
植物生态学报 ›› 2018, Vol. 42 ›› Issue (5): 550-561.DOI: 10.17521/cjpe.2017.0263
孙小伟,杨庆松,刘何铭,王希华( )
)
收稿日期:2017-10-19
修回日期:2018-05-07
出版日期:2018-05-20
发布日期:2018-07-20
通讯作者:
王希华
基金资助:
SUN Xiao-Wei,YANG Qing-Song,LIU He-Ming,WANG Xi-Hua()
Received:2017-10-19
Revised:2018-05-07
Online:2018-05-20
Published:2018-07-20
Contact:
Xi-Hua WANG
Supported by:摘要:
为获取能够代表浙江天童山的森林植被典型群丛类型, 同时也为植被分类中如何发现过渡类型和确定典型类型提供参考, 该研究利用天童20 hm 2森林大样地资料, 运用双向指示种分析(TWINSPAN)与除趋势对应分析(DCA), 剔除过渡群落, 进行群丛划分。结果表明, 去除过渡地段后更利于研究区域典型群丛类型的确定。大样地的植被类型可划分为宜昌荚蒾-厚皮香/港柯+云山青冈群丛(Viburnum erosum-Ternstroemia gymnanthera/Lithocarpus harlandii + Cyclobalanopsis sessilifolia Ass.); 虎皮楠-柯/木荷+米槠群丛(Daphniphyllum oldhami-Lithocarpus glaber/Schima superba + Castanopsis carlesii Ass.); 红毒茴-紫楠/南酸枣+薄叶润楠群丛(Illicium lanceolatum-Phoebe sheareri/Choerospondias axillaries + Machilus leptophylla Ass.)。DCA排序同时能反映各群丛类型分布与环境的相关关系, 结果显示, 海拔和凹凸度对群丛分布有较大影响, 坡度和坡向对群丛分布影响较小。
孙小伟, 杨庆松, 刘何铭, 王希华. 基于浙江天童20 hm2常绿阔叶林动态监测样地的群丛划分. 植物生态学报, 2018, 42(5): 550-561. DOI: 10.17521/cjpe.2017.0263
SUN Xiao-Wei, YANG Qing-Song, LIU He-Ming, WANG Xi-Hua. Classification of plant associations based on a 20 hm2 dynamics plot of evergreen broad- leaved forest in Mt. Tiantong, Zhejiang, China. Chinese Journal of Plant Ecology, 2018, 42(5): 550-561. DOI: 10.17521/cjpe.2017.0263
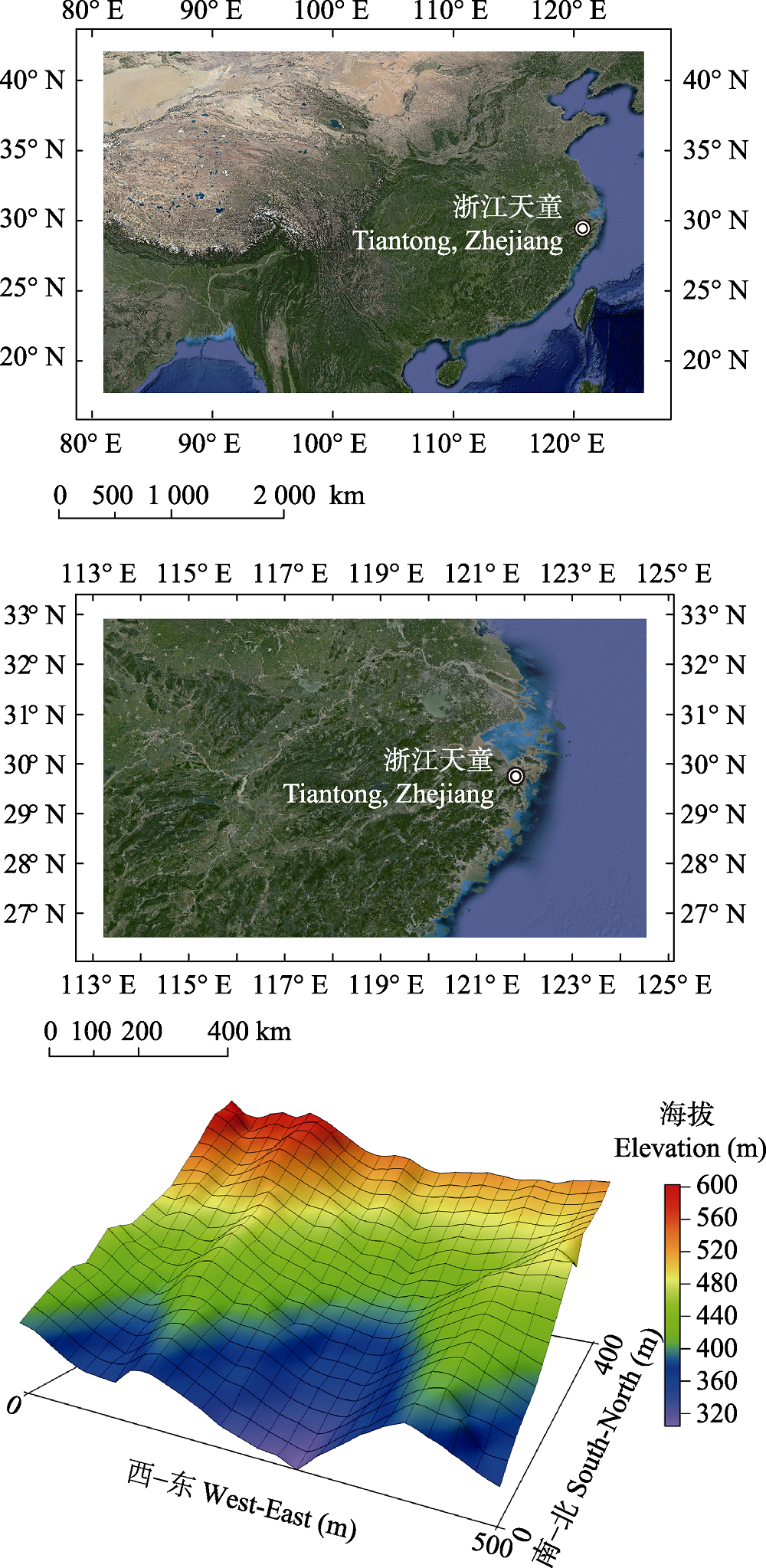
图1 浙江省天童20 hm2森林动态样地所在位置及三维地形图。
Fig. 1 The location and topography of the 20 hm2 forest dynamics plot in Mt. Tiantong, Zhejiang Province.
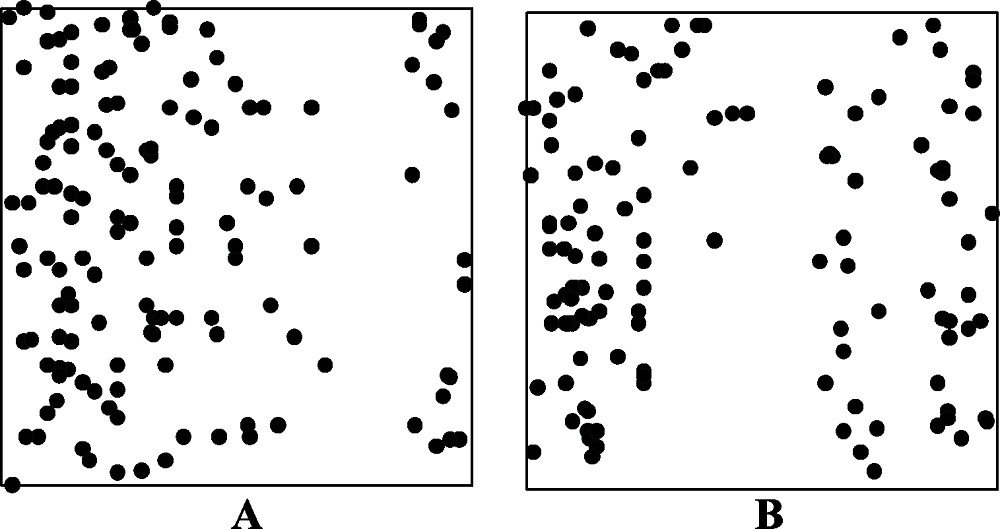
图2 物种分布不均匀的样方(A, 样方编号83, B, 样方编号324)。圆点表示样方中的植物个体。
Fig. 2 Quadrats with uneven distribution of species (A, stands for quadrat 83; B, stands for quadrat 324). Dots represent the plant individuals in each quadrat.
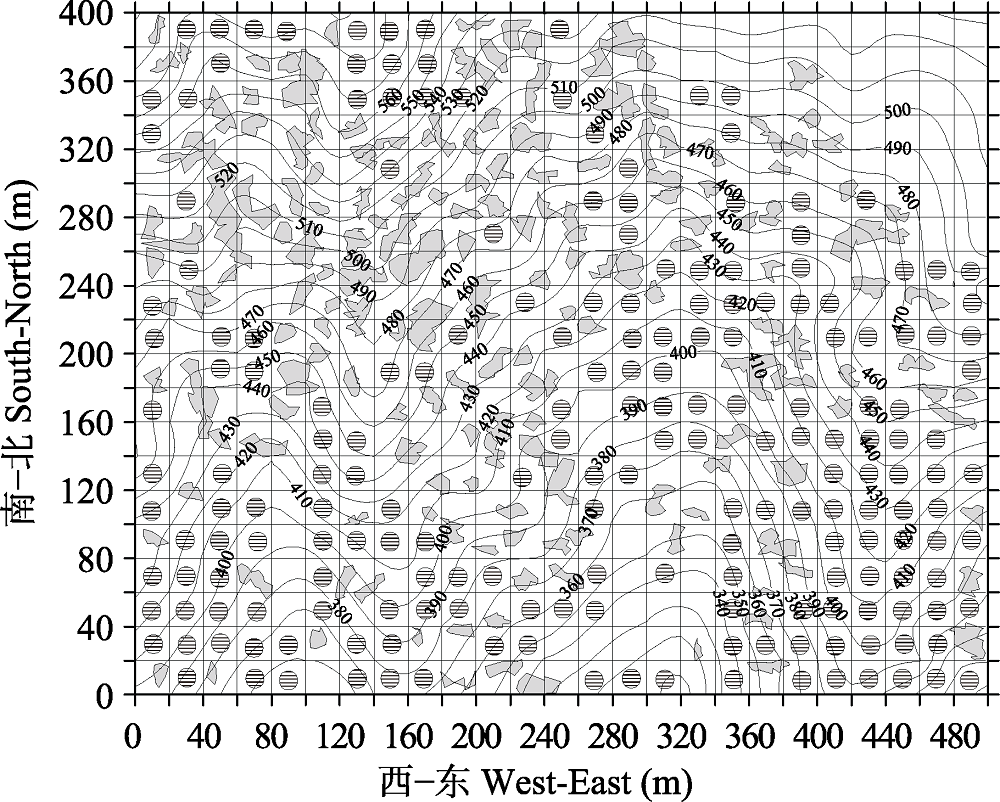
图3 天童20 hm2样地中用于分类的20 m × 20 m的样方分布图(198个样方, 等高线间距为10 m)。灰色阴影部分为林窗, 含有阴影圆圈的样方为用于分类的样方。
Fig. 3 Map of quadrats for classification at the scale of 20 m × 20 m in Tiantong 20 hm2 plot (198 quadrats in total) (The contour lines with 10 m intervals were shown). Gray polygons represent forest gaps, and shadow circles stand for the quadrats used for classification.
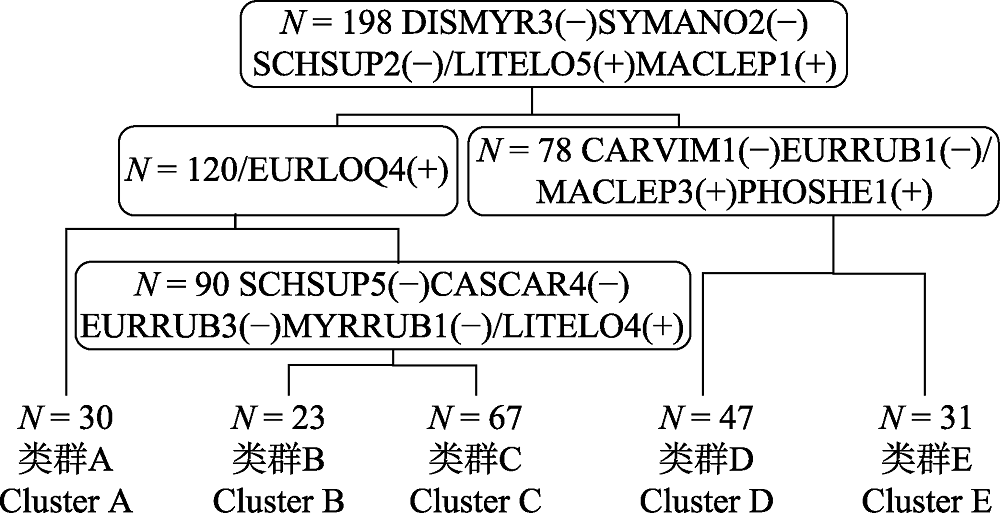
图4 天童20 hm2样地内用于分类的198个样方的TWINSPAN分类结果树状示意图。N表示样方总数, 方框里的物种表示各分类水平的正负指示种, 两者用“/”区分。CARVIM, 雷公鹅耳枥; CASCAR, 米槠; DISMYR, 杨梅叶蚊母树; EURLOQ, 细枝柃; EURRUB, 窄基红褐柃; LITELO, 黄丹木姜子; MACLEP, 薄叶润楠; MYRRUB, 杨梅; PHOSHE, 紫楠; SCHSUP, 木荷; SYMANO, 薄叶山矾。
Fig. 4 The result of TWINSPAN classification for 198 quadrats in Tiantong 20 hm2 plot. N represents total number of quadrats. Species in the box are indicator species, and a negative group and a positive group are included at each level of classification, they are separated by “/”. CARVIM, Carpinus viminea; CASCAR, Castanopsis carlesii; DISMYR, Distylium myricoides; EURLOQ, Eurya loquaiana; EURRUB, Eurya rubiginosa var. attenuata; LITELO, Litsea elongata; MACLEP, Machilus leptophylla; MYRRUB, Myrica rubra; PHOSHE, Phoebe sheareri; SCHSUP, Schima superba; SYMANO, Symplocos anomala.

图5 天童20 hm2样地5个类群的20 m × 20 m的样方分布图。A, B, C, D, E分别表示表示类群A-E包含的样方在样地中的分布。
Fig. 5 Distributions of five clusters at the scale of 20 m × 20 m in Tiantong 20 hm2 plot. A, B, C, D, E represent the distributions of the quadrats from clusters A-E, respectively.
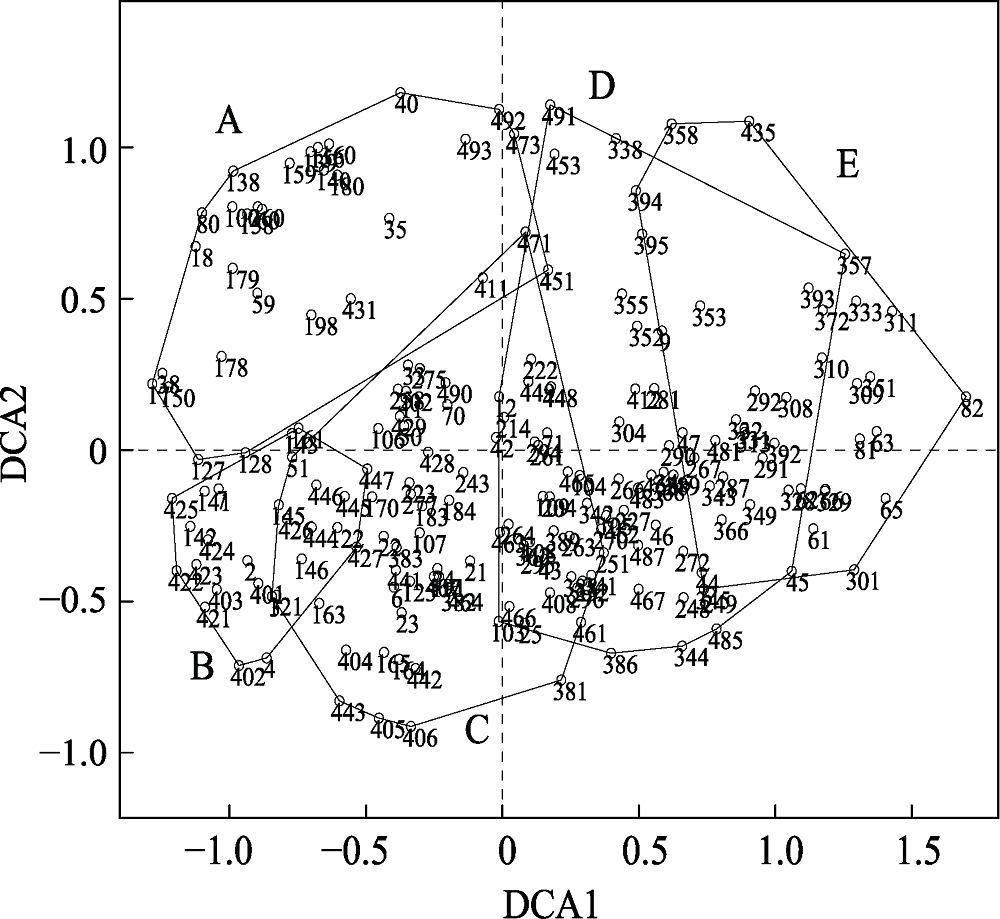
图6 天童20 hm2样地内用于分类的198个样方的DCA排序图。阿拉伯数字表示样方编号。A, 类群A; B, 类群B; C, 类群C; D, 类群D; E, 类群E。
Fig. 6 DCA ordination diagram for the classification of 198 quadrats in Tiantong 20 hm2 plot with the five clusters. The numbers in the figure are the identification code for each quadrat. A, B, C, D, E represent the distributions of the quadrats in clusters A-E, respectively.
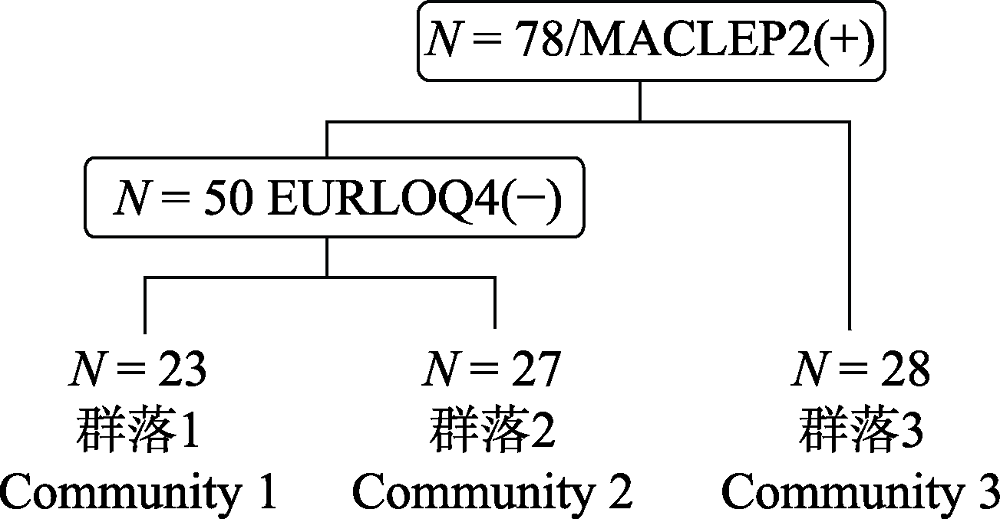
图7 去除过渡群落后剩余78个样方的TWINSPAN分类结果树状示意图。N表示样方总数, EURLOQ, 细枝柃; MACLEP, 薄叶润楠。
Fig. 7 The results of TWINSPAN classification for 78 quadrats after removing the transitional quadrats. N represents total number of quadrats. EURLOQ, Eurya loquaiana; MACLEP, Machilus leptophylla.
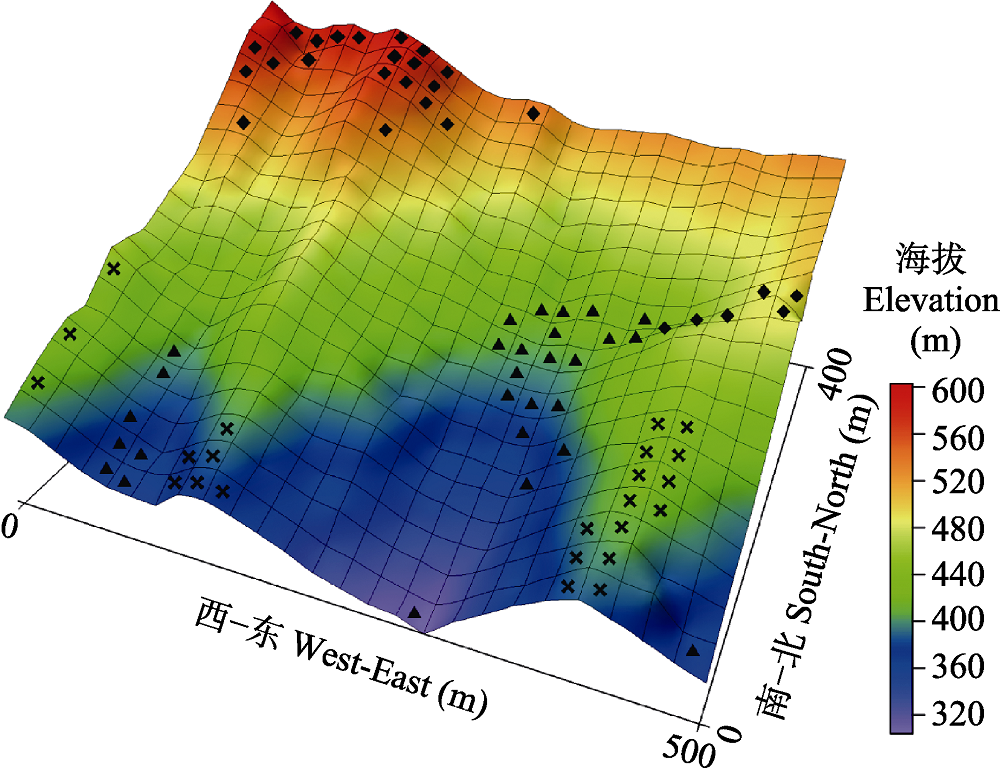
图8 天童20 hm2样地3种群落类型的20 m × 20 m的样方分布图。, , 分别表示群落1、群落2、群落3中包含的样方在样地中的分布。
Fig. 8 Distributions of three community types at the scale of 20 m × 20 m in Tiantong 20 hm2 plot. ,, and represent the distributions of the communities 1-3, respectively.
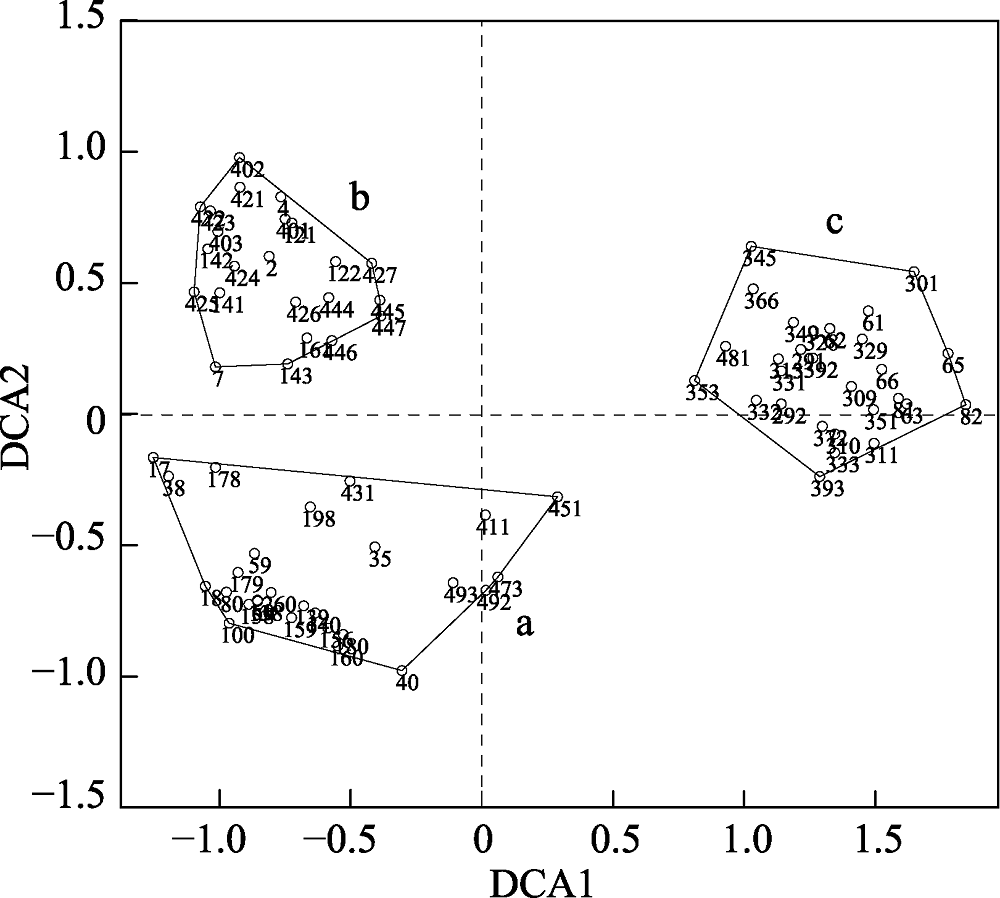
图9 去除过渡群落后剩余78个样方的DCA排序图。阿拉伯数字表示样方编号。a, 群落1; b, 群落2; c, 群落3。
Fig. 9 DCA ordination diagram of 78 quadrats after removing the transitional quadrats. The numbers in the figure are the identification code for each quadrat. a, community 1; b, community 2; c, community 3.
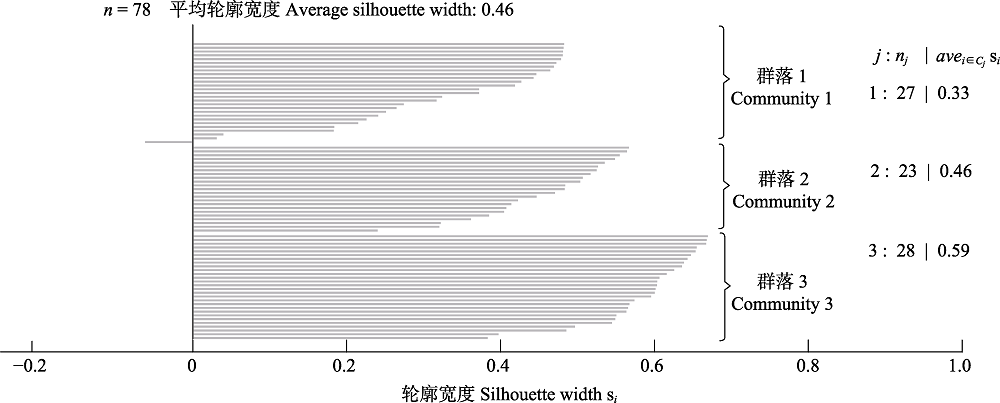
图10 3种群落类型的轮廓宽度值图。每一条水平线的长度代表一个样方的轮廓宽度值。n, 样方总数; j, 各类型编号; nj, 各群落内的样方总数; avei∈cj si, 各群落内的平均轮廓宽度值。
Fig. 10 The silhouette width of three community types. The length of each horizontal line represents the silhouette width of one quadrat. n, total number of quadrats; j, cluster number; nj, total number of quadrats in the community j; avei∈cj si, the average silhouette width in the community j.
| 物种 Species | 重要值 Important value (%) | 生长型 Growth form |
|---|---|---|
| 群落1 Community 1 | ||
| 港柯 Lithocarpus harlandii | 12.20 | 乔木 Tree |
| 杨梅叶蚊母树 Distylium myricoides | 10.97 | 小乔木 Small tree |
| 云山青冈 Cyclobalanopsis sessilifolia | 9.27 | 乔木 Tree |
| 木荷 Schima superba | 6.10 | 乔木 Tree |
| 马银花 Rhododendron ovatum | 6.06 | 小乔木 Small tree |
| 群落2 Community 2 | ||
| 细枝柃 Eurya loquaiana | 13.38 | 小乔木 Small tree |
| 木荷 Schima superba | 12.46 | 乔木 Tree |
| 杨梅叶蚊母树 Distylium myricoides | 12.12 | 小乔木 Small tree |
| 米槠 Castanopsis carlesii | 10.04 | 乔木 Tree |
| 栲 Castanopsis fargesii | 6.01 | 乔木Tree |
| 群落3 Community 3 | ||
| 黄丹木姜子 Litsea elongata | 21.46 | 小乔木 Small tree |
| 薄叶润楠 Machilus leptophylla | 12.06 | 乔木 Tree |
| 南酸枣 Choerospondias axillaris | 10.48 | 乔木 Tree |
| 细枝柃 Eurya loquaiana | 9.34 | 小乔木 Small tree |
| 云山青冈 Cyclobalanopsis sessilifolia | 5.90 | 乔木 Tree |
表1 天童20 hm2样地3种群落类型内物种的重要值排序(前5)
Table 1 The order of species important value in three community types in Tiantong 20 hm2 plot (top 5)
| 物种 Species | 重要值 Important value (%) | 生长型 Growth form |
|---|---|---|
| 群落1 Community 1 | ||
| 港柯 Lithocarpus harlandii | 12.20 | 乔木 Tree |
| 杨梅叶蚊母树 Distylium myricoides | 10.97 | 小乔木 Small tree |
| 云山青冈 Cyclobalanopsis sessilifolia | 9.27 | 乔木 Tree |
| 木荷 Schima superba | 6.10 | 乔木 Tree |
| 马银花 Rhododendron ovatum | 6.06 | 小乔木 Small tree |
| 群落2 Community 2 | ||
| 细枝柃 Eurya loquaiana | 13.38 | 小乔木 Small tree |
| 木荷 Schima superba | 12.46 | 乔木 Tree |
| 杨梅叶蚊母树 Distylium myricoides | 12.12 | 小乔木 Small tree |
| 米槠 Castanopsis carlesii | 10.04 | 乔木 Tree |
| 栲 Castanopsis fargesii | 6.01 | 乔木Tree |
| 群落3 Community 3 | ||
| 黄丹木姜子 Litsea elongata | 21.46 | 小乔木 Small tree |
| 薄叶润楠 Machilus leptophylla | 12.06 | 乔木 Tree |
| 南酸枣 Choerospondias axillaris | 10.48 | 乔木 Tree |
| 细枝柃 Eurya loquaiana | 9.34 | 小乔木 Small tree |
| 云山青冈 Cyclobalanopsis sessilifolia | 5.90 | 乔木 Tree |
| 物种 Species | 特异性Specificity | 确限度Fidelity | 指示值Indicator value | p |
|---|---|---|---|---|
| 群落1 Community 1 | ||||
| 杜鹃 Rhododendron simsii | 0.9659 | 0.7037 | 0.824 | 0.001*** |
| 宜昌荚蒾 Viburnum erosum | 0.9298 | 0.5556 | 0.719 | 0.001*** |
| 厚皮香 Ternstroemia gymnanthera | 0.9643 | 0.4444 | 0.655 | 0.001*** |
| 迎春樱桃 Cerasus discoidea | 1.0000 | 0.3704 | 0.609 | 0.001*** |
| 山鸡椒 Litsea cubeba | 0.9070 | 0.4074 | 0.608 | 0.002** |
| 青冈 Cyclobalanopsis glauca | 0.8421 | 0.3704 | 0.558 | 0.003** |
| 大青 Clerodendrum cyrtophyllum | 0.8750 | 0.3333 | 0.540 | 0.007** |
| 群落2 Community 2 | ||||
| 虎皮楠 Daphniphyllum oldhami | 0.9894 | 0.6957 | 0.830 | 0.001*** |
| 柯 Lithocarpus glaber | 0.9792 | 0.6087 | 0.772 | 0.001*** |
| 杨梅 Myrica rubra | 0.8667 | 0.6087 | 0.726 | 0.001*** |
| 总状山矾 Symplocos botryantha | 0.9375 | 0.4783 | 0.670 | 0.001*** |
| 刺毛越桔 Vaccinium trichocladum | 0.8889 | 0.4783 | 0.652 | 0.001*** |
| 铁冬青 Ilex rotunda | 0.5833 | 0.5217 | 0.552 | 0.045* |
| 群落3 Community 3 | ||||
| 薄叶润楠 Machilus leptophylla | 0.9931 | 1.0000 | 0.997 | 0.001*** |
| 红毒茴 Illicium lanceolatum | 0.9695 | 0.8214 | 0.892 | 0.001*** |
| 紫楠 Phoebe sheareri | 1.0000 | 0.5714 | 0.756 | 0.001*** |
| 黄牛奶树 Symplocos cochinchinensis var. laurina | 0.8482 | 0.6429 | 0.738 | 0.001*** |
| 西川朴 Celtis vandervoetiana | 1.0000 | 0.3214 | 0.567 | 0.001*** |
| 锐角枫 Acer acutum | 1.0000 | 0.2500 | 0.500 | 0.002** |
| 胡桃楸 Juglans mandshurica | 1.0000 | 0.2500 | 0.500 | 0.001*** |
表2 3种群落类型内物种指示值(大于0.5)排序
Table 2 The order of species indicator value (>0.5) in three community types in Tiantong 20 hm2 plot
| 物种 Species | 特异性Specificity | 确限度Fidelity | 指示值Indicator value | p |
|---|---|---|---|---|
| 群落1 Community 1 | ||||
| 杜鹃 Rhododendron simsii | 0.9659 | 0.7037 | 0.824 | 0.001*** |
| 宜昌荚蒾 Viburnum erosum | 0.9298 | 0.5556 | 0.719 | 0.001*** |
| 厚皮香 Ternstroemia gymnanthera | 0.9643 | 0.4444 | 0.655 | 0.001*** |
| 迎春樱桃 Cerasus discoidea | 1.0000 | 0.3704 | 0.609 | 0.001*** |
| 山鸡椒 Litsea cubeba | 0.9070 | 0.4074 | 0.608 | 0.002** |
| 青冈 Cyclobalanopsis glauca | 0.8421 | 0.3704 | 0.558 | 0.003** |
| 大青 Clerodendrum cyrtophyllum | 0.8750 | 0.3333 | 0.540 | 0.007** |
| 群落2 Community 2 | ||||
| 虎皮楠 Daphniphyllum oldhami | 0.9894 | 0.6957 | 0.830 | 0.001*** |
| 柯 Lithocarpus glaber | 0.9792 | 0.6087 | 0.772 | 0.001*** |
| 杨梅 Myrica rubra | 0.8667 | 0.6087 | 0.726 | 0.001*** |
| 总状山矾 Symplocos botryantha | 0.9375 | 0.4783 | 0.670 | 0.001*** |
| 刺毛越桔 Vaccinium trichocladum | 0.8889 | 0.4783 | 0.652 | 0.001*** |
| 铁冬青 Ilex rotunda | 0.5833 | 0.5217 | 0.552 | 0.045* |
| 群落3 Community 3 | ||||
| 薄叶润楠 Machilus leptophylla | 0.9931 | 1.0000 | 0.997 | 0.001*** |
| 红毒茴 Illicium lanceolatum | 0.9695 | 0.8214 | 0.892 | 0.001*** |
| 紫楠 Phoebe sheareri | 1.0000 | 0.5714 | 0.756 | 0.001*** |
| 黄牛奶树 Symplocos cochinchinensis var. laurina | 0.8482 | 0.6429 | 0.738 | 0.001*** |
| 西川朴 Celtis vandervoetiana | 1.0000 | 0.3214 | 0.567 | 0.001*** |
| 锐角枫 Acer acutum | 1.0000 | 0.2500 | 0.500 | 0.002** |
| 胡桃楸 Juglans mandshurica | 1.0000 | 0.2500 | 0.500 | 0.001*** |
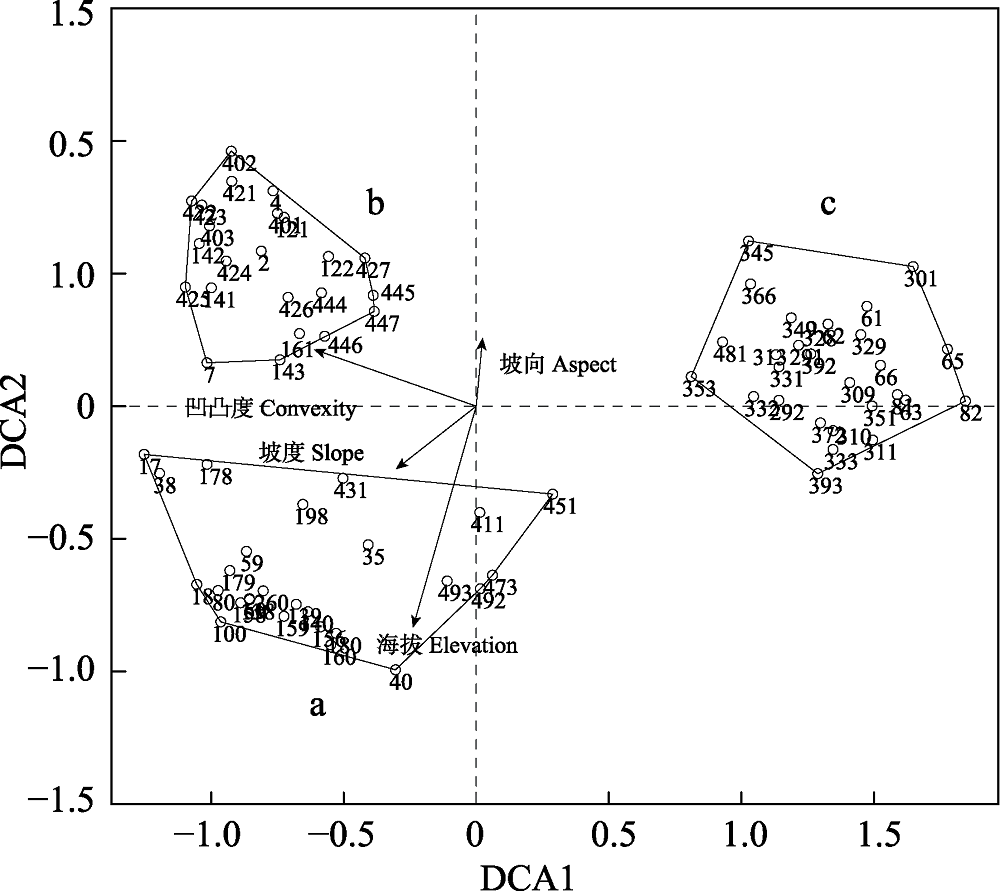
图11 78个样方的DCA排序轴与环境因子的相关性图。阿拉伯数字表示样方编号。a, 宜昌荚蒾-厚皮香/港柯+云山青冈群丛; b, 虎皮楠-柯/木荷+米槠群丛; c, 红毒茴-紫楠/南酸枣+薄叶润楠群丛。
Fig. 11 DCA ordination diagram of 78 quadrats with environmental variables. The numbers in the figure are the identification code for each quadrat. a, Viburnum erosum-Ternstroemia gymnanthera/Lithocarpus harlandii + Cyclobalanopsis sessilifolia Association; b, Daphniphyllum oldhami-Lithocarpus glaber/Schima superba + Castanopsis carlesii Association; c, Illicium lanceolatum-Phoebe sheareri/Choerospondias axillaris + Machilus leptophylla Association.
| 物种 Species | 特异性Specificity | 确限度Fidelity | 指示值Indicator value | p | ||
|---|---|---|---|---|---|---|
| 类群A Cluster A | ||||||
| 杜鹃 Rhododendron simsii | 0.8788 | 0.6667 | 0.765 | 0.001*** | ||
| 宜昌荚蒾 Viburnum erosum | 0.8281 | 0.5000 | 0.643 | 0.001*** | ||
| 厚皮香 Ternstroemia gymnanthera | 0.8056 | 0.5000 | 0.635 | 0.001*** | ||
| 山鸡椒 Litsea cubeba | 0.5362 | 0.4000 | 0.463 | 0.001*** | ||
| 迎春樱桃 Cerasus discoidea | 0.6071 | 0.3333 | 0.450 | 0.001*** | ||
| 南烛 Vaccinium bracteatum | 0.5143 | 0.3333 | 0.414 | 0.003** | ||
| 豆腐柴 Premna microphylla | 0.6000 | 0.1333 | 0.283 | 0.045* | ||
| 野鸦椿 Euscaphis japonica | 0.7500 | 0.1000 | 0.274 | 0.026* | ||
| 木蜡树 Toxicodendron sylvestre | 1.0000 | 0.0667 | 0.258 | 0.029* | ||
| 类群B Cluster B | ||||||
| 柯 Lithocarpus glaber | 0.7833 | 0.6087 | 0.691 | 0.001*** | ||
| 杨梅 Myrica rubra | 0.6047 | 0.6087 | 0.607 | 0.001*** | ||
| 刺毛越桔 Vaccinium trichocladum | 0.5333 | 0.4783 | 0.505 | 0.001*** | ||
| 总状山矾 Symplocos botryantha | 0.4688 | 0.4783 | 0.473 | 0.001*** | ||
| 类群E Cluster E | ||||||
| 紫楠 Phoebe sheareri | 0.9000 | 0.5806 | 0.723 | 0.001*** | ||
| 胡桃楸 Juglans mandshurica | 0.9167 | 0.2581 | 0.486 | 0.001*** | ||
| 锐角槭 Acer acutum | 0.8333 | 0.2258 | 0.434 | 0.001*** | ||
| 中华卫矛 Euonymus nitidus | 0.7000 | 0.1613 | 0.336 | 0.006** | ||
| 枳椇 Hovenia acerba | 0.8750 | 0.1290 | 0.336 | 0.002** | ||
| 类群B + C Cluster B + C | ||||||
| 虎皮楠 Daphniphyllum oldhami | 0.8685 | 0.5889 | 0.715 | 0.001*** | ||
| 皱柄冬青 Ilex kengii | 0.7812 | 0.1889 | 0.384 | 0.013* | ||
| 类群D + E Cluster D + E | ||||||
| 薄叶润楠 Machilus leptophylla | 0.9809 | 0.7436 | 0.854 | 0.001*** | ||
| 红毒茴 Illicium lanceolatum | 0.9031 | 0.6539 | 0.768 | 0.001*** | ||
| 黄牛奶树 Symplocos cochinchinensis var. laurina | 0.8273 | 0.5513 | 0.675 | 0.001*** | ||
| 西川朴 Celtis vandervoetiana | 0.9730 | 0.3077 | 0.547 | 0.001*** | ||
| 无患子 Sapindus saponaria | 0.8529 | 0.2564 | 0.468 | 0.001*** | ||
| 糙叶树 Aphananthe aspera | 0.8846 | 0.2308 | 0.452 | 0.001*** | ||
| 紫弹树 Celtis biondii | 0.8889 | 0.1923 | 0.413 | 0.003** | ||
| 杭州榆 Ulmus changii | 0.9853 | 0.1410 | 0.373 | 0.007** | ||
| 青钱柳 Cyclocarya paliurus | 0.8667 | 0.1282 | 0.333 | 0.009** | ||
| 大叶旱樱 Cerasus subhirtella | 0.8750 | 0.0897 | 0.280 | 0.04* | ||
| 黐花 Mussaenda esquirolii | 1.0000 | 0.0769 | 0.277 | 0.036* | ||
表3 5个类群内物种指示值排序
Table 3 The order of species indicator value in five clusters in Tiantong 20 hm2 plot
| 物种 Species | 特异性Specificity | 确限度Fidelity | 指示值Indicator value | p | ||
|---|---|---|---|---|---|---|
| 类群A Cluster A | ||||||
| 杜鹃 Rhododendron simsii | 0.8788 | 0.6667 | 0.765 | 0.001*** | ||
| 宜昌荚蒾 Viburnum erosum | 0.8281 | 0.5000 | 0.643 | 0.001*** | ||
| 厚皮香 Ternstroemia gymnanthera | 0.8056 | 0.5000 | 0.635 | 0.001*** | ||
| 山鸡椒 Litsea cubeba | 0.5362 | 0.4000 | 0.463 | 0.001*** | ||
| 迎春樱桃 Cerasus discoidea | 0.6071 | 0.3333 | 0.450 | 0.001*** | ||
| 南烛 Vaccinium bracteatum | 0.5143 | 0.3333 | 0.414 | 0.003** | ||
| 豆腐柴 Premna microphylla | 0.6000 | 0.1333 | 0.283 | 0.045* | ||
| 野鸦椿 Euscaphis japonica | 0.7500 | 0.1000 | 0.274 | 0.026* | ||
| 木蜡树 Toxicodendron sylvestre | 1.0000 | 0.0667 | 0.258 | 0.029* | ||
| 类群B Cluster B | ||||||
| 柯 Lithocarpus glaber | 0.7833 | 0.6087 | 0.691 | 0.001*** | ||
| 杨梅 Myrica rubra | 0.6047 | 0.6087 | 0.607 | 0.001*** | ||
| 刺毛越桔 Vaccinium trichocladum | 0.5333 | 0.4783 | 0.505 | 0.001*** | ||
| 总状山矾 Symplocos botryantha | 0.4688 | 0.4783 | 0.473 | 0.001*** | ||
| 类群E Cluster E | ||||||
| 紫楠 Phoebe sheareri | 0.9000 | 0.5806 | 0.723 | 0.001*** | ||
| 胡桃楸 Juglans mandshurica | 0.9167 | 0.2581 | 0.486 | 0.001*** | ||
| 锐角槭 Acer acutum | 0.8333 | 0.2258 | 0.434 | 0.001*** | ||
| 中华卫矛 Euonymus nitidus | 0.7000 | 0.1613 | 0.336 | 0.006** | ||
| 枳椇 Hovenia acerba | 0.8750 | 0.1290 | 0.336 | 0.002** | ||
| 类群B + C Cluster B + C | ||||||
| 虎皮楠 Daphniphyllum oldhami | 0.8685 | 0.5889 | 0.715 | 0.001*** | ||
| 皱柄冬青 Ilex kengii | 0.7812 | 0.1889 | 0.384 | 0.013* | ||
| 类群D + E Cluster D + E | ||||||
| 薄叶润楠 Machilus leptophylla | 0.9809 | 0.7436 | 0.854 | 0.001*** | ||
| 红毒茴 Illicium lanceolatum | 0.9031 | 0.6539 | 0.768 | 0.001*** | ||
| 黄牛奶树 Symplocos cochinchinensis var. laurina | 0.8273 | 0.5513 | 0.675 | 0.001*** | ||
| 西川朴 Celtis vandervoetiana | 0.9730 | 0.3077 | 0.547 | 0.001*** | ||
| 无患子 Sapindus saponaria | 0.8529 | 0.2564 | 0.468 | 0.001*** | ||
| 糙叶树 Aphananthe aspera | 0.8846 | 0.2308 | 0.452 | 0.001*** | ||
| 紫弹树 Celtis biondii | 0.8889 | 0.1923 | 0.413 | 0.003** | ||
| 杭州榆 Ulmus changii | 0.9853 | 0.1410 | 0.373 | 0.007** | ||
| 青钱柳 Cyclocarya paliurus | 0.8667 | 0.1282 | 0.333 | 0.009** | ||
| 大叶旱樱 Cerasus subhirtella | 0.8750 | 0.0897 | 0.280 | 0.04* | ||
| 黐花 Mussaenda esquirolii | 1.0000 | 0.0769 | 0.277 | 0.036* | ||
| [1] | Braun-Blanquet J ( 1964). Pflanzensoziologie: Grundzüge der Vegetationskunde. Springer, Wien,New York. |
| [2] | Condit R ( 1998). Tropical Forest Census Plots: Methods and Results from Barro Colorado Island, Panama and Comparison with Other Plots. Springer,Berlin. |
| [3] | de Cáceres M, Jansen F ( 2016). Indicspecies: Relationship Between Species and Groups of Sites. |
| [4] |
de Cáceres M, Legendre P ( 2009). Associations between species and groups of sites: Indices and statistical inference. Ecology, 90, 3566-3574.
DOI URL PMID |
| [5] |
de Cáceres M, Legendre P, Moretti M ( 2010). Improving indicator species analysis by combining groups of sites. Oikos, 119, 1674-1684.
DOI URL |
| [6] | Dufrêne M, Legendre P ( 1997). Species assemblages and indicator species: The need for a flexible asymmetrical approach. Ecological Monographs, 67, 345-366. |
| [7] |
Fang JY, Wang XP, Shen ZH, Tang ZY, He JS, Yu D, Jiang Y, Wang ZH, Zheng CY, Zhu JL, Guo ZD ( 2009). Methods and protocols for plant community inventory. Biodiversity Science, 17, 533-548.
DOI URL |
|
[ 方精云, 王襄平, 沈泽昊, 唐志尧, 贺金生, 于丹, 江源, 王志恒, 郑成洋, 朱江玲, 郭兆迪 ( 2009). 植物群落清查的主要内容、方法和技术规范. 生物多样性, 17, 533-548.]
DOI URL |
|
| [8] | Federal Geographic Data Committee, Vegetation Subcommittee ( FGDC-VS) ( 2008). The National Vegetation Classification Standard. Version 2. FGDC-STD-005-2008 (Version 2). Cited:2008-02. |
| [9] |
Harms KE, Condit R, Hubbell SP, Foster RB ( 2001). Habitat associations of trees and shrubs in a 50-ha neotropical forest plot. Journal of Ecology, 89, 947-959.
DOI URL |
| [10] | Hill MO ( 1979). TWINSPAN-A FORTRAN Program for Arranging Multivariate Data in an Ordered Two-Way Table by Classification of the Individuals and Attributes. Cornell University, Ithaca, USA. |
| [11] |
Huang FZ, Wang B, Ding T, Xiang WS, Li XK, Zhou AP ( 2014). Numerical classification of associations in a northern tropical karst seasonal rain forest and the relationships of these associations with environmental factors. Biodiversity Science, 22, 157-166.
DOI URL |
|
[ 黄甫昭, 王斌, 丁涛, 向悟生, 李先琨, 周爱萍 ( 2014). 弄岗北热带喀斯特季节性雨林群系数量分类及与环境的关系. 生物多样性, 22, 157-166.]
DOI URL |
|
| [12] |
Jennings MD, Faber-Langendoen D, Louckes OL, Peet RK, Roberts D ( 2009). Standards for associations and alliances of the U.S. National Vegetation Classification. Ecological Monographs, 79, 173-199.
DOI URL |
| [13] |
Lai JS, Mi XC, Ren HB, Ma KP ( 2010). Numerical classification of associations in subtropical evergreen broad-leaved forest based on multivariate regression trees―A case study of 24 hm 2 Gutianshan forest plot in China . Chinese Journal of Plant Ecology, 34, 761-769.
DOI URL |
|
[ 赖江山, 米湘成, 任海保, 马克平 ( 2010). 基于多元回归树的常绿阔叶林群丛数量分类——以古田山24公顷森林样地为例. 植物生态学报, 34, 761-769.]
DOI URL |
|
| [14] | Lepš J, Šmilauer P ( 2003). Multivariate Analysis of Ecological Data Using CANOCO. Cambridge University Press, Cambridge, UK. |
| [15] | Liu HM, Yang QS, Fang XF, Ma ZP, Shen GC, Zhang ZG, Wang ZH, Wang XH ( 2015). Influences on gap species richness in a subtropical evergreen broadleaved forest. Biodiversity Science, 23, 149-156. |
| [ 刘何铭, 杨庆松, 方晓峰, 马遵平, 沈国春, 张志国, 王樟华, 王希华 ( 2015). 亚热带常绿阔叶林林窗物种丰富度的影响因素. 生物多样性, 23, 149-156.] | |
| [16] | Maechler M, Rousseeuw P, Struyf A, Hubert M, Hornik K ( 2017). Cluster: Cluster Analysis Basics and Extensions. |
| [17] | Mueller-Dombois D, Ellenberg H ( 1974). Aims and Methods of Vegetation Ecology. John Wiley,New York. |
| [18] |
Nee S ( 2005). The neutral theory of biodiversity: Do the numbers add up? Functional Ecology, 19, 173-176.
DOI URL |
| [19] | Oksanen J, Blanchet FG, Friendly M, Kindt R, Legendre P, McGlinn D, Minchin PR, O’Hara RB, Simpson GL, Solymos P, Stevens MHH, Szoecs E, Wagner H ( 2017). Vegan: Ordination, Diversity and Dissimilarities. |
| [20] |
Rousseeuw PJ ( 1987). Silhouettes: A graphical aid to the interpretation and validation of cluster analysis. Journal of Computational & Applied Mathematics, 20, 53-65.
DOI URL |
| [21] |
Song YC ( 2011). Recognition and proposal on the vegetation classification system of China. Chinese Journal of Plant Ecology, 35, 882-892.
DOI URL |
|
[ 宋永昌 ( 2011). 对中国植被分类系统的认知和建议. 植物生态学报, 35, 882-892.]
DOI URL |
|
| [22] | Song YC ( 2017). Vegetation Ecology.2nd edn. Higher Education Press, Beijing. |
| [ 宋永昌 ( 2017). 植被生态学. 第二版. 高等教育出版社, 北京.] | |
| [23] | Song YC, Wang XH, Yan NR ( 2013). Evergreen Broad-Leaved Forest of China—Classification, Ecology, Conservation. Science Press, Beijing. |
| [ 宋永昌, 王希华, 阎恩荣 ( 2013). 中国常绿阔叶林——分类, 生态, 保育. 科学出版社, 北京.] | |
| [24] | Song YC, Wang XR ( 1995). Vegetation and Flora of Tiantong National Forest Park, Zhejiang Province. Shanghai Science and Technology Literature Press, Shanghai. 1-16. |
| [ 宋永昌, 王祥荣 ( 1995). 浙江天童国家森林公园的植被与区系. 上海科学技术文献出版社, 上海. 1-16.] | |
| [25] |
Song YC, Yan NR, Song K ( 2015). Synthetic comparison of eight dynamics plots in evergreen broadleaf forests, China. Biodiversity Science, 23, 139-148.
DOI URL |
|
[ 宋永昌, 阎恩荣, 宋坤 ( 2015). 中国常绿阔叶林8大动态监测样地植被的综合比较. 生物多样性, 23, 139-148.]
DOI URL |
|
| [26] |
Song YC, Yan NR, Song K ( 2017). An update of the vegetation classification in China. Chinese Journal of Plant Ecology, 41, 269-278.
DOI URL |
|
[ 宋永昌, 阎恩荣, 宋坤 ( 2017). 再议中国的植被分类系统. 植物生态学报, 41, 269-278.]
DOI URL |
|
| [27] | Su SH, Chang-Yang CH, Lu CL, Tsui CC, Lin TT, Lin CL, Chiou WL, Kuan LH, Chen ZS, Hsieh CF ( 2007). Fushan Subtropical Forest Dynamics Plot: Tree Species Characteristics and Distribution Patterns. Taiwan Forestry Research Institute, Taipei, China. |
| [28] |
Valencia R, Foster RB, Villa G, Condit R, Svenning JC, Hernandez C, Romoleroux K, Losos E, Magard E, Balslev H ( 2004). Tree species distributions and local habitat variation in the Amazon: Large forest plot in eastern Ecuador. Journal of Ecology, 92, 214-229.
DOI URL |
| [29] | van der Maarel E, Franklin J ( 2013). Vegetation Ecology. 2nd edn. John Wiley & Sons, Chichester, UK. 28-103. |
| [30] | Westhoff V, van der Maarel E ( Translated by Zheng HY ) ( 1985). The Braun-Blanquet approach. In: Whittaker RH ed. Classification of Plant Communities. Science Press, Beijing. 223-322. |
| [ 郑慧莹 ( 译)( 1985). 植物群落分类. 科学出版社, 北京. 223-322.] | |
| [31] | Whittaker RH ( Translated by Zhou JL, Li B, Jiang YX, Chen CD, Li SY, Zheng HY, Hu JQ)( 1985). Classification of Plant Communities. Science Press, Beijing. 18. |
| [ 周纪纶, 李博, 蒋有绪, 陈昌笃, 李世英, 郑慧莹, 胡加琪 ( 译)( 1985). 植物群落分类. 科学出版社, 北京. 18] | |
| [32] | Wu ZY ( 1980). Vegetation of China. Science Press, Beijing. |
| [ 吴征镒 ( 1980). 中国植被. 科学出版社, 北京.] | |
| [33] |
Xie YB, Ma ZP, Yang QS, Fang XF, Zhang ZG, Yan ER, Wang XH ( 2012). Coexistence mechanisms of evergreen and deciduous trees based on topographic factors in Tiantong region, Zhejiang Province, eastern China. Biodiversity Science, 20, 159-167.
DOI URL |
|
[ 谢玉彬, 马遵平, 杨庆松, 方晓峰, 张志国, 阎恩荣, 王希华 ( 2012). 基于地形因子的天童地区常绿树种和落叶树种共存机制研究. 生物多样性, 20, 159-167.]
DOI URL |
|
| [34] |
Yamakura T, Kanzake M, Itoh A, Ohkubo T, Ogino K, Chai EOK, Lee HS, Ashton PS ( 1995). Topography of a large-scale research plot established within a tropical rain forest at Lambir, Sarawak. Tropics, 5, 41-56.
DOI URL |
| [35] | Yang QS, Ma ZP, Xie YB, Zhang ZG, Wang ZH, Liu HM, Li P, Zhang N, Wang DL, Yang HB, Fang XF, Yan ER, Wang XH ( 2011). Community structure and species composition of an evergreen broad-leaved forest in Tiantong’s 20 ha dynamics plot, Zhejiang Province, eastern China. Biodiversity Science, 19, 215-223. |
| [ 杨庆松, 马遵平, 谢玉彬, 张志国, 王樟华, 刘何铭, 李萍, 张娜, 王达力, 杨海波, 方晓峰, 阎恩荣, 王希华 ( 2011). 浙江天童20 ha常绿阔叶林动态监测样地的群落特征. 生物多样性, 19, 215-223.] | |
| [36] | Zhang JT ( 2011). Quantitative Ecology. 2nd edn. Science Press,Beijing. 156-160. |
| [ 张金屯 ( 2011). 数量生态学. 第二版. 科学出版社, 北京. 156-160.] | |
| [37] | Zhang ZG, Ma ZP, Liu HM, Zheng ZM, Fang XF, Wang XH ( 2013). Topographic distribution patterns of forest gap within an evergreen broad-leaved forest in Tiantong region of Zhejiang Province, Eastern China. Chinese Journal of Applied Ecology, 24, 621-625. |
| [ 张志国, 马遵平, 刘何铭, 郑泽梅, 方晓峰, 王希华 ( 2013). 天童常绿阔叶林林窗的地形分布格局. 应用生态学报, 24, 621-625.] |
| [1] | 文佳 张新娜 王娟 赵秀海 张春雨. 性状调节幼苗存活率对邻体竞争和环境的响应 [J]. 植物生态学报, 2024, 48(预发表): 0-0. |
| [2] | 董劭琼, 侯东杰, 曲孝云, 郭柯. 柴达木盆地植物群落样方数据集[J]. 植物生态学报, 2024, 48(4): 534-540. |
| [3] | 马常钦, 黄海龙, 彭政淋, 吴纯泽, 韦庆钰, 贾红涛, 卫星. 水曲柳雌雄株复叶类型及光合功能对不同生境的响应[J]. 植物生态学报, 2023, 47(9): 1287-1297. |
| [4] | 冯珊珊, 黄春晖, 唐梦云, 蒋维昕, 白天道. 细叶云南松针叶形态和显微性状地理变异及其环境解释[J]. 植物生态学报, 2023, 47(8): 1116-1130. |
| [5] | 冯可, 刘冬梅, 张琦, 安菁, 何双辉. 旅游干扰对松山油松林土壤微生物多样性及群落结构的影响[J]. 植物生态学报, 2023, 47(4): 584-596. |
| [6] | 石荡, 郭传超, 蒋南林, 唐莹莹, 郑凤, 王瑾, 廖康, 刘立强. 新疆野杏天然更新幼株的个体特征及空间分布格局[J]. 植物生态学报, 2023, 47(4): 515-529. |
| [7] | 汪晶晶, 王嘉浩, 黄致云, Vanessa Chiamaka OKECHUKW, 胡蝶, 祁珊珊, 戴志聪, 杜道林. 不同氮水平下内生固氮菌对入侵植物南美蟛蜞菊生长策略的影响[J]. 植物生态学报, 2023, 47(2): 195-205. |
| [8] | 金伊丽, 王皓言, 魏临风, 侯颖, 胡景, 吴铠, 夏昊钧, 夏洁, 周伯睿, 李凯, 倪健. 青藏高原植物群落样方数据集[J]. 植物生态学报, 2022, 46(7): 846-854. |
| [9] | 余秋伍, 杨菁, 沈国春. 浙江天童常绿阔叶林林冠结构与群落物种组成的关系[J]. 植物生态学报, 2022, 46(5): 529-538. |
| [10] | 王国宏, 郭柯, 谢宗强, 唐志尧, 蒋延玲, 方精云. 《中国植被志》研编规范的若干说明、补充与修订[J]. 植物生态学报, 2022, 46(3): 368-372. |
| [11] | 孟庆静, 樊卫国. 刺梨的适钙类型及对高钙生境的适应性[J]. 植物生态学报, 2022, 46(12): 1562-1572. |
| [12] | 贺忠权, 刘长成, 蔡先立, 郭柯. 黔中高原喀斯特常绿与落叶阔叶混交林类型及群落特征[J]. 植物生态学报, 2021, 45(6): 670-680. |
| [13] | 钟雨辰, 王斌, 方中平, 徐小忠, 于明坚. 片段化景观中壳斗科植物种子捕食和扩散模式[J]. 植物生态学报, 2021, 45(2): 154-162. |
| [14] | 韩大勇, 张维, 努尔买买提•依力亚斯, 杨允菲. 植物种群更新的补充限制[J]. 植物生态学报, 2021, 45(1): 1-12. |
| [15] | 乔鲜果, 郭柯, 赵利清, 王孜, 刘长成. 中国长芒草群系的群落特征[J]. 植物生态学报, 2020, 44(9): 986-994. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
Copyright © 2022 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19