
植物生态学报 ›› 2018, Vol. 42 ›› Issue (6): 692-702.DOI: 10.17521/cjpe.2017.0247
所属专题: 凋落物
• 研究论文 • 上一篇
魏翠翠,刘小飞,林成芳( ),李先锋,李艳,郑裕雄
),李先锋,李艳,郑裕雄
收稿日期:2017-09-28
修回日期:2018-03-15
出版日期:2018-06-20
发布日期:2018-06-20
通讯作者:
林成芳
基金资助:
WEI Cui-Cui,LIU Xiao-Fei,LIN Cheng-Fang(),LI Xian-Feng,LI Yan,ZHENG Yu-Xiong
Received:2017-09-28
Revised:2018-03-15
Online:2018-06-20
Published:2018-06-20
Contact:
Cheng-Fang LIN
Supported by:摘要:
酶在土壤有机质分解中起重要作用。为深入了解全球变化背景下森林凋落物产量的改变对森林生态系统过程的影响, 以亚热带米槠(Castanopsis carlesii)人促更新次生林(米槠人促林)和米槠次生林为研究对象, 设置凋落物加倍(DL)、凋落物去除(NL)和对照(CT) 3种处理, 探讨土壤6种胞外酶活性的变化。研究结果表明: 米槠次生林中土壤纤维素水解酶(CBH)、β-N-乙酰氨基葡萄糖苷酶(NAG)、酚氧化酶(PhOx)和过氧化酶(PerOx)活性高于米槠人促林, 而酸性磷酸酶(AP)和β-葡萄糖苷酶(βG)活性没有差异; NL和DL处理均降低了两种不同更新方式森林土壤的AP、βG和NAG活性, CBH和PerOx活性均无显著变化, 而PhOx活性仅在DL处理后降低; 除NAG活性外, 米槠人促林的AP、βG、PhOx活性在凋落物处理后下降的幅度均高于次生林; Pearson相关分析和冗余分析表明, 土壤酶活性与土壤含水量、碳(C)、氮(N)含量和微生物生物量碳(MBC)、氮(MBN)含量显著相关。因此, 凋落物输入的改变(无论增加和减少), 引起了土壤含水量、C、N以及MBC和MBN含量的下降, 进而可能会导致亚热带米槠次生林和米槠人促林土壤某些胞外酶(如AP、βG和NAG)活性降低。从土壤酶活性角度看, 米槠次生林比米槠人促林更有利于亚热带森林生态系统C、N养分循环。
魏翠翠, 刘小飞, 林成芳, 李先锋, 李艳, 郑裕雄. 凋落物输入改变对亚热带两种米槠次生林土壤酶活性的影响. 植物生态学报, 2018, 42(6): 692-702. DOI: 10.17521/cjpe.2017.0247
WEI Cui-Cui, LIU Xiao-Fei, LIN Cheng-Fang, LI Xian-Feng, LI Yan, ZHENG Yu-Xiong. Response of soil enzyme activities to litter input changes in two secondary Castanopsis carlessii forests in subtropical China. Chinese Journal of Plant Ecology, 2018, 42(6): 692-702. DOI: 10.17521/cjpe.2017.0247
| 项目 Item | 米槠次生林 Secondary Castanopsis carlesii forest | 米槠人促更新次生林 Human-assisted naturally regenerated Castanopsis carlesii forest |
|---|---|---|
| 平均树高 Average tree height (m) | 10.80 | 13.70 |
| 平均胸径 Average breast diameter (cm) | 12.20 | 16.80 |
| 林分密度 Stand density (株?hm-2) | 3788 | 2158 |
| 年凋落物量 Annual litter fall (g?m-2) | 698 | 658 |
| 细根生物量 Fine root biomass (kg?m-3) | 0.93 | 0.70 |
| 全C含量 Total carbon content (g?kg-1) | 20.98 | 16.68 |
| 全N含量 Total nitrogen content (g?kg-1) | 1.42 | 1.14 |
| 土壤容重 Soil bulk density (g?cm-3) | 0.95 | 1.10 |
表1 试验地林分基本特征和土壤表层(0-10 cm)理化性质
Table 1 Main characteristics of the experiment site and properties of the topsoil (0-10 cm)
| 项目 Item | 米槠次生林 Secondary Castanopsis carlesii forest | 米槠人促更新次生林 Human-assisted naturally regenerated Castanopsis carlesii forest |
|---|---|---|
| 平均树高 Average tree height (m) | 10.80 | 13.70 |
| 平均胸径 Average breast diameter (cm) | 12.20 | 16.80 |
| 林分密度 Stand density (株?hm-2) | 3788 | 2158 |
| 年凋落物量 Annual litter fall (g?m-2) | 698 | 658 |
| 细根生物量 Fine root biomass (kg?m-3) | 0.93 | 0.70 |
| 全C含量 Total carbon content (g?kg-1) | 20.98 | 16.68 |
| 全N含量 Total nitrogen content (g?kg-1) | 1.42 | 1.14 |
| 土壤容重 Soil bulk density (g?cm-3) | 0.95 | 1.10 |
| 酶 Enzyme | 缩写 Abbreviation | 编号 Code | 底物 Substrate | 类型 Type |
|---|---|---|---|---|
| 酸性磷酸酶 Acid phosphatase | AP | 3.1.3.2 | 4-甲基伞形酮磷酸酯 4-MUB-phosphate | P获得水解酶 P-targeting hydrolytic |
| β-葡萄糖苷酶 β-1,4-glucosidase | βG | 3.2.1.21 | 4-甲基伞形酮-β-D-葡萄糖苷 4-MUB-β-D-glucoside | C获得水解酶 C-targeting hydrolytic |
| 纤维素水解酶 Cellobiohydrolase | CBH | 3.2.1.91 | 4-甲基伞形酮-β-D-纤维素二糖苷 4-MUB-β-D-cellobioside | C获得水解酶 C-targeting hydrolytic |
| β-N-乙酰氨基葡萄糖苷酶 β-1,4-N-acetylglucosaminidase | NAG | 3.1.6.1 | 4-甲基伞形酮-2-乙酰氨基-2-脱氧-β-D-吡喃葡萄糖苷 4-MUB-N-acetyl-β-D-lucosaminide | N获得水解酶 N-targeting hydrolytic |
| 酚氧化酶 Phenol oxidase | PhOx | 1.10.3.2 | L-二羟基苯 L-DOPA | C获得氧化酶 C-targeting oxidase |
| 过氧化物酶 Peroxidase | PerOx | 1.11.1.7 | L-二羟基苯 L-DOPA | C获得氧化酶 C-targeting oxidase |
表2 土壤酶的种类、底物、缩写和类型
Table 2 The substrates, abbreviations and types of the examined soil enzymes
| 酶 Enzyme | 缩写 Abbreviation | 编号 Code | 底物 Substrate | 类型 Type |
|---|---|---|---|---|
| 酸性磷酸酶 Acid phosphatase | AP | 3.1.3.2 | 4-甲基伞形酮磷酸酯 4-MUB-phosphate | P获得水解酶 P-targeting hydrolytic |
| β-葡萄糖苷酶 β-1,4-glucosidase | βG | 3.2.1.21 | 4-甲基伞形酮-β-D-葡萄糖苷 4-MUB-β-D-glucoside | C获得水解酶 C-targeting hydrolytic |
| 纤维素水解酶 Cellobiohydrolase | CBH | 3.2.1.91 | 4-甲基伞形酮-β-D-纤维素二糖苷 4-MUB-β-D-cellobioside | C获得水解酶 C-targeting hydrolytic |
| β-N-乙酰氨基葡萄糖苷酶 β-1,4-N-acetylglucosaminidase | NAG | 3.1.6.1 | 4-甲基伞形酮-2-乙酰氨基-2-脱氧-β-D-吡喃葡萄糖苷 4-MUB-N-acetyl-β-D-lucosaminide | N获得水解酶 N-targeting hydrolytic |
| 酚氧化酶 Phenol oxidase | PhOx | 1.10.3.2 | L-二羟基苯 L-DOPA | C获得氧化酶 C-targeting oxidase |
| 过氧化物酶 Peroxidase | PerOx | 1.11.1.7 | L-二羟基苯 L-DOPA | C获得氧化酶 C-targeting oxidase |
| 项目 Item | 米槠次生林 Secondary Castanopsis carlesii forest | 米槠人促更新次生林 Human-assisted naturally regenerated Castanopsis carlesii forest | ||||
|---|---|---|---|---|---|---|
| 对照 Control | 凋落物去除 No litter | 凋落物添加 Double litter | 对照 Control | 凋落物去除 No litter | 凋落物添加 Double litter | |
| 含水量 Moisture content (%) | 0.30 ± 0.00aA | 0.23 ± 0.01bA | 0.29 ± 0.03abA | 0.26 ± 0.06aA | 0.21 ± 0.01aA | 0.25 ± 0.03aA |
| pH值 pH value | 5.11 ± 0.14aA | 5.10 ± 0.04aA | 4.91 ± 0.16aA | 5.25 ± 0.17aA | 5.11 ± 0.02aA | 5.13 ± 0.10aA |
| 全碳 Total carbon (g·kg-1) | 20.98 ± 2.75aA | 12.41 ± 1.75bA | 16.49 ± 0.47bB | 16.68 ± 0.44aB | 12.77 ± 0.09bA | 14.71 ± 3.00abA |
| 全氮 Total nitrogen (g·kg-1) | 1.42 ± 0.09aA | 0.93 ± 0.12cA | 1.14 ± 0.02bB | 1.14 ± 0.09aB | 0.88 ± 0.07bA | 1.02 ± 0.19abA |
| 可溶性有机碳 Dissolved organic carbon (mg·kg-1) | 49.15 ± 8.37aA | 29.77 ± 9.46bA | 55.58 ± 8.7aA | 48.86 ± 6.76aA | 47.11 ± 10.09aA | 49.52 ± 6.46aA |
| 可溶性有机氮 Dissolved organic nitrogen (mg·kg-1) | 2.08 ± 0.02aA | 1.85 ± 0.09aA | 0.74 ± 0.15bB | 2.26 ± 0.36aA | 2.18 ± 0.43aA | 1.86 ± 0.44aA |
| NH4+ (mg·kg-1) | 37.49 ± 0.69bA | 26.5 ± 3.21cA | 52.6 ± 0.6aA | 26.99 ± 4.01bB | 22.04 ± 3.12aA | 29.63 ± 4.41aB |
| 矿质氮 Mineral nitrogen (mg·kg-1) | 38.08 ± 0.52bA | 27.22 ± 3.31cA | 53.05 ± 0.82aA | 27.96 ± 3.94aB | 22.37 ± 3.24bA | 30.98 ± 4.41aB |
| 微生物生物量碳 Microbial biomass carbon (mg·kg-1) | 315 ± 33abA | 225 ± 66bA | 371 ± 57aA | 258 ± 17aB | 171 ± 5cA | 221 ± 23bB |
| 微生物生物量氮 Microbial biomass nitrogen (mg·kg-1) | 29.33 ± 6.30aA | 13.41 ± 2.27bA | 26.38 ± 5.36aA | 36.56 ± 3.49aA | 14.61 ± 2.58cA | 23.05 ± 5.07bA |
表3 两种米槠林添加和去除凋落物处理土壤理化性质和微生物生物量碳、微生物生物量氮含量的变化(平均值±标准偏差, n = 3)
Table 3 Soil physical, chemical properties and microbial biomass carbon and nitrogen contents in the litter removal, control, and litter addition treatments plots in the two Castanopsis carlesii forests (mean ± SD, n = 3)
| 项目 Item | 米槠次生林 Secondary Castanopsis carlesii forest | 米槠人促更新次生林 Human-assisted naturally regenerated Castanopsis carlesii forest | ||||
|---|---|---|---|---|---|---|
| 对照 Control | 凋落物去除 No litter | 凋落物添加 Double litter | 对照 Control | 凋落物去除 No litter | 凋落物添加 Double litter | |
| 含水量 Moisture content (%) | 0.30 ± 0.00aA | 0.23 ± 0.01bA | 0.29 ± 0.03abA | 0.26 ± 0.06aA | 0.21 ± 0.01aA | 0.25 ± 0.03aA |
| pH值 pH value | 5.11 ± 0.14aA | 5.10 ± 0.04aA | 4.91 ± 0.16aA | 5.25 ± 0.17aA | 5.11 ± 0.02aA | 5.13 ± 0.10aA |
| 全碳 Total carbon (g·kg-1) | 20.98 ± 2.75aA | 12.41 ± 1.75bA | 16.49 ± 0.47bB | 16.68 ± 0.44aB | 12.77 ± 0.09bA | 14.71 ± 3.00abA |
| 全氮 Total nitrogen (g·kg-1) | 1.42 ± 0.09aA | 0.93 ± 0.12cA | 1.14 ± 0.02bB | 1.14 ± 0.09aB | 0.88 ± 0.07bA | 1.02 ± 0.19abA |
| 可溶性有机碳 Dissolved organic carbon (mg·kg-1) | 49.15 ± 8.37aA | 29.77 ± 9.46bA | 55.58 ± 8.7aA | 48.86 ± 6.76aA | 47.11 ± 10.09aA | 49.52 ± 6.46aA |
| 可溶性有机氮 Dissolved organic nitrogen (mg·kg-1) | 2.08 ± 0.02aA | 1.85 ± 0.09aA | 0.74 ± 0.15bB | 2.26 ± 0.36aA | 2.18 ± 0.43aA | 1.86 ± 0.44aA |
| NH4+ (mg·kg-1) | 37.49 ± 0.69bA | 26.5 ± 3.21cA | 52.6 ± 0.6aA | 26.99 ± 4.01bB | 22.04 ± 3.12aA | 29.63 ± 4.41aB |
| 矿质氮 Mineral nitrogen (mg·kg-1) | 38.08 ± 0.52bA | 27.22 ± 3.31cA | 53.05 ± 0.82aA | 27.96 ± 3.94aB | 22.37 ± 3.24bA | 30.98 ± 4.41aB |
| 微生物生物量碳 Microbial biomass carbon (mg·kg-1) | 315 ± 33abA | 225 ± 66bA | 371 ± 57aA | 258 ± 17aB | 171 ± 5cA | 221 ± 23bB |
| 微生物生物量氮 Microbial biomass nitrogen (mg·kg-1) | 29.33 ± 6.30aA | 13.41 ± 2.27bA | 26.38 ± 5.36aA | 36.56 ± 3.49aA | 14.61 ± 2.58cA | 23.05 ± 5.07bA |
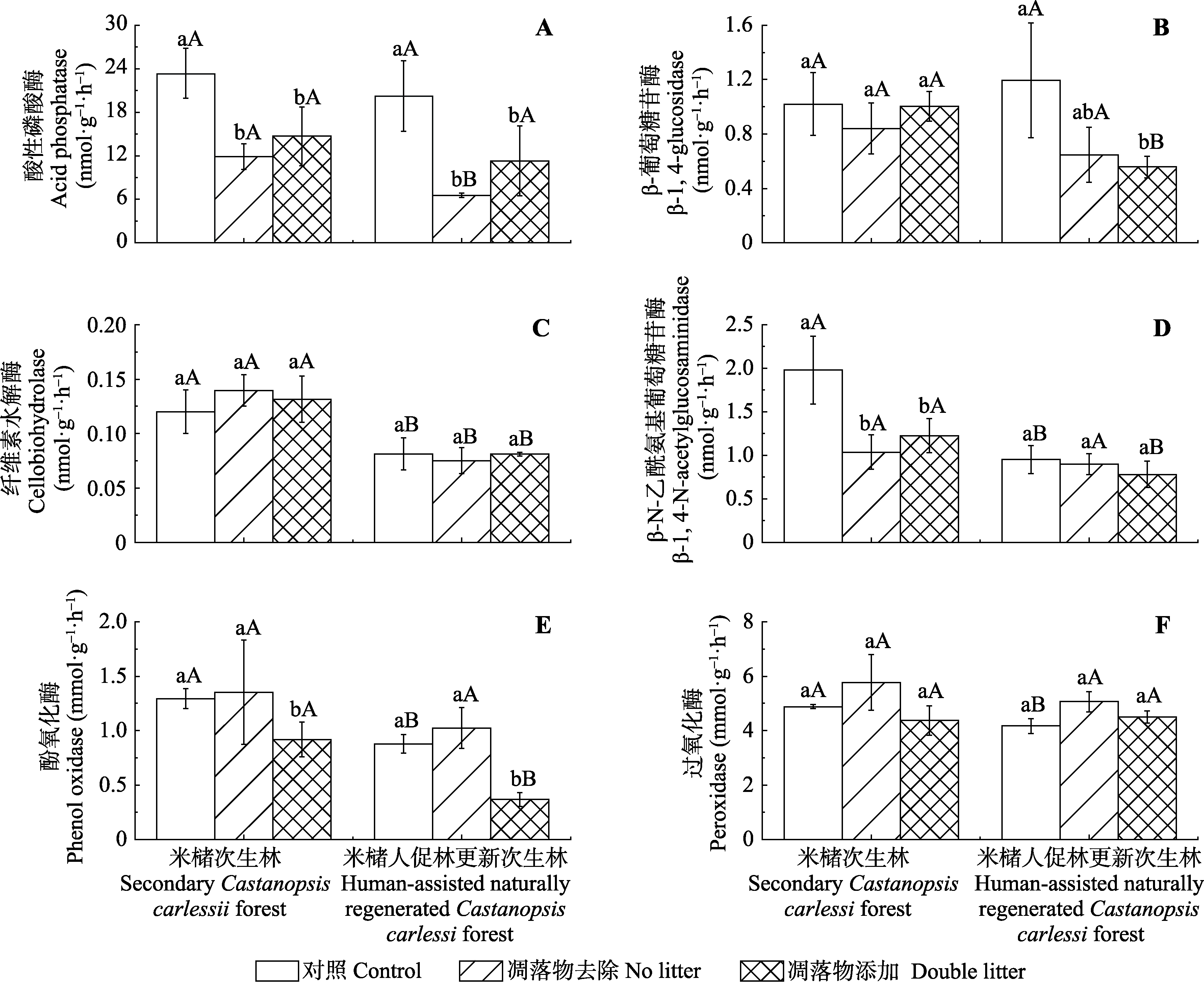
图1 次生林和人促林中凋落物添加和去除后土壤酶活性的变化(平均值±标准偏差, n = 3)。小写字母表示同一林分下不同处理间的差异性(p < 0.05), 大写字母表示不同林分同一处理间的差异性(p < 0.05)。
Fig. 1 Soil enzyme activities under litter removal, control, and litter addition treatments in the two Castanopsis carlesii forests (mean ± SD, n = 3). The lowercase letters mean significant differences among treatments in the same forests (p < 0.05), the capital letters mean significant differences between the two forests under the same treatment (p < 0.05).
| 项目 Item | 酸性磷酸酶 Acid phosphatase | β-葡萄糖苷酶 β-1,4-glucosidase | 纤维素水解酶 Cellobiohydrolase | β-N-乙酰氨基葡萄糖苷酶 β-1,4-N-acetylglucosam- inidase | 酚氧化酶 Phenol oxidase | 过氧化酶 Peroxidase |
|---|---|---|---|---|---|---|
| 含水量 Moisture content (%) | 0.545* | 0.327 | 0.344 | 0.504* | 0.221 | -0.230 |
| 全碳 Total carbon | 0.638** | 0.312 | -0.003 | 0.751** | 0.119 | -0.164 |
| 全氮 Total nitrogen | 0.830** | 0.499* | 0.248 | 0.807** | 0.245 | -0.109 |
| 可溶性有机碳 Dissolved organic carbon | 0.274 | 0.225 | -0.175 | 0.195 | -0.391 | -0.624** |
| 可溶性有机氮 Dissolved organic nitrogen | 0.060 | -0.140 | -0.534* | -0.139 | -0.205 | 0.105 |
| NH4+ | 0.362 | 0.296 | 0.527* | 0.411 | 0.035 | -0.279 |
| 微生物生物量碳 Microbial biomass carbon | 0.523* | 0.488* | 0.531* | 0.524* | 0.252 | -0.163 |
| 微生物生物量碳 Microbial biomass nitrogen | 0.648** | 0.498* | -0.118 | 0.231 | -0.150 | -0.608** |
表4 土壤酶活性与土壤理化性质和微生物生物量碳、微生物生物量氮的相关关系
Table 4 Correlation between soil enzyme activity and soil physical, chemical properties, microbial biomass carbon, microbial biomass nitrogen
| 项目 Item | 酸性磷酸酶 Acid phosphatase | β-葡萄糖苷酶 β-1,4-glucosidase | 纤维素水解酶 Cellobiohydrolase | β-N-乙酰氨基葡萄糖苷酶 β-1,4-N-acetylglucosam- inidase | 酚氧化酶 Phenol oxidase | 过氧化酶 Peroxidase |
|---|---|---|---|---|---|---|
| 含水量 Moisture content (%) | 0.545* | 0.327 | 0.344 | 0.504* | 0.221 | -0.230 |
| 全碳 Total carbon | 0.638** | 0.312 | -0.003 | 0.751** | 0.119 | -0.164 |
| 全氮 Total nitrogen | 0.830** | 0.499* | 0.248 | 0.807** | 0.245 | -0.109 |
| 可溶性有机碳 Dissolved organic carbon | 0.274 | 0.225 | -0.175 | 0.195 | -0.391 | -0.624** |
| 可溶性有机氮 Dissolved organic nitrogen | 0.060 | -0.140 | -0.534* | -0.139 | -0.205 | 0.105 |
| NH4+ | 0.362 | 0.296 | 0.527* | 0.411 | 0.035 | -0.279 |
| 微生物生物量碳 Microbial biomass carbon | 0.523* | 0.488* | 0.531* | 0.524* | 0.252 | -0.163 |
| 微生物生物量碳 Microbial biomass nitrogen | 0.648** | 0.498* | -0.118 | 0.231 | -0.150 | -0.608** |
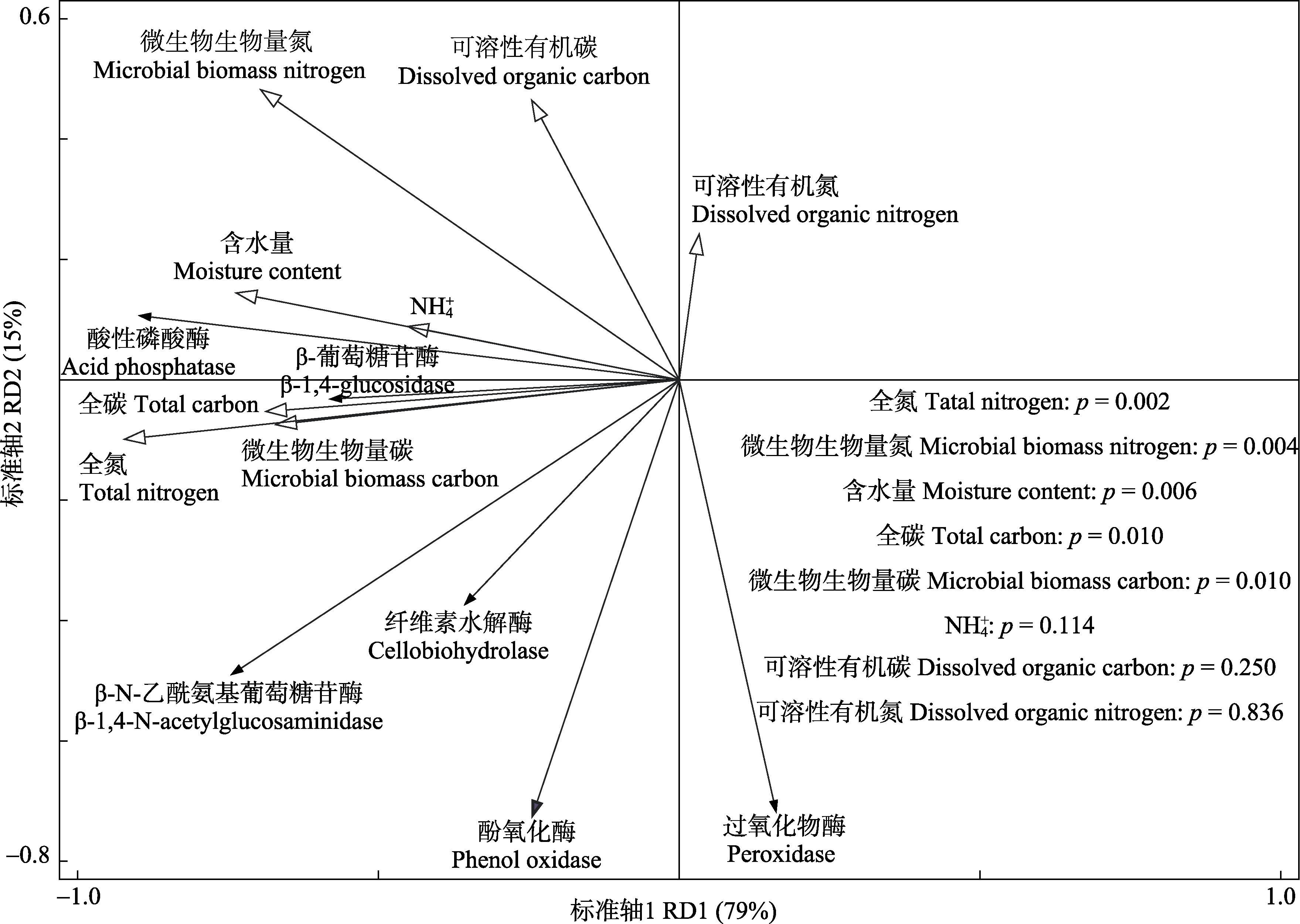
图2 土壤酶活性与土壤理化性质和微生物生物量碳、微生物生物量氮冗余分析。
Fig. 2 Redundancy analysis on the relationship of soil enzyme activity and soil physical, chemical properties, microbial biomass carbon, microbial biomass nitrogen.
| [1] |
Allison SD, Vitousek PM ( 2005). Responses of extracellular enzymes to simple and complex nutrient inputs. Soil Biology & Biochemistry, 37, 937-944.
DOI URL |
| [2] |
Baldrian P ( 2009). Microbial enzyme-catalyzed processes in soils and their analysis. Plant Soil and Environment, 55, 370-378.
DOI URL |
| [3] | Blazier MA, Hennessey TC, Deng S ( 2005). Effects of fertilization and vegetation control on microbial biomass carbon and dehydrogenase activity in a juvenile loblolly pine plantation. Forest Science, 51, 449-459. |
| [4] | Brockett BF, Prescott CE, Grayston SJ ( 2012). Soil moisture is the major factor influencing microbial community structure and enzyme activities across seven biogeoclimatic zones in western Canada. Soil Biology & Biochemistry, 44, 9-20. |
| [5] | Cao FQ, Liu ZH, Liu M, Cui JF ( 2010). Research progress on the forest litterfall and its decomposition process. Guangxi Agricultural Sciences, 41, 693-697. |
| [ 曹富强, 刘朝晖, 刘敏, 崔俊峰 ( 2010). 森林凋落物及其分解过程的研究进展. 广西农业科学, 41, 693-697.] | |
| [6] | Cao H, Sun H, Yang H, Sun B, Zhao QG ( 2003). A review soil enzyme activity and its indication for soil quality. Chinese Journal of Applied and Environmental Biology, 9, 105-109. |
| [ 曹慧, 孙辉, 杨浩, 孙波, 赵其国 ( 2003). 土壤酶活性及其对土壤质量的指示研究进展. 应用与环境生物学报, 9, 105-109.] | |
| [7] |
Cernusak LA, Winter K, Dalling JW, Holtum JAM, Jaramillo C, K?rner C, Leakey ADW, Norby RJ, Poulter B, Turner BL, Wright S ( 2013). Tropical forest responses to increasing atmospheric CO2: Current knowledge and opportunities for future research. Functional Plant Biology, 40, 531-551.
DOI URL |
| [8] |
Chen SS, Chen SR, Ma XQ ( 2001). Effects of different regeneration pattern of secondary broadleaved forest on stand composition and soil fertility. Scientia Silvae Sinicae, 37(6), 113-117.
DOI |
|
[ 陈绍栓, 陈淑容, 马祥庆 ( 2001). 次生阔叶林不同更新方式对林分组成及土壤肥力的影响. 林业科学, 37(6), 113-117.]
DOI |
|
| [9] |
Ding SY, She JY, Yang QP, Wang SL ( 2015). Effects of thinning and pruning on soil microbial biomass carbon and soilenzyme activities in Chinese fir plantation. Journal of Central South University of Forestry & Technology, 35(6), 75-79.
DOI URL |
|
[ 丁思一, 佘济云, 杨庆朋, 汪思龙 ( 2015). 间伐和修枝对杉木人工林土壤微生物量碳和酶活性的影响. 中南林业科技大学学报, 35(6), 75-79.]
DOI URL |
|
| [10] |
Doughty CE, Metcalfe DB, Girardin CA, Girardin F, Amezquita FF, Cabrera DG, Huasco WH, Silva-Espejo JE, Araujo-Murakami A, Costa MC, Rocha W, Feldpausch TR, Mendoza ALM, Costa ACL, Meir P, Phillips OL, Malhi Y ( 2015). Drought impact on forest carbon dynamics and fluxes in Amazonia. Nature, 519, 78-82.
DOI URL PMID |
| [11] | Fekete I, Varga C, Kotroczó Z, Tóth JA, Várbiró G ( 2011). The relation between various detritus inputs and soil enzyme activities in a Central European deciduous forest. Geoderma, 167- 168, 15-21. |
| [12] | Gray DB, Mary KT, Julie EJ ( 2002). Interactions between crop residue and soil organic matter quality and the functional diversity of soil microbial communities. Soil Biology & Biochemistry, 34, 1073-1082. |
| [13] |
Hernandez DL, Hobbie SE ( 2010). The effects of substrate composition, quantity, and diversity on microbial activity. Plant and Soil, 335, 397-411.
DOI URL |
| [14] | Huang QL, Zheng QR, Rong JT, Zhuo MX, Guan XH ( 2012). Selective logging techniques of natural broad-leaved timber forest in the mid-subtropical zone of Fujian, China. Journal of Mountain Science, 30(2), 180-185. |
| [ 黄清麟, 郑群瑞, 戎建涛, 卓鸣秀, 官晓辉 ( 2012). 福建中亚热带天然阔叶用材林择伐技术I. 基于树种特征的目标树种清单. 山地学报, 30(2), 180-185.] | |
| [15] |
Huang YM, Yang WQ, Zhang J, Lu CT, Liu X, Wang W, Guo W, Zhang DJ ( 2014). Response of soil microorganism and soil enzyme activity to understory plant removal in the subalpine coniferous plantation of western Sichuan. Acta Ecologica Sinica, 34, 4183-4192.
DOI URL |
|
[ 黄玉梅, 杨万勤, 张健, 卢昌泰, 刘旭, 王伟, 郭伟, 张丹桔 ( 2014). 川西亚高山针叶林土壤微生物及酶对林下植物去除的响应. 生态学报, 34, 4183-4192.]
DOI URL |
|
| [16] | Jenkinson DS, Brookes PC, Powlson DS ( 2004). Measuring soil microbial biomass. Soil Biology & Biochemistry, 36, 5-7. |
| [17] |
Kivlin SN, Treseder KK ( 2014). Soil extracellular enzyme activities correspond with abiotic factors more than fungal community composition. Biogeochemistry, 117, 23-37.
DOI URL |
| [18] |
Kotroczó Z, Veres Z, Fekete I, Krakomperger K, Tóth JA Lajtha K, Tóthmérész B ( 2014). Soil enzyme activity in response to long term organic matter manipulation. Soil Biology & Biochemistry, 70, 237-243.
DOI URL |
| [19] |
Li XJ, Liu XF, Xiong DC, Lin WS, Lin TW, Shi YW ( 2016). Impact of litterfall addition and exclusion on soil respiration in Cunninghamia lanceolata plantation and secondary Castanopsis carlesii forest in mid-subtropical China. Chinese Journal of Plant Ecology, 40, 447-457.
DOI URL |
|
[ 李晓杰, 刘小飞, 熊德成, 林伟盛, 林廷武, 施友文 ( 2016). 中亚热带杉木人工林和米槠次生林凋落物添加与去除对土壤呼吸的影响. 植物生态学报, 40, 447-457.]
DOI URL |
|
| [20] | Liu XF, Lin TC, Yang Z, Vadeboncoeur MA, Lin CF, Xiong DC, Lin WS, Chen GS, Xie JS, Li YQ, Yang YS ( 2017). Increased litter in subtropical forests boosts soil respiration in natural forests but not plantations of Castanopsis carlesii. Plant and Soil, 418, 141-148. |
| [21] | Liu X, Wang N, Zhao B, Zhang Q, Zhao XH ( 2014). Effects of carbon input changes on soil enzyme activities in a Pinus tabulaeformis forest at the Taiyue Mountain. Chinese Journal of Applied and Environmental Biology, 20, 655-661. |
| [ 刘星, 王娜, 赵博, 张青, 赵秀海 ( 2014). 改变碳输入对太岳山油松林土壤酶活性的影响. 应用与环境生物学报, 20, 655-661.] | |
| [22] | Lu RK 2000). Soil Agrichemistry Analysis Protocols.China Agriculture Science Press, Beijing |
| [ 鲁如坤 ( 2000). 土壤农业化学分析方法.中国农业科学出版社,北京.] | |
| [23] |
Olander LP, Vitousek PM ( 2000). Regulation of soil phosphatase and chitinase activity by N and P availability. Biogeochemistry, 49, 175-191.
DOI URL |
| [24] | Qi G, Wang QL, Wang XC, Yu DP, Zhou L, Zhou WM ( 2013). Soil organic carbon storage in different aged Larix gmelinii plantations in Great Xing’an Mountains of Northeast China. Chinese Journal of Applied Ecology, 24, 10-16. |
| [ 齐光, 王庆礼, 王新闯, 于大炮, 周莉, 周旺明 ( 2013). 大兴安岭林区兴安落叶松人工林土壤有机碳贮量. 应用生态学报, 24, 10-16.] | |
| [25] |
Rutigliano FA, Castaldi S, D’Ascoli R, Papa S, Carfora A, Marzaioli R ( 2009). Soil activities related to nitrogen cycle under three plant cover types in Mediterranean environment . Applied Soil Ecology, 43, 40-46.
DOI URL |
| [26] | Saiya-Cork KR, Sinsabaugh RL, Zak DR ( 2002). The effects of long term nitrogen deposition on extracellular enzyme activity in an Acer saccharum forest soil. Soil Biology & Biochemistry, 34, 1309-1315. |
| [27] |
Sayer EJ, Powers JS, Tanner EVJ ( 2007). Increased litterfall in tropical forests boosts the transfer of soil CO2 to the atmosphere. PLOS ONE, 2, e1299. DOI: 10. 1371/journal. pone.?0001299.
DOI URL PMID |
| [28] |
Shu YY, Huang JS, Zhao GJ, Bao WK, Pang XY ( 2016). Effects of a forestation with different tree species on soil enzyme activities and nutrient content in eastern Qinghai-?Tibetan Plateau, China. Acta Ecologica Sinica, 36, 394-402.
DOI URL |
|
[ 舒媛媛, 黄俊胜, 赵高卷, 包维楷, 李根前, 庞学勇 ( 2016). 青藏高原东缘不同树种人工林对土壤酶活性及养分的影响. 生态学报, 36, 394-402.
DOI URL |
|
| [29] |
Sinsabaugh RL, Lauber CL, Weintraub MN, Ahmed B, Allison SD, Crenshaw C, Contosta AS, Cusack D, Frey S, Gallo ME, Gartner TB, Hobbie SE, Holland K, Keeler BL, Powers JS, Stursova M, Takacs-Vesbach C, Waldrop MP, Wallenstein MD, Zak DR, Zeglin LH ( 2008). Stoichiometry of soil enzyme activity at global scale . Ecology Letters, 11, 1252-1264.
DOI URL PMID |
| [30] | Song XY, Niu SK, Peng B, Ke YQ ( 2006). A study on species diversity of different regeneration patterns in Castanopsis carlesii forest in Sanming, Fujian Province. Subtropical Plant Science, 35(2), 31-34. |
| [ 宋晓英, 牛树奎, 彭彪, 柯玉琼 ( 2006). 福建三明不同更新方式的米槠林物种多样性研究. 亚热带植物科学, 35(2), 31-34.] | |
| [31] |
Thomas H, Benjamin S, Icolin P, Kristina M, Paul M, Almut A, Martint S ( 2008). CO2 fertilization in temperate FACE experiments not representative of boreal and tropical forests. Global Change Biology, 14, 1531-1542.
DOI URL |
| [32] |
Vance ED, Brooks PC, Jenkinson DS ( 1987). An extraction method for measuring soil microbial biomass. Soil Biology & Biochemistry, 19, 703-707.
DOI URL |
| [33] |
Veres Z, Kotroczó Z, Fekete I, Tóth JA, Lajtha K, Townsend K, Tóthmérész B ( 2015). Soil extracellular enzyme activities are sensitive indicators of detrital inputs and carbon availability. Applied Soil Ecology, 92, 18-23.
DOI URL |
| [34] | Wang ZH, Duan CQ, Hou YP, Yang JS ( 2006). The relationship of plant species diversity to ecosystem function in relation to soil conservation in semi-humid evergreen forests, Yunnan Province, China. Journal of Plant Ecology (Chinese Version), 30, 392-403. |
| [ 王震洪, 段昌群, 侯永平, 杨建松 ( 2006). 植物多样性与生态系统土壤保持功能关系及其生态学意义. 植物生态学报, 30, 392-403.] | |
| [35] |
Weintraub SR, Wieder WR, Cleveland CC, Townsend AR ( 2013). Organic matter inputs shift soil enzyme activity and allocation patterns in a wet tropical forest. Biogeochemistry, 114, 313-326.
DOI URL |
| [36] | Xiong YM, Xia HP, Li ZA, Cai XA, Fu SL ( 2008). Impacts of litter and understory removal on soil properties in a subtropical Acacia mangium plantation in China. Plant and Soil, 304, 179-188. |
| [37] | Yang JT, Su ZX, Hu JY, Wu QG, He J ( 2010). Path analysis for soil organic matters and enzyme activities of Davidia involucrate Virgin forest. Chinese Journal of Applied and Environmental Biology, 16, 164-167. |
| [ 杨敬天, 苏智先, 胡进耀吴庆贵, 贺静 ( 2010). 珙桐林土壤有机质与酶活性的通径分析. 应用与环境生物学报, 16, 164-167.] | |
| [38] |
Yang WQ, Wang KY ( 2004). Advances in forest soil enzymology. Scientia Silvae Sinicae, 40(2), 152-159.
DOI URL |
|
[ 杨万勤, 王开运 ( 2004). 森林土壤酶的研究进展. 林业科学, 40(2), 152-159.]
DOI URL |
|
| [39] |
Yang WQ, Zhong ZC, Tao JP, He WM ( 2001). Study on relationship between soil enzyme activities and plant species diversity in forest ecosystem of Mt. Jinyun. Scientia Silvae Sinicae, 37(4), 124-128.
DOI |
|
[ 杨万勤, 钟章成, 陶建平, 何维明 ( 2001). 缙云山森林土壤酶活性与植物多样性的关系. 林业科学, 37(4), 124-128.]
DOI |
|
| [40] | Yang Y, Wang JF, Zhang XY, Li DD, Wang HM, Chen FS, Sun XM, Wen XF ( 2016). Mechanism of litter and understory vegetation effects on soil carbon and nitrogen hydrolase activities in Chinese fir forests. Acta Ecologica Sinica, 36, 8102-8110. |
| [ 杨洋, 王继富, 张心昱, 李丹丹, 王辉民, 陈伏生, 孙晓敏, 温学发 ( 2016). 凋落物和林下植被对杉木林土壤碳氮水解酶活性的影响机制. 生态学报, 36, 8102-8110.] | |
| [41] | Yin P, Hu X, Wu Y ( 2015). Responses of soil enzyme activity on snow cover and litter input in alpine area. Journal of Jinan University, 36, 115-118. |
| [ 尹鹏, 胡霞, 吴彦 ( 2015). 川西高原土壤酶活性对雪被覆盖和凋落物添加的响应. 暨南大学学报, 36, 115-118.] | |
| [42] | Yuan XC, Lin WS, Pu XT, Yang ZR, Zheng W, Chen YM ( 2016). Effects of forest regeneration patterns on the quantity and chemical structure of soil solution dissolved organic matter in a subtropical forest. Chinese Journal of Applied Ecology, 27, 1845-1852. |
| [ 元晓春, 林伟盛, 蒲晓婷, 杨智榕, 郑蔚, 陈岳民 ( 2016). 更新方式对亚热带森林土壤溶液可溶性有机质数量及化学结构的影响. 应用生态学报, 27, 1845-1852.] | |
| [43] | Zhang P, Tian XJ, He XB, Song FQ, Ren LL ( 2007). Enzyme activities in litter, fragmentation and humus layers of subtropical forests. Ecology and Environment, 16, 1024-1029. |
| [ 张鹏, 田兴军, 何兴兵, 宋富强, 任利利 ( 2007). 亚热带森林凋落物层土壤酶活性的季节动态. 生态环境学报, 16, 1024-1029.] | |
| [44] | Zhao J ( 2016). Effects of Nitrogen Addition and Litter on Soil Microorganism and Enzyme Activities. Master degree dissertation, Beijing Forestry University, Beijing. |
| [ 赵静 ( 2016). 氮添加与凋落物对土壤微生物和酶活性的影响. 硕士学位论文, 北京林业大学. 北京.] | |
| [45] | Zheng WG, Xue L, Xu PB, Liang LL, Feng HF ( 2011). Soil response to litter in a Pinus caribaea woodland. Journal of South China Agricultural University, 32(1), 21-25. |
| [ 郑卫国, 薛立, 许鹏波, 梁丽丽, 冯慧芳 ( 2011). 加勒比松林地土壤对凋落物的响应. 华南农业大学学报, 32(1), 21-25.] | |
| [46] | Zhu TB, Zhuge YP, Liu SJ, Lou YH ( 2008). Effects of different fertilizer treatments and water regimes on soil enzyme activities. Shandong Agricultural Sciences,( 3), 74-78. |
| [ 朱同彬, 诸葛玉平, 刘少军, 娄燕宏 ( 2008). 不同水肥条件对土壤酶活性的影响. 山东农业科学, ( 3), 74-78.] |
| [1] | 刘瑶 钟全林 徐朝斌 程栋梁 郑跃芳 邹宇星 张雪 郑新杰 周云若. 不同大小刨花楠细根功能性状与根际微环境关系[J]. 植物生态学报, 2024, 48(预发表): 0-0. |
| [2] | 吴君梅, 曾泉鑫, 梅孔灿, 林惠瑛, 谢欢, 刘苑苑, 徐建国, 陈岳民. 土壤磷有效性调控亚热带森林土壤酶活性和酶化学计量对凋落叶输入的响应[J]. 植物生态学报, 2024, 48(2): 242-253. |
| [3] | 汪子微, 万松泽, 蒋洪毛, 胡扬, 马书琴, 陈有超, 鲁旭阳. 青藏高原不同高寒草地类型土壤酶活性及其影响因子[J]. 植物生态学报, 2021, 45(5): 528-538. |
| [4] | 解梦怡, 冯秀秀, 马寰菲, 胡汗, 王洁莹, 郭垚鑫, 任成杰, 王俊, 赵发珠. 秦岭锐齿栎林土壤酶活性与化学计量比变化特征及其影响因素[J]. 植物生态学报, 2020, 44(8): 885-894. |
| [5] | 邹瓒, 陈劲松, 李洋, 宋会兴. 光合产物传输方向对蓉城竹根际微生物过程的影响[J]. 植物生态学报, 2018, 42(8): 863-872. |
| [6] | 王祥, 朱亚琼, 郑伟, 关正翾, 盛建东. 昭苏山地草甸4种典型土地利用方式下的土壤呼吸特征[J]. 植物生态学报, 2018, 42(3): 382-396. |
| [7] | 卢玉鹏, 许纪元, 张晓曦, 王博雅, 谢博, 刘增文. 林下药用植物淋出物对红桦和杜仲枯落物分解及土壤酶活性的影响[J]. 植物生态学报, 2017, 41(6): 639-649. |
| [8] | 李晓杰, 刘小飞, 熊德成, 林伟盛, 林廷武, 施友文, 谢锦升, 杨玉盛. 中亚热带杉木人工林和米槠次生林凋落物添加与去除对土壤呼吸的影响[J]. 植物生态学报, 2016, 40(5): 447-457. |
| [9] | 刘成刚, 薛建辉. 喀斯特石漠化山地不同类型人工林土壤的基本性质和综合评价[J]. 植物生态学报, 2011, 35(10): 1050-1060. |
| [10] | 袁丽环, 闫桂琴. 丛枝菌根化翅果油树幼苗根际土壤微环境[J]. 植物生态学报, 2010, 34(6): 678-686. |
| [11] | 蒋智林, 刘万学, 万方浩, 李正跃. 紫茎泽兰与非洲狗尾草单、混种群落土壤酶活性和土壤养分的比较[J]. 植物生态学报, 2008, 32(4): 900-907. |
| [12] | 刘恩科, 赵秉强, 李秀英, 姜瑞波, 李燕婷, Hwat Bing So. 长期施肥对土壤微生物量及土壤酶活性的影响[J]. 植物生态学报, 2008, 32(1): 176-182. |
| [13] | 李传荣, 许景伟, 宋海燕, 李春艳, 郑莉, 王卫东, 王月海. 黄河三角洲滩地不同造林模式的土壤酶活性[J]. 植物生态学报, 2006, 30(5): 802-809. |
| [14] | 杨玉盛, 李振问, 俞新妥, 何宗明. 南平溪后杉木林取代杂木林后土壤肥力变化的研究[J]. 植物生态学报, 1994, 18(3): 236-242. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
Copyright © 2022 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19