
植物生态学报 ›› 2018, Vol. 42 ›› Issue (2): 164-172.DOI: 10.17521/cjpe.2017.0203
所属专题: 全球变化与生态系统
张鑫1,邢亚娟1,2,闫国永1,王庆贵1,*( )
)
出版日期:2018-02-20
发布日期:2018-04-16
通讯作者:
王庆贵
基金资助:
ZHANG Xin1,XING Ya-Juan1,2,YAN Guo-Yong1,WANG Qing-Gui1,*()
Online:2018-02-20
Published:2018-04-16
Contact:
Qing-Gui WANG
Supported by:摘要:
细根对土壤水分含量变化十分敏感, 增加和减少降水直接影响土壤水分含量。为探索细根对降水变化的响应, 该文从48篇已发表的国内外研究论文中搜集到202组数据, 通过meta分析的方法揭示细根生物量、生产量、周转率、根长度密度、比根长及细根分解对增加和减少降水的一般响应规律, 用加权响应比评价降水对细根各指标的影响效应, 降水变化对细根分解的影响用土壤微生物生物量碳的响应比衡量。结果表明: 1)不同类型植物的细根对降水变化的响应程度不同, 灌木细根的响应强于乔木。2)细根各指标对降水变化的响应存在土层空间异质性, 并且降水变化量为50%时细根响应最显著。降水增加50%时, 显著增加20-40 cm土层的细根生物量和0-10 cm土层的细根比根长, 降水减少50%时, 显著减少20-40 cm土层的细根生产量和增加0-10 cm土层的细根根长度密度。3)降水变化实验持续时间的长短会影响细根的响应程度, 短期实验中细根通过形态适应对降水变化做出应对, 而长期实验中细根通过重新分配生物量对降水变化做出响应。4)增加降水促进了细根养分归还, 致使土壤微生物得到了充足的底物资源, 提高了自身活性, 使细根分解加快。
张鑫, 邢亚娟, 闫国永, 王庆贵. 细根对降水变化响应的meta分析. 植物生态学报, 2018, 42(2): 164-172. DOI: 10.17521/cjpe.2017.0203
ZHANG Xin, XING Ya-Juan, YAN Guo-Yong, WANG Qing-Gui. Response of fine roots to precipitation change: A meta-analysis. Chinese Journal of Plant Ecology, 2018, 42(2): 164-172. DOI: 10.17521/cjpe.2017.0203
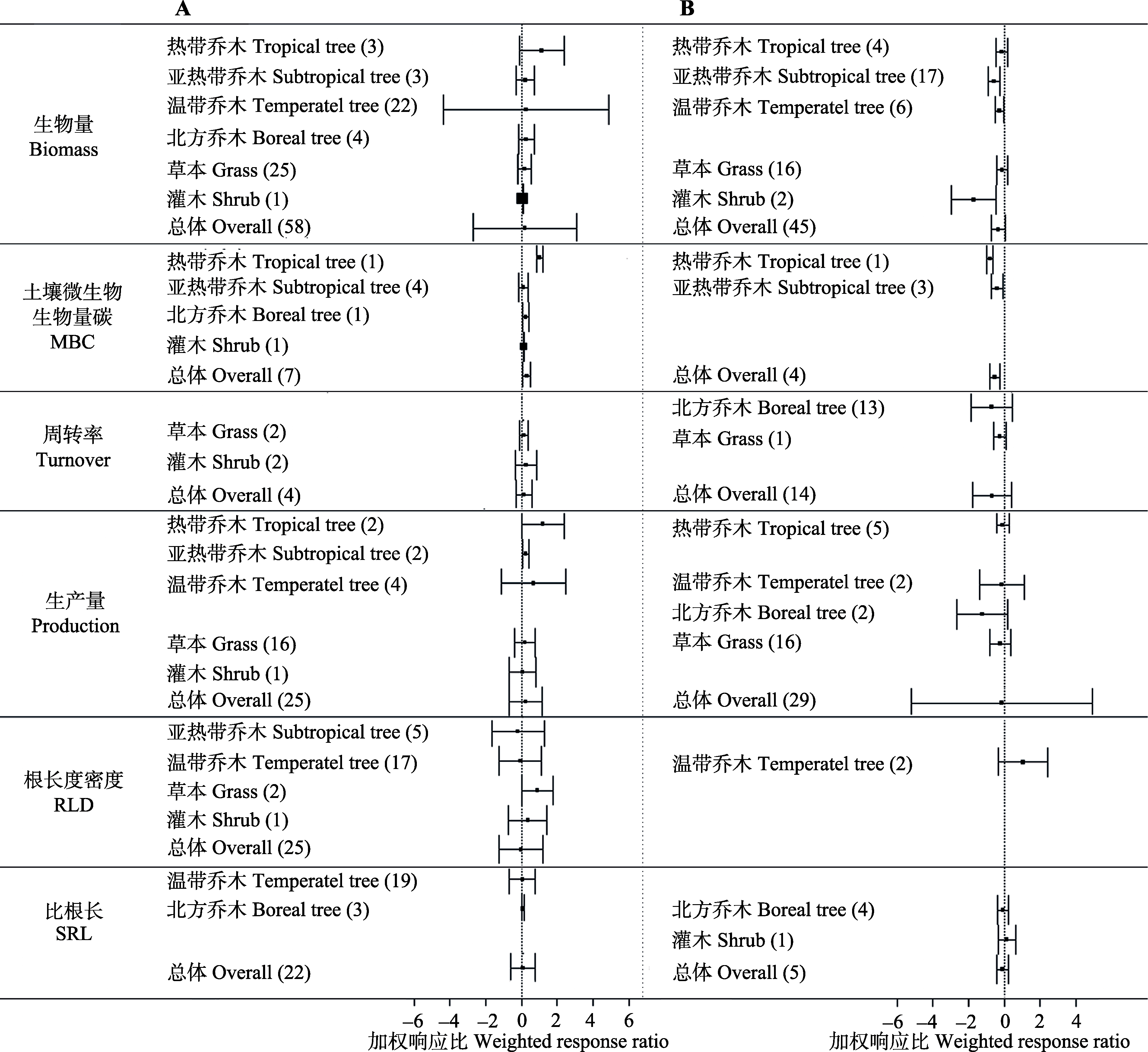
图1 不同类型植物细根和土壤微生物生物量碳对增加(A)或减少(B)降水的加权响应比。括号内数字代表样本量, 误差线代表95%置信区间。
Fig. 1 Weighted response ratio of increasing (A) or reducing (B) precipitation on fine root of different plant type and soil microbial biomass carbon. The variables are categorized into different groups depending on plant types. The number in parentheses represents the sample size for each variable. Error bars represent 95% confidence intervals. MBC, soil microbial biomass carbon; RLD, root length density; SRL, specific root length.
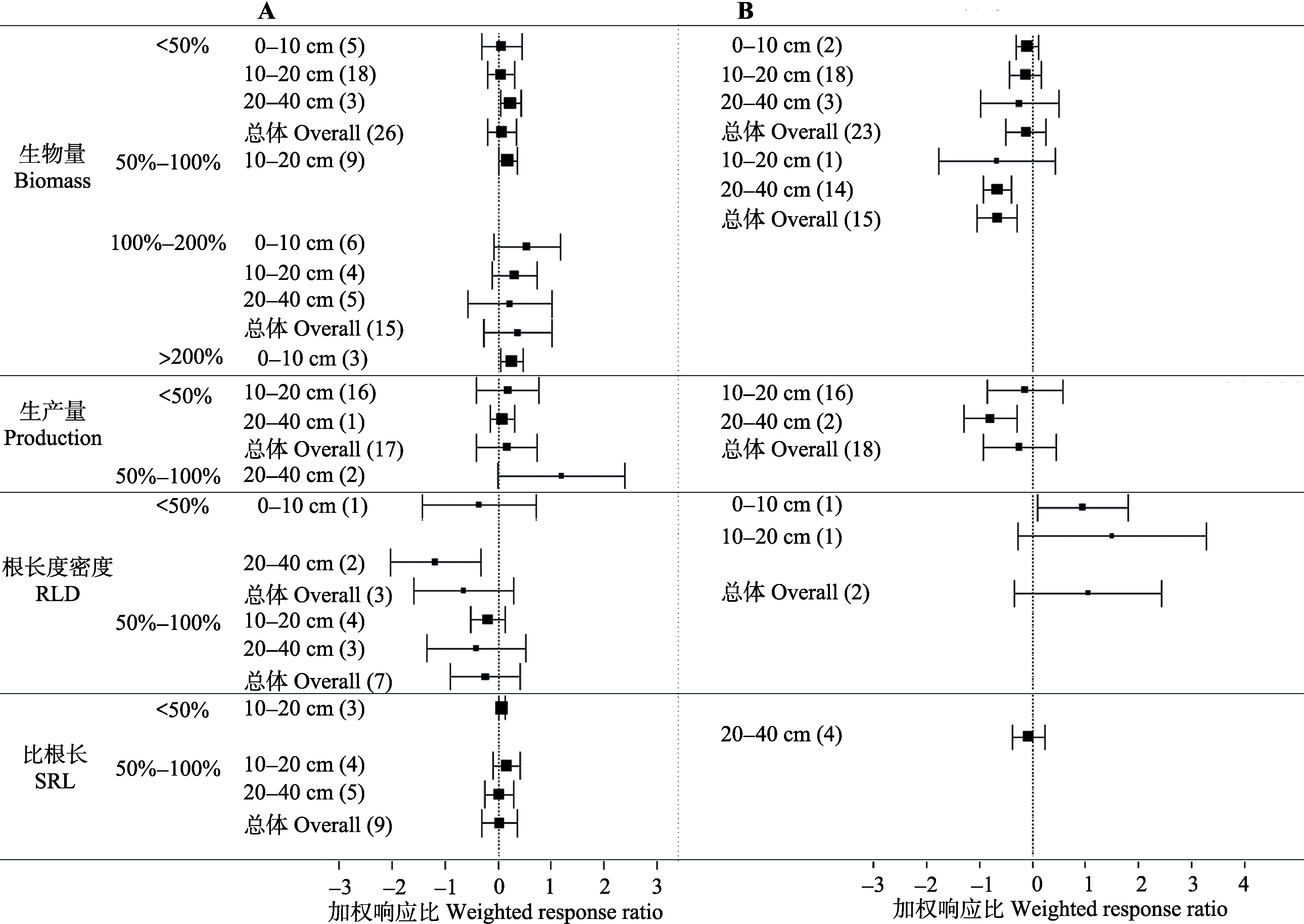
图2 各土层细根指标对不同增加(A)或减少(B)降水量的加权响应比。括号内数字代表样本量, 误差线代表95%置信区间。
Fig. 2 Weighted response ratio (RR++) of different increasing (A) or reducing (B) precipitation amount on each soil layer fine root. The variables are categorized into different groups depending on duration. The number in parentheses represents the sample size for each variable. Error bars represent 95% confidence intervals. RLD, root length density; SRL, specific root length.

图3 不同实验持续时间下细根和土壤微生物生物量碳对增加(A)或减少(B)降水的加权响应比。括号内数字代表样本量, 误差线代表95%置信区间。
Fig. 3 Weighted response ratio (RR++) of increasing (A) or reducing (B) precipitation on fine root and soil microbial biomass carbon under different duration of experiment . The variables are categorized into different groups depending on duration. The number in parentheses represents the sample size for each variable. Error bars represent 95% confidence intervals. MBC, soil microbial biomass carbon; RLD, root length density; SRL, specific root length.
| [1] |
Ansley RJ, Boutton TW, Jacoby PW ( 2014). Root biomass and distribution patterns in a semi-arid mesquite savanna: Responses to long-term rainfall manipulation. Rangeland Ecology & Management, 67, 206-218.
DOI URL |
| [2] |
Bai W, Wan S, Niu S, Liu W, Chen Q, Wang Q, Zhang W, Han X, Li L ( 2010). Increased temperature and precipitation interact to affect root production, mortality, and turnover in a temperate steppe: Implications for ecosystem C cycling. Global Change Biology, 16, 1306-1316.
DOI URL |
| [3] | Berg B, McClaugherty C ( 2003). Plant Litter: Decomposition, Humus Formation, Carbon Sequestration. Springer, Berlin. |
| [4] |
Bloomfield J, Vogt DJ ( 1993). Decay rate and substrate quality of fine roots and foliage of two tropical tree species in the Luquillo Experimental Forest, Puerto Rico. Plant and Soil, 150, 233-245.
DOI URL |
| [5] |
Chang WJ, Guo DL ( 2008). Variation in root diameter among 45 common tree species in temperate, subtropical and tropical forest in China. Journal of Plant Ecology (Chinese Version), 32, 1248-1257.
DOI URL |
|
[ 常文静, 郭大立 ( 2008). 中国温带、亚热带和热带森林45个常见树种细根直径变异. 植物生态学报, 32, 1248-1257.]
DOI URL |
|
| [6] | Chapin FSI, Matson PAI, Mooney HA ( 2002). Principles of Terrestrial Ecosystem Ecology. Springer-Verlag, New York. |
| [7] |
Chen G, Yang Y, Robinson D ( 2013). Allocation of gross primary production in forest ecosystems: Allometric constraints and environmental responses. New Phytologist, 200, 1176-1186.
DOI URL PMID |
| [8] | Chen MT, Zhao Z ( 2011). Effect of drought on root characteristics and mass allocation in each part of seedlings of four tree species. Journal of Beijing Forest University, 33(1), 16-22. |
| [ 陈明涛, 赵忠 ( 2011). 干旱对四种苗木根系特征及各部分物质分配的影响. 北京林业大学学报, 33(1), 16-22.] | |
| [9] | Chen X, Zhang D, Liang G, Qiu Q, Liu J, Zhou G, Liu J, Zhou G, Liu S, Chu G, Yan J ( 2015). Effects of precipitation on soil organic carbon fractions in three subtropical forests in southern China. Journal of Plant Ecology, 9, 10-19. |
| [10] |
Coleman M ( 2007). Spatial and temporal patterns of root distribution in developing stands of four woody crop species grown with drip irrigation and fertilization. Plant and Soil, 299, 195-213.
DOI URL |
| [11] |
de Visser PHB, Beier C, Rasmussen L, Kreutzer K, Steinberg N, Bredemeier M, Blanck K, Farrell EP, Cummins T ( 1994). Biological response of ?ve forest ecosystems in the EXMAN project to input changes of water, nutrients and atmospheric loads. Forest Ecology, 68, 15-29.
DOI URL |
| [12] |
Dong BF ( 2015). Root distribution characteristics of three kinds of forest lands in loess hilly region of northern Shaanxi. Journal of Changjiang Engineering Vocational College, 4, 24-26.
DOI URL |
|
[ 董宾芳 ( 2015). 陕北黄土丘陵区三种林地根系分布特征. 长江工程职业技术学院学报, 4, 24-26.]
DOI URL |
|
| [13] |
Fiala K, T?ma I, Holub P ( 2009). Effect of manipulated rainfall on root production and plant belowground dry mass of different grassland ecosystems. Ecosystems, 12, 906-914.
DOI URL |
| [14] |
Fiala K, T?ma I, Holub P ( 2012). Interannual variation in root production in grasslands affected by artificially modified amount of rainfall. The Scientific World Journal, 2, 805298. DOI: 10.1100/2012/805298.
DOI URL PMID |
| [15] |
Ford CR, McGee J, Scandellari F, Hobbie EA, Mitchell RJ ( 2012). Long- and short-term precipitation effects on soil CO2 efflux and total belowground carbon allocation. Agricultural and Forest Meteorology, 156, 54-64.
DOI URL |
| [16] |
García-Palacios P, Prieto I, Ourcival JM, H?ttenschwiler S ( 2016). Disentangling the litter quality and soil microbial contribution to leaf and fine root litter decomposition responses to reduced rainfall. Ecosystems, 19, 490-503.
DOI URL |
| [17] |
Gei MG, Powers JS ( 2015). The influence of seasonality and species effects on surface fine roots and nodulation in tropical legume tree plantations. Plant and Soil, 388, 187-196.
DOI URL |
| [18] |
Han YY, Ye YH, Wang ZH, Wei LP, Lin L ( 2014). Root biomass, specific root length and root length density of Sophora moorcroftian in Tibet. Journal of Northeast Forestry University, 42(2), 39-41.
DOI URL |
|
[ 韩艳英, 叶彦辉, 王贞红, 魏丽萍, 林玲 ( 2014). 西藏砂生槐根系生物量、比根长和根长密度. 东北林业大学学报, 42(2), 39-41.]
DOI URL |
|
| [19] |
Hedges LV, Gurevitch J, Curtis PS ( 2008). The meta-analysis of response ratios in experimental ecology. Ecology, 80, 1150-1156.
DOI URL |
| [20] |
Herzog C, Steffen J, Pannatier EG, Hajdas I, Brunner I ( 2014). Nine years of irrigation cause vegetation and fine root shifts in a water-limited pine forest. PLOS ONE, 9, e96321. DOI: 10.1371/journal.pone.0096321.
DOI URL PMID |
| [21] |
Hertel D, Strecker T, Müller-Haubold H, Leuschner C ( 2013). Fine root biomass and dynamics in beech forests across a precipitation gradient—Is optimal resource partitioning theory applicable to water-limited mature trees? Journal of Ecology, 101, 1183-1200.
DOI URL |
| [22] |
Hu JZ, Zheng JL, Shen JY ( 2005). Discussion of root ecological niche and root distribution characteristics of artificial phyto-communities in rehabilitated fields. Acta Ecologica Sinica, 25, 481-490.
DOI URL |
|
[ 胡建忠, 郑佳丽, 沈晶玉 ( 2005). 退耕地人工植物群落根系生态位及其分布特征. 生态学报, 25, 481-490.]
DOI URL |
|
| [23] | Imada S, Taniguchi T, Acharya K, Yamanaka N ( 2013). Vertical distribution of fine roots of Tamarix ramosissima in an arid region of southern Nevada. Journal of Arid Environments, 92(3), 46-52. |
| [24] | IPCC (Intergovernmental Panel on Climate Change) (2013). Contribution of working group 1 to the fifth assessment report of the intergovernmental panel on climate change. In: Stocker TF, Qin DPlattner G-K, Tignor M, Allen SK, Boschung J, Nauels A, Xia Y eds. Climate Change in 2013: The Physical Science Basis. Cambridge University Press Cambridge, UK. |
| [25] |
Jerbi A, Nissim WG, Fluet R, Labrecque M ( 2015). Willow root development and morphology changes under different irrigation and fertilization regimes in a vegetation filter. Bioenergy Research, 8, 775-787.
DOI URL |
| [26] |
Jiang H, Bai Y, Du H, Hu Y, Rao Y, Chen C, Cai Y ( 2016). The spatial and seasonal variation characteristics of fine roots in different plant configuration modes in new reclamation saline soil of humid climate in China. Ecological Engineering, 86, 231-238.
DOI URL |
| [27] |
Kong DL, Lü XT, Jiang LL, Wu HF, Miao Y, Kardol P ( 2013). Extreme rainfall events can alter inter-annual biomass responses to water and N enrichment. Biogeosciences, 10, 8129-8138.
DOI URL |
| [28] |
Kon?pka B, Lukac M, Andrea V ( 2012). Moderate drought alters biomass and depth distribution of fine roots in Norway spruce. Forest Pathology, 43, 115-123.
DOI URL |
| [29] |
Larsen KS, Jonasson S, Michelsen A ( 2002). Repeated freeze thaw cycles and their effects on biological processes in two arctic ecosystem types. Applied Soil Ecology, 21, 187-195.
DOI URL |
| [30] |
Li YL, Yang FF, Ou YX, Zhang DQ, Liu JX, Chu GW, Zhang YR, Otieno D, Zhou GY ( 2013). Changes in forest soil properties in different successional stages in lower tropical China. PLOS ONE, 8, e81359. DOI: 10.1371/journal.pone.0081359.
DOI URL PMID |
| [31] | Lin Y, Hu HT, Qiu LJ, Lin SZ, He ZM, Zhang Y, Huang Z, Huang XY ( 2017). Microbial biomass and its influence factors in topsoil of three different plantations on a sandy coastal plain. Journal of Northeast Forestry University, 5, 85-90. |
| [ 林宇, 胡欢甜, 邱岭军, 林思祖, 何宗明, 张勇, 黄政, 黄秀勇 ( 2017). 滨海沙地3种人工林表层土壤微生物量及其影响因素. 东北林业大学学报, 5, 85-90.] | |
| [32] |
Liu Y, Liu S, Wan S, Wang J, Wang H, Liu K ( 2017). Effects of experimental throughfall reduction and soil warming on fine root biomass and its decomposition in a warm temperate oak forest. The Science of the Total Environment, 574, 1448-1455.
DOI URL PMID |
| [33] |
Luo D, Liu S, Shi ZM, Feng QH, Liu QL, Zhang L, Huang Q, He JS ( 2017). Soil microbial community structure in Picea asperata plantations with different ages in subalpine of western Sichuan, Southwest China. Chinese Journal of Applied Ecology, 28, 519-527.
DOI URL |
|
[ 罗达, 刘顺, 史作民, 冯秋红, 刘千里, 张利, 黄泉, 何建社 ( 2017). 川西亚高山不同林龄云杉人工林土壤微生物群落结构. 应用生态学报, 28, 519-527.]
DOI URL |
|
| [34] |
Martin PH, Sherman RE, Fahey TJ ( 2004). Forty years of tropical forest recovery from agriculture: Structure and floristics of secondary and old-growth riparian forests in the Dominican Republic. Biotropica, 36, 297-317.
DOI URL |
| [35] |
McClaugherthy CA, Aber JD, Melillo JM ( 1982). The role of fine roots in the organic matter and nitrogen budgets of two forested ecosystems. Ecology, 63, 1481-1490.
DOI URL |
| [36] |
McCormack ML, Dickie IA, Eissenstat DM, Fahey TJ, Fernandez CW, Guo D, Helmisaari HS, Hobbie EA, Iversen CM, Jackson RB, Lepp?lammi-Kujansuu J, Norby RJ, Phillips RP, Pregitzer KS, Pritchard SG, Rewald B, Zadworny M ( 2015). Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphere processes. New Phytologist, 207, 505-518.
DOI URL PMID |
| [37] |
Moser G, Leuschner C, Hertel D, H?lscher D, K?hler M, Leitner D, Michalzik B, Prihastanti E, Tjitrosemito S, Schwendenmann L ( 2010). Response of cocoa trees ( Theobroma cacao) to a 13-month desiccation period in Sulawesi, Indonesia. Agroforestry Systems, 79, 171-187.
DOI URL |
| [38] |
Moser G, Schuldt B, Hertel D, Horna V, Coners H, Barus H, Leuschner C ( 2014). Replicated throughfall exclusion experiment in an Indonesian perhumid rainforest: Wood production, litter fall and fine root growth under simulated drought. Global Change Biology, 20, 1481-1497.
DOI URL PMID |
| [39] | Olesinski J, Lavigne MB, Krasowski MJ ( 2011). Effects of soil moisture manipulations on fine root dynamics in a mature balsam fir ( Abies balsamea L. Mill.) forest. Tree Physiology, 31, 339-348. |
| [40] |
Ostertag R, Marín-Spiotta E, Silver WL, Schulten J ( 2008). Litterfall and decomposition in relation to soil carbon pools along a secondary forest chronosequence in Puerto Rico. Ecosystems, 11, 701-714.
DOI URL |
| [41] |
Starr G, Oberbauer SF ( 2008). Photosynthesis of Arctic evergreens under snow implications for tundra ecosystem carbon balance. Ecology, 84, 1415-1420.
DOI URL |
| [42] |
Taylor JP, Wilson B, Mills MS, Burns RG ( 2002). Comparison of microbial numbers and enzymatic activities in surface soils and subsoils using various techniques. Soil Biology & Biochemistry, 34, 387-401.
DOI URL |
| [43] |
Verburg PS, Young AC, Stevenson BA, Glanzmann I, Arnone JA, Marion GM, Holmes C, Nowak RS ( 2013). Do increased summer precipitation and N deposition alter fine root dynamics in a Mojave Desert ecosystem? Global Change Biology, 19, 948-956.
DOI URL PMID |
| [44] | Vogt KA, Vogt DJ, Palmiotto PA, Boon P, O’Hara J, Asbjornsen H ( 1996). Review of root dynamics in forest ecosystems grouped by climate, climatic forest type, and species. Plant and Soil, 187, 159-219. |
| [45] | Wu ZC, Wu FZ, Yang WQ, Wei YY, Wang A, Liu JL ( 2012). Dynamics of soil microbial biomass during early fine roots decomposition of three species in alpine region. Acta Ecologica Sinica, 32, 4094-4102. |
| [ 武志超, 吴福忠, 杨万勤, 魏圆云, 王奥, 刘金玲 ( 2012). 高山森林三种细根分解初期微生物生物量动态. 生态学报, 32, 4094-4102.] | |
| [46] |
Yuan ZY, Chen H ( 2010). Fine root biomass, production, turnover rates, and nutrient contents in boreal forest ecosystems in relation to species, climate, fertility, and stand age: Literature review and meta-analyses. Critical Reviews in Plant Sciences, 29, 204-21.
DOI URL |
| [47] |
Zhang JR, Zhang LY, Liu F, Yao B ( 2014). Research progress in effect of rainfall on soil microbe in arid and semi-arid area. World Forest Research, 27(4), 6-12.
DOI URL |
|
[ 张静茹, 张雷一, 刘方, 姚斌 ( 2014). 降雨对干旱半干旱地区土壤微生物影响研究进展. 世界林业研究, 27(4), 6-12.]
DOI URL |
|
| [48] |
Zhong BY, Xiong DC, Shi SZ, Feng JX, Xu CS, Deng F, Chen YY, Chen GS ( 2016). Effects of precipitation exclusion on fine-root biomass and functional traits of Cunninghamia lanceolata seedlings. Chinese Journal of Applied Ecology, 27, 2807-2814.
DOI URL |
|
[ 钟波元, 熊德成, 史顺增, 冯建新, 许辰森, 邓飞, 陈云玉, 陈光水 ( 2016). 隔离降水对杉木幼苗细根生物量和功能特征的影响. 应用生态学报, 27, 2807-2814.]
DOI URL |
|
| [49] | Zi H, Xiang Z, Wang G, Luji A, Wang C ( 2017). Profile of soil microbial community under different stand types in Qinghai Province. Scientia Silvae Sinicae, 53(3), 21-32. |
| [1] | 张文瑾 佘维维 秦树高 乔艳桂 张宇清. 氮和水分添加对黑沙蒿群落优势植物叶片氮磷化学计量特征的影响[J]. 植物生态学报, 2024, 48(5): 590-600. |
| [2] | 耿雪琪, 唐亚坤, 王丽娜, 邓旭, 张泽凌, 周莹. 氮添加增加中国陆生植物生物量并降低其氮利用效率[J]. 植物生态学报, 2024, 48(2): 147-157. |
| [3] | 李娜, 唐士明, 郭建英, 田茹, 王姗, 胡冰, 罗永红, 徐柱文. 放牧对内蒙古草地植物群落特征影响的meta分析[J]. 植物生态学报, 2023, 47(9): 1256-1269. |
| [4] | 张潇, 武娟娟, 贾国栋, 雷自然, 张龙齐, 刘锐, 吕相融, 代远萌. 降水控制对侧柏液流变化特征及其水分来源的影响[J]. 植物生态学报, 2023, 47(11): 1585-1599. |
| [5] | 魏春雪, 杨璐, 汪金松, 杨家明, 史嘉炜, 田大栓, 周青平, 牛书丽. 实验增温对陆地生态系统根系生物量的影响[J]. 植物生态学报, 2021, 45(11): 1203-1212. |
| [6] | 刘雪飞, 吴林, 王涵, 洪柳, 熊莉军. 鄂西南亚高山湿地泥炭藓的生长与分解[J]. 植物生态学报, 2020, 44(3): 228-235. |
| [7] | 苗百岭, 梁存柱, 史亚博, 梁茂伟, 刘钟龄. 降水变化对内蒙古典型草原地上生物量的影响[J]. 植物生态学报, 2019, 43(7): 557-565. |
| [8] | 顾伟平, 刘瑞鹏, 李兴欢, 孙涛, 张子嘉, 昝鹏, 温璐宁, 马鹏宇, 毛子军. 四个典型温带树种不同根序细根分解速率及其主要影响因素[J]. 植物生态学报, 2018, 42(9): 955-962. |
| [9] | 何为, 杨雪英, 肖娟, 张子良, 蒋铮, 袁远爽, 王东, 刘庆, 尹华军. 氮素富集对青藏高原东缘窄叶鲜卑花灌丛根系分泌物碳输入的影响[J]. 植物生态学报, 2017, 41(6): 610-621. |
| [10] | 杨青霄, 田大栓, 曾辉, 牛书丽. 降水格局改变背景下土壤呼吸变化的主要影响因素及其调控过程[J]. 植物生态学报, 2017, 41(12): 1239-1250. |
| [11] | 陈冠陶, 彭勇, 郑军, 李顺, 彭天驰, 邱细容, 涂利华. 氮添加对亚热带次生常绿阔叶林扁刺栲细根生物量、寿命和形态的短期影响[J]. 植物生态学报, 2017, 41(10): 1041-1050. |
| [12] | 陈云玉, 熊德成, 黄锦学, 王韦韦, 胡双成, 邓飞, 许辰森, 冯建新, 史顺增, 钟波元, 陈光水. 中亚热带不同演替阶段的马尾松和米槠人工林的细根生产量研究[J]. 植物生态学报, 2015, 39(11): 1071-1081. |
| [13] | 杨秀云, 韩有志, 武小钢. 华北落叶松林细根生物量对土壤水分、氮营养空间异质性改变的响应[J]. 植物生态学报, 2012, 36(9): 965-972. |
| [14] | 王慧, 周广胜, 蒋延玲, 石耀辉, 许振柱. 降水与CO2浓度协同作用对短花针茅光合特性的 影响[J]. 植物生态学报, 2012, 36(7): 597-606. |
| [15] | 刘佳, 项文化, 徐晓, 陈瑞, 田大伦, 彭长辉, 方晰. 湖南会同5个亚热带树种的细根构型及功能特征分析[J]. 植物生态学报, 2010, 34(8): 938-945. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
Copyright © 2022 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19