
植物生态学报 ›› 2016, Vol. 40 ›› Issue (3): 200-211.DOI: 10.17521/cjpe.2015.0161
所属专题: 生物多样性
韩杰1, 应凌霄1, 李贵祥2, 沈泽昊1,*( )
)
收稿日期:2015-05-07
修回日期:2016-12-27
出版日期:2016-03-10
发布日期:2016-03-25
通讯作者:
沈泽昊
基金资助:
Jie HAN1, Ling-Xiao YING1, Gui-Xiang LI2, Ze-Hao SHEN1,*()
Received:2015-05-07
Revised:2016-12-27
Online:2016-03-10
Published:2016-03-25
Contact:
Ze-Hao SHEN
摘要:
云南松(Pinus yunnanensis)林是我国西南地区最为广布的针叶林。火烧是云南松林常见的一种自然(或人为)干扰, 但对云南松林火后更新群落的物种多样性特征很少研究。该研究以云南禄丰县勤丰镇2013年“4.13”火灾后云南松混交林火烧迹地更新早期群落草本层为研究对象, 设置了47条1000 m2样带进行群落和环境因子调查, 探讨火烧后更新早期群落草本层的物种构成与丰富度的空间格局及其影响因素。结果表明: 火后更新草本层按物种构成可分为4个群落类型; 草本层物种丰富度受地形和火前群落冠层的影响显著, 与立木生物量和地形坡度正相关, 而与坡位和海拔负相关, 但与火烧强度没有显著的相关性。样带内草本群落β多样性与火前立木胸面积及火烧强度显著正相关, 而与海拔和坡位负相关。总体上, 生境地形特征、火前冠层和火烧强度构成了火烧后早期群落草本层物种构成与丰富度变化的主要控制因子。此外, 一年生和多年生草本物种的更新格局及其对环境的响应存在显著差异。
韩杰, 应凌霄, 李贵祥, 沈泽昊. 云南松混交林火烧迹地更新早期草本层物种多样性的空间格局. 植物生态学报, 2016, 40(3): 200-211. DOI: 10.17521/cjpe.2015.0161
Jie HAN, Ling-Xiao YING, Gui-Xiang LI, Ze-Hao SHEN. Spatial patterns of species diversity in the herb layer of early post-fire regeneration in mixed Pinus yunnanensis forests. Chinese Journal of Plant Ecology, 2016, 40(3): 200-211. DOI: 10.17521/cjpe.2015.0161
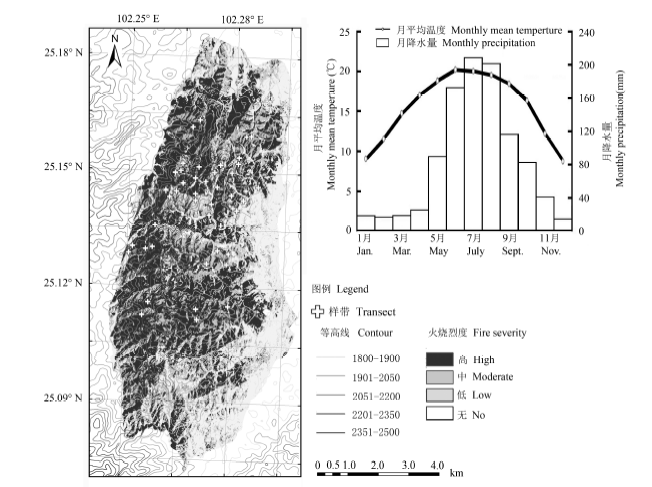
图1 研究区域的地理区位、气候、地貌特征和火烧面积(2013年)及其调查样点分布。
Fig. 1 The geographical location, climate, geomorphic feature of the burnt area (in 2013) and the distribution of sampling sites.
| 类型 Type | 变量 Variable | 定义及范围 Definition & value range |
|---|---|---|
| 火烧烈度 Fire severity | 火烧比例 Burn ratio | 0-1 |
| 火前植被 Pre-fire vegetation | ln(胸面积和) ln(Sum of basal area) | 样方中死树胸面积和的自然对数值 Natural logarithm of sum of basal area of dead trees in sample plot |
| 地形 Topography | 海拔 Elevation (m) | 1 928-2 331 m |
| 坡度 Slope gradient (°) | 5°-52° | |
| 坡向 Slope aspect (°) | 0°(N) -180°(S) | |
| 坡位 Slope position | 2-下坡位; 3-中坡位; 4-上坡位; 5-山顶 2-lower slope; 3-midslope; 4-upper slope; 5-top | |
| 坡形 Slope shape | 1-凹; 2-平; 3-凸 1-concave; 2-flat; 3-convex |
表1 火后植被与生境调查的变量
Table 1 Variables of post-fire vegetation and habitat investigations
| 类型 Type | 变量 Variable | 定义及范围 Definition & value range |
|---|---|---|
| 火烧烈度 Fire severity | 火烧比例 Burn ratio | 0-1 |
| 火前植被 Pre-fire vegetation | ln(胸面积和) ln(Sum of basal area) | 样方中死树胸面积和的自然对数值 Natural logarithm of sum of basal area of dead trees in sample plot |
| 地形 Topography | 海拔 Elevation (m) | 1 928-2 331 m |
| 坡度 Slope gradient (°) | 5°-52° | |
| 坡向 Slope aspect (°) | 0°(N) -180°(S) | |
| 坡位 Slope position | 2-下坡位; 3-中坡位; 4-上坡位; 5-山顶 2-lower slope; 3-midslope; 4-upper slope; 5-top | |
| 坡形 Slope shape | 1-凹; 2-平; 3-凸 1-concave; 2-flat; 3-convex |
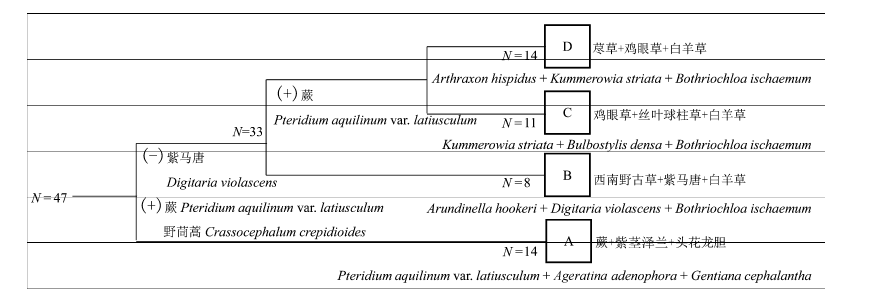
图2 火后更新草本层的TWINSPAN群落分类。N代表样方个数。A、B、C、D代表4种火后草本更新群落类型。A代表蕨+紫茎泽兰+头花龙胆群落; B代表西南野古草+紫马唐+白羊草群落; C代表鸡眼草+丝叶球柱草+白羊草群落; D代表荩草+鸡眼草+白羊草群落。(+)表示与分类聚组正相关; (-)表示与分类聚组负相关。
Fig. 2 The classification diagram of TWINSPAN for the herb layer of post-fire regenerated communities. N represents the number of transects. A, B, C, and D designate four types of post-fire regenerated herb communities. A, Pteridium aquilinum var. latiusculum + Ageratina adenophora + Gentiana cephalantha community; B, Arundinella hookeri + Digitaria violascens + Bothriochloa ischaemum community; C, Kummerowia striata + Bulbostylis densa + Bothriochloa ischaemum community; D, Arthraxon hispidus + Kummerowia striata + Bothriochloa ischaemum community. (+) means it is positive with classification; (-) means it is negetive with classification.
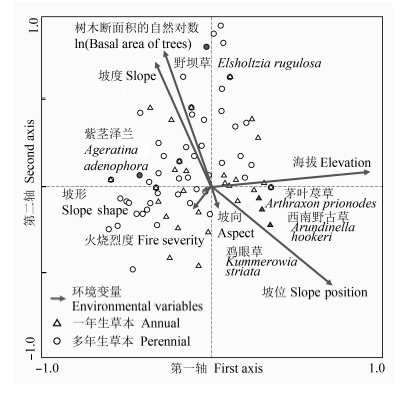
图3 火后更新植物群落中一年生(∆)和多年生(○)草本物种的典范对应分析排序格局。
Fig. 3 The canonical correspondence analysis ordination pattern for the annual (∆) and perennial (○) plant species in the early post-fire regenerated communities.
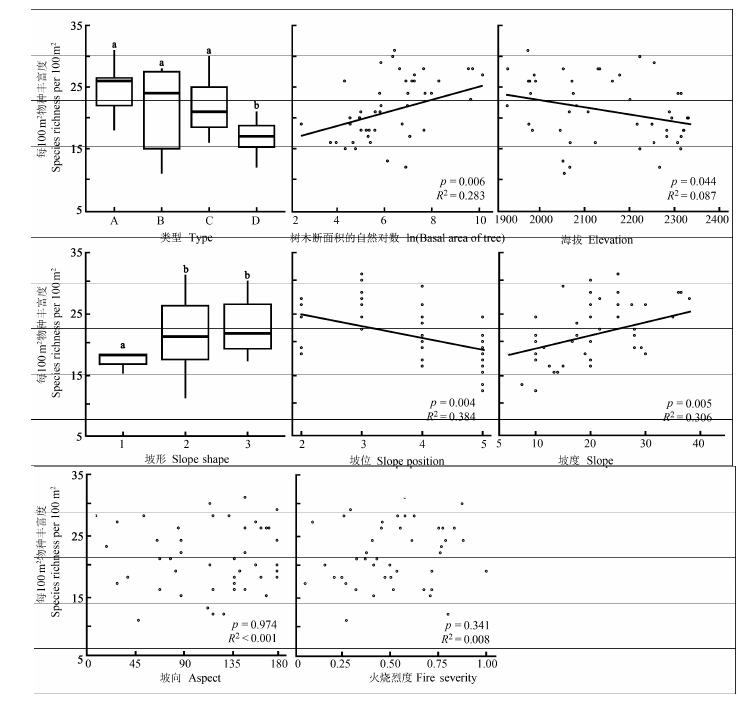
图4 火后植物群落样带(1 000 m2)草本层物种丰富度与生境因子的关系。所有小图中标注的a、b代表类型间t检验显著的不同水平。A、B、C、D代表4种火后草本更新群落类型。A代表蕨+紫茎泽兰+头花龙胆群落; B代表西南野古草+紫马唐+白羊草群落; C代表鸡眼草+丝叶球柱草+白羊草群落; D代表荩草+鸡眼草+白羊草群落。
Fig. 4 The relationships between habitat factors and the species richness in herb layer of the transects of post-fire plant communities. The letters a and b above all box-plots refer to significantly different levels in t-test. A, B, C, and D designate four types of post-fire regenerated herb communities. A, Pteridium aquilinum var. latiusculum + Ageratina adenophora + Gentiana cephalantha community; B, Arundinella hookeri + Digitaria violascens + Bothriochloa ischaemum community; C, Kummerowia striata + Bulbostylis densa + Bothriochloa ischaemum community; D, Arthraxon hispidus + Kummerowia striata + Bothriochloa ischaemum community.
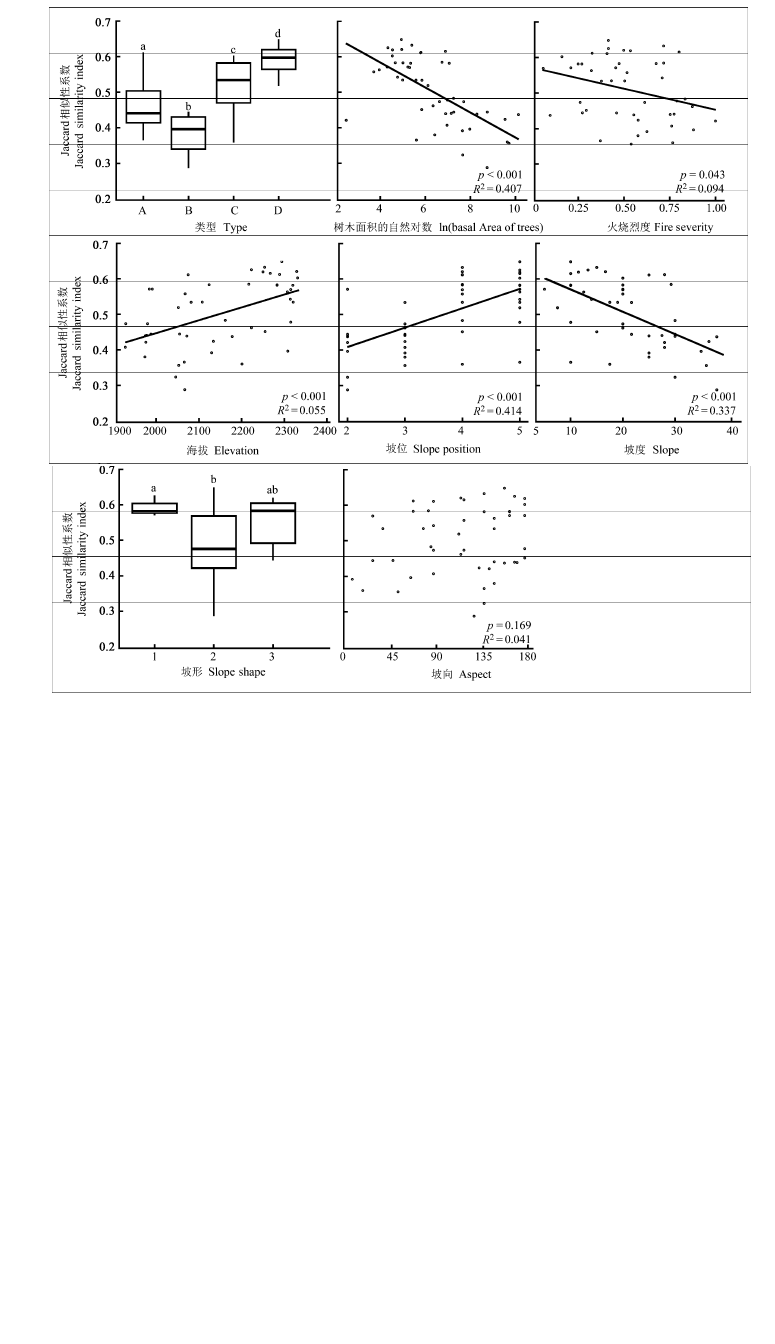
图5 火后植物群落草本层β多样性格局。所有小图中标注的a、b表示类型间t检验显著的不同水平。A、B、C、D 代表4种火后草本更新群落类型。第一幅小图中A代表蕨+紫茎泽兰+头花龙胆群落; B代表西南野古草+紫马唐+白羊草群落; C代表鸡眼草+丝叶球柱草+白羊草群落; D代表荩草+鸡眼草+白羊草群落。
Fig. 5 The patterns of β-diversity in herb layer of post-fire plant communities. The letters a and b above all box-plots refer to significantly different levels in t-test. A, B, C, and D in the first graph panel designate four types of post-fire regenerated herb communities. A, Pteridium aquilinum var. latiusculum + Ageratina adenophora + Gentiana cephalantha community; B, Arundinella hookeri + Digitaria violascens + Bothriochloa ischaemum community; C, Kummerowia striata + Bulbostylis densa + Bothriochloa ischaemum community; D, Arthraxon hispidus + Kummerowia striata + Bothriochloa ischaemum community.
| 生境因子 Habitat factor | α多样性 α diversity | β多样性 β diversity | |||
|---|---|---|---|---|---|
| 样方尺度 Plot scale | 样带尺度 Transect scale | Jaccard系数 Jaccard index | Morisita-Horn系数 Morisita-Horn index | ||
| 海拔 Elevation | 3.8 | 12.0 | 16.8* | 1.9 | |
| 坡向 Aspect | 1.6 | 1.9 | 1.6 | 9.2 | |
| 坡形 Slope shape | 17.9* | 6.4 | 1.5 | 5.3 | |
| 坡位 Slope position | 54.7* | 30.6* | 20.7* | 13.4 | |
| 乔木胸面积对数 ln(sum of tree basal area) | 3.4 | 26.6* | 38.6* | 41.6* | |
| 坡度 Slope | 8.7* | 20.9* | 11.9* | 19.5* | |
| 火烧强度 Fire severity | 9.9* | 1.5 | 8.9* | 9.0 | |
| 调整的决定系数 Adjusted coefficient of determination | 0.172 | 0.645 | 0.678 | 0.638 | |
表2 生境因子对火后植物群落草本层物种多样性的影响
Table 2 Effects of habitat factors on the species diversity in herb layer of post-fire plant communities
| 生境因子 Habitat factor | α多样性 α diversity | β多样性 β diversity | |||
|---|---|---|---|---|---|
| 样方尺度 Plot scale | 样带尺度 Transect scale | Jaccard系数 Jaccard index | Morisita-Horn系数 Morisita-Horn index | ||
| 海拔 Elevation | 3.8 | 12.0 | 16.8* | 1.9 | |
| 坡向 Aspect | 1.6 | 1.9 | 1.6 | 9.2 | |
| 坡形 Slope shape | 17.9* | 6.4 | 1.5 | 5.3 | |
| 坡位 Slope position | 54.7* | 30.6* | 20.7* | 13.4 | |
| 乔木胸面积对数 ln(sum of tree basal area) | 3.4 | 26.6* | 38.6* | 41.6* | |
| 坡度 Slope | 8.7* | 20.9* | 11.9* | 19.5* | |
| 火烧强度 Fire severity | 9.9* | 1.5 | 8.9* | 9.0 | |
| 调整的决定系数 Adjusted coefficient of determination | 0.172 | 0.645 | 0.678 | 0.638 | |
| [1] | Abella SR, Covington WW (2006). Vegetation-environment relationships and ecological species groups of an Arizona Pinus ponderosa landscape, USA.Plant Ecology, 185, 255-268. |
| [2] | Ackerly D (2004). Functional strategies of chaparral shrubs in relation to seasonal water deficit and disturbance.Ecological Monographs, 74, 25-44. |
| [3] | Bond W, Keeley J (2005). Fire as a global ‘herbivore’: The ecology and evolution of flammable ecosystems.Trends in Ecology & Evolution, 20, 387-394. |
| [4] | Boneau CA (1960). The effects of violations of assumptions underlying the t test.Psychological Bulletin, 57, 49-64. |
| [5] | Bormann FH, Likens GE (2012). Pattern and Process in a Forested Ecosystem: Disturbance, Development and the Steady State Based on the Hubbard Brook Ecosystem Study. Springer Science & Business Media, New York. |
| [6] | Boucher D, Gauthier S, Noël J, Greene DF, Bergeron Y (2014). Salvage logging affects early post-fire tree composition in Canadian boreal forest.Forest Ecology and Management, 325,118-127. |
| [7] | Bowman DM, Balch JK, Artaxo P, Bond WJ, Carlson JM, Cochrane MA, D’Antonio CM, DeFries RS, Doyle JC, Harrison SP, Johnston FH, Keeley JE, Krawchuk MA, Kull CA, Marston JB, Moritz MA, Prentice IC, Roos CI, Scott AC, Swetnam TW, van der Werf GR, Pyne SJ (2009). Fire in the Earth system.Science, 324, 481-484. |
| [8] | Bowman DM, Balch JK, Artaxo P, Bond WJ, Cochrane MA, D’Antonio CM, DeFries R, Johnston FH, Keeley JE, Krawchuk MA, Kull CA, Mack M, Moritz MA, Pyne S, Roos CI, Scott AC, Sodhi NS, Swetnam TW (2011). The human dimension of fire regimes on Earth.Journal of Biogeography, 38, 2223-2236. |
| [9] | Broncano MJ, Retana J (2004). Topography and forest composition affecting the variability in fire severity and post-fire regeneration occurring after a large fire in the Mediterranean basin.International Journal of Wildland Fire, 13, 209-216. |
| [10] | Camac JS, Williams RJ, Wahren C, Morris WK, Morgan JW (2013). Post-fire regeneration in alpine heathland: Does fire severity matter?Austral Ecology, 38, 199-207. |
| [11] | Casady GM, van Leeuwen WJD, Marsh SE (2010). Evaluating post-wildfire vegetation regeneration as a response to multiple environmental determinants.Environmental Modeling & Assessment, 15, 295-307. |
| [12] | Crotteau JS, Morgan Varner J, Ritchie MW (2013). Post-fire regeneration across a fire severity gradient in the southern Cascades.Forest Ecology and Management, 287, 103-112. |
| [13] | Dai CR, Gu BW, Li JH (2011). The relationship between the severely affected forest fire season in Yunnan and the abnormal change of the Pacific Ocean surface temperature. Journal of Yunnan University (Natural Sciences), 33(S1), 188-191. (in Chinese with English abstract)[戴丛蕊, 顾本文, 李金槐 (2011). 云南林火重灾年与太平洋海温异常变化关系的研究. 云南大学学报(自然科学版), 33(S1), 188-191.] |
| [14] | Forrestel EJ, Donoghue MJ, Smith MD (2014). Convergent phylogenetic and functional responses to altered fire regimes in mesic savanna grasslands of North America and South Africa.New Phytologist, 203, 1000-1011. |
| [15] | Fox DM, Maselli F, Carrega P (2008). Using SPOT images and field sampling to map burn severity and vegetation factors affecting post forest fire erosion risk.Catena, 75, 326-335. |
| [16] | Guo Q (2001). Early post-fire succession in California chaparral: Changes in diversity, density, cover and biomass.Ecological Research, 16, 471-485. |
| [17] | Han J, Shen ZH, Ying LX, Chen A (2015). Early post-fire regeneration of a fire-prone subtropical Pinus mixed forest in Yunnan, southwest China: The effects of pre-fire vegetation, fire severity and topographic factors.Forest Ecology & Management, 356, 31-40. |
| [18] | Hill MO (1979). TWINSPAN—A FORTRAN Program for Arranging Multivariate Data in an Ordered Two-Way Table by Classification of the Individuals and Attributes. Cornell University Press, New York. |
| [19] | Hurtt GC, Pacala SW (1995). The consequences of recruitment limitation: reconciling chance, history and competitive differences between plants.Journal of Theoretical Biology, 176, 1-12. |
| [20] | Jaccard P (1912). The distribution of the flora in the alpine zone.New Phytologist, 11, 37-50. |
| [21] | Jin ZZ, Peng J (1998). Vegetation of Kunming. Yunnan Science and Technology Press, Kunming. (in Chinese)[金振洲, 彭鉴 (1988). 昆明植被. 云南科技出版社, 昆明.] |
| [22] | Keeley JE, Fotheringham CJ, Baer-Keeley M (2005). Factors affecting plant diversity during post-fire recovery and succession of Mediterranean-climate shrublands in California, USA.Diversity Distributions, 11, 525-537. |
| [23] | Kurkowski TA, Mann DH, Rupp TS, Verbyla DL (2008). Relative importance of different secondary successional pathways in an Alaskan boreal forest.Canadian Journal of Forest Research, 38, 1911-1923. |
| [24] | Lee J, Lee S, Lim J, Won M, Lee H (2014). Effects of heterogeneity of pre-fire forests and vegetation burn severity on short-term post-fire vegetation density and regeneration in Samcheok, Korea.Landscape and Ecological Engineering, 10, 215-228. |
| [25] | Liu CJ, Yang ZZ, Li Q, Lu ZX, Zhang W (2012). Impact of fire disturbance on Carabid beetles communities in forest of Pinus yunnanensis in Cangshan.Sichuan Journal of Zoology, 3, 387-392. (in Chinese with English abstract)[刘春菊, 杨自忠, 李巧, 卢志兴, 张威 (2012). 火干扰对苍山云南松林步甲群落的影响. 四川动物, 3, 387-392.] |
| [26] | Lobo J (2001). Spatial and environmental determinants of vascular plant species richness distribution in the Iberian Peninsula and Balearic Islands.Biological Journal of the Linnean Society, 73, 233-253. |
| [27] | Maia P, Keizer J, Vasques A, Abrantes N, Roxo L, Fernandes P, Ferreira A, Moreira F (2014). Post-fire plant diversity and abundance in pine and eucalypt stands in Portugal: Effects of biogeography, topography, forest type and post-fire management.Forest Ecology and Management, 334,154-162. |
| [28] | Maia P, Pausas JG, Vasques A, Keizer JJ (2012). Fire severity as a key factor in post-fire regeneration of Pinus pinaster (Ait.) in Central Portugal.Annals of Forest Science, 69, 489-498. |
| [29] | Mayer H, Swetnam TW (2000). Century scale climate forcing of fire regimes in the American Southwest.The Holocene, 10, 213-220. |
| [30] | Mouillot F, Ratte J, Joffre R, Mouillot D, Serge Rambal A (2005). Long-term forest dynamic after land abandonment in a fire prone Mediterranean landscape (central Corsica, France).Landscape Ecology, 20, 101-112. |
| [31] | Oliveira S, Pereira JMC, San-Miguel-Ayanz J, Lourenço L (2014). Exploring the spatial patterns of fire density in Southern Europe using Geographically Weighted Regression.Applied Geography, 51, 143-157. |
| [32] | Pausas JG (1994). Species richness patterns in the understory of Pyrenean Pinus sylvestris forest.Journal of Vegetation Science, 5, 517-524. |
| [33] | Roder A, Hill J, Duguy B, Alloza J, Vallejo R (2008). Using long time series of Landsat data to monitor fire events and post-fire dynamics and identify driving factors. A case study in the Ayora region (eastern Spain).Remote Sensing of Environment, 112, 259-273. |
| [34] | Roleček J, Tichý L, Zelený D, Chytrý M (2009). Modified TWINSPAN classification in which the hierarchy respects cluster heterogeneity.Journal of Vegetation Science, 20, 596-602. |
| [35] | Shen ZH, Fei SL, Feng JM, Liu YN, Liu ZL, Tang ZY, Wang XP, Wu XP, Zheng CY, Zhu B, Fang JY (2012). Geographical patterns of community-based tree species richness in Chinese mountain forests: The effects of contemporary climate and regional history.Ecography, 35, 1134-1146. |
| [36] | Shen ZH, Zhang XS, Jin YX (2000a). Spatial pattern analysis and topographical interpretation of species diversity in the forests of Dalaoling in the region of the Three Gorges.Acta Botanica Sinica, 42, 620-627. (in Chinese with English abstract)[沈泽昊, 张新时, 金义兴 (2000a). 三峡大老岭森林物种多样性的空间格局分析及其地形解释. 植物学报, 42, 620-627.] |
| [37] | Shen, ZH, Jin YX, Zhao ZE, Wu JQ, Huang HD (2000b). A study on the quantitative classification of forest communities of Dalaoling region at the Three Gorges.Journal of Wuhan Botanical Research, 18, 99-107. (in Chinese with English abstract)[沈泽昊, 金义兴, 赵子恩, 吴金清, 黄汉东 (2000b). 三峡大老岭地区森林群落的数量分类研究. 武汉植物学研究, 18, 99-107.] |
| [38] | Su WH, Shi Z, Zhou R, Zhao YJ, Zhang GF (2015). Thr role of fire in the Central Yunnan Plateau ecosystem, southwestern China.Forest Ecology and Management, 356, 22-30. |
| [39] | Swanson FJ, Kratz TK, Caine N (1988). Landform effects on ecosystem patterns and processes.BioScience, 38, 92-98. |
| [40] | Tang RR, Zhou RL (2007). Study on the fuel loading forecast models of Pinus yunnanensis.Shandong Forestry Science and Technology, 1, 1-4. (in Chinese with English abstract)[唐荣逸, 周汝良 (2007). 云南松可燃物载量预测模型研究. 山东林业科技, 1, 1-4.] |
| [41] | Vallejo VR, Alloza JA (2012). Post-fire management in the Mediterranean Basin.Israel Journal of Ecology and Evolution, 58, 251-264. |
| [42] | Walsh C, Mac Nally R (. |
| [43] | Wang QH, Shu LF, Li SY (2011). Study on spotting of Pinus yunnanensis forest during burning.Journal of Safety Science and Technology, 7, 49-53. (in Chinese with English abstract)[王秋华, 舒立福, 李世友 (2011). 云南松林燃烧过程中飞火的研究.中国安全生产科学技术, 7, 49-53.] |
| [44] | Wang S, Niu SK, Li D, Wang JH, Chen F, Sun W (2013). Vertical distribution of fuels in Pinus yunnanensis forest and related affecting factors. Chinese Journal of Applied Ecology, 24, 331-337. (in Chinese with English abstract)[王叁, 牛树奎, 李德, 王景华, 陈锋, 孙武 (2013). 云南松林可燃物的垂直分布及影响因子. 应用生态学报, 24, 331-337.] |
| [45] | Whigham DF (2004). Ecology of woodland herbs in temperate deciduous forests.Annual Review of Ecology, Evolution, and Systematics, 35, 583-621. |
| [46] | Wolda H (1981). Similarity indices, sample size and diversity.Oecologia, 50, 296-302. |
| [47] | Zhang YT, Duan X, Li JF (1994). Climatic regionalization of forest fires in the central Yunnan region.Journal of Southwest Forestry College, 3, 122-127. (in Chinese with English abstract)[张映堂, 段旭, 李俊峰 (1994). 滇中地区林火气候区划. 西南林学院学报, 3, 122-127.] |
| [1] | 李帅锋, 郎学东, 黄小波, 王艳红, 刘万德, 徐崇华, 苏建荣. 云南普洱30 hm2季风常绿阔叶林动态监测样地群丛数量分类[J]. 植物生态学报, 2020, 44(3): 236-247. |
| [2] | 王艳红, 李帅锋, 郎学东, 黄小波, 刘万德, 徐崇华, 苏建荣. 地形异质性对云南普洱季风常绿阔叶林物种多样性的影响[J]. 植物生态学报, 2020, 44(10): 1015-1027. |
| [3] | 陈云, 王婷, 李培坤, 姚成亮, 袁志良, 叶永忠. 河南木札岭温带落叶阔叶林群落特征及主要乔木空间分布格局[J]. 植物生态学报, 2016, 40(11): 1179-1188. |
| [4] | 许洺山, 赵延涛, 杨晓东, 史青茹, 周刘丽, 阎恩荣. 浙江天童木本植物叶片性状空间变异的地统计学分析[J]. 植物生态学报, 2016, 40(1): 48-59. |
| [5] | 史青茹, 许洺山, 赵延涛, 周刘丽, 张晴晴, 马文济, 赵绮, 阎恩荣. 浙江天童木本植物Corner法则的检验: 微地形的影响[J]. 植物生态学报, 2014, 38(7): 665-674. |
| [6] | 杨婧, 褚鹏飞, 陈迪马, 王明玖, 白永飞. 放牧对内蒙古典型草原α、β和γ多样性的影响机制[J]. 植物生态学报, 2014, 38(2): 188-200. |
| [7] | 梁爽, 许涵, 林家怡, 李意德, 林明献. 尖峰岭热带山地雨林优势树种白颜树空间分布格局[J]. 植物生态学报, 2014, 38(12): 1273-1282. |
| [8] | 余敏,周志勇,康峰峰,欧阳帅,米湘成,孙建新. 山西灵空山小蛇沟林下草本层植物群落梯度分析及环境解释[J]. 植物生态学报, 2013, 37(5): 373-383. |
| [9] | 许玥, 沈泽昊, 吕楠, 唐圆圆, 李道新, 王功芳, 谭家林, 刘益平. 湖北三峡大老岭自然保护区光叶水青冈群落种子雨10年观测: 种子雨密度、物种构成及其与群落的关系[J]. 植物生态学报, 2012, 36(8): 708-716. |
| [10] | 龙文兴, 丁易, 臧润国, 杨民, 陈少伟. 海南岛霸王岭热带云雾林雨季的环境特征[J]. 植物生态学报, 2011, 35(2): 137-146. |
| [11] | 赖江山, 米湘成, 任海保, 马克平. 基于多元回归树的常绿阔叶林群丛数量分类——以古田山24公顷森林样地为例[J]. 植物生态学报, 2010, 34(7): 761-769. |
| [12] | 苏志尧, 刘刚, 区余端, 戴朝晖, 李镇魁. 车八岭山地常绿阔叶林冰灾后林木受损的生态学评估[J]. 植物生态学报, 2010, 34(2): 213-222. |
| [13] | 吴大千, 刘建, 王炜, 丁文娟, 王仁卿. 黄河三角洲植被指数与地形要素的多尺度分析[J]. 植物生态学报, 2009, 33(2): 237-245. |
| [14] | 胡志伟, 沈泽昊, 吕楠, 赵俊, 李道兴, 陈华, 王功芳. 地形对森林群落年龄及其空间格局的影响[J]. 植物生态学报, 2007, 31(5): 814-824. |
| [15] | 杨永川, 达良俊. 丘陵地区地形梯度上植被格局的分异研究概述[J]. 植物生态学报, 2006, 30(3): 504-513. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
Copyright © 2022 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19