
植物生态学报 ›› 2005, Vol. 29 ›› Issue (3): 386-393.DOI: 10.17521/cjpe.2005.0051
蔡永立, 宋永昌
收稿日期:2004-01-15
接受日期:2004-07-24
出版日期:2005-01-15
发布日期:2005-05-30
基金资助:CAI Yong-Li, SONG Yong-Chang
Received:2004-01-15
Accepted:2004-07-24
Online:2005-01-15
Published:2005-05-30
摘要:
选用节间长、节数和支持木大小作为攀援能力的指标, 单株攀援的支持木种类、数量及水平扩展面积作为单株攀援效率的指标, 对浙江天童常绿阔叶林中 5类 12种大中型木质藤本的攀援能力及生态适应性和其中 2种藤本单株攀援效率进行了分析, 结果发现 :1) 不同攀援类型的节间长和节数存在显著差异, 对支持木大小要求不同, 反映出攀援能力在类型间的差异 ;没有一类藤本兼具所有优势, 攀援能力是节间长、节数和支持木大小不同组合的结果 ;卷须类的广东蛇葡萄 (Ampelopsiscantoniensis) 和暗色菝葜 (Smilaxlanceifoliavar.opaca) 节间最长、节数居中、要求较小支持木 ;枝卷类的香港黄檀 (Dalbergiamillettii) 节数和支持木大小与卷须类接近, 但节间较短 ;不定根类的中华常春藤 (Hederanepalensisvar.sinensis) 、扶芳藤 (Euonymusfortunei) 、薜荔 (Ficuspumila) 和络石 (Trachlospermumjasmi noides) 以及吸盘类的异叶爬山虎 (Parthnocissusheterophylla) 节间最短、但节数最多, 要求较大支持木 ;茎缠绕类的白花野木瓜 (Stauntonialeucantha) 、羊角藤 (Morindaumbellata) 、忍冬 (Lonicerajaponica) 和薯莨 (Dioscoreacirrhosa) 节间长居中, 节数最少, 要求中等但幅度较宽的支持木。 2 ) 同类型不同藤本的节间长、节数和支持木大小的差异反映出由非亲缘关系植物演化形成的藤本具有自身特性的一面 ;同种藤本个体间差异反映出植物的可塑性特点, 是个体对群落内异质生境响应的结果。 3) 广东蛇葡萄单株平均攀援支持木 (乔木 ) 种类、株数和水平扩展面积大于香港黄檀, 反映出前者单株攀援效率大于后者 ;两种藤本的单株攀援效率不与年龄呈正相关, 反映出影响因素的复杂性 ;单株攀援效率可用来指示它们在群落中作用大小。
蔡永立, 宋永昌. 浙江天童常绿阔叶林藤本植物的适应生态学研究 II.攀援能力和单株攀援效率. 植物生态学报, 2005, 29(3): 386-393. DOI: 10.17521/cjpe.2005.0051
CAI Yong-Li, SONG Yong-Chang. ADAPTIVE ECOLOGY OF LIANAS IN AN EVERGREEN BROAD-LEAVED FOREST OF TIANTONG NATIONAL FOREST PARK, ZHEJIANG II. CLIMBING CAPACITY AND CLIMBING EFFICIENCY. Chinese Journal of Plant Ecology, 2005, 29(3): 386-393. DOI: 10.17521/cjpe.2005.0051
| 种类 (编码) Species (Code) | 攀援机制 Climbing mechanics | 生活型 Life_type | 在群落中的分布 Distribution in community |
|---|---|---|---|
| 羊角藤 (MU) Morinda umbellate | 茎缠绕 Stem_twining | 中型常绿木质藤本 Medium EL | 乔木中下层 Under canopy |
| 薯莨 (DC) Dioscorea cirrhosa | 茎缠绕 Stem_twining | 中型常绿木质藤本 Medium EL | 乔木亚层 In subcanopy |
| 白花野木瓜 (SL) Stauntonia leucantha | 茎缠绕 Stem_twining | 大中型落叶木质藤本 Big_medium DL | 冠层之上 Above canopy |
| 忍冬 (LJ) Lonicera japonica | 茎缠绕 Stem_twining | 中型半常绿木质藤本 Medium HEL | 林缘 At edge |
| 络石 (TJ) Trachlospermum jasminoides | 不定根攀援 Adventitious root_climbing | 中型常绿木质藤本 Medium EL | 冠层内 In canopy |
| 扶芳藤 (EF) Euonymus fortunei | 不定根攀援 Adventitious root_climbing | 大型常绿木质藤本 Big EL | 冠层内、石壁 In canopy, on stone |
| 中华常春藤 (HS) Hedera nepalensis var. sinensis | 不定根攀援 Adventitious root_climbing | 大型常绿木质藤本 Big EL | 冠层内 In canopy |
| 薜荔 (FP) Ficus pumila | 不定根攀援 Adventitious root_climbing | 大型常绿木质藤本 Big EL | 冠层内、林缘 In canopy, at edge |
| 异叶爬山虎 (PH) Parthenocissus heterophylla | 吸盘攀援 Adhesive disc_climbed | 大型木质藤本 Big EL | 冠层内、林缘 In canopy, at edge |
| 暗色菝葜 (SO) Smilax lanceiofolia var.opaca | 卷须攀援 Tendril_curled | 大中型常绿木质藤本 Big_medium EL | 冠层之上 Above canopy |
| 广东蛇葡萄 (AC) Ampelopsis cantoniensis | 卷须攀援 Tendril_curled | 大型落叶木质藤本 Big DL | 冠层之上 Above canopy |
| 香港黄檀 (DM) Dalbergia millettii | 小枝攀援 Branch_curled | 大型落叶木质藤本 Big DL | 冠层之上 Above canopy |
表1 浙江天童常绿阔叶林12种木质藤本的概况
Table 1 Summary of 12 species of lianas in Tiantong National Forest Park, Zhejiang
| 种类 (编码) Species (Code) | 攀援机制 Climbing mechanics | 生活型 Life_type | 在群落中的分布 Distribution in community |
|---|---|---|---|
| 羊角藤 (MU) Morinda umbellate | 茎缠绕 Stem_twining | 中型常绿木质藤本 Medium EL | 乔木中下层 Under canopy |
| 薯莨 (DC) Dioscorea cirrhosa | 茎缠绕 Stem_twining | 中型常绿木质藤本 Medium EL | 乔木亚层 In subcanopy |
| 白花野木瓜 (SL) Stauntonia leucantha | 茎缠绕 Stem_twining | 大中型落叶木质藤本 Big_medium DL | 冠层之上 Above canopy |
| 忍冬 (LJ) Lonicera japonica | 茎缠绕 Stem_twining | 中型半常绿木质藤本 Medium HEL | 林缘 At edge |
| 络石 (TJ) Trachlospermum jasminoides | 不定根攀援 Adventitious root_climbing | 中型常绿木质藤本 Medium EL | 冠层内 In canopy |
| 扶芳藤 (EF) Euonymus fortunei | 不定根攀援 Adventitious root_climbing | 大型常绿木质藤本 Big EL | 冠层内、石壁 In canopy, on stone |
| 中华常春藤 (HS) Hedera nepalensis var. sinensis | 不定根攀援 Adventitious root_climbing | 大型常绿木质藤本 Big EL | 冠层内 In canopy |
| 薜荔 (FP) Ficus pumila | 不定根攀援 Adventitious root_climbing | 大型常绿木质藤本 Big EL | 冠层内、林缘 In canopy, at edge |
| 异叶爬山虎 (PH) Parthenocissus heterophylla | 吸盘攀援 Adhesive disc_climbed | 大型木质藤本 Big EL | 冠层内、林缘 In canopy, at edge |
| 暗色菝葜 (SO) Smilax lanceiofolia var.opaca | 卷须攀援 Tendril_curled | 大中型常绿木质藤本 Big_medium EL | 冠层之上 Above canopy |
| 广东蛇葡萄 (AC) Ampelopsis cantoniensis | 卷须攀援 Tendril_curled | 大型落叶木质藤本 Big DL | 冠层之上 Above canopy |
| 香港黄檀 (DM) Dalbergia millettii | 小枝攀援 Branch_curled | 大型落叶木质藤本 Big DL | 冠层之上 Above canopy |
| 种类 Species | 节间长度 Intemode length (cm) Mean±SD | 变异幅度 CV (%) | 最大值 Maximum (cm) | 节数 No. of internodes Mean±SD | 变异幅度 CV (%) | 最大值 Maximum (cm) |
|---|---|---|---|---|---|---|
| 茎缠绕 Stem_twinning | ||||||
| 羊角藤MU | 14.0±5.3a | 37.9 | 24.0 | 23±12 | 52.0 | 38 |
| 薯莨DC | 10.2±2.8ab | 27.5 | 19.0 | 19±7 | 36.8 | 27 |
| 白花野木瓜SL | 7.9±1.6b | 20.3 | 11.3 | 15±4a | 26.7 | 26 |
| 忍冬LJ | 5.1±2.1c | 41.2 | 7.5 | 21±5 | 23.8 | 28 |
| 平均Mean | 9.3±3.8C | 40.9 | 20±3B | 15.0 | ||
| 不定根攀援 Adventitious root_climbing | ||||||
| 络石TJ | 6.7±2.0a | 29.9 | 11.1 | 24±8a | 33.3 | 34 |
| 扶芳藤EF | 3.5±1.0b | 28.6 | 5.0 | 29±7a | 24.1 | 39 |
| 中华常春藤HS | 2.5±0.6b | 24.0 | 3.8 | 52±5b | 9.6 | 68 |
| 薜荔FP | 1.9±0.7c | 36.8 | 3.6 | 42±16b | 38.1 | 66 |
| 平均Mean | 3.7±2.1B, D | 56.8 | 37±12C | 32.4 | ||
| 吸盘攀援 Adhesive disc_climbing | ||||||
| 异叶爬山虎PH | 1.9±0.5B, D | 26.3 | 2.2 | 65±4C | 6.2 | 90 |
| 卷须攀援 Tendril curling | ||||||
| 暗色菝葜SO | 23.1±5.3a | 22.9 | 33.5 | 27±8 | 29.6 | 35 |
| 广东蛇葡萄AC | 14.8±6.0b | 40.5 | 25.5 | 34±16 | 47.1 | 54 |
| 平均Mean | 19.0±5.9A | 31.1 | 31±5A | 15.8 | ||
| 小枝攀援 Branch curling | ||||||
| 香港黄檀DM | 4.6±1.2B | 26.1 | 8.5 | 25±6A, B | 24.0 | 39 |
| 总平均Total mean | 7.7±6.8 | 88.3 | 36±16 | 44.4 |
表2 浙江天童5类12种木质藤本攀援茎形态的比较
Table 2 Comparison of internode length and number of climbing branches among 12 lianas belonged to five climbing mechanics in Tiantong National Forest Park, Zhejiang
| 种类 Species | 节间长度 Intemode length (cm) Mean±SD | 变异幅度 CV (%) | 最大值 Maximum (cm) | 节数 No. of internodes Mean±SD | 变异幅度 CV (%) | 最大值 Maximum (cm) |
|---|---|---|---|---|---|---|
| 茎缠绕 Stem_twinning | ||||||
| 羊角藤MU | 14.0±5.3a | 37.9 | 24.0 | 23±12 | 52.0 | 38 |
| 薯莨DC | 10.2±2.8ab | 27.5 | 19.0 | 19±7 | 36.8 | 27 |
| 白花野木瓜SL | 7.9±1.6b | 20.3 | 11.3 | 15±4a | 26.7 | 26 |
| 忍冬LJ | 5.1±2.1c | 41.2 | 7.5 | 21±5 | 23.8 | 28 |
| 平均Mean | 9.3±3.8C | 40.9 | 20±3B | 15.0 | ||
| 不定根攀援 Adventitious root_climbing | ||||||
| 络石TJ | 6.7±2.0a | 29.9 | 11.1 | 24±8a | 33.3 | 34 |
| 扶芳藤EF | 3.5±1.0b | 28.6 | 5.0 | 29±7a | 24.1 | 39 |
| 中华常春藤HS | 2.5±0.6b | 24.0 | 3.8 | 52±5b | 9.6 | 68 |
| 薜荔FP | 1.9±0.7c | 36.8 | 3.6 | 42±16b | 38.1 | 66 |
| 平均Mean | 3.7±2.1B, D | 56.8 | 37±12C | 32.4 | ||
| 吸盘攀援 Adhesive disc_climbing | ||||||
| 异叶爬山虎PH | 1.9±0.5B, D | 26.3 | 2.2 | 65±4C | 6.2 | 90 |
| 卷须攀援 Tendril curling | ||||||
| 暗色菝葜SO | 23.1±5.3a | 22.9 | 33.5 | 27±8 | 29.6 | 35 |
| 广东蛇葡萄AC | 14.8±6.0b | 40.5 | 25.5 | 34±16 | 47.1 | 54 |
| 平均Mean | 19.0±5.9A | 31.1 | 31±5A | 15.8 | ||
| 小枝攀援 Branch curling | ||||||
| 香港黄檀DM | 4.6±1.2B | 26.1 | 8.5 | 25±6A, B | 24.0 | 39 |
| 总平均Total mean | 7.7±6.8 | 88.3 | 36±16 | 44.4 |
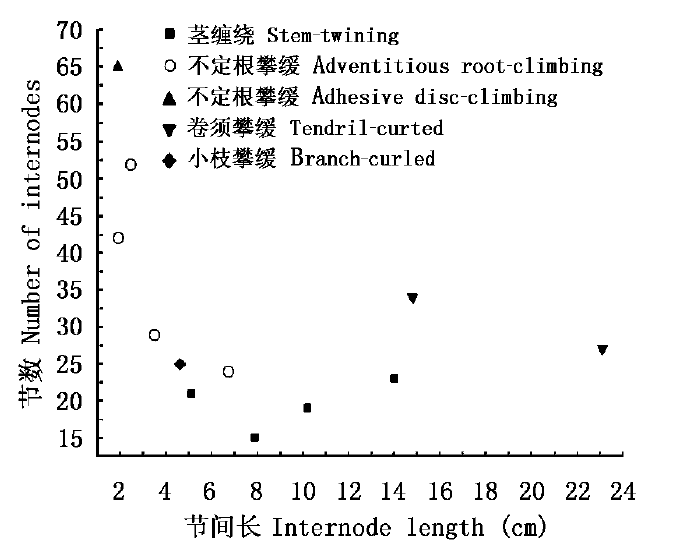
图1 攀援茎节间长与节数之间的相关性
Fig.1 Correlation between mean internode length and number of internodes
| 种类 Species | 样本数 No. of samples | 支持木直径 (cm) Support diameter | 种类 Species | 样本数 No. of samples | 支持木直径 (cm) Support diameter |
|---|---|---|---|---|---|
| 茎缠绕 Stem_twining | 薜荔FP | 50 | > 4.8 | ||
| 薯莨DC | 150 | 0.6~30.0 | 吸盘攀援 Adhesive disc-climbing | ||
| 白花野木瓜SL | 150 | 0.3~11.0 | 异叶爬山虎PH | 45 | > 2.0 |
| 羊角藤MU | 150 | 0.9~11.0 | 卷须攀援 Tendril_curled | ||
| 忍冬LJ | 65 | 0.5~8.0 | 广东蛇葡萄AC | 150 | 0.8~1.4 |
| 不定根攀援 Adventitious root-climbing | 暗色拔葜SO | 150 | 0.8~2.3 | ||
| 络石TJ | 150 | 4.0~54.0 | 小枝攀援 Branch curled | ||
| 扶芳藤EF | 45 | > 5.3 | 香港黄檀DM | 150 | 0.9~1.2 |
| 中华常春藤HS | 150 | > 4.5 |
表3 不同攀援类型藤本对支持木直径大小的要求
Table 3 Sizes of support diameters required by different lianas
| 种类 Species | 样本数 No. of samples | 支持木直径 (cm) Support diameter | 种类 Species | 样本数 No. of samples | 支持木直径 (cm) Support diameter |
|---|---|---|---|---|---|
| 茎缠绕 Stem_twining | 薜荔FP | 50 | > 4.8 | ||
| 薯莨DC | 150 | 0.6~30.0 | 吸盘攀援 Adhesive disc-climbing | ||
| 白花野木瓜SL | 150 | 0.3~11.0 | 异叶爬山虎PH | 45 | > 2.0 |
| 羊角藤MU | 150 | 0.9~11.0 | 卷须攀援 Tendril_curled | ||
| 忍冬LJ | 65 | 0.5~8.0 | 广东蛇葡萄AC | 150 | 0.8~1.4 |
| 不定根攀援 Adventitious root-climbing | 暗色拔葜SO | 150 | 0.8~2.3 | ||
| 络石TJ | 150 | 4.0~54.0 | 小枝攀援 Branch curled | ||
| 扶芳藤EF | 45 | > 5.3 | 香港黄檀DM | 150 | 0.9~1.2 |
| 中华常春藤HS | 150 | > 4.5 |
| 种名 Species | 样本数 No. of samples | 藤本直径 DBH of individual | 支持木种类 Kinds of trellis | 支持木数量 No. of trellis | 水平扩展面积 Horizontal extensive area (m2) |
|---|---|---|---|---|---|
| 香港黄檀 DM | 17 | 4.9±1.1 | 3.2±1.2 | 4.9+±3.1 | 42.5±15.1 |
| 广东蛇葡萄AC | 15 | 6.4±1.5 | 4.8±2.9 | 8.4±6.2 | 119.7±160.9 |
表4 广东蛇葡萄和香港黄檀单株攀援效率比较
Table 4 Comparison of climbing efficiency between individual AC liana and individual DM liana
| 种名 Species | 样本数 No. of samples | 藤本直径 DBH of individual | 支持木种类 Kinds of trellis | 支持木数量 No. of trellis | 水平扩展面积 Horizontal extensive area (m2) |
|---|---|---|---|---|---|
| 香港黄檀 DM | 17 | 4.9±1.1 | 3.2±1.2 | 4.9+±3.1 | 42.5±15.1 |
| 广东蛇葡萄AC | 15 | 6.4±1.5 | 4.8±2.9 | 8.4±6.2 | 119.7±160.9 |
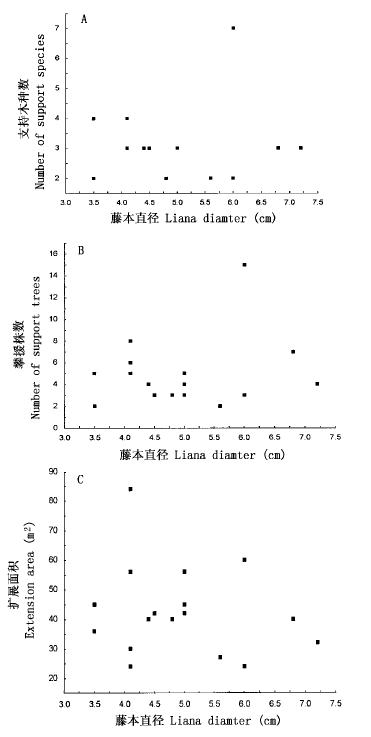
图2 香港黄檀单株攀援效率
Fig.2 Climbing efficiency of individual Dalbergia millettii liana
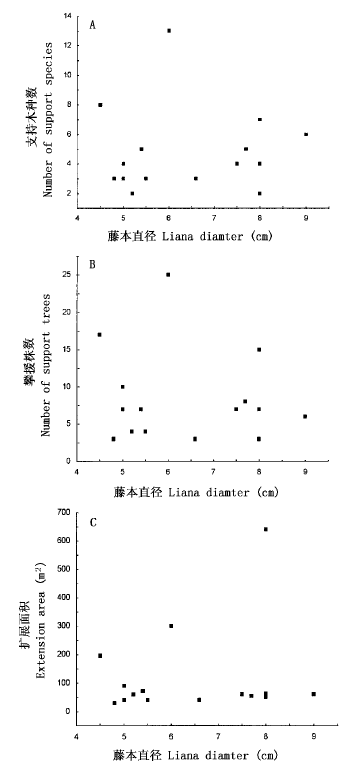
图3 广东蛇葡萄单株攀援效率
Fig.3 Climbing efficiency of individual Ampelopsis cantoniensis liana
| [1] | Bradshaw AD (1965). Evolutionarysignificanceofphenotypicplas ticityinplants. AdvancedGenetics, 13,115-155. |
| [2] | Caballe G (1984). Essaisurladynamiquedespeuplementsdelianesligneusesd'uneforetduNord_EstduGabon. Revued'E cologie (TerreetVie), 39,3-35. |
| [3] | Cai YL (蔡永立), Song YC (宋永昌) (2000). Therevisionofvinelifeformsystemandanalysisoflifeformsofvinesinthesub tropicalzoneofEastChina (SZEC). ActaEcologicaSinica (生态学报), 20,808-814. (inChinesewithEnglishabstract). |
| [4] | Cai YL (蔡永立), Song YC (宋永昌) (2001a). Adaptiveecolo gyoflianasinTiantongevergreenbroad_leavedforest, Zhejiang, ChinaI. Leafanatomicalcharacters.ActaPhytoecologicaSinica (植物生态学报), 25,90-98. (inChinesewithEnglishab stract). |
| [5] | Cai YL (蔡永立), Song YC (宋永昌) (2001b). Adaptivechar acteristicsandbehaviorofDalbergiamillettiilianainsubtropicalevergreenbroad_leavedforestofeasternChina. ActaEcologicaSinica (生态学报), 21,216-224. (inChinesewithEnglishabstract). |
| [6] |
Castellanos RL, Durn R, Guzman S, Briones O, Feria M (1992). Three_dimensionalspaceutilizationoflianas:amethodology. Biotropica, 24,396-401.
DOI URL |
| [7] | Darwin C (1867). Onthemovementsandhabitsofclimbingplants. JournaloftheLinneanSociety (Botany), 9,1-118. |
| [8] | denDubbelden KC, Oosterheek B (1995). Theavailabilityofex ternalsupportaffectspatternsandmorphologyinherbaceousclimbingplants. FunctionalEcology, 9,628-634. |
| [9] | Larson KC (2000). Circumnutationbehaviorofanexotichoneysucklevineanditsnativecongener:influenceonclonalmobility. AmericanJournalofBotany, 87,533-538. |
| [10] | Pealosa J (1983). ShootdynamicsandadaptivemorphologyofIpomoeaphillomega (Vell). House (Convolvulaceae), atropicalrainforestliana.AnnalsofBotany, 52,737-754. |
| [11] |
Putz FE (1984). ThenaturalhistoryoflianasonBarroColoradoIs land, Panama. Ecology, 65,1713-1724.
DOI URL |
| [12] | Putz FE, Mooney HA (1991). TheBiologyofVines.CambridgePress, Cambridge,1-353. |
| [13] | Qu ZX (曲仲湘) (1964). ApreliminarystudyonthetwinningvinesinsouthernmountainforestofChina. ActaPhytoecologica&GeobotanicaSinica (植物生态学与地植物学丛刊), 2,1-9. (inChinesewithEnglishabstract). |
| [14] | Ray TS (1992). Foragingbehaviorintropicalherbaceousclimbers (Araceae). JournalofEcology, 80,189-203. |
| [15] | Richards PW (1952). TropicalRainForest.CambridgeUniversityPress, London,102-108. |
| [16] | Scheiner SM (1993). Geneticsandevolutionofphenotypicplasticity. AnnualReviewofEcology&Systematics, 24,35-68. |
| [17] | Schlichting CD (1986). Theevolutionofphenotypicplasticityinplants. AnnualReviewofEcology&Systematics, 17,667-693. |
| [18] |
Strong DR, Ray TS (1975). Hosttreelocationbehaviorofatropi calvine (Monsteragigantea) byskototropism. Science, 190,804-806.
DOI URL |
| [19] | Tao JP (陶建平), Zhong ZC (钟章成) (2003). Effectsofsup portanglesonthemorphologyandgrowthintheherbaceousclimberTrichosantheskirilowii. ActaEcologicaSinica (生态学报), 23,1-7. (inChinesewithEnglishabstract). |
| [20] |
Tao JP (陶建平), Zhong ZC (钟章成), Huang L (黄林) (2003). TheresponsesofMomordicacharantiaatdifferentmod ularlevelstothechangesofsupportdiameter. ActaPhytoecologi caSinica (植物生态学报), 27,86-92. (inChinesewithEnglishabstract).
DOI |
| [1] | 冯珊珊, 黄春晖, 唐梦云, 蒋维昕, 白天道. 细叶云南松针叶形态和显微性状地理变异及其环境解释[J]. 植物生态学报, 2023, 47(8): 1116-1130. |
| [2] | 白天道, 余春兰, 甘泽朝, 赖海荣, 杨隐超, 黄厚宸, 蒋维昕. 细叶云南松种实性状变异与地理气象因子的关联[J]. 植物生态学报, 2020, 44(12): 1224-1235. |
| [3] | 蒋裕良, 李先琨, 郭屹立, 丁涛, 王斌, 向悟生. 广西弄岗喀斯特季节性雨林藤本种子植物多样性及繁殖习性[J]. 植物生态学报, 2017, 41(7): 716-728. |
| [4] | 周晓旋, 蔡玲玲, 傅梅萍, 洪礼伟, 沈英嘉, 李庆顺. 红树植物胎生现象研究进展[J]. 植物生态学报, 2016, 40(12): 1328-1343. |
| [5] | 覃凤飞,李强,崔棹茗,李洪萍,杨智然. 越冬期遮阴条件下3个不同秋眠型紫花苜蓿品种叶片解剖结构与其光生态适应性[J]. 植物生态学报, 2012, 36(4): 333-345. |
| [6] | 江浩, 周国逸, 黄钰辉, 刘世忠, 唐旭利. 南亚热带常绿阔叶林林冠不同部位藤本植物的光合生理特征及其对环境因子的适应[J]. 植物生态学报, 2011, 35(5): 567-576. |
| [7] | 袁春明, 刘文耀, 李小双, 杨国平. 哀牢山湿性常绿阔叶林木质藤本植物地上部分 生物量及其对人为干扰的响应[J]. 植物生态学报, 2009, 33(5): 852-859. |
| [8] | 蔡颖, 关保华, 安树青, 申瑞玲, 蒋金辉, 董蕾. 克隆植物乌菱对底泥磷含量及植株密度的表型可塑性响应[J]. 植物生态学报, 2007, 31(4): 599-606. |
| [9] | 毛建丰, 李悦, 刘玉军, 刘灏, 王晓茹. 高山松种实性状与生殖适应性[J]. 植物生态学报, 2007, 31(2): 291-299. |
| [10] | 杨九艳, 杨劼, 杨明博, 梁娜, 孔维雪. 鄂尔多斯高原锦鸡儿属植物叶表皮特征及生态适应性[J]. 植物生态学报, 2005, 29(6): 961-967. |
| [11] | 蔡永立, 宋永昌. 浙江天童常绿阔叶林藤本植物的适应生态学I. 叶片解剖特征的比较[J]. 植物生态学报, 2001, 25(1): 90-98. |
| [12] | 贺善安, 刘友良, 郝日明, 钱俊秋. 鹅掌楸种群间光生态适应性的分化[J]. 植物生态学报, 1999, 23(1): 40-47. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
Copyright © 2022 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19