
Chin J Plant Ecol ›› 2023, Vol. 47 ›› Issue (10): 1407-1421.DOI: 10.17521/cjpe.2022.0276 cstr: 32100.14.cjpe.2022.0276
Special Issue: 植物功能性状
• Research Articles • Previous Articles Next Articles
CHEN Tu-Qiang, XU Gui-Qing( ), LIU Shen-Si, LI Yan
), LIU Shen-Si, LI Yan
Received:2022-07-04
Accepted:2023-03-13
Online:2023-10-20
Published:2023-11-23
Contact:
* (Supported by:CHEN Tu-Qiang, XU Gui-Qing, LIU Shen-Si, LI Yan. Hydraulic traits adjustments and nonstructural carbohydrate dynamics of Haloxylon ammodendron under drought stress[J]. Chin J Plant Ecol, 2023, 47(10): 1407-1421.
Add to citation manager EndNote|Ris|BibTeX
URL: https://www.plant-ecology.com/EN/10.17521/cjpe.2022.0276
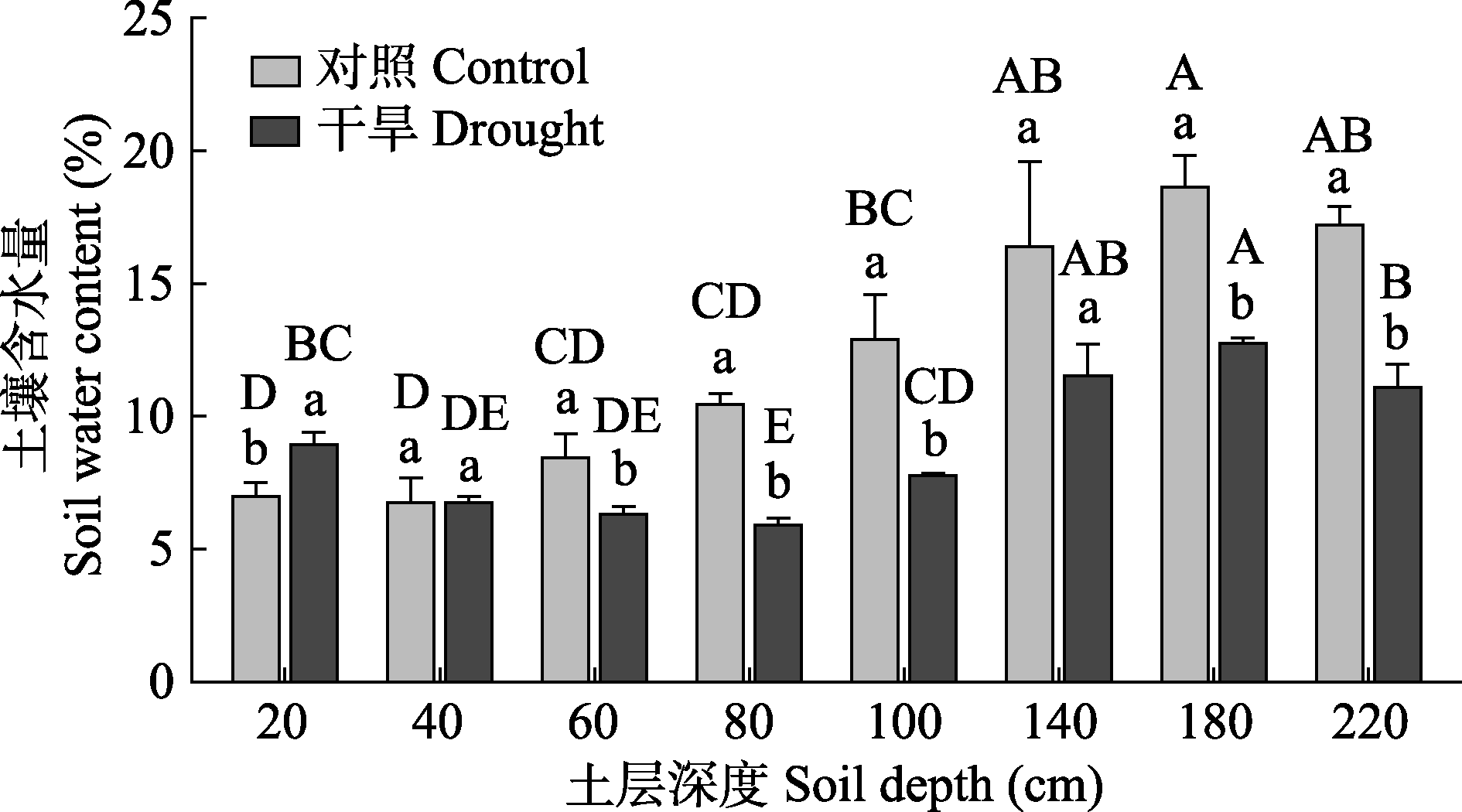
Fig. 1 Soil water content at different depths in Haloxylon ammodendron sampling plot (mean ± SE). Different uppercase letters indicate significant differences among soil depths of the same treatment (p < 0.05), and different lowercase letters indicate significant differences between treatments of the same soil depth (p < 0.05).
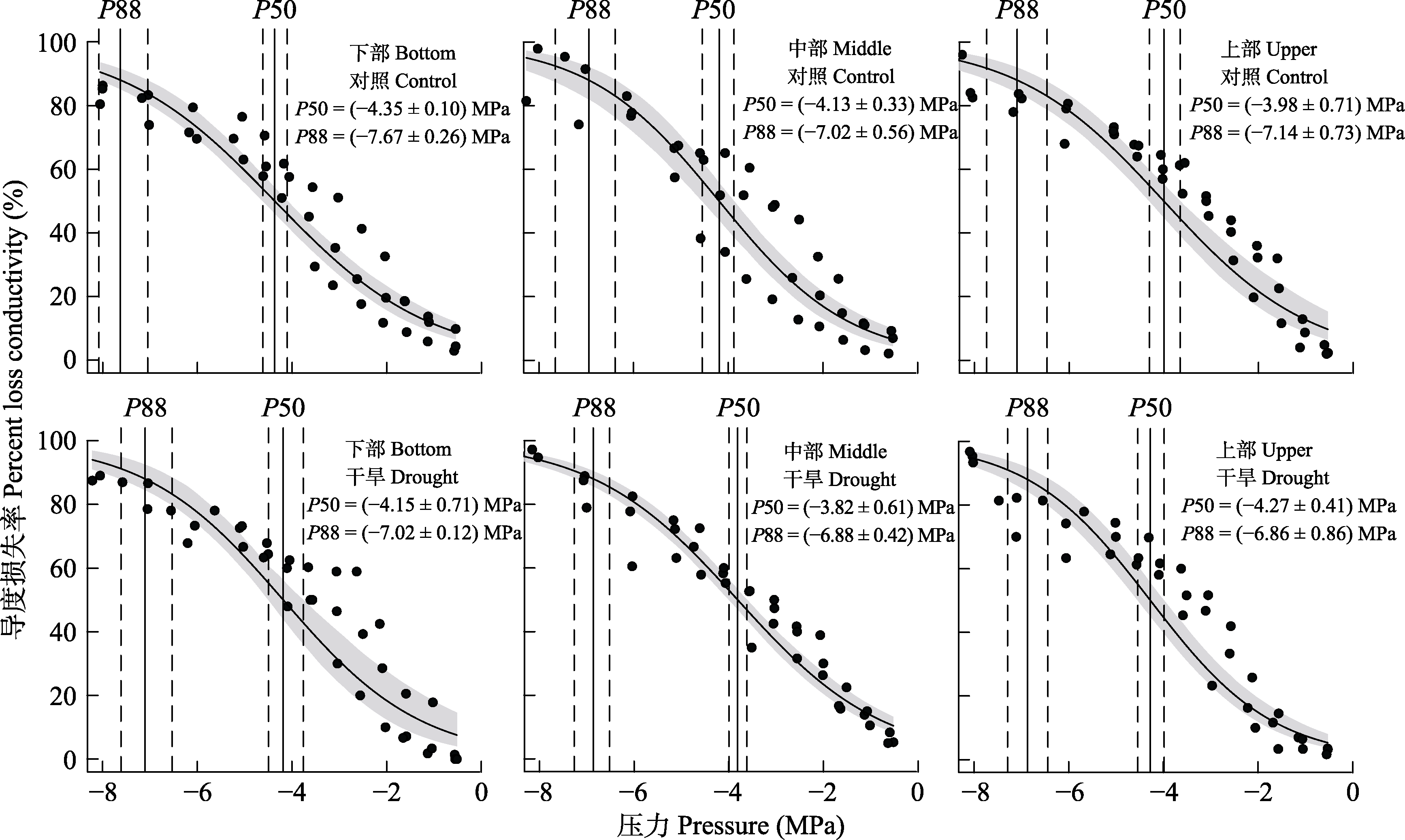
Fig. 2 Percent loss of xylem conductivity for Haloxylon ammodendron. The vertical solid lines indicate the pressure on the xylem (i.e. the water potential of the xylem) for 50% and 88% loss of maximum hydraulic conductivity (P50, P88), respectively; the dashed lines and shaded areas are 95% confidence intervals.
| P50 (MPa) | P88 (MPa) | |||||
|---|---|---|---|---|---|---|
| 下部 Bottom | 中部 Middle | 上部 Upper | 下部 Bottom | 中部 Middle | 上部 Upper | |
| 对照组 Control | -4.35 ± 0.10Aa | -4.13 ± 0.33Aa | -3.98 ± 0.17Aa | -7.67 ± 0.26Aa | -7.02 ± 0.56Aa | -7.10 ± 0.07Aa |
| 干旱组 Drought | -4.15 ± 0.17Aa | -3.82 ± 0.16Aa | -4.27 ± 0.14Aa | -7.03 ± 0.21Aa | -6.88 ± 0.24Aa | -6.86 ± 0.09Aa |
Table 1 Xylem water potential for 50% and 88% loss of maximum hydraulic conductivity (P50, P88) in upper, middle and lower branches of control and drought-treated Haloxylon ammodendron (mean ± SE)
| P50 (MPa) | P88 (MPa) | |||||
|---|---|---|---|---|---|---|
| 下部 Bottom | 中部 Middle | 上部 Upper | 下部 Bottom | 中部 Middle | 上部 Upper | |
| 对照组 Control | -4.35 ± 0.10Aa | -4.13 ± 0.33Aa | -3.98 ± 0.17Aa | -7.67 ± 0.26Aa | -7.02 ± 0.56Aa | -7.10 ± 0.07Aa |
| 干旱组 Drought | -4.15 ± 0.17Aa | -3.82 ± 0.16Aa | -4.27 ± 0.14Aa | -7.03 ± 0.21Aa | -6.88 ± 0.24Aa | -6.86 ± 0.09Aa |
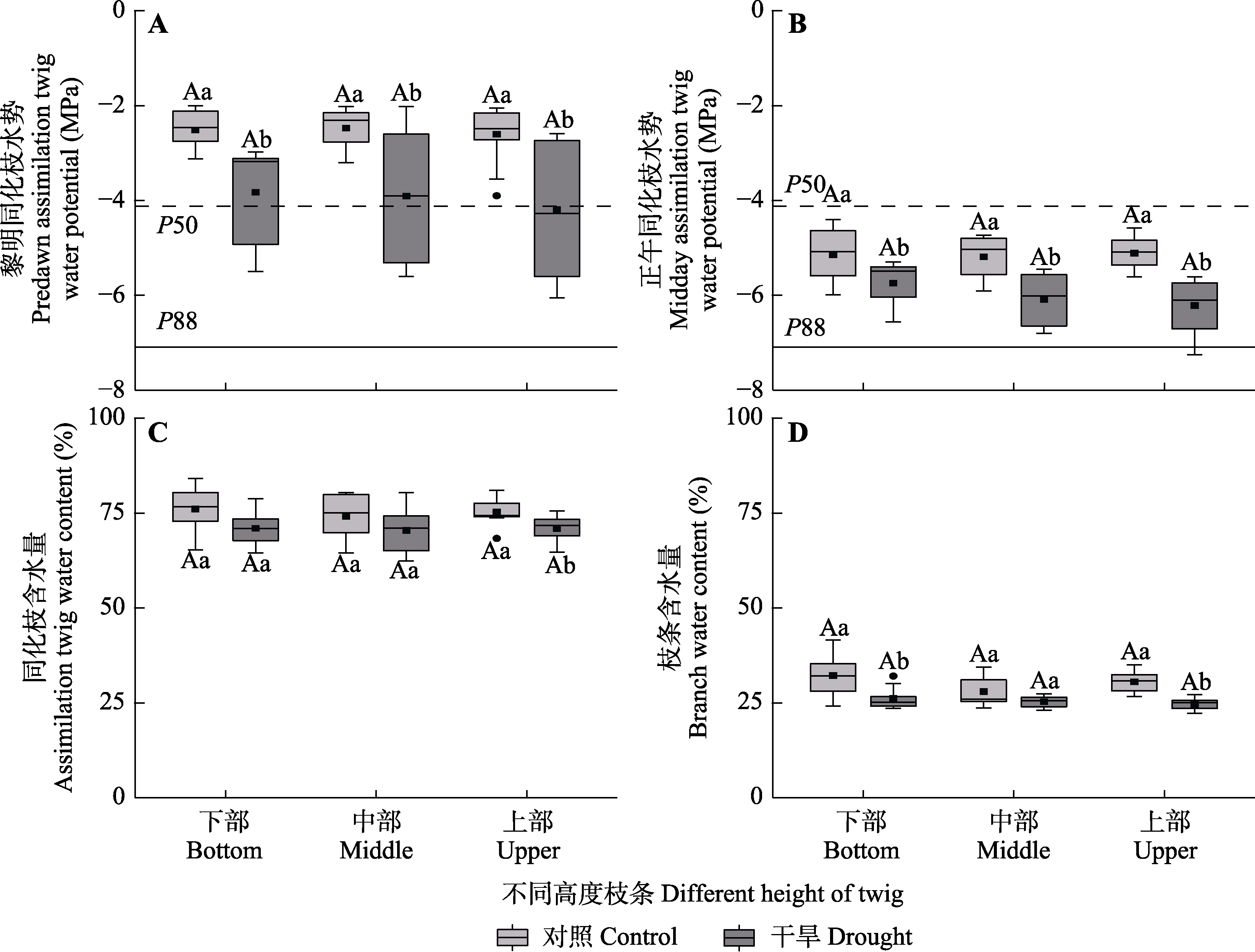
Fig. 3 Predawn assimilation twig water potential, midday assimilation twig water potential, twig water content and branch water content among different heights of Haloxylon ammodendron in control and drought treatment groups (mean ± SE). P50 and P88 are the xylem water potentials for 50% and 88% loss of maximum hydraulic conductivity, respectively. Different uppercase letters indicate significant differences among different heights of the same treatment (p < 0.05) and different lowercase letters indicate significant differences between different treatments of the same height (p < 0.05).
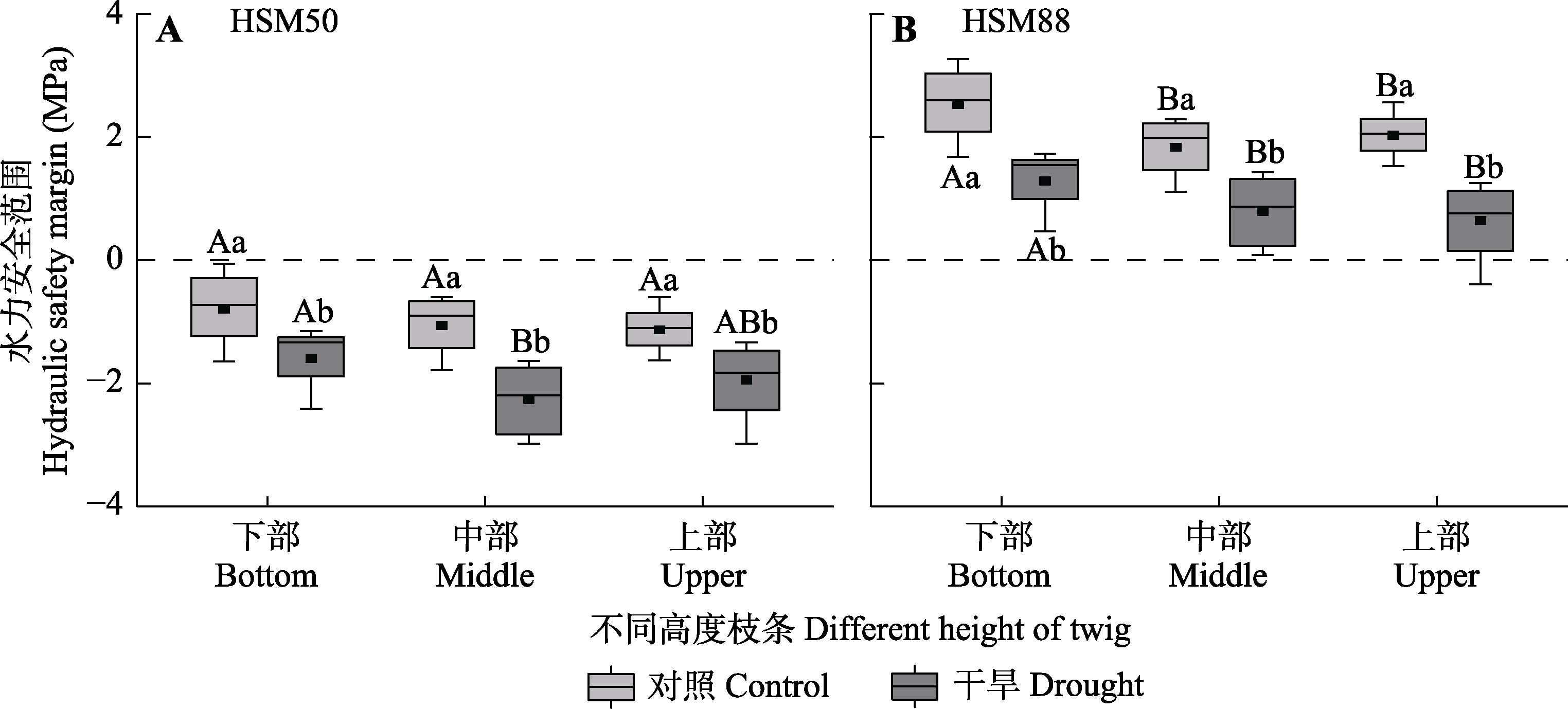
Fig. 4 Hydraulic safety margin among different heights of Haloxylon ammodendron in control and drought treatment groups (mean ± SE). HSM50 and HSM88 are the difference between the midday assimilation twig water potential and the xylem water potential for 50% and 88% loss of maximum hydraulic conductivity, respectively. Different uppercase letters indicate significant differences among different heights of the same treatment (p < 0.05) and different lowercase letters indicate significant differences between different treatments of the same height (p < 0.05).
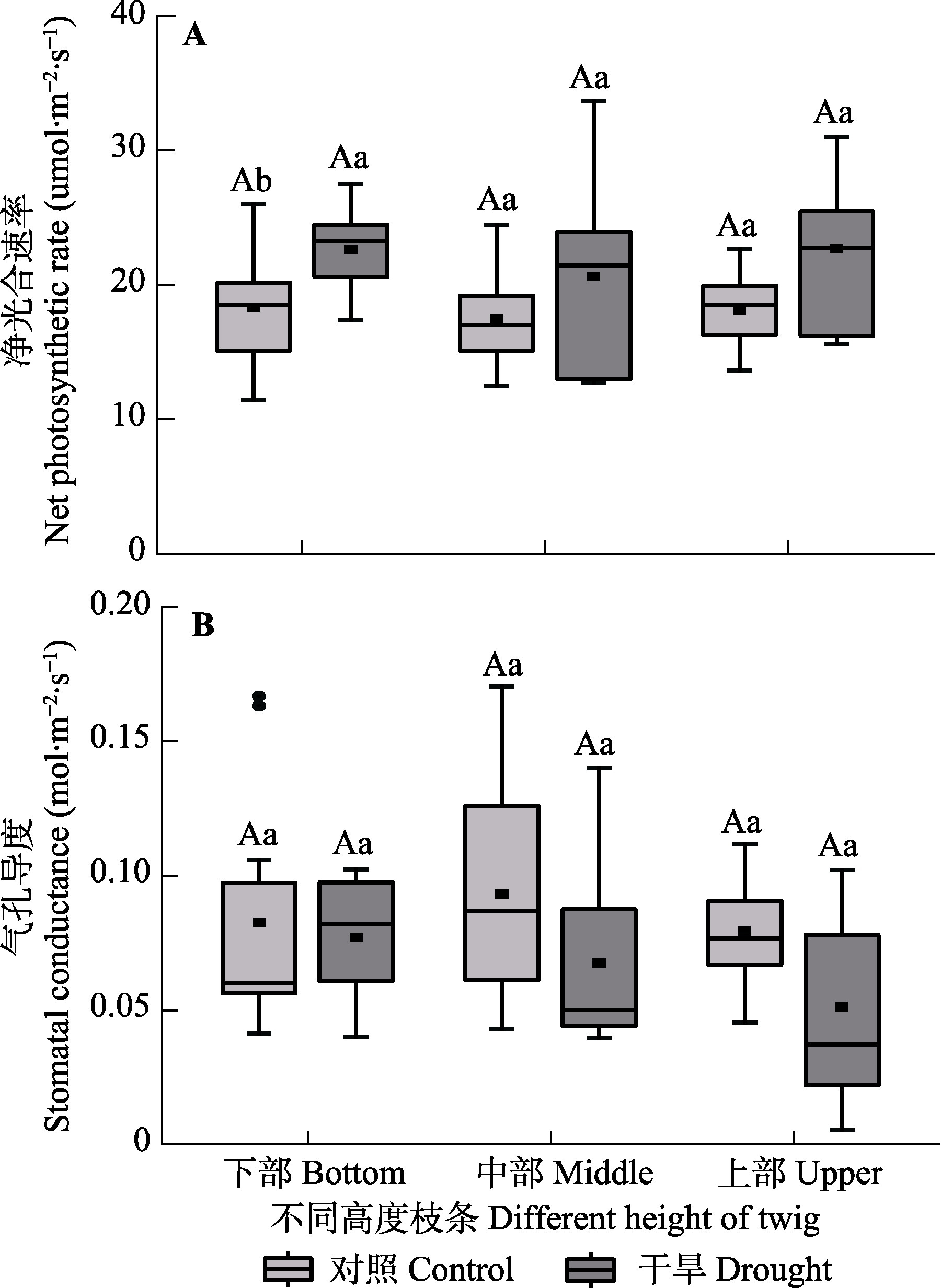
Fig. 5 Net photosynthetic rate and stomatal conductance among different heights of Haloxylon ammodendron in control and drought treatment groups (mean ± SE). Different uppercase letters indicate significant differences among different heights of the same treatment (p < 0.05) and different lowercase letters indicate significant differences between different treatments of the same height (p < 0.05).
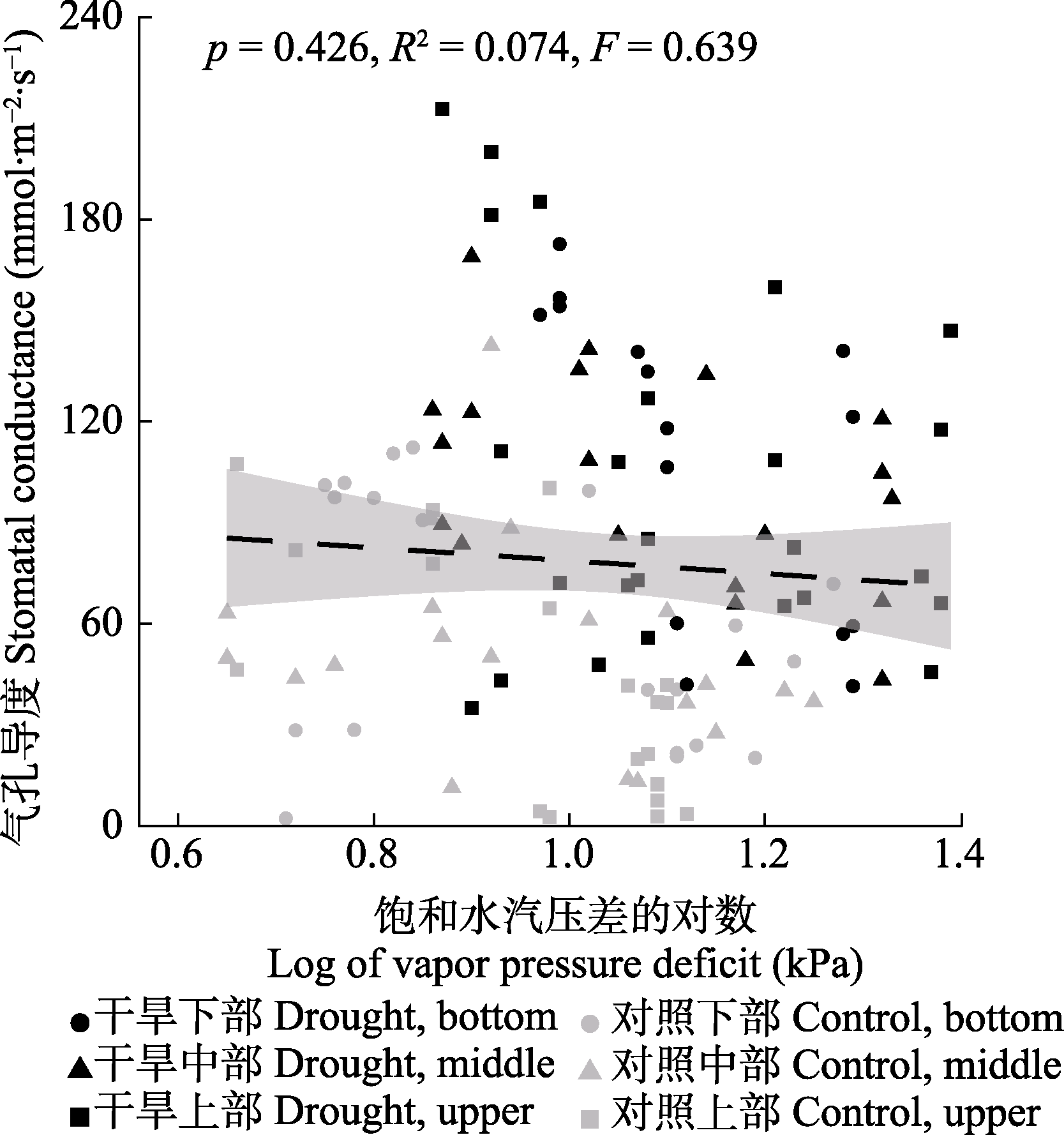
Fig. 6 Response of Haloxylon ammodendron stomatal conductance to vapor pressure deficit. The shaded area are 95% confidence intervals.
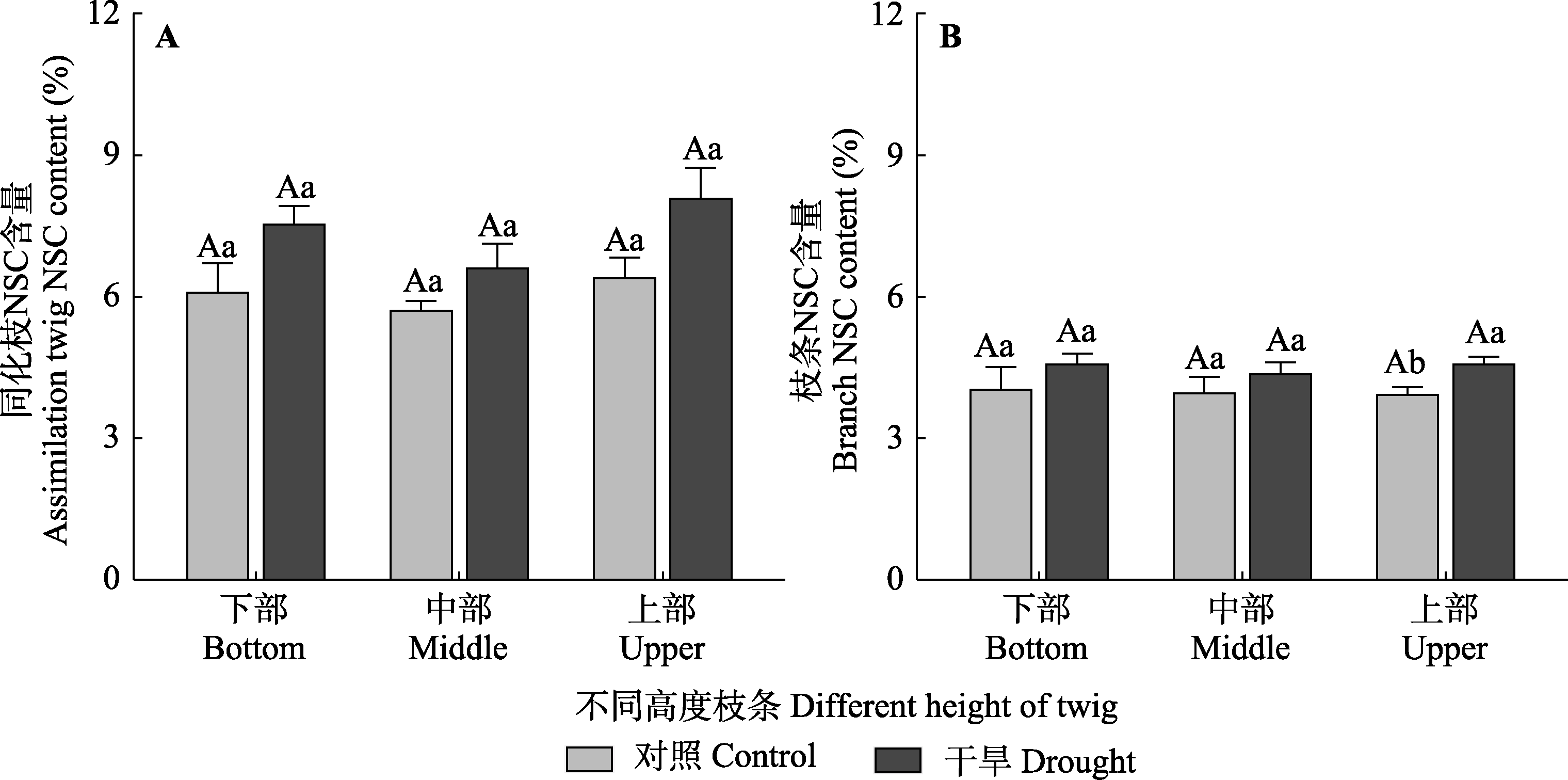
Fig. 7 Nonstructural carbohydrate (NSC) content of assimilation twigs and branches among different heights of Haloxylon ammodendron in control and drought treatment groups (mean ± SE). Different uppercase letters indicate significant differences among different heights of the same treatment (p < 0.05) and different lowercase letters indicate significant differences between different treatments of the same height (p < 0.05).
| 处理 Treatment | 高度 Height | 同化枝 Assiassimilation twig | 枝条 Branch | ||
|---|---|---|---|---|---|
| 可溶性糖 Soluble sugar (%) | 淀粉 Starch (%) | 可溶性糖 Soluble sugar (%) | 淀粉 Starch (%) | ||
| 对照 Control | 下部 Bottom | 2.48 ± 0.45Ab | 3.61 ± 0.62Aa | 2.62 ± 0.70Aa | 1.41 ± 0.17Aa |
| 中部 Middle | 2.57 ± 0.14Ab | 3.13 ± 0.30Aa | 2.21 ± 0.39Ab | 1.75 ± 0.28Aa | |
| 上部 Upper | 2.83 ± 0.24Ab | 3.57 ± 0.59Aa | 2.22 ± 0.20Ab | 1.71 ± 0.29Aa | |
| 干旱 Drought | 下部 Bottom | 3.81 ± 0.37Aa | 3.72 ± 0.56Aa | 3.01 ± 0.27Aa | 1.56 ± 0.07Aa |
| 中部 Middle | 3.48 ± 0.58Aa | 3.12 ± 0.59Aa | 3.01 ± 0.14Aa | 1.34 ± 0.33Aa | |
| 上部 Upper | 4.43 ± 0.49Aa | 3.65 ± 0.71Aa | 3.03 ± 0.19Aa | 1.53 ± 0.13Aa | |
Table 2 Soluble sugar and starch contents of assimilation twigs and branches among different heights of Haloxylon ammodendron in control and drought treatment groups (mean ± SE)
| 处理 Treatment | 高度 Height | 同化枝 Assiassimilation twig | 枝条 Branch | ||
|---|---|---|---|---|---|
| 可溶性糖 Soluble sugar (%) | 淀粉 Starch (%) | 可溶性糖 Soluble sugar (%) | 淀粉 Starch (%) | ||
| 对照 Control | 下部 Bottom | 2.48 ± 0.45Ab | 3.61 ± 0.62Aa | 2.62 ± 0.70Aa | 1.41 ± 0.17Aa |
| 中部 Middle | 2.57 ± 0.14Ab | 3.13 ± 0.30Aa | 2.21 ± 0.39Ab | 1.75 ± 0.28Aa | |
| 上部 Upper | 2.83 ± 0.24Ab | 3.57 ± 0.59Aa | 2.22 ± 0.20Ab | 1.71 ± 0.29Aa | |
| 干旱 Drought | 下部 Bottom | 3.81 ± 0.37Aa | 3.72 ± 0.56Aa | 3.01 ± 0.27Aa | 1.56 ± 0.07Aa |
| 中部 Middle | 3.48 ± 0.58Aa | 3.12 ± 0.59Aa | 3.01 ± 0.14Aa | 1.34 ± 0.33Aa | |
| 上部 Upper | 4.43 ± 0.49Aa | 3.65 ± 0.71Aa | 3.03 ± 0.19Aa | 1.53 ± 0.13Aa | |
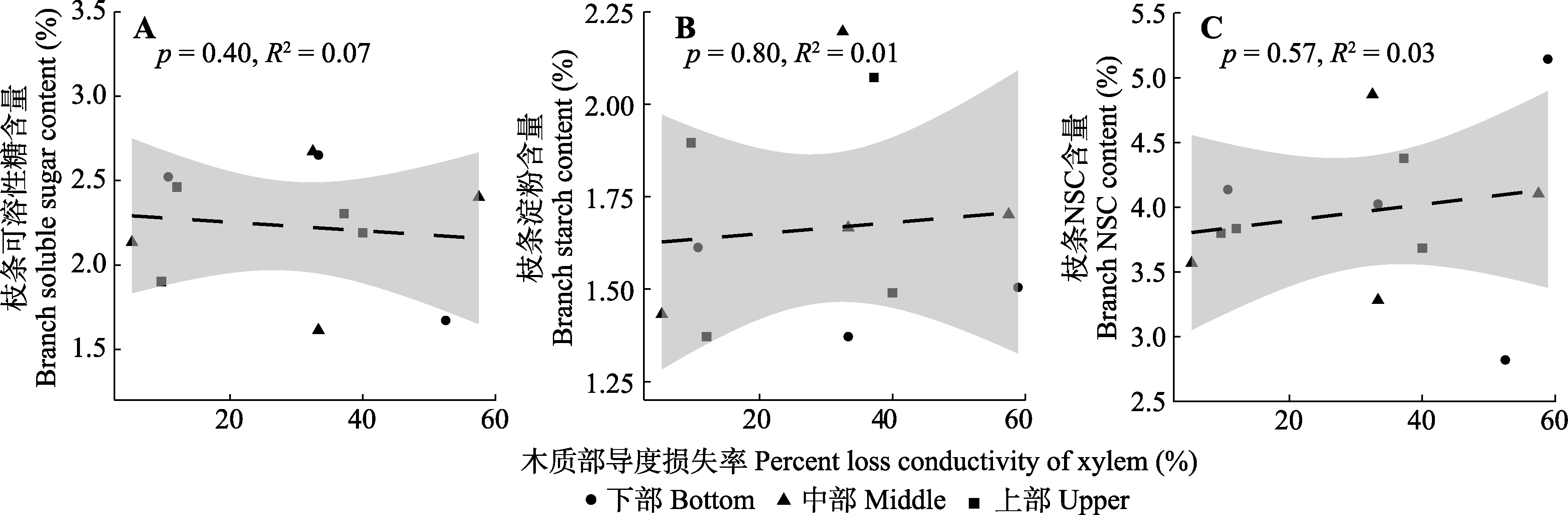
Fig. 8 Relationship between branch xylem hydraulic conductivity loss rate and nonstructural carbohydrate (NSC) content of Haloxylon ammodendron in drought treatment group.
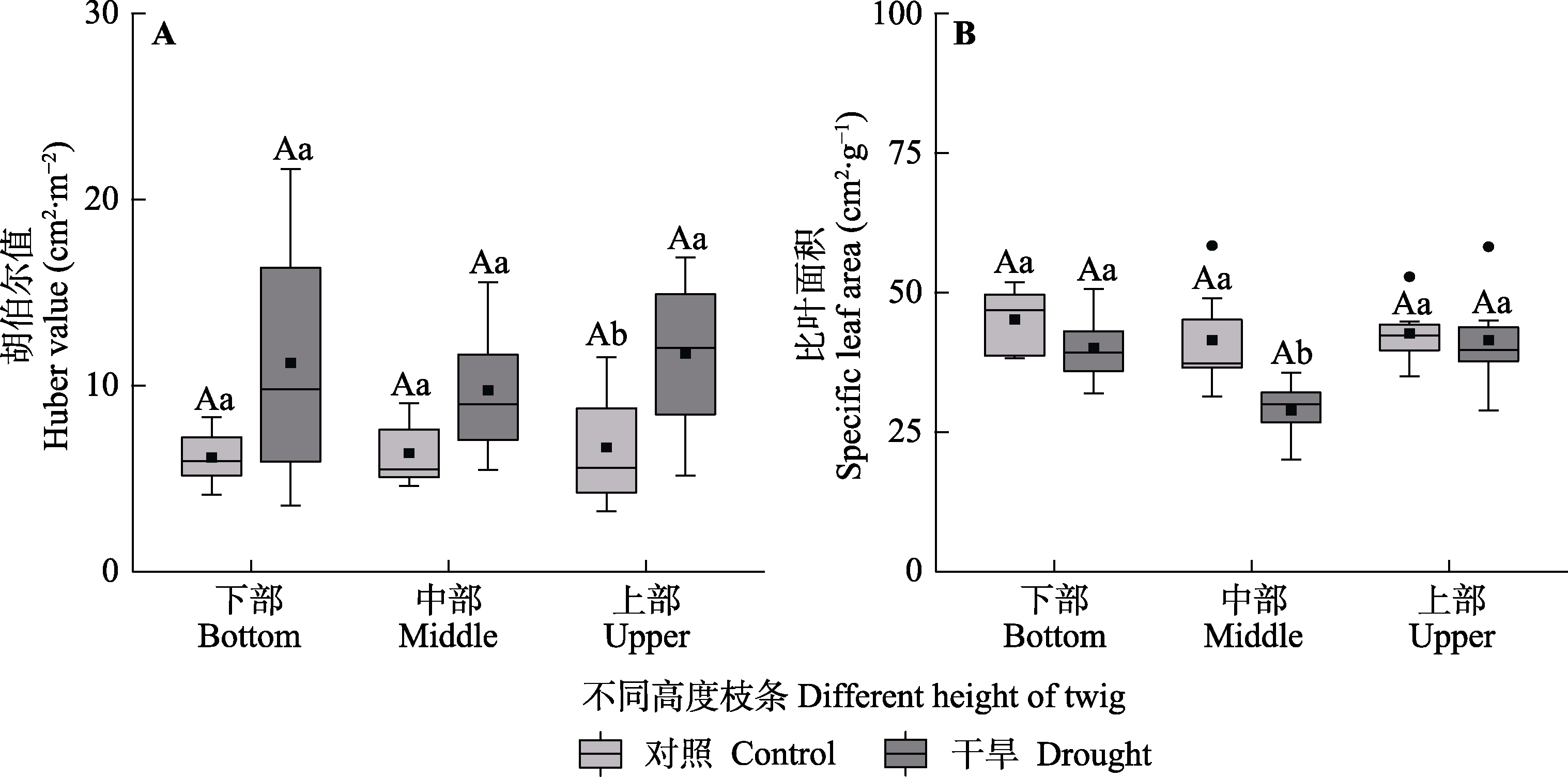
Fig. 9 Huber value and specific leaf area among different heights of Haloxylon ammodendron in control and drought treatment groups (mean ± SE). Different uppercase letters indicate significant differences among different heights of the same treatment (p < 0.05) and different lowercase letters indicate significant differences between different treatments of the same height (p < 0.05).
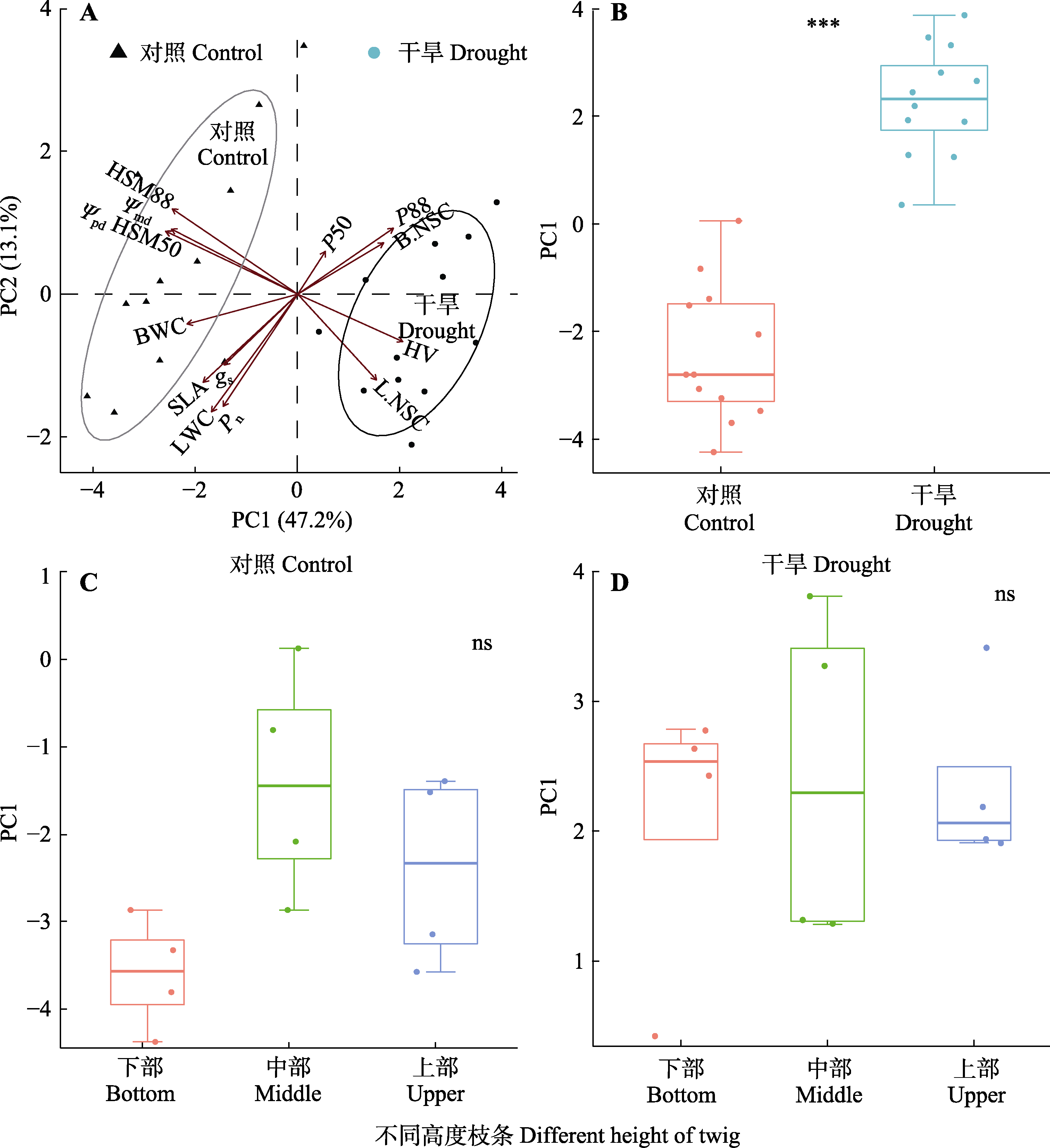
Fig. 10 Principal component (PC) analysis of each trait of drought resistance strategies for Haloxylon ammodendron. ns, no significant difference; ***, p < 0.001. Ψmd, midday assimilation twig water potential; Ψpd, predawn assimilation twig water potential; B.NSC, nonstructural carbohydrate content of branch; BWC, branch water content; gs, stomatal conductance; HSM50, Ψmd - P50; HSM88, Ψmd - P88; HV, Huber value; L.NSC, nonstructural carbohydrate content of assimilation twigs; LWC, assimilation twig water content; P50, xylem water potentials for 50% loss of maximum hydraulic conductivity; P88, xylem water potentials for 88% loss of maximum hydraulic conductivity; Pn, net photosynthetic rate; SLA, specific leaf area.
| [1] |
Allen CD, Macalady AK, Chenchouni H, Bachelet D, McDowell N, Vennetier M, Kitzberger T, Rigling A, Breshears DD, Hogg EH, Gonzalez P, Fensham R, Zhang Z, Castro J, Demidova N, et al. (2010). A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. Forest Ecology and Management, 259, 660-684.
DOI URL |
| [2] |
Anderegg WRL, Berry JA, Smith DD, Sperry JS, Anderegg LDL, Field CB (2012). The roles of hydraulic and carbon stress in a widespread climate-induced forest die-off. Proceedings of the National Academy of Sciences of the United States of America, 109, 233-237.
DOI PMID |
| [3] |
Anderegg WRL, Klein T, Bartlett M, Sack L, Pellegrini AFA, Choat B, Jansen S (2016). Meta-analysis reveals that hydraulic traits explain cross-species patterns of droughtinduced tree mortality across the globe. Proceedings of the National Academy of Sciences of the United States of America, 113, 5024-5029.
DOI PMID |
| [4] |
Anderegg WRL, Konings AG, Trugman AT, Yu KL, Bowling DR, Gabbitas R, Karp DS, Pacala S, Sperry JS, Sulman BN, Zenes N (2018). Hydraulic diversity of forests regulates ecosystem resilience during drought. Nature, 561, 538-541.
DOI |
| [5] | Anest A, Charles-Dominique T, Maurin O, Millan M, Edelin C, Tomlinson KW (2021). Evolving the structure: climatic and developmental constraints on the evolution of plant architecture. A case study in Euphorbia. New Phytologist, 231, 1278-1295. |
| [6] | Beikircher B, Mayr S (2008). The hydraulic architecture of Juniperus communis L. ssp. communis: shrubs and trees compared. Plant, Cell & Environment, 31, 1545-1556. |
| [7] |
Bond WJ, Midgley JJ (2001). Ecology of sprouting in woody plants: the persistence niche. Trends in Ecology & Evolution, 16, 45-51.
DOI URL |
| [8] |
Breshears DD, Myers OB, Meyer CW, Barnes FJ, Zou CB, Allen CD, McDowell NG, Pockman WT (2009). Tree die-off in response to global change-type drought: mortality insights from a decade of plant water potential measurements. Frontiers in Ecology and the Environment, 7, 185-189.
DOI URL |
| [9] |
Brodribb TJ, Cochard H (2009). Hydraulic failure defines the recovery and point of death in water-stressed conifers. Plant Physiology, 149, 575-584.
DOI PMID |
| [10] | Brodribb TJ, Feild TS (2000). Stem hydraulic supply is linked to leaf photosynthetic capacity: evidence from New Caledonian and Tasmanian rainforests. Plant, Cell & Environment, 23, 1381-1388. |
| [11] |
Carter JL, White DA (2009). Plasticity in the Huber value contributes to homeostasis in leaf water relations of a mallee Eucalypt with variation to groundwater depth. Tree Physiology, 29, 1407-1418.
DOI PMID |
| [12] |
Chaves MM, Pereira JS, Maroco J, Rodrigues ML, Ricardo CP, Osório ML, Carvalho I, Faria T, Pinheiro C (2002). How plants cope with water stress in the field. Photosynthesis and growth. Annals of Botany, 89, 907-916.
DOI URL |
| [13] |
Chen YJ, Choat B, Sterck F, Maenpuen P, Katabuchi M, Zhang SB, Tomlinson KW, Oliveira RS, Zhang YJ, Shen JX, Cao KF, Jansen S (2021). Hydraulic prediction of drought-induced plant dieback and top-kill depends on leaf habit and growth form. Ecology Letters, 24, 2350-2363.
DOI URL |
| [14] |
Choat B, Jansen S, Brodribb TJ, Cochard H, Delzon S, Bhaskar R, Bucci SJ, Feild TS, Gleason SM, Hacke UG, Jacobsen AL, Lens F, Maherali H, Martínez-Vilalta J, Mayr S, et al. (2012). Global convergence in the vulnerability of forests to drought. Nature, 491, 752-755.
DOI |
| [15] |
Cooper DJ, D’Amico DR, Scott ML (2003). Physiological and morphological response patterns of Populus deltoides to alluvial groundwater pumping. Environmental Management, 31, 215-226.
PMID |
| [16] |
Dai AG (2013). Increasing drought under global warming in observations and models. Nature Climate Change, 3, 52-58.
DOI |
| [17] |
Dai Y, Zheng XJ, Tang LS, Li Y (2015). Stable oxygen isotopes reveal distinct water use patterns of two Haloxylon species in the Gurbantonggut Desert. Plant and Soil, 389, 73-87.
DOI URL |
| [18] |
Davis SD, Ewers FW, Sperry JS, Portwood KA, Crocker MC, Adams GC (2002). Shoot dieback during prolonged drought in Ceanothus (Rhamnaceae) chaparral of California: a possible case of hydraulic failure. American Journal of Botany, 89, 820-828.
DOI URL |
| [19] | Duursma R, Choat B (2017). fitplc-an R package to fit hydraulic vulnerability curves. Journal of Plant Hydraulics, 4, e002. DOI: 10.20870/jph.2017.e002. |
| [20] | Fang LD, Ning QR, Guo JJ, Gong XW, Zhu JJ, Hao GY (2021). Hydraulic limitation underlies the dieback of Populus pseudosimonii trees in water-limited areas of Northern China. Forest Ecology and Management, 483, 118764. DOI: 10.1016/j.foreco.2020.118764. |
| [21] |
Fu PL, Jiang YJ, Wang AY, Brodribb TJ, Zhang JL, Zhu SD, Cao KF (2012). Stem hydraulic traits and leaf water-stress tolerance are co-ordinated with the leaf phenology of angiosperm trees in an Asian tropical dry karst forest. Annals of Botany, 110, 189-199.
DOI URL |
| [22] | Garcia-Forner N, Adams HD, Sevanto S, Collins AD, Dickman LT, Hudson PJ, Zeppel MJB, Jenkins M, Powers H, Martinez-Vilalta J, McDowell NG (2016). Responses of two semiarid conifer tree species to reduced precipitation and warming reveal new perspectives for stomatal regulation. Plant, Cell & Environment, 39, 38-49. |
| [23] |
Giorgi F, Coppola E, Raffaele F (2014). A consistent picture of the hydroclimatic response to global warming from multiple indices: models and observations. Journal of Geophysical Research: Atmospheres, 119, 11695-11708.
DOI URL |
| [24] |
Gong CM, Wang JJ, Hu CX, Wang JH, Ning PB, Bai J (2015). Interactive response of photosynthetic characteristics in Haloxylon ammodendron and Hedysarum scoparium exposed to soil water and air vapor pressure deficits. Journal of Environmental Sciences (China), 34, 184-196.
DOI URL |
| [25] |
Gong XW, Guo JJ, Fang LD, Bucci SJ, Goldstein G, Hao GY (2021). Hydraulic dysfunction due to root-exposure- initiated water stress is responsible for the mortality of Salix gordejevii shrubs on the windward slopes of active sand dunes. Plant and Soil, 459, 185-201.
DOI |
| [26] |
Grossiord C, Sevanto S, Dawson TE, Adams HD, Collins AD, Dickman LT, Newman BD, Stockton EA, McDowell NG (2017). Warming combined with more extreme precipitation regimes modifies the water sources used by trees. New Phytologist, 213, 584-596.
DOI PMID |
| [27] |
Hartmann H, Moura CF, Anderegg WRL, Ruehr NK, Salmon Y, Allen CD, Arndt SK, Breshears DD, Davi H, Galbraith D, Ruthrof KX, Wunder J, Adams HD, Bloemen J, Cailleret M, et al. (2018). Research frontiers for improving our understanding of drought induced tree and forest mortality. New Phytologist, 218, 15-28.
DOI PMID |
| [28] |
Hartmann H, Trumbore S (2016). Understanding the roles of nonstructural carbohydrates in forest trees-from what we can measure to what we want to know. New Phytologist, 211, 386-403.
DOI PMID |
| [29] |
Hochberg U, Windt CW, Ponomarenko A, Zhang YJ, Gersony J, Rockwell FE, Holbrook NM (2017). Stomatal closure, basal leaf embolism, and shedding protect the hydraulic integrity of grape stems. Plant Physiology, 174, 764-775.
DOI PMID |
| [30] |
Hoover DL, Duniway MC, Belnap J (2015). Pulse-drought atop press-drought: unexpected plant responses and implications for dryland ecosystems. Oecologia, 179, 1211-1221.
DOI PMID |
| [31] |
Huang JP, Ma JR, Guan XD, Li Y, He YL (2019). Progress in semi-arid climate change studies in China. Advances in Atmospheric Sciences, 36, 922-937.
DOI |
| [32] |
Huang JP, Yu HP, Guan XD, Wang GY, Guo RX (2016). Accelerated dryland expansion under climate change. Nature Climate Change, 6, 166-171.
DOI |
| [33] | Johnson DM, McCulloh KA, Meinzer FC, Woodruff DR, Eissenstat DM, Phillips N (2011). Hydraulic patterns and safety margins, from stem to stomata, in three eastern US tree species. Tree Physiology, 31, 659-668. |
| [34] |
Koch GW, Sillett SC, Jennings GM, Davis SD (2004). The limits to tree height. Nature, 428, 851-854.
DOI |
| [35] |
Li BF, Chen YN, Shi X, Chen ZS, Li WH (2013). Temperature and precipitation changes in different environments in the arid region of northwest China. Theoretical and Applied Climatology, 112, 589-596.
DOI URL |
| [36] |
Li JY, Chang H, Liu T, Zhang C (2019). The potential geographical distribution of Haloxylon across Central Asia under climate change in the 21st century. Agricultural and Forest Meteorology, 275, 243-254.
DOI URL |
| [37] | Li Y, Zheng XJ, Wang YG, Xu GQ, Liu R (2021). Experiment and simulation platform for oasis-desert symbiotic relationship (ODP). Bulletin of Chinese Academy of Sciences, 36, 1506-1514. |
| [李彦, 郑新军, 王玉刚, 徐贵青, 刘冉 (2021). 绿洲-荒漠共生关系实验模拟平台(绿洲-荒漠平台). 中国科学院院刊, 36, 1506-1514.] | |
| [38] | Liu H, Gleason SM, Hao GY, Hua L, He PC, Goldstein G, Ye Q (2019). Hydraulic traits are coordinated with maximum plant height at the global scale. Science Advances, 5, eaav1332. DOI: 10.1126/sciadv.aav1332. |
| [39] |
López R, Cano FJ, Martin-StPaul NK, Cochard H, Choat B (2021). Coordination of stem and leaf traits define different strategies to regulate water loss and tolerance ranges to aridity. New Phytologist, 230, 497-509.
DOI PMID |
| [40] |
Lü XP, Gao HJ, Zhang L, Wang YP, Shao KZ, Zhao Q, Zhang JL (2019). Dynamic responses of Haloxylon ammodendron to various degrees of simulated drought stress. Plant Physiology and Biochemistry, 139, 121-131.
DOI URL |
| [41] | Martínez-Vilalta J, Garcia-Forner N (2017). Water potential regulation, stomatal behaviour and hydraulic transport under drought: deconstructing the iso/anisohydric concept. Plant, Cell & Environment, 40, 962-976. |
| [42] |
Martínez-Vilalta J, Sala AN, Asensio D, Galiano L, Hoch G, Palacio S, Piper FI, Lloret F (2016). Dynamics of non-structural carbohydrates in terrestrial plants: a global synthesis. Ecological Monographs, 86, 495-516.
DOI URL |
| [43] |
Mayr S, Beikircher B, Obkircher MA, Schmid P (2010). Hydraulic plasticity and limitations of alpine Rhododendron species. Oecologia, 164, 321-330.
DOI URL |
| [44] |
McDowell N, Pockman WT, Allen CD, Breshears DD, Cobb N, Kolb T, Plaut J, Sperry J, West A, Williams DG, Yepez EA (2008). Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? New Phytologist, 178, 719-739.
DOI PMID |
| [45] |
McDowell NG (2011). Mechanisms linking drought, hydraulics, carbon metabolism, and vegetation mortality. Plant Physiology, 155, 1051-1059.
DOI PMID |
| [46] |
Noy-Meir I (1973). Desert ecosystems: environment and producers. Annual Review of Ecology and Systematics, 4, 25-51.
DOI URL |
| [47] |
Powers JS, Vargas GG, Brodribb TJ, Schwartz NB, Pérez- Aviles D, Smith-Martin CM, Becknell JM, Aureli F, Blanco R, Calderón-Morales E, Calvo-Alvarado JC, Calvo- Obando AJ, Chavarría MM, Carvajal-Vanegas D, Jiménez-Rodríguez CD, et al. (2020). A catastrophic tropical drought kills hydraulically vulnerable tree species. Global Change Biology, 26, 3122-3133.
DOI PMID |
| [48] |
Rosner S, Heinze B, Savi T, Dalla-Salda G (2019). Prediction of hydraulic conductivity loss from relative water loss: new insights into water storage of tree stems and branches. Physiologia Plantarum, 165, 843-854.
DOI URL |
| [49] |
Rosner S, Klein A, Wimmer R, Karlsson B (2006). Extraction of features from ultrasound acoustic emissions: a tool to assess the hydraulic vulnerability of Norway spruce trunkwood? New Phytologist, 171, 105-116.
DOI PMID |
| [50] |
Sage RF, Sultmanis S (2016). Why are there no C4 forests? Journal of Plant Physiology, 203, 55-68.
DOI URL |
| [51] |
Schenk HJ, Espino S, Goedhart CM, Nordenstahl M, Cabrera HIM, Jones CS (2008). Hydraulic integration and shrub growth form linked across continental aridity gradients. Proceedings of the National Academy of Sciences of the United States of America, 105, 11248-11253.
DOI PMID |
| [52] |
Scholander PF, Bradstreet ED, Hemmingsen EA, Hammel HT (1965). Sap pressure in vascular plants. Science, 148, 339-346.
PMID |
| [53] | Sevanto S, McDowell NG, Dickman LT, Pangle R, Pockman WT (2014). How do trees Die? A test of the hydraulic failure and carbon starvation hypotheses. Plant, Cell & Environment, 37, 153-161. |
| [54] | Song CW, Li CJ, Halik Ü, Xu XW, Lei JQ, Zhou ZB, Fan JL (2021). Spatial distribution and structural characteristics for Haloxylon ammodendron plantation on the southwestern edge of the gurbantünggüt desert. Forests, 12, 633. DOI: 10.3390/f12050633. |
| [55] | Stovall AEL, Shugart H, Yang X (2019). Tree height explains mortality risk during an intense drought. Nature Communications, 10, 4385. DOI: 10.1038/s41467-019-12380-6. |
| [56] |
Tiemuerbieke B, Min XJ, Zang YX, Xing P, Ma JY, Sun W (2018). Water use patterns of co-occurring C3 and C4shrubs in the Gurbantonggut Desert in northwestern China. Science of the Total Environment, 634, 341-354.
DOI URL |
| [57] |
Trugman AT, Anderegg LDL, Anderegg WRL, Das AJ, Stephenson NL (2021). Why is tree drought mortality so hard to predict? Trends in Ecology & Evolution, 36, 520-532.
DOI URL |
| [58] |
Werner C, Correia O, Beyschlag W (1999). Two different strategies of Mediterranean macchia plants to avoid photoinhibitory damage by excessive radiation levels during summer drought. Acta Oecologica, 20, 15-23.
DOI URL |
| [59] |
Wu X, Zheng XJ, Li Y, Xu GQ (2019). Varying responses of two Haloxylon species to extreme drought and groundwater depth. Environmental and Experimental Botany, 158, 63-72.
DOI URL |
| [60] |
Xu GQ, Li Y (2008). Rooting depth and leaf hydraulic conductance in the xeric tree Haloxyolon ammodendron growing at sites of contrasting soil texture. Functional Plant Biology, 35, 1234-1242.
DOI PMID |
| [61] |
Xu GQ, McDowell NG, Li Y (2016). A possible link between life and death of a xeric tree in desert. Journal of Plant Physiology, 194, 35-44.
DOI URL |
| [62] |
Xu GQ, Yu DD, Li Y (2017). Patterns of biomass allocation in Haloxylon persicum woodlands and their understory herbaceous layer along a groundwater depth gradient. Forest Ecology and Management, 395, 37-47.
DOI URL |
| [63] | Yang XY, Lu MQ, Wang YF, Wang YR, Liu ZJ, Chen S (2021). Response mechanism of plants to drought stress. Horticulturae, 7, 50. DOI: 10.3390/horticulturae7030050. |
| [64] |
Yu X, Lei JQ, Gao X (2022). An over review of desertification in Xinjiang, Northwest China. Journal of Arid Land, 14, 1181-1195.
DOI |
| [65] |
Zhang HY, Wang CK, Wang XC (2014). Spatial variations in non-structural carbohydrates in stems of twelve temperate tree species. Trees, 28, 77-89.
DOI URL |
| [66] | Zhang L, Liu L, Zhao H, Jiang ZM, Cai J (2020). Differences in near isohydric and anisohydric behavior of contrasting poplar hybrids (I-101 (Populus alba L.) × 84K (Populus alba L. × Populus glandulosa Uyeki)) under drought- rehydration treatments. Forests, 11, 402. DOI: 10.3390/f11040402. |
| [67] | Zhang T, Cao Y, Chen YM, Liu GB (2015). Non-structural carbohydrate dynamics in Robinia pseudoacacia saplings under three levels of continuous drought stress. Trees, 29, 1837-1849. |
| [68] |
Zolfaghar S, Villalobos-Vega R, Zeppel M, Eamus D (2015). The hydraulic architecture of Eucalyptus trees growing across a gradient of depth-to-groundwater. Functional Plant Biology, 42, 888-898.
DOI PMID |
| [1] | ZHANG Bin, ZHANG Hao-Cheng, QIAO Tian, LÜ Zhi-Bing, XU Ya-Nan, LI Xue-Qin, YUAN Xiang-Yang, FENG Mei-Chen, ZHANG Mei-Jun. Effect of arbuscular mycorrhizal fungi inoculation on non-structural carbohydrates and C, N and P stoichiometry in oat plants under drought stress [J]. Chin J Plant Ecol, 2025, 49(7): 1082-1095. |
| [2] | SHAO Chang-Chang, DUAN Hong-Lang, ZHAO Xi-Zhou, DING Gui-Jie. Research progress on the prediction of drought death point and the mechanism of drought- induced tree mortality [J]. Chin J Plant Ecol, 2025, 49(2): 221-231. |
| [3] | ZHANG Wang, TAN Si-Yi, TU Wen-Qin, LOU An-Ru. Genetic diversity and genetic structure of Haloxylon ammodendron in the Turpan and Hami area, Xinjiang, China [J]. Chin J Plant Ecol, 2025, 49(11): 1858-1868. |
| [4] | HAN Yu-Qing, XIONG Wei, WU Bo, LU Qi, YANG Wen-Bin, LIU Ya-Li, ZHANG Jing-Bo, XIN Zhi-Ming, MA Ying-Bin, LIAN Hong-Lin, WANG Si-Han. Responses of stem sap flow of Haloxylon ammodendron to rainfall pulses in Ulan Buh Desert [J]. Chin J Plant Ecol, 2024, 48(9): 1172-1179. |
| [5] | OUYANG Yi-Lei, GONG Xue-Wei, DUAN Chun-Yang, ZHANG Chi, MA Chen-Yang, HAN Peng, ZHANG Yuan-Ming, HAO Guang-You. Water- and carbon-related physiological mechanisms underlying the decline of wild apricot trees in Ili, Xinjiang, China [J]. Chin J Plant Ecol, 2024, 48(9): 1192-1201. |
| [6] | LIAO Su-Hui, NI Long-Kang, QIN Jia-Shuang, TAN Yu, GU Da-Xing. Hydraulic regulation strategies of karst forest species exhibit variation across different successional stages in the mid-subtropical zone [J]. Chin J Plant Ecol, 2024, 48(9): 1223-1231. |
| [7] | LONG Ji-Lan, JIANG Zheng, LIU Ding-Qin, MIAO Yu-Xuan, ZHOU Ling-Yan, FENG Ying, PEI Jia-Ning, LIU Rui-Qiang, ZHOU Xu-Hui, FU Yu-Ling. Effects of drought on plant root exudates and associated rhizosphere priming effect: review and prospect [J]. Chin J Plant Ecol, 2024, 48(7): 817-827. |
| [8] | CHEN Yu-Ting, MA Song-Mei, ZHANG Dan, ZHANG Lin, WANG Chun-Cheng. Diversity pattern and formation mechanism of sympatric Haloxylon ammodendron and Haloxylon persicum in Xinjiang, China [J]. Chin J Plant Ecol, 2024, 48(1): 56-67. |
| [9] | ZHOU Jie, YANG Xiao-Dong, WANG Ya-Yun, LONG Yan-Xin, WANG Yan, LI Bo-Rui, SUN Qi-Xing, SUN Nan. Difference in adaptation strategy between Haloxylon ammodendron and Alhagi sparsifolia to drought [J]. Chin J Plant Ecol, 2022, 46(9): 1064-1076. |
| [10] | QIN Hui-Jun, JIAO Liang, ZHOU Yi, XUE Ru-Hong, QI Chang-Liang, DU Da-Shi. Effects of altitudes on non-structural carbohydrate allocation in different dominate trees in Qilian Mountains, China [J]. Chin J Plant Ecol, 2022, 46(2): 208-219. |
| [11] | REN Jin-Pei, LI Jun-Peng, WANG Wei-Feng, DAI Yong-Xin, WANG Lin. Responses of leaf hydraulic traits to water conditions in eight tree species and the driving factors [J]. Chin J Plant Ecol, 2021, 45(9): 942-951. |
| [12] | LUO Dan-Dan, WANG Chuan-Kuan, JIN Ying. Response mechanisms of hydraulic systems of woody plants to drought stress [J]. Chin J Plant Ecol, 2021, 45(9): 925-941. |
| [13] | LIU Li-Yan, FENG Jin-Xia, LIU Wen-Xin, WAN Xian-Chong. Effects of drought stress on photosynthesis, growth and root structure of transgenic PtPIP2;8 poplar 84K (Populus alba × P. glandulosa) [J]. Chin J Plant Ecol, 2020, 44(6): 677-686. |
| [14] | SONG Lin, LUO Wen-Tao, MA Wang, HE Peng, LIANG Xiao-Sa, WANG Zheng-Wen. Extreme drought effects on nonstructural carbohydrates of dominant plant species in a meadow grassland [J]. Chin J Plant Ecol, 2020, 44(6): 669-676. |
| [15] | LUO Jin-Huan, TAN Zhao-Yuan, CHEN Bin, CHEN Guang-Wu, JIANG Kai, HEI Qi-Fang, ZHANG Hui. Key characteristics for facilitating Leucaena leucocephala to successfully invade pioneer communities of tropical rain forests [J]. Chin J Plant Ecol, 2020, 44(12): 1215-1223. |
| Viewed | ||||||
|
Full text |
|
|||||
|
Abstract |
|
|||||
Copyright © 2026 Chinese Journal of Plant Ecology
Tel: 010-62836134, 62836138, E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
![]()