
|
|
||
|
晋西黄土区典型乔灌木短期水分利用效率对环境因子的响应
植物生态学报
2021, 45 (12):
1350-1364.
DOI: 10.17521/cjpe.2021.0220
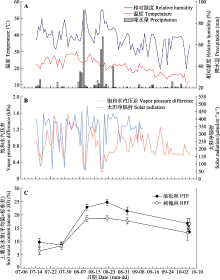
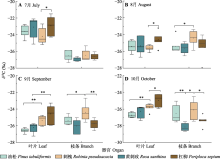
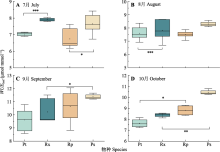
为明确晋西黄土区植物的水分利用规律及对半干旱区的适应策略, 提高黄土地区植被建设效益, 该研究对该地区典型乔灌木短期水分利用效率随环境因子的变化进行了探究。以典型乔木油松(Pinus tabuliformis)、刺槐(Robinia pseudoacacia)及其林下灌木黄刺玫(Rosa xanthina)、杠柳(Periploca sepium)为研究对象, 测定叶片可溶性糖稳定碳同位素比值(δ13Cleaf)与枝条渗出液稳定碳同位素比值(δ13Cbranch), 使用δ13Cleaf推导计算7-10月叶片尺度下植物短期水分利用效率(WUEleaf)变化趋势, 使用δ13Cbranch明确植物光合作用后分馏情况, 确定半干旱区植物在生长季的水分变化规律对环境因子变化的响应。结果表明: (1) 7-10月4种植物δ13Cleaf总体呈现降低趋势, δ13Cbranch呈现先升高后降低趋势。δ13Cleaf在种间和生活型中均存在差异。具体表现为: 灌木>乔木, 常绿乔木(油松) >落叶乔木(刺槐)。研究过程中未发现明显的碳同位素在光合作用后发生分馏的情况。(2) 4种植物WUEleaf在7-8月保持稳定, 9-10月逐渐升高。21.5 ℃、0.9 kPa、52.4%分别为WUEleaf随温度(Ta)、饱和水汽压差(VPD)、相对湿度(RH)变化的突变点, 突变点之后4种植物WUEleaf均表现出稳定的变化趋势, 不再随Ta、VPD、RH升高而降低。(3) WUEleaf与Ta、RH、VPD之间存在显著负相关关系, Ta通过非气孔因素, 即酶的作用改变光合速率, 引起WUEleaf变化。RH、VPD等水分因子则通过改变气孔开度, 影响蒸腾, 进而改变WUEleaf。随着土壤含水量(SWC)的升高, WUEleaf呈现先升高后降低的趋势。油松林和刺槐林在SWC分别达到15%-18%、13%-14%时, WUEleaf达到最高值。经过混合线性模型(LMM)分析得到, 油松和刺槐WUEleaf主导环境因子分别为RH和VPD, 黄刺玫和杠柳WUEleaf主导环境因子均为Ta。该研究得到了黄土地区典型乔灌木生长季水分利用效率变化的规律和主要环境影响因子, 明确了黄土地区植物对气候因子变化的适应机制。 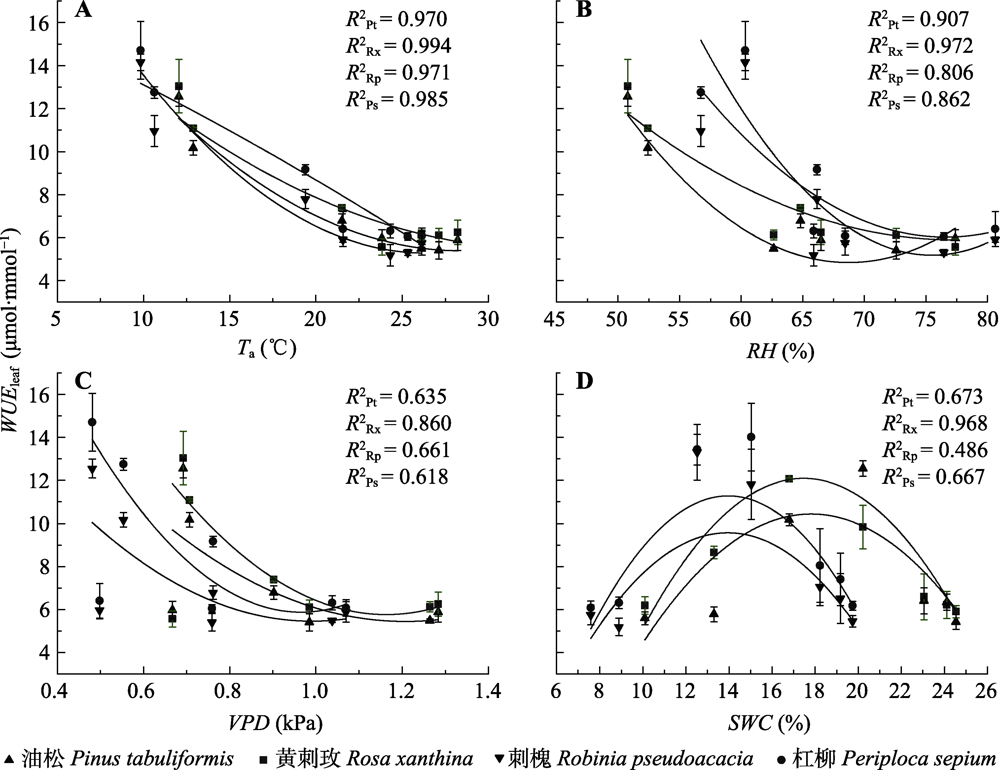 View image in article
图7
晋西黄土区短期水分利用效率(WUEleaf)随日平均气温(Ta)(A)、相对湿度(RH)(B)、饱和水汽压差(VPD)(C)和土壤含水量(SWC)(D)变化趋势(平均值±标准差)。Ps, 杠柳; Pt, 油松; Rp, 刺槐; Rx, 黄刺玫。
正文中引用本图/表的段落
为研究WUEleaf对各环境因子变化的响应, 故量化WUEleaf与各气象因子及土壤含水量之间的关系(图7)。不同植物种WUEleaf与Ta、VPD和RH均存在显著负相关关系(p < 0.05)。经过Mann-Kendall检验得到, 21.5 ℃、0.9 kPa、52.4%分别为WUEleaf随Ta、VPD、RH变化的突变点, 突变点之后4种植物WUEleaf均表现出稳定的变化趋势, 不再随Ta、VPD、RH升高而降低。不同物种WUEleaf随着SWC增加表现为先增加后降低的变化趋势。当SWC极低的时候, 植物WUEleaf保持较低的水平。随后植物WUEleaf随SWC升高而逐渐升高。当SWC达到14%左右时, 刺槐WUEleaf达到最高点(图7D); SWC达到17%左右时, 油松WUEleaf达到最高点(图7D), 随后逐渐降低。
当水分短缺时, 样地内的水热条件等环境因子主导植物WUEleaf变化(图6)。水分因子是影响油松和刺槐两种乔木植物WUEleaf的关键因素(表5)。根据赵丹阳等(2021)对晋西黄土区不同林分土壤水分的研究, 油松林的年内平均土壤储水量大于刺槐林。这说明刺槐林土壤水分状况差且刺槐耗水能力强。吴旭等(2015)也证明刺槐为降雨敏感型植物, 蒸腾耗水对土壤水分的需求大。本研究中刺槐林SWC低于油松林(图1)且林分密度高于油松林(表1), 对水分竞争激烈。为维持植物体内水分平衡和避免碳饥饿现象产生, 在SWC低的情况下, 植物会减少蒸腾耗水, 进而WUE升高(李群等, 2017)。刺槐、杠柳叶片大且薄, 可获得光照面积较大。相比之下, 油松为针叶, 其林下灌木黄刺玫叶片较小, 对光照的接受面积小, 光合速率低, 导致在产生水分胁迫时, 油松样地乔灌木WUEleaf小于刺槐样地乔灌木。以往研究表明植物在干旱的情况下会通过提高自身的WUEleaf以维持自身的生理活动, 降低干旱胁迫带来的损失(张桂玲等, 2021)。然而, 本研究发现植物WUEleaf随SWC变化呈现先增加后降低的变化趋势(图7D)。张永娥等(2017)也发现SWC升高不会必然造成WUE的升高。类似的实现WUEleaf最优值的土壤水分区间在青海云杉(Picea crassifolia)的研究中也得到了证明(童翠芸等, 2020)。由于WUE的变化取决于光合速率与蒸腾速率变化速度的相对大小, 本研究结果说明不同土壤水分条件会造成光合速率和蒸腾速率增长速率的差异, 进而造成WUEleaf的阶段性变化。
线性拟合(图7)及LMM模型(表4)结果显示4种植物WUEleaf对环境因子的响应均存在种间差异。其中, 杠柳、黄刺玫两种灌木植物WUEleaf均受到Ta的显著影响。Ta可以通过影响酶的活性间接影响植物光合速率(路伟伟等, 2018)。同时, Ta也可以通过影响气孔开度影响光合速率。判断光合速率是否受气孔控制要根据Ci与Ls的变化, 且参考Ci的是变化方向, 而不是幅度(曹生奎等, 2009)。气孔关闭会导致Ci下降, 而酶活性下降会导致Ci升高。本研究观测期间Ci变化为先上升后保持稳定(图2), Ls保持下降状态(图5), 故本研究中Ta通过非气孔因素, 也就是酶的作用影响光合速率。随着Ta升高, 光合速率下降, 进而导致WUEleaf降低。相比之下, 油松和刺槐WUEleaf主要受到RH和VPD的影响。谢会成和姜志林(2010)的研究也得到了类似的结论。陈高路等(2021)利用通径分析也得到VPD是乔木WUE主要限制因子的结论。
本文的其它图/表
|



{kind=link}