
|
|
||
|
竹笋期竹箨和笋体的日间蒸腾特性及其对水分运输的影响
植物生态学报
2021, 45 (12):
1365-1379.
DOI: 10.17521/cjpe.2021.0164
竹子的高速生长主要发生在无枝无叶的笋期, 并对水分需求巨大。水分不仅参与植物体内各种代谢, 而且水分转运可促进光合产物、矿质元素、生长激素等物质流动。竹子夜间主要由根压驱动水分转运, 但在日间尤其是下午根压基本为负值, 明确竹笋日间蒸腾作用发生机制及其对水分运输的影响对竹林培育有重要意义。该研究以不同伸长阶段的慈竹(Bambusa emeiensis)笋为材料, 研究了茎秆和竹箨的气孔特征、气孔导度与蒸腾速率等生理特征及在离体条件下竹笋的水分转运速率。结果表明: 1)不同发育状态的竹笋茎秆及箨鞘表面均分布有大量气孔, 气孔小且凹陷, 光合速率及叶绿素a、b含量极低, 但气孔导度和蒸腾速率均显著高于成熟叶片, 表明笋体和箨鞘是竹笋主要的呼吸和蒸腾部位。2)离体条件下竹笋的番红示踪表明, 高生长阶段的竹笋茎秆中番红上升速率较快, 有着较强的蒸腾。竹箨分离后, 番红仍然能够扩散和运输, 表明笋体茎秆也存在一定的蒸腾, 但与竹箨包裹的竹笋相比, 番红在分离竹箨后的笋体中上升速度显著下降, 表明竹箨对笋体内水分运输影响较大。3)箨环处的组织解剖发现, 节间的纵向维管束在竹节处特化形成一个类板状结构, 弯曲伸入竹箨, 是竹箨影响笋体内水分运输的重要结构基础。上述结果表明, 日间竹笋水分通过茎秆和竹箨表面的气孔大量蒸散, 产生蒸腾拉力驱动笋体内水分转运。该研究也发现, 随着茎秆成熟, 竹箨松动并开始脱落, 茎秆表面的气孔宽度增加, 加大了气孔的开口大小, 增大了节间气孔与大气水气交换的有效面积, 在一定程度上弥补了竹箨脱落时减少的蒸腾拉力。 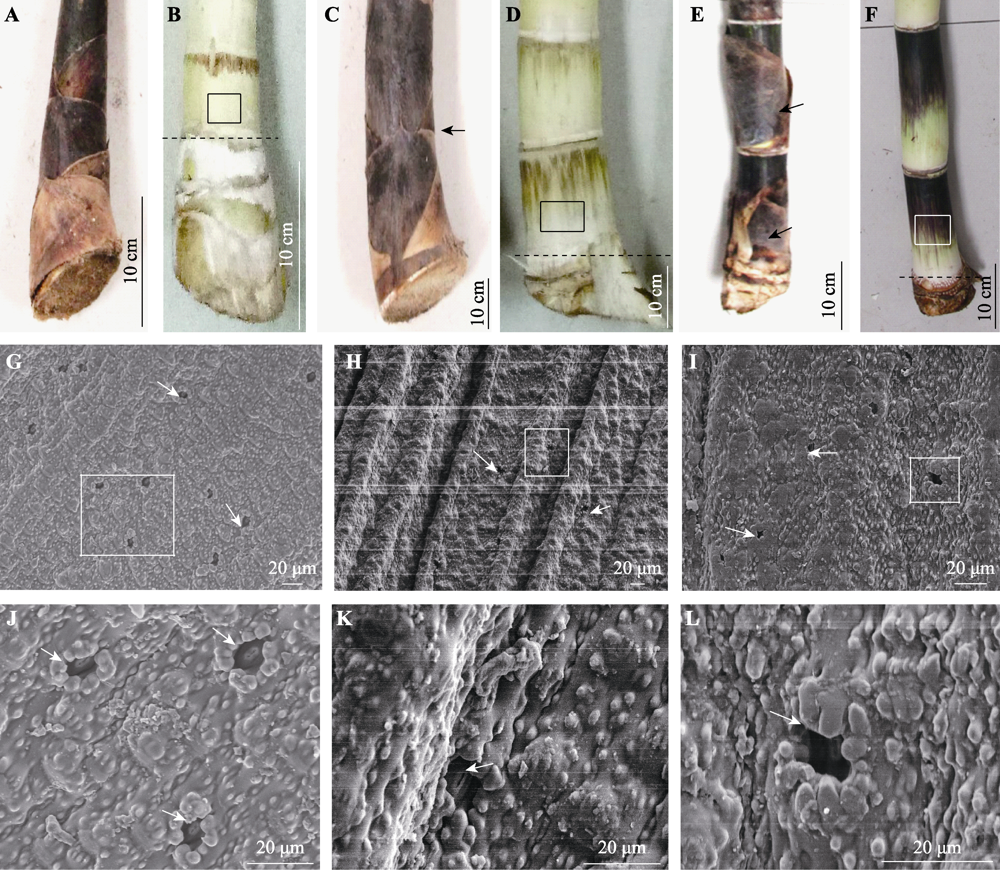 View image in article
图2
慈竹不同发育阶段笋体(茎秆)的气孔形态。A-F, 不同高度竹笋茎秆基部形态。基部第一节间正在伸长(A, B); 基部第一节间完全伸长, 节间部分露出竹箨(C, D), 箭头指向节间露出位置(C); 基部第一节间成熟, 竹箨松动并开始脱落(E, F), 箭头指向竹箨松动处(E)。B, D, F中的方框为取样部位, 箭头指向气孔, 虚线以上为无根节, 虚线以下为有根节。G-L, 扫描电镜下竹笋茎秆基部第一节间的气孔形态。G-I分别为节间的伸长期, 完全伸长期和成熟期; J-L分别为G-I中方框位置的局部放大图。
正文中引用本图/表的段落
分析处于伸长期(竹箨紧密包裹)、完全伸长期(有部分笋体露出竹箨)和成熟期(竹箨开始脱落)的竹笋基部第1节间的茎秆表面气孔特征, 结果表明: 有无箨体包裹的笋体表面都观察到气孔的存在(图2), 气孔形态与箨鞘相似, 气孔小且凹陷。并且, 基部第1节间从伸长到完全伸长, 节长和节宽增加2倍左右, 节间表面积增加5.77倍(p < 0.05); 随着节间伸长, 与之相对应的笋体表面的气孔长度、宽度上升, 气孔密度下降: 气孔长度增加2.11倍, 气孔宽度增加1.75倍, 气孔密度减小至22.10% (p < 0.05)(表2), 这种气孔长度与宽度显著正相关, 气孔大小与密度显著负相关的现象与已有的不同植物的报道(Franks et al., 2009; Casson & Hetherington, 2010; Liu et al., 2018)规律相似。
植物表皮上的气孔调控着植物与外界环境进行水气交换, 对植物的蒸腾与光合作用起着决定性的作用(Belin et al., 2010; Tayloy et al., 2012)。不同植物及同一植物的不同部位的气孔特征都存在差异, 如不同种类的变叶木(Codiaeum variegatum)叶表皮存在不同特征的气孔(杨洋等, 2013), 蚕豆(Vicia faba)茎上下表皮的气孔随环境光/暗变化表现出不同特征(任筠等, 2010)。在本研究中, 发现慈竹笋有/无竹箨包裹的茎秆及竹箨上均存在气孔, 茎秆和箨鞘表面的气孔小而凹陷(图1, 图2), 这种结构可以减少水分散失, 且更易储存大气中的水分(石辉等, 2011), 箨叶表面的气孔与成熟叶片相似, 大且分布浅。这些气孔特征表明慈竹茎秆及箨鞘上的气孔与外界环境的水分交换密切相关。
本文的其它图/表
|


{kind=link}