|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
滤除自然光中UV-B辐射成分对高山植物美丽风毛菊光合生理的影响
植物生态学报
2011, 35 (2):
176-186.
DOI: 10.3724/SP.J.1258.2011.00176
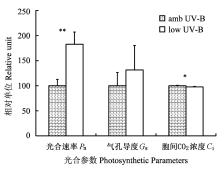
采用滤除自然光谱中UV-B辐射成分的方法, 探讨了高山植物美丽风毛菊(Saussurea superba)光合机构对青藏高原强UV-B辐射的响应和适应特性。结果表明, 强太阳光中的UV-B成分能引起净光合速率的降低。连续16天不同天气下的观测表明, 滤除UV-B处理时3 min暗适应的光化学量子效率有升高的趋势; 晴天下稳态光化学效率的分析也显示滤除UV-B处理的实际光化学量子效率和光化学猝灭系数有升高趋势, 意味着自然光中的UV-B成分可限制美丽风毛菊叶片PSII反应中心的激发能捕获效率。PSII有效光化学量子效率的增加和非光化学猝灭系数的降低进一步表明, UV-B辐射能导致有效光化学效率的降低和非光化学能量耗散的增加。由上可知, 自然强UV-B辐射是限制美丽风毛菊叶片光合作用的一个因素。滤除UV-B辐射处理对光合色素含量的影响较小, 无论以叶面积还是叶鲜重为基础的滤除UV-B处理仅有微弱的增加趋势, 说明强UV-B辐射具有加速光合色素的光氧化进程, 促进细胞成熟和叶片衰亡的潜在作用。同样UV-B吸收物质的含量也几乎没有变化, 表明强太阳辐射环境下生活的高山植物美丽风毛菊叶表皮层中已具有较多的紫外线屏蔽物质, 足以抵御目前环境中强太阳UV-B辐射可能引起的伤害, 较少受UV-B辐射波动的影响。
表2
2008年8月滤除自然光中UV-B成分处理对美丽风毛菊叶片光合色素的影响(平均值±标准误差, n = 15; p = 0.05)
正文中引用本图/表的段落
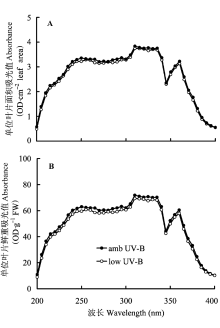
取经UV-B辐射处理后的成熟叶片数枚, 用直径0.7 cm的打孔器取叶圆片, 浸入内盛10 mL 79%酸化甲醇的样品瓶中, 盖紧密封, 避光低温浸提约10天。UV-B吸收物质的含量以10 mL酸化甲醇提取液中, 单位叶面积或叶鲜重在200-400 nm范围内的紫外吸收曲线表示, 即OD·cm-2·10 mL-1或OD·g-1 FW·10 mL-1 (Shi et al., 2004)。
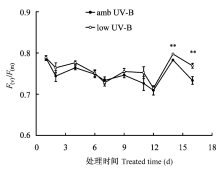
连续16天的滤除试验表明, 除第7天外, low UV-B处理时美丽风毛菊叶片的F(v)/F(m)有升高趋势, 即自然光谱中的UV-B辐射对暗适应3 min后的PSII光化学量子效率F(v)/F(m)有负影响; 试验后期尽管田间采样和叶片趋于枯黄等导致了供试样本数目的降低, 但第14天(阴天)和第16天(晴天)都表现出极显著差异(图2)。
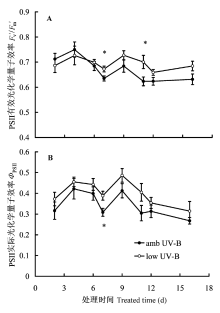
图3表明, low UV-B处理时, 除第2天和第4天的两次测定(p分别为0.474 9和0.651 6; n分别为10和4) 外, 美丽风毛菊叶片Fv′/Fm′高于对照amb UV-B (p分别为0.516 6、0.021 4、0.168 3、0.041 4、0.103 5和0.136 5; n分别为16、15、9、5、5和4) (图3A); 而ΦPSII在整个处理期间均较高(p分别为0.304 3、0.567 1、0.311 6、0.010 1、0.138 7、0.113 8、0.347 1和0.398 3; n分别为10、4、16、15、9、5、5和4) (图3B)。
表2表明, 尽管叶片光合色素的含量(分别用单位叶片面积和单位叶片鲜重表示)都没有达到显著性差异水平, 但与对照amb UV-B相比, low UV-B处理后美丽风毛菊叶片中光合色素含量均有升高的趋势; 叶绿素a和叶绿素b的比值Chl a/b以及类胡萝卜素和总叶绿素的比值Car/Chl也都略有升高的趋势。
由叶绿素基础荧光参数得到的诸多PSII光化学效率参数, 是评价光合机构PSII光化学活性的有利工具(张守仁, 1999)。最大光化学效率(Fv/Fm), 也称PSII光化学最大量子效率或开放PSII反应中心的能量捕获效率, 通常以没有遭受环境胁迫并经过充分暗适应的叶片进行测定, 用于估测植物叶片PSII反应中心的内禀光能转化效率。在胁迫环境中或没有充分暗适应恢复时, Fv/Fm将不再代表最大意义的光化学效率, 而是反映光合机构的光化学活性受抑制后开放PSII反应中心可能具有的能量捕获效率(许大全, 2002)。本文采用暗适应3 min后的PSII光化学量子效率F(v)/F(m)来探测UV-B辐射对叶片光化学效率的影响, 一定时间内F(v)/F(m)的相对变化可以用来判定PSII反应中心的失活或损伤程度(Galvez-Valdivieso et al., 2009)。3 min暗适应能保证依赖类囊体膜内外质子浓度差而产生的非光化学猝灭的快相组分完全恢复, 但植物叶片的潜在量子效率尚未能完全恢复, 这主要由光合作用光抑制有关的慢相组分以及状态转换的中间相组分所致, 尤其对于遭受胁迫的植物叶片(Quick & Stitt, 1989)。图2表明, 尽管天气状况不尽相同, 连续16天的滤除试验显示amb UV-B处理能引起F(v)/F(m)不同程度的降低, 说明高原强自然太阳光中的UV-B成分能影响美丽风毛菊植物的光化学活性。但连续滤除试验时并没有观测到UV-B辐射效应的累积, 即没有观测到amb UV-B处理下F(v)/F(m)逐渐降低以及与low UV-B处理间差异渐趋显著的现象, 这说明具有较多UV-B成分在内的强太阳短波辐射能引起F(v)/F(m)降低加剧, 但对生长在青藏高原恶劣环境中的土著高山植物来讲, 这主要是一种PSII反应中心的光失活或光诱导天线猝灭的增加, 而非PSII反应中心的光伤害加剧。PSII反应中心是UV-B辐射的敏感位点(van Rensen et al., 2007), 受损D1蛋白亚基的降解和祛除以及新D1蛋白亚基的合成都受温度降低的制约(Baker, 1996)。高原地区植物生长季节的多数时间昼夜温差很大, 夜间温度常在0 ℃以下, 但连续amb UV-B和low UV-B处理的美丽风毛菊叶片F(v)/F(m)都没有出现逐渐降低的现象, 表明F(v)/F(m)能快速恢复, 强太阳光引起的光化学效率降低主要是一种PSII天线蛋白复合体中依赖光的激发能猝灭的弛豫, F(v)/ F(m)的快速恢复显然不可能完全来自PSII反应中心的修复。图2中第1天、第4天和第14天为多云或较阴天的测定, 不同天气状况下两处理的F(v)/F(m)差异具有相同的变化趋势, 也进一步验证了UV-B辐射的负效应。同时, 3个阴天的F(v)/F(m)相对较高也表明晴天时强可见光辐射(通常大于1 500 mol·m-2·s-1)可诱导光合作用的光抑制甚至光损伤, 而自然光谱中的UV-B辐射成分并不是光抑制现象的主要因素。以上结果也说明, 高山植物的叶片D1蛋白亚基的光伤害速率和光伤害后PSII复合体的修复速率是相匹配的, 因此能维持稳定的有效PSII反应中心复合体组成, 反映了高山植物对包括强UV-B辐射在内的高原强太阳短波辐射的适应性。
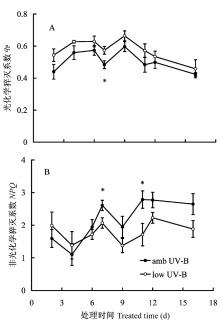
稳态PSII光化学效率受测定时自然光强的影响较大, 故计算时仅挑选了晴天的数据。ΦPSII 为作用光下的PSII实际光化学量子效率, 代表PSII反应中心电荷分离的实际量子效率。ΦPSII与通过PSII反应中心的线性电子传递通量(liner electron flux, LEF)有直接的联系, 因此又被称为PSII运行效率(PSII operating efficiency), 可快速确定不同光下或其他环境条件下PSII反应中心的作用效率(Baker, 2008)。qP表示开放PSII反应中心占总PSII反应中心的比例, 为PSII反应中心初级接受体QA氧化还原状态的相对度量, 其数值与开放PSII反应中心呈非线性比例, 反映了QA氧化程度。氧化态的QA是PSII电子传递的原初醌受体, 决定着PSII激发能的捕获效率。qP也被称作PSII效率因子(PSII efficiency factor), 它将稳态作用光下的Fv′/Fm′和ΦPSII联系在一起(Baker, 2008)。短期滤除UV-B试验表明, low UV-B处理时ΦPSII和qP都增高(图3B, 图4A), 说明自然光中的UV-B成分依然是限制美丽风毛菊叶片光化学效率的一个因素。除第2天和第4天外, low UV-B处理后也引起了Fv′/Fm′的增加(图3A); 与Fv′/Fm′变化趋势相反, 除第2天和第4天外, low UV-B处理后导致了NPQ的降低(图4B)。Fv′/Fm′与反映整个光合作用过程的ΦPSII不同, 仅表示开放PSII反应中心的激发能捕获效率, 显然, Fv′/Fm′也能够用来评估非光化学猝灭对ΦPSII变化的贡献(Baker, 2008)。滤除自然光谱中UV-B成分处理时, NPQ以及Fv′/Fm′的变化表明, 自然光中的UV-B辐射成分能导致非光化学能量耗散的增加。进一步分析可知, 除滤除试验的第2天和第4天外, amb UV-B处理的Fs和Fm′都相对较低(图表未列), 显然Fm′是引起NPQ以及Fv′/Fm′变化的原因。
2008年8月的试验数据表明, 无论以单位叶片面积为基础还是以单位叶片鲜重为基础, low UV-B处理后美丽风毛菊叶片光合色素含量均略有增加的趋势, 且以单位叶片面积表示时略微明显(表2)。叶绿素含量的高低在一定程度上能反映植物利用光能及制造有机物的能力。已有的文献中既有UV-B辐射降低光合色素含量的报道(Lizana et al., 2009), 也有中性甚至极少数正影响的报道(Middleton & Teramura, 1993)。滤除自然光谱中的UV-B成分处理对美丽风毛菊叶片的光合色素几无影响或略有增加, 似乎与2009年7月的测定以及返青期的多数测定并不一致。通常认为较强的UV-B辐射能破坏叶绿体的结构及其前体, 或使它的合成受阻, 从而降低叶绿素的含量; 而叶绿素含量的下降将减少叶片对光的吸收, 使植物能免于强辐射的损伤。叶绿素水平对UV-B的响应依赖于植物的发育阶段和环境条件, 强UV-B辐射下光合色素的降低可能缘于对光合色素合成过程的限制或(和)它的光氧化作用(Strid & Porra, 1992; Day & Neale, 2002)。8月份海北高寒草甸短暂的无霜期即将结束, 高山植物在逐渐频繁的夜间低温的影响下叶片更易衰老, 表现为叶尖和叶缘褐斑的出现和叶片枯黄。显然, 自然太阳光谱中的UV-B成分加速了这种过程, amb UV-B处理的美丽风毛菊叶片边缘和叶尖常有较多褐斑出现, 光合色素含量也趋于降低, 说明UV-B辐射能加速细胞成熟和促进衰亡过程(Gassi-Lit et al., 1997)。low UV-B处理时Car/Chl和Chl a/b都呈增加趋势(图4B), 即自然强UV-B辐射具有潜在的降低Car/Chl和Chl a/b比值的作用, 这与Lizana等(2009)在小麦(Triticum aestivum)不同物候期的UV-B试验结果一致。有实验证明, UV-B辐射降低Chl a/b比值与光捕获复合系统(LHC)对PSII和PSI核复合体相对数量的增加有关(?prtová et al., 2003)。
本文的其它图/表
|