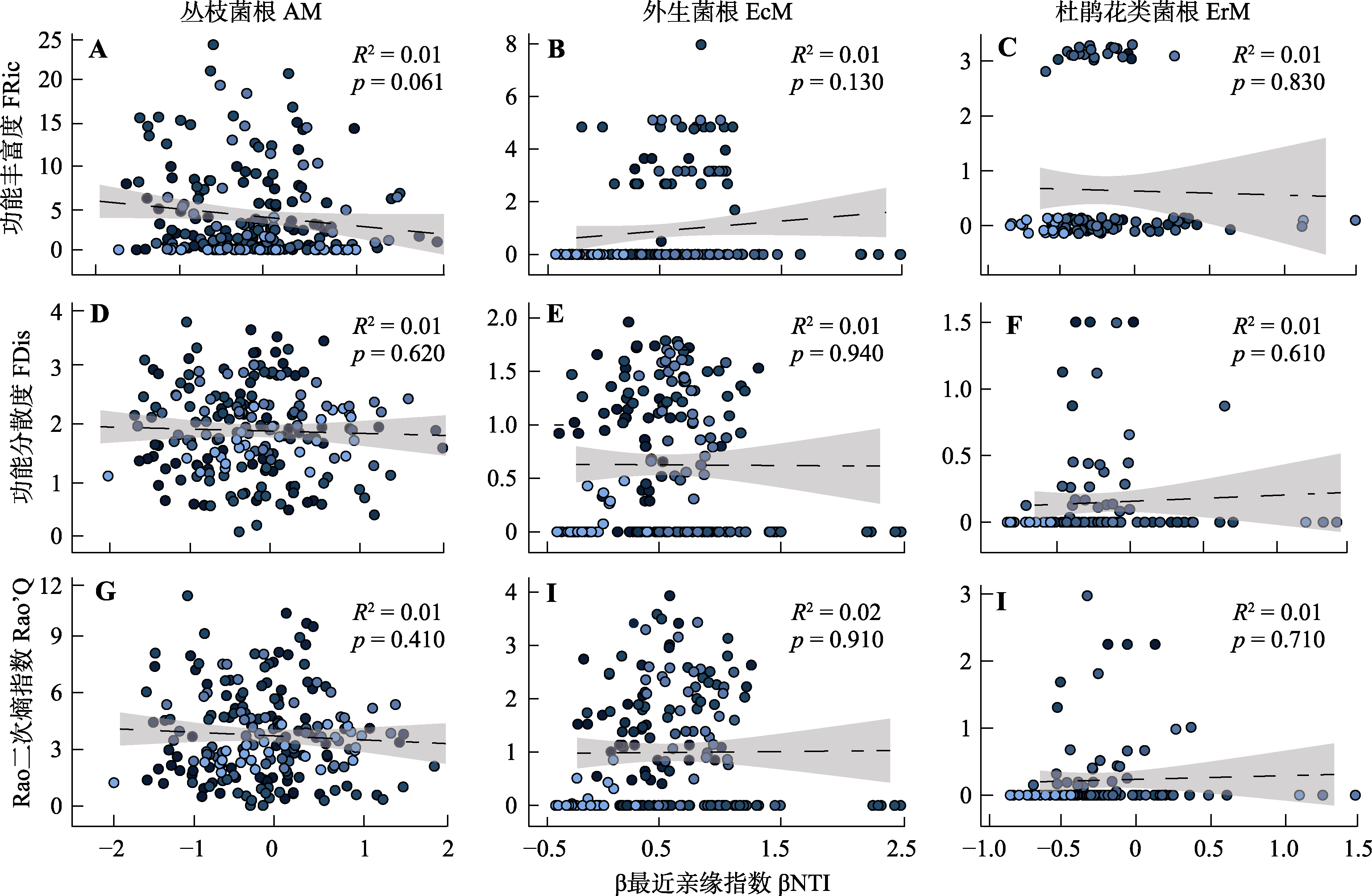
图3 丛枝菌根、外生菌根、杜鹃花类菌根植物的功能丰富度指数、功能分散度指数、Rao二次熵指数与β最近亲缘指数之间的线性关系。p < 0.05表示差异显著; p > 0.05表示没有显著差异。不同颜色圆点代表不同海拔高度, 颜色越深, 海拔越高。
Fig. 3 Linear relationships between functional richness index (FRic), functional dispersion index (FDis), Rao’s quadratic entropy index (Rao’Q) and community assembly processes (Beta Nearest Taxon Index, βNTI) for arbuscular mycorrhizal (AM), ectomycorrhizal (EcM), and ericoid mycorrhizal (ErM) plant species. p < 0.05 indicates significant differences; p > 0.05 indicates no significant differences. Different colored dots represent different altitudes; the darker color, the higher altitude.
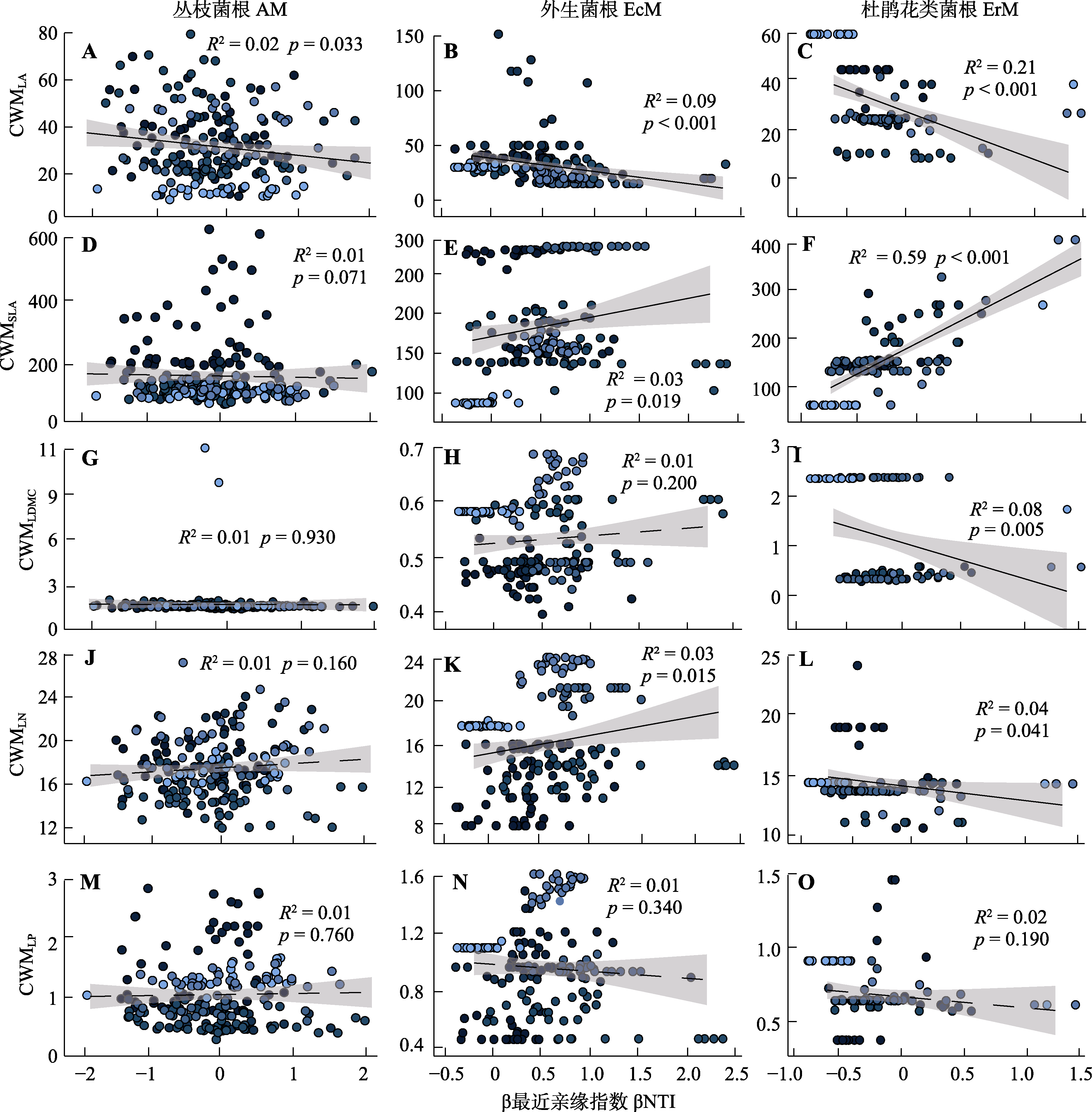
图4 丛枝菌根、外生菌根和杜鹃花类菌根植物的群落加权平均值与β最近亲缘指数之间的线性关系。p < 0.05表示差异显著; p > 0.05表示没有显著差异。CWM, 群落加权平均值; LA, 叶面积; LDMC, 叶干物质含量; LN, 叶氮含量; LP, 叶磷含量; SLA, 比叶面积。不同颜色圆点代表不同海拔高度, 颜色越深, 海拔越高。
Fig. 4 Linear relationships between community-weighted means (CWMLA, CWMSLA, CWMLDMC, CWMLN, CWMLP) and Beta Nearest Taxon Index, βNTI) for arbuscular mycorrhizal (AM), ectomycorrhizal (EcM), and ericoid mycorrhizal (ErM) plant species. p < 0.05 indicates significant differences; p > 0.05 indicates no significant difference. CWM, community weighted mean; LA, leaf area; LDMC, leaf dry matter content; LN, leaf nitrogen content; LP, leaf phosphorus content; SLA, specific leaf area. Different colored dots represent different altitudes; the darker color, the higher altitude.
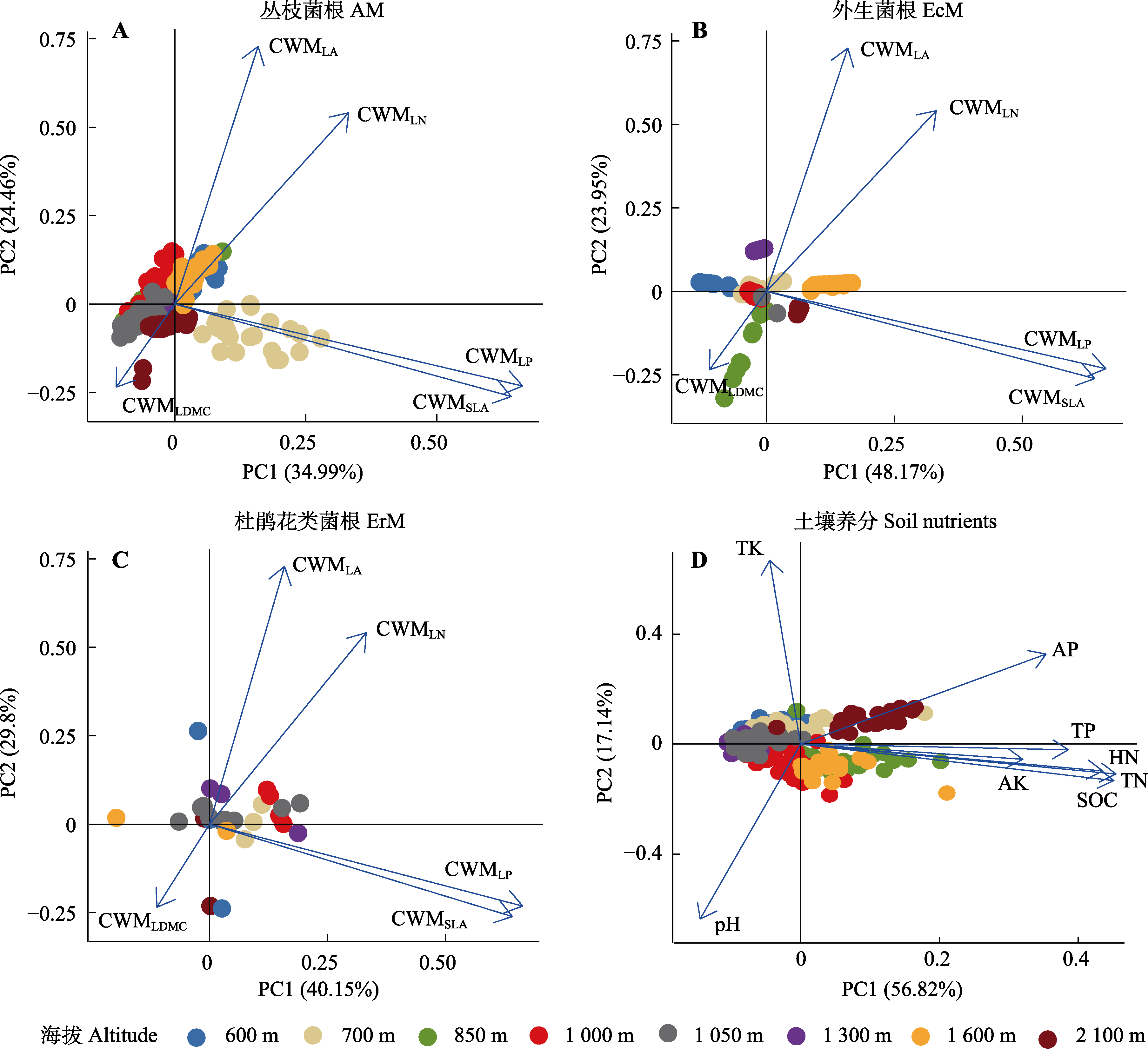
图5 丛枝菌根、外生菌根、杜鹃花类菌根植物的功能多样性以及土壤养分基于海拔高度的主成分(PC)分析。AK, 速效钾含量; AP, 速效磷含量; CWM, 群落加权平均值; HN, 水解性氮含量; LA, 叶面积; LDMC, 叶干物质含量; LN, 叶氮含量; LP, 叶磷含量; SLA, 比叶面积; SOC, 土壤有机碳含量; TK, 全钾含量; TN, 全氮含量; TP, 全磷含量。
Fig. 5 Principal component (PC) analysis of the functional diversity of arbuscular mycorrhizal (AM), ectomycorrhizal (EcM), and ericoid mycorrhizal (ErM) plant species, along with soil nutrient variables, across the altitudinal gradient. AK, available potassium content; AP, available phosphoru content; CWM, community weighted mean; HN, hydrolysable nitrogen content; LA, leaf area; LDMC, leaf dry matter content; LN, leaf nitrogen content; LP, leaf phosphorus content; SLA, specific leaf area; SOC, soil organic carbon content; TK, total potassium content; TN, total nitrogen content; TP, total phosphorus content.
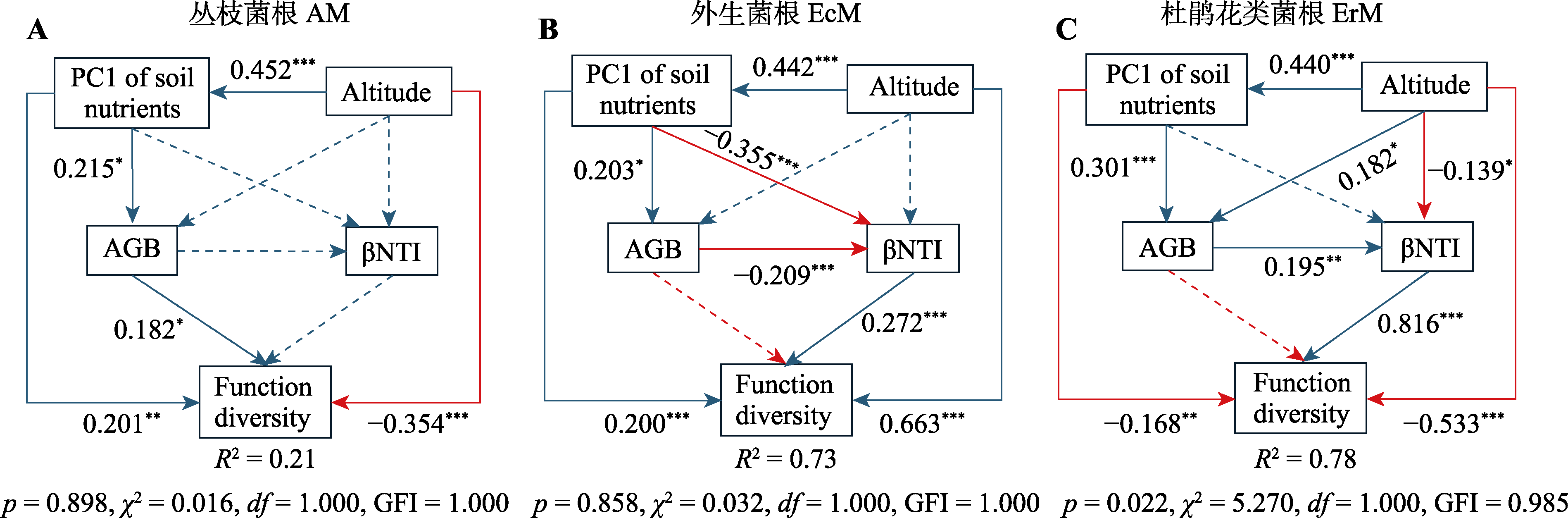
图6 海拔(Altitude)、土壤养分(PC1 of soil nutrients)含量、地上生物量(AGB)、β最近亲缘指数(βNTI)对丛枝菌根、外生菌根、杜鹃花类菌根植物功能多样性(Function diversity)的综合影响及效应值。绿线代表正效应, 红线代表负效应。虚线表示无显著影响(p > 0.05), 实线表示显著影响(p < 0.05)。*, p < 0.05; **, p < 0.01; ***, p < 0.001。土壤养分包括: HN, 水解性氮含量; SOC, 土壤有机碳含量; TN, 全氮含量。GFI, 模型拟合优度, GFI > 0.95代表模型拟合较好; 卡方检验p > 0.05代表模型具有较好的拟合效果。
Fig. 6 Structural equation model showing the combined effects and standardized path coefficients of altitude, PC1 of soil nutrients contents, aboveground biomass (AGB), and community assembly (Beta Nearest Taxon Index, βNTI) on the functional diversity of arbuscular mycorrhizal (AM), ectomycorrhizal (EcM), and ericoid mycorrhizal (ErM) plant species. Green lines represent positive effects and red lines represent negative effects. Dashed lines indicate no significant effect (p > 0.05), and solid lines indicate significant effect (p < 0.05). *, p < 0.05; **, p < 0.01; ***, p < 0.001. Soil nutrients including: HN, hydrolysable nitrogen content; SOC, soil organic carbon content; TN, total nitrogen content. GFI, model goodness of fit, GFI > 0.95 represents a better model fit; p > 0.05 in chi-square test, represents a better model fit.
| [1] |
Bahram M, Hildebrand F, Forslund SK, Anderson JL, Soudzilovskaia NA, Bodegom PM, Bengtsson-Palme J, Anslan S, Coelho LP, Harend H, Huerta-Cepas J, Medema MH, Maltz MR, Mundra S, Olsson PA, et al. (2018). Structure and function of the global topsoil microbiome. Nature, 560, 233-237.
DOI |
| [2] |
Bever JD, Dickie IA, Facelli E, Facelli JM, Klironomos J, Moora M, Rillig MC, Stock WD, Tibbett M, Zobel M (2010). Rooting theories of plant community ecology in microbial interactions. Trends in Ecology & Evolution, 25, 468-478.
DOI URL |
| [3] |
Bogar LM, Tavasieff OS, Raab TK, Peay KG (2022). Does resource exchange in ectomycorrhizal symbiosis vary with competitive context and nitrogen addition? New Phytologist, 233, 1331-1344.
DOI URL |
| [4] | Bongers FJ, Schmid B, Bruelheide H, Bongers F, Li S, von Oheimb G, Li Y, Cheng A, Ma K, Liu X (2021). Functional diversity effects on productivity increase with age in a forest biodiversity experiment. Nature Ecology & Evolution, 5, 1594-1603. |
| [5] |
Carmona CP, Azcárate FM, de Bello F, Ollero HS, Lepš J, Peco B (2012). Taxonomical and functional diversity turnover in Mediterranean grasslands: interactions between grazing, habitat type and rainfall. Journal of Applied Ecology, 49, 1084-1093.
DOI URL |
| [6] |
Cayuela L, Granzow-de la Cerda Í, Albuquerque FS, Golicher DJ (2012). Taxonstand: an r package for species names standardisation in vegetation databases. Methods in Ecology and Evolution, 3, 1078-1083.
DOI URL |
| [7] | Chen XL, Chen HYH, Chang SX (2022). Meta-analysis shows that plant mixtures increase soil phosphorus availability and plant productivity in diverse ecosystems. Nature Ecology & Evolution, 6, 1112-1121. |
| [8] |
Cornwell WK, Ackerly DD (2009). Community assembly and shifts in plant trait distributions across an environmental gradient in coastal California. Ecological Monographs, 79, 109-126.
DOI URL |
| [9] | de Bello F, Lavorel S, Hallett LM, Valencia E, Garnier E, Roscher C, Conti L, Galland T, Goberna M, Májeková M, Montesinos-Navarro A, Pausas JG, Verdú M, E-Vojtkó A, Götzenberger L, Lepš J (2021). Functional trait effects on ecosystem stability: assembling the jigsaw puzzle. Trends Trends in Ecology & Evolution, 36, 822-836. |
| [10] |
Díaz S, Purvis A, Cornelissen JHC, Mace GM, Donoghue MJ, Ewers RM, Jordano P, Pearse WD (2013). Functional traits, the phylogeny of function, and ecosystem service vulnerability. Ecology and Evolution, 3, 2958-2975.
DOI PMID |
| [11] | Dini-Andreote F, Stegen JC, van Elsas JD, Salles JF (2015). Disentangling mechanisms that mediate the balance between stochastic and deterministic processes in microbial succession. Proceedings of the National Academy of Sciences of the United States of America, 112, E1326-E1332. |
| [12] | Dong SK, Tang L, Zhang XF, Liu SL, Liu QR, Su XK, Zhang Y, Wu XY, Zhao ZZ, Li Y, Sha W (2017). Relationship between plant species diversity and functional diversity in alpine grasslands. Acta Ecologica Sinica, 37, 1472-1483. |
| [董世魁, 汤琳, 张相锋, 刘世梁, 刘全儒, 苏旭坤, 张勇, 武晓宇, 赵珍珍, 李钰, 沙威 (2017). 高寒草地植物物种多样性与功能多样性的关系. 生态学报, 37, 1472-1483.] | |
| [13] |
Duan S, Feng G, Limpens E, Bonfante P, Xie X, Zhang L (2024). Cross-kingdom nutrient exchange in the plant-arbuscular mycorrhizal fungus-bacterium continuum. Nature Reviews Microbiology, 22, 773-790.
DOI |
| [14] | Fan Y, Chen J, Shirkey G, John R, Wu SR, Park H, Shao C (2016). Applications of structural equation modeling (SEM) in ecological studies: an updated review. Ecological Processes, 5, 19. DOI: 10.1186/s13717-016-0063-3. |
| [15] |
Gao WQ, Lei XD, Gao DL, Li YT (2021). Mass-ratio and complementarity effects simultaneously drive aboveground biomass in temperate Quercus forests through stand structure. Ecology and Evolution, 11, 16806-16816.
DOI URL |
| [16] | Gilbert B, Levine JM (2017). Ecological drift and the distribution of species diversity. Proceedings of the Royal Society B: Biological Sciences, 284, 20170507. DOI: 10.1098/rspb.2017.0507. |
| [17] |
Gonzalez-Akre E, Piponiot C, Lepore M, Herrmann V, Lutz JA, Baltzer JL, Dick CW, Gilbert GS, He F, Heym M, Huerta AI, Jansen PA, Johnson DJ, Knapp N, Král K, et al. (2022). Allodb: an R package for biomass estimation at globally distributed extratropical forest plots. Methods in Ecology and Evolution, 13, 330-338.
DOI |
| [18] |
Jin Y, Qian H (2022). V. PhyloMaker2: an updated and enlarged R package that can generate very large phylogenies for vascular plants. Plant Diversity, 44, 335-339.
DOI |
| [19] |
Karandashov V, Bucher M (2005). Symbiotic phosphate transport in arbuscular mycorrhizas. Trends in Plant Science, 10, 22-29.
DOI PMID |
| [20] | Lei H, Chen L, Wang H, Qi X, Liu J, Ouyang S, Deng X, Lei P, Lin G, Kuzyakov Y, Xiang W (2022). Dominant mycorrhizal association of trees determines soil nitrogen availability in subtropical forests. Geoderma, 427, 116135. DOI: 10.1016/j.geoderma.2022.116135. |
| [21] |
Li SF, Lang XD, Huang XB, Wang YH, Liu WD, Xu CH, Su JR (2020). Association classification of a 30 hm2 dynamics plot in the monsoon broad-leaved evergreen forest in Pu’er, Yunnan, China. Chinese Journal of Plant Ecology, 44, 236-247.
DOI URL |
|
[李帅锋, 郎学东, 黄小波, 王艳红, 刘万德, 徐崇华, 苏建荣 (2020). 云南普洱30 hm2季风常绿阔叶林动态监测样地群丛数量分类. 植物生态学报, 44, 236-247.]
DOI |
|
| [22] | Li YP, Pan LJ, Chen J, Xu H, Yang LX (2024). Response and influencing factors of leaf functional traits to forest succession in subtropical mixed plantations. Biodiversity Science, 32(7), 6-18. |
| [李艳朋, 盘李军, 陈洁, 许涵, 杨立新 (2024). 亚热带人工混交林叶功能性状对森林演替的响应规律及影响因素. 生物多样性, 32(7), 6-18.] | |
| [23] | Liu Z, Fang J, Song B, Yang Y, Yu Z, Hu J, Dong K, Takahashi K, Adams JM (2023). Stochastic processes dominate soil arbuscular mycorrhizal fungal community assembly along an elevation gradient in central Japan. Science of the Total Environment, 855, 158941. DOI: 10.1016/j.scitotenv.2022.158941. |
| [24] |
Mammola S, Carmona CP, Guillerme T, Cardoso P (2021). Concepts and applications in functional diversity. Functional Ecology, 35, 1869-1885.
DOI URL |
| [25] | Mao Z, Wiegand T, Corrales A, Fang S, Hao Z, Lin F, Ye J, Yuan Z, Wang X (2024). Mycorrhizal types regulate tree spatial associations in temperate forests: ectomycorrhizal trees might favour species coexistence. Ecology Letters, 27, e70005. DOI: 10.1111/ele.70005. |
| [26] |
Martínez-Camilo R, González-Espinosa M, Ramírez-Marcial N, Cayuela L, Pérez-Farrera MÁ (2018). Tropical tree species diversity in a mountain system in southern Mexico: local and regional patterns and determinant factors. Biotropica, 50, 499-509.
DOI URL |
| [27] | Mi X, Feng G, Hu Y, Zhang J, Chen L, Corlett RT, Hughes AC, Pimm S, Schmid B, Shi S, Svenning JC, Ma K (2021). The global significance of biodiversity science in China: an overview. National Science Review, 8, nwab032. DOI: 10.1093/nsr/nwab032. |
| [28] |
Miller JED, Li D, LaForgia ML, Harrison SP (2019). Functional diversity is a passenger but not driver of drought-related plant diversity losses in annual grasslands. Journal of Ecology, 107, 2033-2039.
DOI |
| [29] | Niu KC, Liu YN, Shen ZH, He FL, Fang JY (2009). Community assembly: the relative importance of neutral theory and niche theory. Biodiversity Science, 17, 579. DOI: 10.3724/SP.J.1003.2009.09142. |
| [30] | Ray T, Delory BM, Beugnon R, Bruelheide H, Cesarz S, Eisenhauer N, Ferlian O, Quosh J, von Oheimb G, Fichtner A (2023). Tree diversity increases productivity through enhancing structural complexity across mycorrhizal types. Science Advances, 9, eadi2362. DOI: 10.1126/sciadv.adi2362. |
| [31] |
Rixen C, Wipf S, Rumpf SB, Giejsztowt J, Millen J, Morgan JW, Nicotra AB, Venn S, Zong S, Dickinson KJM, Freschet GT, Kurzböck C, Li J, Pan H, Pfund B, et al. (2022). Intraspecific trait variation in alpine plants relates to their elevational distribution. Journal of Ecology, 110, 860-875.
DOI URL |
| [32] |
Sachsenmaier L, Schnabel F, Dietrich P, Eisenhauer N, Ferlian O, Quosh J, Richter R, Wirth C (2024). Forest growth resistance and resilience to the 2018-2020 drought depend on tree diversity and mycorrhizal type. Journal of Ecology, 112, 1787-1803.
DOI URL |
| [33] | Shen KP, He YJ, Xia TT, Guo Y, Wu BL, Han X, Chen HC, Zhao Y, Wu P, Liu Y (2024). Arbuscular mycorrhizal fungi improve the competitive advantage of a native plant relative to a congeneric invasive plant in growth and nutrition. Ecology and Evolution, 14, e11459. DOI: 10.1002/ece3.11459. |
| [34] | Smith SE, Read D (2008). The symbionts forming arbuscular mycorrhizas//Smith SE, Read D. Mycorrhizal Symbiosis. Academic Press, New York. 13-41. |
| [35] | Soudzilovskaia NA, Vaessen S, Barcelo M, He J, Rahimlou S, Abarenkov K, Brundrett MC, Gomes SIF, Merckx V, Tedersoo L (2020). FungalRoot: global online database of plant mycorrhizal associations. New Phytologist, 227, 955-966. |
| [36] |
Teste FP, Kardol P, Turner BL, Wardle DA, Zemunik G, Renton M, Laliberté E (2017). Plant-soil feedback and the maintenance of diversity in Mediterranean-climate shrublands. Science, 355, 173-176.
DOI PMID |
| [37] |
van der Heijden MGA, Bardgett RD, van Straalen NM (2008). The unseen majority: soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems. Ecology Letters, 11, 296-310.
DOI PMID |
| [38] | Veresoglou SD, Xi J, Peñuelas J (2024). Mechanisms of coexistence: exploring species sorting and character displacement in woody plants to alleviate belowground competition. Ecology Letters, 27, e14489. DOI: 10.1111/ele.14489. |
| [39] | Wang H, Wu AJ, Liu BX, Liu RJ, Chen YL (2020). Interactions between mycorrhizal fungal diversity and plant diversity: a review. Microbiology China, 47, 3918-3932. |
| [王浩, 吴爱姣, 刘保兴, 刘润进, 陈应龙 (2020). 菌根真菌多样性与植物多样性的相互作用研究进展. 微生物学通报, 47, 3918-3932.] | |
| [40] | Ward EB, Duguid MC, Kuebbing SE, Lendemer JC, Bradford MA (2022). The functional role of ericoid mycorrhizal plants and fungi on carbon and nitrogen dynamics in forests. New Phytologist, 235, 1701-1718. |
| [41] | Ward EB, Polussa A, Bradford MA (2023). Depth-dependent effects of ericoid mycorrhizal shrubs on soil carbon and nitrogen pools are accentuated under arbuscular mycorrhizal trees. Global Change Biology, 29, 5924-5940. |
| [42] |
Washburne AD, Morton JT, Sanders J, McDonald D, Zhu Q, Oliverio AM, Knight R (2018). Methods for phylogenetic analysis of microbiome data. Nature Microbiology, 3, 652-661.
DOI PMID |
| [43] | Willing CE, Wan J, Yeam JJ, Cessna AM, Peay KG (2024). Arbuscular mycorrhizal fungi equalize differences in plant fitness and facilitate plant species coexistence through niche differentiation. Nature Ecology & Evolution, 8, 2058-2071. |
| [44] |
Xia TT, Shen KP, Guo Y, Chen DM, Han X, Wu BL, Zhao Y, Chen HC, Zhao Y, Yang XH, Lei ML, Xiao JH, He YJ (2023). AM fungus improves the competitiveness of a native plant against an invasive plant under moderate soil P supply. Plant and Soil, 492, 541-556.
DOI |
| [45] | Xu JS, Zhang YH, Liu SQ, Chai YF, Dang H, Yue M, Liu X, Guo YX (2021). Patterns of diversity and community assembly change across local to regional scales: an evidence of deterministic assembly processes along resource availability gradient at temperate forest. Ecological Indicators, 132, 108261. DOI: 10.1016/j.ecolind.2021.108261. |
| [46] |
Yan GY, Fan CN, Zheng JQ, Liu GC, Yu JH, Guo ZL, Cao W, Wang LH, Wang WJ, Meng QF, Zhang JH, Li Y, Zheng JP, Cui XY, Wang XC, et al. (2024). Forest carbon stocks increase with higher dominance of ectomycorrhizal trees in high latitude forests. Nature Communications, 15, 5959. DOI: 10.1038/s41467-024-50423-9.
PMID |
| [47] |
Yang JR, Dai D, Chen JF, Wu X, Liu XL, Liu Y (2023). Insight into recent studies on the diversity of arbuscular mycorrhizal fungi in shaping plant community assembly and maintaining rare species. Chinese Journal of Plant Ecology, 47, 745-755.
DOI URL |
|
[杨佳绒, 戴冬, 陈俊芳, 吴宪, 刘啸林, 刘宇 (2023). 丛枝菌根真菌多样性对植物群落构建和稀有种维持的研究进展. 植物生态学报, 47, 745-755.]
DOI |
|
| [48] |
Yi H, Eisenhauer N, Austen JC, Rebollo R, Ray T, Bönisch E, von Oheimb G, Fichtner A, Schuldt A, Patoine G, Ferlian O (2024). Tree diversity and mycorrhizal type co-determine multitrophic ecosystem functions. Journal of Ecology, 112, 528-546.
DOI URL |
| [49] | Zhang GL, Jia J, Zhao QQ, Wang W, Wang DW, Bai JH (2023a). Seasonality and assembly of soil microbial communities in coastal salt marshes invaded by a perennial grass. Journal of Environmental Management, 331, 117247. DOI: 10.1016/j.jenvman.2023.117247. |
| [50] |
Zhang L, Liu X, Sun Z, Bu W, Bongers FJ, Song X, Yang J, Sun Z, Li Y, Li S, Cao M, Ma K, Swenson NG (2023b). Functional trait space and redundancy of plant communities decrease toward cold temperature at high altitudes in Southwest China. Science China Life Sciences, 66, 376-384.
DOI |
| [51] | Zhao L, Wang S, Shen R, Gong Y, Wang C, Hong P, Reuman DC (2022a). Biodiversity stabilizes plant communities through statistical-averaging effects rather than compensatory dynamics. Nature Communications, 13, 7804. DOI: 10.1038/s41467-022-35514-9. |
| [52] | Zhao LJ, Xiang WH, Li JX, Liu WQ, Hu YT, Wu HL, Zhang YL, Cheng X, Wang WJ, Wang WT, Ouyang S (2022b). “Realistic strategies” and neutral processes drive the community assembly based on leaf functional traits in a subtropical evergreen broad-leaved forest. Ecology and Evolution, 12, e9323. DOI: 10.1002/ece3.9323. |
| [53] |
Zhong Y, Chu C, Myers JA, Gilbert GS, Lutz JA, Stillhard J, Zhu K, Thompson J, Baltzer JL, He F, LaManna JA, Davies SJ, Aderson-Teixeira KJ, Burslem DFRP, Alonso A, et al. (2021). Arbuscular mycorrhizal trees influence the latitudinal beta-diversity gradient of tree communities in forests worldwide. Nature Communications, 12, 3137. DOI: 10.1038/s41467-021-23236-3.
PMID |
| [54] | Zhu LY, Luan L, Chen Y, Wang XY, Zhou SG, Zou WX, Han XR, Duan YH, Zhu B, Li Y, Liu WZ, Zhou JZ, Zhang JB, Jiang YJ, Sun B (2024). Community assembly of organisms regulates soil microbial functional potential through dual mechanisms. Global Change Biology, 30, e17160. DOI: 10.1111/gcb.17160. |
| [1] | 段建林, 孟晟, 陈仁利, 熊林峰, 卢春洋, 席念勋. 全球变化因子多样性对菌根植物性状的影响[J]. , 2026, 50(预发表): 0-. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
Copyright © 2026 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19
![]()