
植物生态学报 ›› 2018, Vol. 42 ›› Issue (7): 774-784.DOI: 10.17521/cjpe.2018.0104
孙慧敏1,姜姜1,*( ),崔莉娜1,张水锋2,张金池1
),崔莉娜1,张水锋2,张金池1
出版日期:2018-07-20
发布日期:2018-09-26
通讯作者:
姜姜
基金资助:
SUN Hui-Min1, JIANG Jiang1,*(), CUI Li-Na1, ZHANG Shui-Feng2, ZHANG Jin-Chi1
Online:2018-07-20
Published:2018-09-26
Contact:
Jiang JIANG
Supported by:摘要:
红树林对全球气候变化敏感, 近年来不少区域又受互花米草(Spartina alterniflora)入侵的影响, 土壤碳库组成发生显著变化, 然而鲜有从有机碳官能团特征角度出发的关于两群落的研究。为了解在红树林群落与互花米草群落下土壤碳库及其有机碳官能团的特征差异, 在福建省云霄县漳江口红树林自然保护区湿地内由内陆到海岸方向选取3条样带, 每条样带依次选取3个样地: 红树林群落(MC)、秋茄(Kandelia obovata)-互花米草过渡带(TC)和互花米草群落(SC), 每个样地选取3个呈品字形分布的采样点, 分5层采集0-100 cm土壤样品, 分析土壤中的总有机碳(TOC)、颗粒有机碳(POC)以及可溶性有机碳(DOC)特征, 并利用核磁共振波谱法测定表层0-15 cm与深层75-100 cm土壤总有机碳官能团特征, 以空间换时间法研究入侵前后土壤碳库变化特征。结果表明: (1)从MC群落到SC群落, 土壤有机碳库显著减小, 各样地总有机碳与颗粒有机碳含量表现为MC > TC > SC, 并随着土层深度增加而减少, DOC含量没有表现出明显的变化趋势。(2)各植被类型土壤有机碳以烷基碳与烷氧碳为主, 其次是芳香碳与羰基碳, N-烷氧碳与酚基碳含量最少, 其中表层0-15 cm土壤从红树林群落到互花米草群落, 烷基碳与烷氧碳含量呈现增加趋势但不显著, 芳香碳与酚基碳含量显著减少, 其余有机碳组分含量无显著差异。在深层75-100 cm随着植被类型的改变, 土壤有机碳组成结构均无显著差异。(3)在0-15 cm土层, 烷基碳/烷氧碳含量表现为: SC > MC > TC; 芳香度表现为SC最小, MC与TC无显著差异; 疏水碳/亲水碳无显著差异; 脂族碳/芳香碳表现为SC显著大于其他两种植被类型, MC与TC无显著差异。在75-100 cm土层, 各比值无显著差异。综上所述, 红树林群落碳储量显著高于互花米草群落, 受植被的影响, 互花米草群落表层土壤有机碳分解程度显著高于红树林群落, 而红树林群落的土壤有机碳分子结构要比互花米草群落更复杂, 以维持其土壤有机碳的稳定性。因此, 互花米草入侵红树林后可能会加快有机碳的分解, 最终稳定在相对简单的分子结构, 降低土壤碳储量。
孙慧敏, 姜姜, 崔莉娜, 张水锋, 张金池. 互花米草入侵对漳江口红树林湿地土壤有机碳官能团特征的影响. 植物生态学报, 2018, 42(7): 774-784. DOI: 10.17521/cjpe.2018.0104
SUN Hui-Min, JIANG Jiang, CUI Li-Na, ZHANG Shui-Feng, ZHANG Jin-Chi. Effects of Spartina alterniflora invasion on soil organic carbon composition of mangrove wetland in Zhangjiang River Estuary. Chinese Journal of Plant Ecology, 2018, 42(7): 774-784. DOI: 10.17521/cjpe.2018.0104

图1 研究区域位置及采样点分布。MC, 红树林群落; SC, 互花米草群落; TC, 秋茄-互花米草过渡带。
Fig. 1 The location of study area and sampling sites. MC, mangrove community; SC, Spartina alterniflora community; TC, Kandelia obovata-S. alterniflora transitional community.
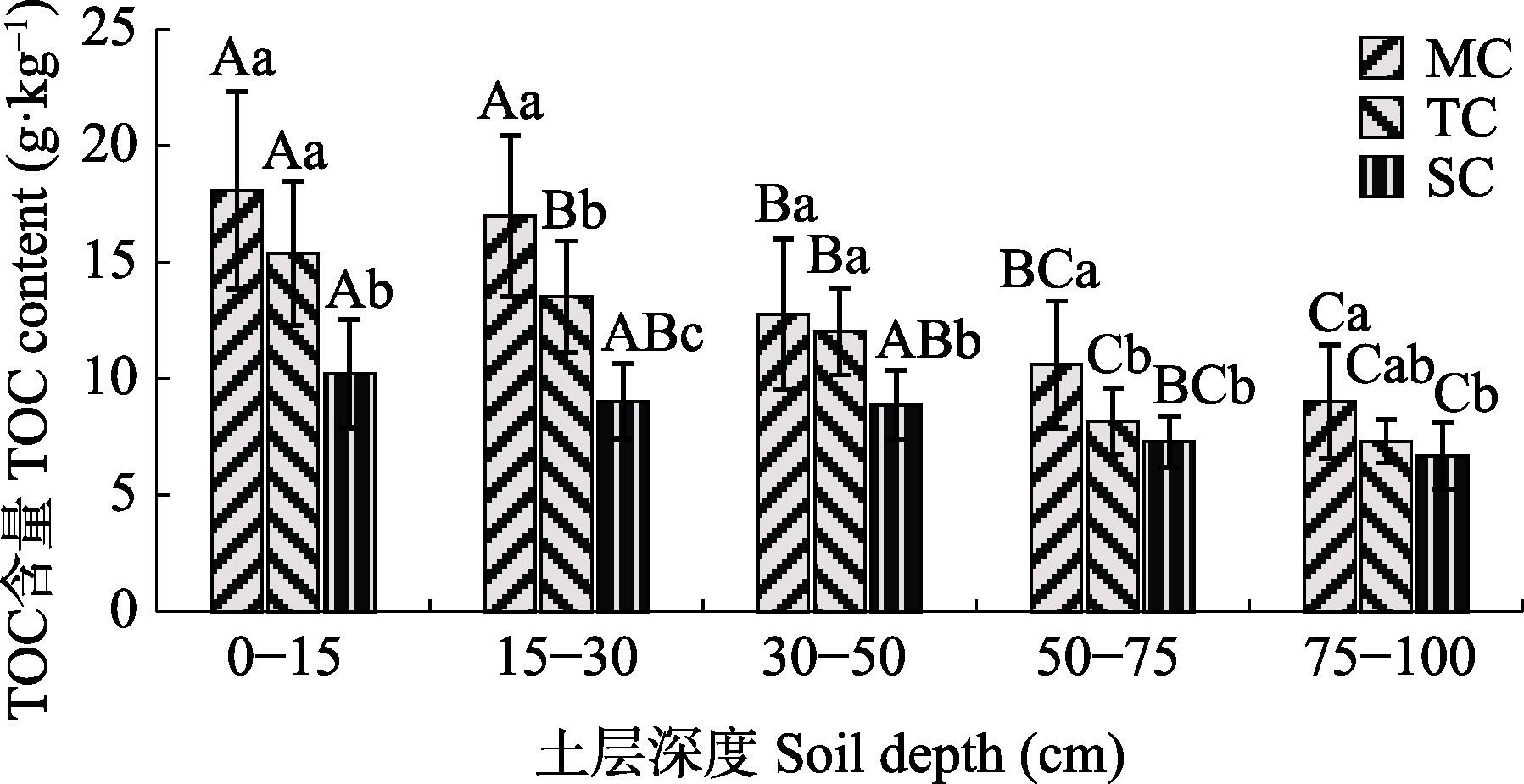
图2 各土层土壤有机碳(TOC)含量(平均值±标准偏差)。不同大写字母表示同一植被类型不同土层间差异显著, 不同小写字母表示同一土层不同植被类型间显著差异(p < 0.05)。MC, 红树林群落; SC, 互花米草群落; TC, 秋茄-互花米草过渡带。
Fig. 2 Total organic carbon (TOC) content at different soil depths (mean ± SD). Different capital letters indicate significant differences in different soil layers of the same vegetation type (p < 0.05); and different lowercase letters indicate significant differences in different vegetation types of the same soil layer (p < 0.05). MC, mangrove community; SC, Spartina alterniflora community; TC, Kandelia obovata-S. alterniflora transitional community.
| 效应 Source of effects | 平方和 Sum of squares (SS) | 自由度 d.f. | 均方 Mean square (MS) | F | p |
|---|---|---|---|---|---|
| 土层深度 Soil depth | 853.659 | 4 | 213.415 | 34.747 | <0.001 |
| 植被类型 Vegetation type | 521.694 | 2 | 260.847 | 42.470 | <0.001 |
| 土层深度×植被类型 Soil depth × Vegetation type | 136.912 | 8 | 17.114 | 2.786 | 0.008 |
表1 土壤有机碳含量的土层深度与植被类型的双因子方差分析
Table 1 Two-way analysis of variance for total organic carbon content in vegetation types and soil depth
| 效应 Source of effects | 平方和 Sum of squares (SS) | 自由度 d.f. | 均方 Mean square (MS) | F | p |
|---|---|---|---|---|---|
| 土层深度 Soil depth | 853.659 | 4 | 213.415 | 34.747 | <0.001 |
| 植被类型 Vegetation type | 521.694 | 2 | 260.847 | 42.470 | <0.001 |
| 土层深度×植被类型 Soil depth × Vegetation type | 136.912 | 8 | 17.114 | 2.786 | 0.008 |

图3 各土层土壤颗粒有机碳(POC)含量(平均值±标准偏差)。不同大写字母表示同一植被类型不同土层间差异显著, 不同小写字母表示同一土层不同植被类型间显著差异(p < 0.05)。MC, 红树林群落; SC, 互花米草群落; TC, 秋茄-互花米草过渡带。
Fig. 3 Particulate organic carbon (POC) content at different soil depths (mean ± SD). Different capital letters indicate significant differences in different soil layers of the same vegetation type (p < 0.05); and different lowercase letters indicate significant differences in different vegetation types of the same soil layer (p < 0.05). MC, mangrove community; SC, Spartina alterniflora community; TC, Kandelia obovata-S. alterniflora transitional community.
| 效应 Source of effects | 平方和 Sum of squares (SS) | 自由度 d.f. | 均方 Mean square (MS) | F | p |
|---|---|---|---|---|---|
| 土层深度 Soil depth | 961.322 | 4 | 240.330 | 525.104 | <0.001 |
| 植被类型 Vegetation type | 440.351 | 3 | 146.784 | 320.711 | <0.001 |
| 土层深度×植被类型 Soil depth × Vegetation type | 118.343 | 8 | 14.793 | 32.321 | <0.001 |
表2 颗粒有机碳含量的土层深度与植被类型的双因子方差分析
Table 2 Two-way analysis of variance for particulate organic carbon content in vegetation types and soil depth
| 效应 Source of effects | 平方和 Sum of squares (SS) | 自由度 d.f. | 均方 Mean square (MS) | F | p |
|---|---|---|---|---|---|
| 土层深度 Soil depth | 961.322 | 4 | 240.330 | 525.104 | <0.001 |
| 植被类型 Vegetation type | 440.351 | 3 | 146.784 | 320.711 | <0.001 |
| 土层深度×植被类型 Soil depth × Vegetation type | 118.343 | 8 | 14.793 | 32.321 | <0.001 |

图4 各土层土壤溶解有机碳(DOC)含量(平均值±标准偏差)。不同大写字母表示同一植被类型不同土层间差异显著, 不同小写字母表示同一土层不同植被类型间显著差异(p < 0.05)。MC, 红树林群落; SC, 互花米草群落; TC, 秋茄-互花米草过渡带。
Fig. 4 Dissolved organic carbon (DOC) content at different soil depths (mean ± SD). Different capital letters indicate significant differences in different soil layers of the same vegetation type (p < 0.05); and different lowercase letters indicate significant differences in different vegetation types of the same soil layer (p < 0.05). MC, mangrove community; SC, Spartina alterniflora community; TC, Kandelia obovata-S. alterniflora transitional community.
| 效应 Source of effects | 平方和 Sum of squares (SS) | 自由度 d.f. | 均方 Mean square (MS) | F | p |
|---|---|---|---|---|---|
| 土层深度 Soil depth | 19 929.215 | 4 | 4 982.304 | 13.063 | <0.001 |
| 植被类型 Vegetation type | 24 698.588 | 2 | 12 349.294 | 32.379 | <0.001 |
| 土层深度×植被类型 Soil depth × Vegetation type | 19 651.572 | 8 | 2 456.446 | 6.441 | <0.001 |
表3 溶解有机碳(DOC)含量的土层深度与植被类型的双因子方差分析
Table 3 Two-way analysis of variance for dissolved organic carbon content in forest types and soil depth
| 效应 Source of effects | 平方和 Sum of squares (SS) | 自由度 d.f. | 均方 Mean square (MS) | F | p |
|---|---|---|---|---|---|
| 土层深度 Soil depth | 19 929.215 | 4 | 4 982.304 | 13.063 | <0.001 |
| 植被类型 Vegetation type | 24 698.588 | 2 | 12 349.294 | 32.379 | <0.001 |
| 土层深度×植被类型 Soil depth × Vegetation type | 19 651.572 | 8 | 2 456.446 | 6.441 | <0.001 |

图5 两个土层3种植被类型土壤有机碳核磁共振波谱图。T1、T2、T3分别为互花米草群落、秋茄-互花米草过渡带和红树林群落样带。
Fig. 5 Nuclear magnetic resonance spectra of three vegetation types at different soil depths. T1, T2 and T3 are three transect lines of S. alterniflora community, transitional community and mangrove community, respectively.
| 土壤深度 Soil depth | 植被类型 Vegetation type | 烷基碳 Alkyl C | N-烷氧碳 N-alkyl C | 烷氧碳 O-alkyl C | 缩醛碳 di-O-alkyl C | 芳香碳 Aromatic C | 酚基碳 Phenolic C | 羰基碳 Carbonyl C | 烷基碳/ 烷氧碳 A/O-A | 芳香度 Aromaticity | 疏水碳/亲水碳 Hydrophobic/ hydrophilic C | 脂族碳/芳香碳 Aliphatic/ aromatic C |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0-15 cm | MC | 0.199 6ab | 0.072 9a | 0.216 5a | 0.108 0ab | 0.172 3a | 0.091 1a | 0.139 7a | 0.503 0ab | 0.3065 a | 0.862 9a | 2.269 7a |
| 0-15 cm | TC | 0.157 2a | 0.080 0a | 0.260 0a | 0.118 5a | 0.185 8b | 0.084 1a | 0.114 3a | 0.343 6a | 0.304 8a | 0.746 0a | 2.281 8a |
| 0-15 cm | SC | 0.240 3b | 0.083 7a | 0.234 9a | 0.095 8b | 0.157 5c | 0.065 6b | 0.122 3a | 0.602 6b | 0.254 7b | 0.867 3a | 2.941 0b |
| 75-100 cm | MC | 0.234 6a | 0.076 5a | 0.228 9a | 0.085 3a | 0.156 9a | 0.068 0a | 0.149 8a | 0.628 2a | 0.264 1a | 0.855 8a | 2.892 9a |
| 75-100 cm | TC | 0.270 4a | 0.088 7a | 0.213 7a | 0.077 9a | 0.150 0a | 0.061 3a | 0.138 1a | 0.710 4a | 0.246 0a | 0.930 2a | 3.161 2a |
| 75-100 cm | SC | 0.216 9a | 0.078 3a | 0.220 2a | 0.082 2a | 0.170 9a | 0.071 4a | 0.160 0a | 0.584 7a | 0.288 6a | 0.854 6a | 2.502 5a |
表4 不同植被类型土壤有机碳官能团比例
Table 4 The ratios of soil organic carbon functional groups for different vegetation types
| 土壤深度 Soil depth | 植被类型 Vegetation type | 烷基碳 Alkyl C | N-烷氧碳 N-alkyl C | 烷氧碳 O-alkyl C | 缩醛碳 di-O-alkyl C | 芳香碳 Aromatic C | 酚基碳 Phenolic C | 羰基碳 Carbonyl C | 烷基碳/ 烷氧碳 A/O-A | 芳香度 Aromaticity | 疏水碳/亲水碳 Hydrophobic/ hydrophilic C | 脂族碳/芳香碳 Aliphatic/ aromatic C |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0-15 cm | MC | 0.199 6ab | 0.072 9a | 0.216 5a | 0.108 0ab | 0.172 3a | 0.091 1a | 0.139 7a | 0.503 0ab | 0.3065 a | 0.862 9a | 2.269 7a |
| 0-15 cm | TC | 0.157 2a | 0.080 0a | 0.260 0a | 0.118 5a | 0.185 8b | 0.084 1a | 0.114 3a | 0.343 6a | 0.304 8a | 0.746 0a | 2.281 8a |
| 0-15 cm | SC | 0.240 3b | 0.083 7a | 0.234 9a | 0.095 8b | 0.157 5c | 0.065 6b | 0.122 3a | 0.602 6b | 0.254 7b | 0.867 3a | 2.941 0b |
| 75-100 cm | MC | 0.234 6a | 0.076 5a | 0.228 9a | 0.085 3a | 0.156 9a | 0.068 0a | 0.149 8a | 0.628 2a | 0.264 1a | 0.855 8a | 2.892 9a |
| 75-100 cm | TC | 0.270 4a | 0.088 7a | 0.213 7a | 0.077 9a | 0.150 0a | 0.061 3a | 0.138 1a | 0.710 4a | 0.246 0a | 0.930 2a | 3.161 2a |
| 75-100 cm | SC | 0.216 9a | 0.078 3a | 0.220 2a | 0.082 2a | 0.170 9a | 0.071 4a | 0.160 0a | 0.584 7a | 0.288 6a | 0.854 6a | 2.502 5a |
| [1] |
Albaladejo J, Ortiz R, Garcia-Franco N, Navarro AR, Almagro M, Pintado JG ( 2013). Land use and climate change impacts on soil organic carbon stocks in semi-arid Spain. Journal of Soils and Sediments, 13, 265-277.
DOI URL |
| [2] |
Alongi DM ( 2014). Carbon cycling and storage in mangrove forests. Annual Review of Marine Science, 6, 195-219.
DOI URL PMID |
| [3] |
Bai J, Yan JY, He DJ, Cai JB, Wang R, You WB, Xiao SH, Hou DL, Li WW ( 2017). Effects of Spartina alterniflora invasion in eastern Fujian coastal wetland on the physicochemical properties and enzyme activities of mangrove soil. Journal of Beijing Forestry University, 39(1), 70-77.
DOI URL |
|
[ 白静, 严锦钰, 何东进, 蔡金标, 王韧, 游巍斌, 肖石红, 侯栋梁, 李威威 ( 2017). 互花米草入侵对闽东滨海湿地红树林土壤理化性质和酶活性的影响. 北京林业大学学报, 39(1), 70-77.]
DOI URL |
|
| [4] |
Biederbeck VO, Janzen HH, Campbell CA, Zentner RP ( 1994). Labile soil organic matter as influenced by cropping practices in an arid environment. Soil Biology & Biochemistry, 26, 1647-1656.
DOI URL |
| [5] |
Bouillon S, Borges AV, Casta?eda-Moya E, Diele K, Dittmar T, Duke NC ( 2008). Mangrove production and carbon sinks: A revision of global budget estimates. Global Biogeochemical Cycles, 22(2), 1-12.
DOI URL |
| [6] |
Cai WJ ( 2011). Estuarine and coastal ocean carbon paradox: CO2 sinks or sites of terrestrial carbon incineration? Annual Review of Marine Science, 3(3), 123-145.
DOI URL PMID |
| [7] |
Chen C ( 2004). Soil carbon pools in adjacent natural and plantation forests of subtropical Australia. Soil Science Society of America Journal, 68, 282-291.
DOI URL |
| [8] | Chen GX, Gao DZ, Chen G, Zeng CS, Wang WQ ( 2017). Effects of Spartina alterniflora invasion on soil carbon fractions in mangrove wetlands of China. Journal of Soil and Water Conservation, 31(6), 249-256. |
| [ 陈桂香, 高灯州, 陈刚, 曾从盛, 王维奇 ( 2017). 互花米草入侵对我国红树林湿地土壤碳组分的影响. 水土保持学报, 31(6), 249-256.] | |
| [9] | Chen ZJ, Han SJ, Zhang JH ( 2016). Effects of land use change on soil organic carbon fractions in mangrove wetland of Zhangjiangkou. Chinese Journal of Ecology, 35, 2379-2385. |
| [ 陈志杰, 韩士杰, 张军辉 ( 2016). 土地利用变化对漳江口红树林土壤有机碳组分的影响. 生态学杂志, 35, 2379-2385.] | |
| [10] | Coleman DC, Callaham MA, Crossley Jr DA ( 2017). Fundamentals of soil ecology. The Quarterly Review of Biology, 161, 321. |
| [11] |
Dou S, Zhang JJ, Li K ( 2008). Effect of organic matter applications on 13C-NMR spectra of humic acids of soil. European Journal of Soil Science, 59, 532-539.
DOI URL |
| [12] |
Doughty CL, Langley JA, Walker WS, Feller IC, Schaub R, Chapman SK ( 2016). Mangrove range expansion rapidly increases coastal wetland carbon storage. Estuaries and Coasts, 39, 1-12.
DOI URL |
| [13] |
Ehrenfeld JG ( 2010). Ecosystem consequences of biological invasions. Annual Review of Ecology, Evolution, and Systematics, 41, 59-80.
DOI URL |
| [14] |
Fontaine S, Barot S, Barré P, Bdioui N, Mary B, Rumpel C ( 2007). Stability of organic carbon in deep soil layers controlled by fresh carbon supply. Nature, 450, 277-280.
DOI URL PMID |
| [15] |
Gregorich EG, Voroney RP, Kachanoski RG ( 1991). Turnover of carbon through the microbial biomass in soils with different texture. Soil Biology & Biochemistry, 23, 799-805.
DOI URL |
| [16] | Hang ZQ ( 2013). Soil Organic Carbon Composition, Source and Structural Characteristics of Spartina alterniflora. Master degree dissertation, Nanjing Normal University, Nanjing. |
| [ 杭子清 ( 2013). 互花米草(Spartina alterniflora)盐沼土壤有机碳组分、来源及结构特征研究. 硕士学位论文, 南京师范大学, 南京.] | |
| [17] | Hang ZQ, Wang GX, Liu JE, Wang G, Wang H ( 2014). Characterization of soil organic carbon fractions at Spartina alterniflora saltmarsh in North Jiangsu. Acta Ecologica Sinica, 34, 4175-4182. |
| [ 杭子清, 王国祥, 刘金娥, 王刚, 王会 ( 2014). 互花米草盐沼土壤有机碳库组分及结构特征. 生态学报, 34, 4175-4182.] | |
| [18] |
Kelleway JJ, Saintilan N, Macreadie PI, Skilbeck CG, Zawadzki A, Ralph PJ ( 2015). Seventy years of continuous encroachment substantially increases “blue carbon” capacity as mangroves replace intertidal salt marshes. Global Change Biology, 22, 1097-1109.
DOI URL PMID |
| [19] | Li GD, Liu GQ, Zhuang SY, Gui RY ( 2010). Changes of organic matter in soils planted lei bamboo with different years. Chinese Journal of Soil Science, 41, 845-849. |
| [ 李国栋, 刘国群, 庄舜尧, 桂仁意 ( 2010). 不同种植年限下雷竹林土壤的有机质转化. 土壤通报, 41, 845-849.] | |
| [20] |
Li SF, Yu YC, He S ( 2002). Summary of research on dissolved organic carbon (DOC). Soil and Environmental Sciences, 11, 422-429.
DOI URL |
|
[ 李淑芬, 俞元春, 何晟 ( 2002). 土壤溶解有机碳的研究进展. 土壤与环境, 11, 422-429.]
DOI URL |
|
| [21] | Li T, Zhao SW, Zhang Y, Ma S, Li XX ( 2011). Effect of revegetation on functional groups of soil organic carbon on the Loess Plateau. Acta Ecologica Sinica, 31, 5199-5206. |
| [ 李婷, 赵世伟, 张扬, 马帅, 李晓晓 ( 2011). 黄土区次生植被恢复对土壤有机碳官能团的影响. 生态学报, 31, 5199-5206.] | |
| [22] |
Li Z, Zhao B, Wang Q, Cao X, Zhang J ( 2015). Differences in chemical composition of soil organic carbon resulting from long-term fertilization strategies. PLOS ONE, 10, e0124359. DOI: 10.1371/journal.pone.0124359.
DOI URL PMID |
| [23] | Liao BW, Zhang QM ( 2014). Area, distribution and species composition of mangroves in China. Wetland Science, 12, 435-440. |
| [ 廖宝文, 张乔民 ( 2014). 中国红树林的分布、面积和树种组成. 湿地科学, 12, 435-440.] | |
| [24] |
Linn DM, Doran JW ( 1984). Aerobic and anaerobic microbial populations in no-till and plowed soils. Soil Science Society of America Journal, 48, 794-799.
DOI URL |
| [25] |
Liu MQ, Hu F, Chen XY ( 2007). A review on mechanisms of soil organic carbon stabilization. Acta Ecologica Sinica, 27, 2642-2650.
DOI URL |
|
[ 刘满强, 胡锋, 陈小云 ( 2007). 土壤有机碳稳定机制研究进展. 生态学报, 27, 2642-2650.]
DOI URL |
|
| [26] |
Mathers NJ, Xu Z ( 2003). Solid-state 13C NMR spectroscopy: Characterization of soil organic matter under two contrasting residue management regimes in a 2-year-old pine plantation of subtropical Australia . Geoderma, 114, 19-31.
DOI URL |
| [27] |
Mathers NJ, Xu Z, Bernersprice SJ, Perera MCS, Saffigna PG ( 2002). Hydrofluoric acid pre-treatment for improving 13C CPMAS NMR spectral quality of forest soils in south-east Queensland, Australia . Soil Research, 40, 665-674.
DOI URL |
| [28] |
Mcleod E, Chmura GL, Bouillon S, Salm R, Bj?rk M, Duarte CM ( 2011). A blueprint for blue carbon: Toward an improved understanding of the role of vegetated coastal habitats in sequestering CO2. Frontiers in Ecology and the Environment, 9, 552-560.
DOI URL |
| [29] |
Mikutta R, Kleber M, Torn MS, Jahn R ( 2006). Stabilization of soil organic matter: Association with minerals or chemical recalcitrance? Biogeochemistry, 77, 25-56.
DOI URL |
| [30] |
Ouyang X, Lee SY, Connolly RM ( 2016). Structural equation modelling reveals factors regulating surface sediment organic carbon content and CO2 efflux in a subtropical mangrove. Science of the Total Environment, 578, 513-522.
DOI URL PMID |
| [31] |
Rumpel C, K?gel-Knabner I ( 2011). Deep soil organic matter— A key but poorly understood component of terrestrial C cycle. Plant and Soil, 338, 143-158.
DOI URL |
| [32] |
Schmidt MW, Torn MS, Abiven S, Dittmar T, Guggenberger G, Janssens IA ( 2011). Persistence of soil organic matter as an ecosystem property. Nature, 478, 49-56.
DOI URL |
| [33] |
Sollins P, Homann P, Caldwell BA ( 1996). Stabilization and destabilization of soil organic matter: Mechanisms and controls. Geoderma, 74(1-2), 65-105.
DOI URL |
| [34] |
Spaccini R, Mbagwu JSC, Conte P, Piccolo A ( 2006). Changes of humic substances characteristics from forested to cultivated soils in Ethiopia. Geoderma, 132, 9-19.
DOI URL |
| [35] |
Tiessen H, Stewart JWB ( 1983). Particle-size fractions and their use in studies of soil organic Matter: II. Cultivation effects on organic matter composition in size fractions. Soil Science Society of America Journal, 47, 509-514.
DOI URL |
| [36] |
Ussiri DAN, Johnson CE ( 2003). Characterization of organic matter in a northern hardwood forest soil by 13C NMR spectroscopy and chemical methods . Geoderma, 111, 123-149.
DOI URL |
| [37] |
Wang G, Yang WB, Wang GX, Liu JE, Hang ZQ ( 2013). The effects of Spartina alterniflora seaward invasion on soil organic carbon fractions, sources and distribution. Acta Ecologica Sinica, 33, 2474-2483.
DOI URL |
|
[ 王刚, 杨文斌, 王国祥, 刘金娥, 杭子清 ( 2013). 互花米草海向入侵对土壤有机碳组分、来源和分布的影响. 生态学报, 33, 2474-2483.]
DOI URL |
|
| [38] |
Wang JM, Ouyang J, Shang Q, Deng ZW ( 2008). Application of the NMR techniques in studies on organic matters in soil. Chinese Journal of Magnetic Resonance, 25, 287-295.
DOI URL |
|
[ 王俊美, 欧阳捷, 尚倩, 邓志威 ( 2008). 土壤有机质研究中的核磁共振技术. 波谱学杂志, 25, 287-295.]
DOI URL |
|
| [39] |
Wang S, Mei H, Shao X, Mickler RA, Li K, Ji J ( 2004). Vertical distribution of soil organic carbon in China. Environmental Management, 33, S200-S209.
DOI URL |
| [40] | Wei D, Dai WH, Tang J ( 2011). Study of soils dissolved organic carbon in different landuse. Chinese Agricultural Science Bulletin, 27(18), 121-124. |
| [ 卫东, 戴万宏, 汤佳 ( 2011). 不同利用方式下土壤溶解性有机碳含量研究. 中国农学通报, 27(18), 121-124.] | |
| [41] | Wu YP ( 2015). In situ Decomposition of Organic Carbon in Litter of Spartina alterniflora. Master degree dissertation, Nanjing Normal University,Nanjing. |
| [ 吴亚萍 ( 2015). 互花米草(Spartina alterniflora)凋落物有机碳原位分解动态. 硕士学位论文, 南京师范大学, 南京.] | |
| [42] |
Wynn JG, Bird MI, Vellen L, Grand-Clement E, Carter J, Berry SL ( 2006). Continental-scale measurement of the soil organic carbon pool with climatic, edaphic, and biotic controls. Global Biogeochemical Cycles, 20, GB1007. DOI: 10.1029/2005GB002576.
DOI URL |
| [43] |
Xue JF, Gao YM, Wang JK, Fu SF, Zhu FC ( 2007). Microbial biomass carbon and nitrogen as an indicator for evaluation of soil fertility. Chinese Journal of Soil Science, 38, 247-250.
DOI URL |
|
[ 薛菁芳, 高艳梅, 汪景宽, 付时丰, 祝凤春 ( 2007). 土壤微生物量碳氮作为土壤肥力指标的探讨. 土壤通报, 38, 247-250.]
DOI URL |
|
| [44] | Yang W, Zhao H, Chen X, Yin S, Cheng X, An S ( 2013). Consequences of short-term C4 plant Spartina alterniflora, invasions for soil organic carbon dynamics in a coastal wetland of eastern China. Ecological Engineering, 61(12), 50-57. |
| [45] | Yang WQ, Deng RJ, Zhang J ( 2007). Forest litter decomposition and its responses to global climate change. Chinese Journal of Applied Ecology, 18, 2889-2895. |
| [ 杨万勤, 邓仁菊, 张健 ( 2007). 森林凋落物分解及其对全球气候变化的响应. 应用生态学报, 18, 2889-2895.] | |
| [46] | Zhang L, Guo ZH, Li ZY ( 2013). Carbon storage and carbon sink of mangrove wetland: Research progress. Chinese Journal of Applied Ecology, 24, 1153-1159. |
| [ 张莉, 郭志华, 李志勇 ( 2013). 红树林湿地碳储量及碳汇研究进展. 应用生态学报, 24, 1153-1159.] | |
| [47] |
Zhang XH, Li DY, Pan GX, Li LQ, Lin F, Xu XW ( 2008). Conservation of wetland soil C stock and climate change of China. Advances in Climate Change Research, 4, 202-208.
DOI URL |
|
[ 张旭辉, 李典友, 潘根兴, 李恋卿, 林凡, 许信旺 ( 2008). 中国湿地土壤碳库保护与气候变化问题. 气候变化研究进展, 4, 202-208.]
DOI URL |
|
| [48] | Zhang YH, Zhang FC, Zhou XD, Xie XJ, Wang XW, Li Q ( 2011). Effects of plant invasion along a Spartina alterniflora chronosequence on organic carbon dynamics in coastal wetland in north Jiangsu. China Environmental Science, 31, 271-276. |
| [ 张耀鸿, 张富存, 周晓冬, 谢晓金, 王小巍, 李强 ( 2011). 互花米草对苏北滨海湿地表土有机碳更新的影响. 中国环境科学, 31, 271-276.] | |
| [49] | Zhou CH, Mao TY, Xu X, Fang CM, Luo YM, Li B ( 2016). Preliminary analysis of C sequestration potential of blue carbon ecosystems on Chinese coastal zone. Scientia Sinica: Vitae, 46, 475-486. |
| [ 周晨昊, 毛覃愉, 徐晓, 方长明, 骆永明, 李博 ( 2016). 中国海岸带蓝碳生态系统碳汇潜力的初步分析. 中国科学: 生命科学, 46, 475-486.] |
| [1] | 盘远方, 潘良浩, 邱思婷, 邱广龙, 苏治南, 史小芳, 范航清. 中国沿海红树林树高变异与环境适应机制[J]. 植物生态学报, 2024, 48(4): 483-495. |
| [2] | 史欢欢, 雪穷, 于振林, 汪承焕. 密度、物种比例对盐沼植物种子萌发阶段种内、种间相互作用的影响[J]. 植物生态学报, 2023, 47(1): 77-87. |
| [3] | 陈权, 马克明. 互花米草入侵对红树林湿地沉积物重金属累积的效应与潜在机制[J]. 植物生态学报, 2017, 41(4): 409-417. |
| [4] | 陈权, 马克明. 红树林生物入侵研究概况与趋势[J]. 植物生态学报, 2015, 39(3): 283-299. |
| [5] | 王丹, 张荣, 熊俊, 郭海强, 赵斌. 互花米草入侵对滨海湿地土壤碳库的贡献——基于稳定同位素的研究[J]. 植物生态学报, 2015, 39(10): 941-949. |
| [6] | 胡楚琦, 刘金珂, 王天弘, 王文琳, 卢山, 周长芳. 三种盐胁迫对互花米草和芦苇光合作用的影响[J]. 植物生态学报, 2015, 39(1): 92-103. |
| [7] | 赵聪蛟, 邓自发, 周长芳, 关保华, 安树青, 陈琳, 陆霞梅. 氮水平和竞争对互花米草与芦苇叶特征的影响[J]. 植物生态学报, 2008, 32(2): 392-401. |
| [8] | 陈琳, 邓自发, 安树青, 赵聪蛟, 周长芳, 智颖飙. 淡咸水轮换浇灌抑制互花米草的克隆生长和繁殖[J]. 植物生态学报, 2007, 31(4): 645-651. |
| [9] | 黄华梅, 张利权. 上海九段沙互花米草种群动态遥感研究[J]. 植物生态学报, 2007, 31(1): 75-82. |
| [10] | 何斌, 温远光, 梁宏温, 李志先, 刘世荣. 英罗港红树植物群落不同演替阶段植物元素分布及其与土壤肥力的关系[J]. 植物生态学报, 2002, 26(5): 518-524. |
| [11] | 叶勇, 谭凤仪, 卢昌义. 土壤结构与光照水平对秋茄某些生长和生理参数的影响[J]. 植物生态学报, 2001, 25(1): 42-49. |
| [12] | 卢昌义, 叶勇, 黄玉山, 谭凤仪. 海南岛东寨港红树林群落甲烷通量研究[J]. 植物生态学报, 2000, 24(1): 87-90. |
| [13] | 林鹏, 王文卿. 盐胁迫下红树植物秋茄(Kandelia candel)热值变化的研究[J]. 植物生态学报, 1999, 23(5): 466-470. |
| [14] | 杨盛昌, 林鹏. 潮滩红树植物抗低温适应的生态学研究[J]. 植物生态学报, 1998, 22(1): 60-67. |
| [15] | 梁士楚. 广西英罗湾红树植物群落的研究[J]. 植物生态学报, 1996, 20(4): 310-321. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
Copyright © 2022 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19