
植物生态学报 ›› 2018, Vol. 42 ›› Issue (8): 841-849.DOI: 10.17521/cjpe.2018.0043
陈怡超,赵莹,宋希强( ),任明迅()
),任明迅()
收稿日期:2018-02-20
出版日期:2018-08-20
发布日期:2018-12-07
通讯作者:
宋希强,任明迅
基金资助:
CHEN Yi-Chao,ZHAO Ying,SONG Xi-Qiang(),REN Ming-Xun()
Received:2018-02-20
Online:2018-08-20
Published:2018-12-07
Contact:
Xi-Qiang SONG,Ming-Xun REN
Supported by:摘要:
河岸带是河流与陆地生态系统的交错带, 孕育了丰富的生物多样性。河流的冲刷与地势的作用使得河岸带往往形成弯道, 弯道内外两侧水流速度、泥沙性质与植物繁殖体聚集程度不同, 影响到植物生长与种群动态, 可能导致河岸带弯道凸岸、凹岸两侧植物空间分布格局和种群结构存在较大差异。该研究以中国特有、狭域分布的海南杜鹃(Rhododendron hainanense)为例, 揭示溪流弯道对其两侧海南杜鹃种群空间分布格局与种群动态的影响。在海南岛3个国家级自然保护区内各设置2个河岸弯道样带, 用方差/平均值法对弯道凸岸、凹岸两侧海南杜鹃的空间分布格局和种群动态进行对比研究。结果显示: (1)海南杜鹃弯道凸岸的种群结构呈增长型, 凹岸基本呈衰退型; (2)凸岸一侧上下游的海南杜鹃种群基本呈增长型, 凹岸一侧上下游的海南杜鹃则出现断龄现象; (3)距离弯顶位置或河岸越远, 各龄级海南杜鹃种群个体数量基本呈下降趋势; (4)在2 m × 2 m尺度下, 弯道凸岸及其下游的海南杜鹃主要呈聚集分布, 上游近似随机分布, 凹岸及其上下游则均呈随机分布。海南杜鹃分布的山区河岸带水流较快、坡度较大, 可能是导致弯道凹岸个体较少、种群结构呈衰退型的主要原因。因此, 河岸弯道内外两侧可能存在个体生长与群体差异, 在开展河岸带植物种群动态与物种保育研究中需给予重视。
陈怡超, 赵莹, 宋希强, 任明迅. 海南杜鹃在河岸带弯道两侧的空间分布格局和年龄结构差异. 植物生态学报, 2018, 42(8): 841-849. DOI: 10.17521/cjpe.2018.0043
CHEN Yi-Chao, ZHAO Ying, SONG Xi-Qiang, REN Ming-Xun. Difference in spatial distribution patterns and population structures of Rhododendron hainanense between both sides of riparian bends. Chinese Journal of Plant Ecology, 2018, 42(8): 841-849. DOI: 10.17521/cjpe.2018.0043
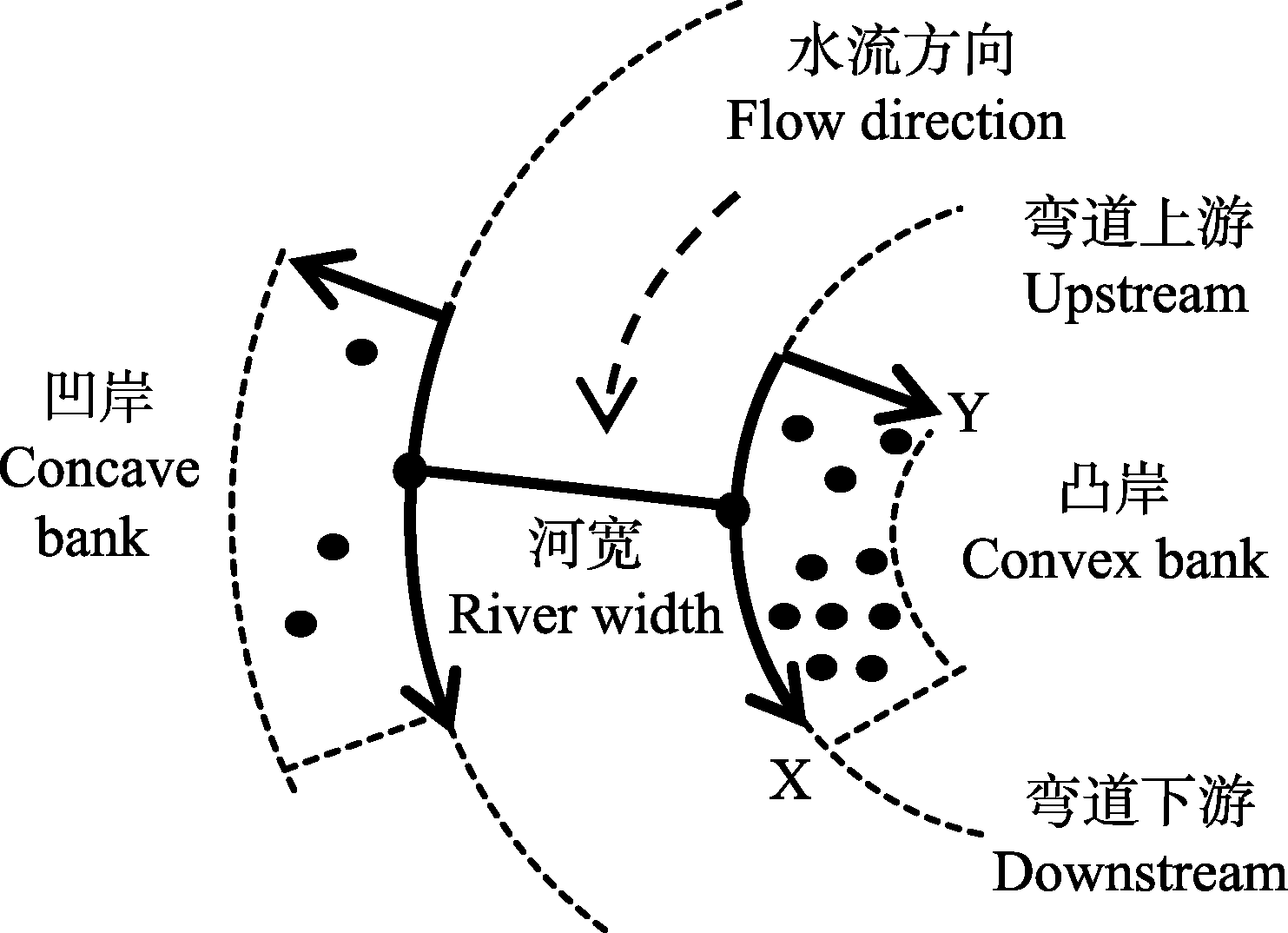
图1 溪流弯道样带设置方法。
Fig. 1 Transect setting method on stream bends.
| 研究地点 Study site | 样带 Transect | 海拔 Altitude (m) | 样带长度 Transect length (m) | 弯曲系数 Transect sinuosity | 弯顶 Top of bend | 河宽 Stream width (m) | 郁闭度 Canopy density (%) |
|---|---|---|---|---|---|---|---|
| 黎母山 Mt. Limu | L1 | 577 | 50 | 1.04 | 20 | 6.5 | 90 |
| L2 | 573 | 16 | 1.07 | 8 | 10.5 | 85 | |
| 吊罗山 Mt. Diaoluo | D1 | 258 | 24 | 1.09 | 12 | 6.5 | 78 |
| D2 | 595 | 10 | 1.20 | 5 | 3.8 | 85 | |
| 尖峰岭 Mt. Jianfeng | J1 | 690 | 20 | 1.43 | 10 | 8 | 75 |
| J2 | 665 | 40 | 1.21 | 15 | 23 | 75 |
表1 河岸带弯道研究样带概况
Table 1 Characteristics of experimental transects along the riparian bends
| 研究地点 Study site | 样带 Transect | 海拔 Altitude (m) | 样带长度 Transect length (m) | 弯曲系数 Transect sinuosity | 弯顶 Top of bend | 河宽 Stream width (m) | 郁闭度 Canopy density (%) |
|---|---|---|---|---|---|---|---|
| 黎母山 Mt. Limu | L1 | 577 | 50 | 1.04 | 20 | 6.5 | 90 |
| L2 | 573 | 16 | 1.07 | 8 | 10.5 | 85 | |
| 吊罗山 Mt. Diaoluo | D1 | 258 | 24 | 1.09 | 12 | 6.5 | 78 |
| D2 | 595 | 10 | 1.20 | 5 | 3.8 | 85 | |
| 尖峰岭 Mt. Jianfeng | J1 | 690 | 20 | 1.43 | 10 | 8 | 75 |
| J2 | 665 | 40 | 1.21 | 15 | 23 | 75 |
| 样带 Transect | 尺度 Scale (m) | 岸型 Bank type | 样本量 Sample size | 扩散系数 C | t检验 t test | Cassie指数 Ca | 平均拥挤度 m* | 丛生指数 I | 聚块性指数 m*/m | 负二项指数 K | 分布型 Pattern |
|---|---|---|---|---|---|---|---|---|---|---|---|
| L1 | 1 × 1 | 凹岸 Concave | 31 | 2.362 | 5.275** | 21.969 | 1.424 | 1.362 | 22.969 | 0.046 | C |
| 凸岸 Convex | 261 | 4.464 | 39.495** | 6.636 | 3.986 | 3.464 | 7.636 | 0.151 | C | ||
| 2 × 2 | 凹岸 Concave | 31 | 0.694 | -1.187 | -4.943 | -0.244 | -0.306 | -3.943 | -0.202 | R | |
| 凸岸 Convex | 261 | 1.549 | 6.258** | 1.051 | 1.071 | 0.549 | 2.051 | 0.951 | C | ||
| 5 × 5 | 凹岸 Concave | 31 | 0.338 | -2.563* | -10.675 | -0.600 | -0.662 | -9.675 | -0.094 | R | |
| 凸岸 Convex | 261 | 0.394 | -6.912** | -1.161 | -0.084 | -0.606 | -0.161 | -0.861 | R | ||
| L2 | 1 × 1 | 凹岸 Concave | 57 | 3.735 | 14.473** | 6.046 | 3.188 | 2.735 | 7.046 | 0.165 | C |
| 凸岸 Convex | 89 | 3.019 | 13.391** | 2.541 | 2.813 | 2.019 | 3.541 | 0.394 | C | ||
| 2 × 2 | 凹岸 Concave | 57 | 1.645 | 3.415** | 1.812 | 1.002 | 0.645 | 2.812 | 0.552 | C | |
| 凸岸 Convex | 89 | 1.982 | 6.512** | 1.412 | 1.677 | 0.982 | 2.412 | 0.708 | C | ||
| 5 × 5 | 凹岸 Concave | 57 | 0.803 | -1.044 | -0.692 | 0.088 | -0.197 | 0.308 | -1.445 | R | |
| 凸岸 Convex | 89 | 0.988 | -0.082 | -0.028 | 0.433 | -0.012 | 0.972 | -36.098 | R | ||
| D1 | 1 × 1 | 凸岸 Convex | 39 | 2.697 | 7.395** | 6.264 | 1.967 | 1.697 | 7.264 | 0.160 | C |
| 2 × 2 | 凸岸 Convex | 39 | 0.704 | -1.291 | -1.093 | -0.025 | -0.296 | -0.093 | -0.915 | R | |
| 5 × 5 | 凸岸 Convex | 39 | 0.307 | -3.021** | -5.332 | -0.563 | -0.693 | -4.332 | -0.188 | R | |
| D2 | 1 × 1 | 凹岸 Concave | 6 | 1.238 | 0.376 | 1.984 | 0.358 | 0.238 | 2.984 | 0.504 | C |
| 凸岸 Convex | 46 | 3.320 | 11.006** | 2.522 | 3.240 | 2.320 | 3.522 | 0.396 | C | ||
| 2 × 2 | 凹岸 Concave | 6 | 0.265 | -1.163 | -8.824 | -0.652 | -0.735 | -7.824 | -0.113 | R | |
| 凸岸 Convex | 46 | 1.050 | 0.237 | 0.078 | 0.689 | 0.050 | 1.078 | 12.811 | C | ||
| 5 × 5 | 凹岸 Concave | 6 | 0.027 | -1.539 | -8.111 | -0.853 | -0.973 | -7.111 | -0.123 | R | |
| 凸岸 Convex | 46 | 0.170 | -3.935** | -0.902 | 0.090 | -0.830 | 0.098 | -1.109 | R | ||
| J1 | 1 × 1 | 凸岸 Convex | 72 | 3.880 | 17.158** | 6.400 | 3.330 | 2.880 | 7.400 | 0.156 | C |
| 2 × 2 | 凸岸 Convex | 72 | 1.768 | 4.575** | 1.706 | 1.218 | 0.768 | 2.706 | 0.586 | C | |
| 5 × 5 | 凸岸 Convex | 72 | 0.678 | -1.918 | -0.894 | 0.038 | -0.322 | 0.106 | -1.118 | R | |
| J2 | 1 × 1 | 凸岸 Convex | 50 | 2.482 | 7.338** | 5.930 | 1.732 | 1.482 | 6.930 | 0.169 | C |
| 2 × 2 | 凸岸 Convex | 50 | 0.936 | -0.315 | -0.320 | 0.135 | -0.064 | 0.680 | -3.122 | R | |
| 5 × 5 | 凸岸 Convex | 50 | 0.173 | -4.092** | -3.307 | -0.577 | -0.827 | -2.307 | -0.302 | R |
表2 不同研究尺度下的弯道两侧海南杜鹃种群的空间分布格局
Table 2 Distribution patterns of Rhododendron hainanense populations on both sides of stream bends at different sampling scales
| 样带 Transect | 尺度 Scale (m) | 岸型 Bank type | 样本量 Sample size | 扩散系数 C | t检验 t test | Cassie指数 Ca | 平均拥挤度 m* | 丛生指数 I | 聚块性指数 m*/m | 负二项指数 K | 分布型 Pattern |
|---|---|---|---|---|---|---|---|---|---|---|---|
| L1 | 1 × 1 | 凹岸 Concave | 31 | 2.362 | 5.275** | 21.969 | 1.424 | 1.362 | 22.969 | 0.046 | C |
| 凸岸 Convex | 261 | 4.464 | 39.495** | 6.636 | 3.986 | 3.464 | 7.636 | 0.151 | C | ||
| 2 × 2 | 凹岸 Concave | 31 | 0.694 | -1.187 | -4.943 | -0.244 | -0.306 | -3.943 | -0.202 | R | |
| 凸岸 Convex | 261 | 1.549 | 6.258** | 1.051 | 1.071 | 0.549 | 2.051 | 0.951 | C | ||
| 5 × 5 | 凹岸 Concave | 31 | 0.338 | -2.563* | -10.675 | -0.600 | -0.662 | -9.675 | -0.094 | R | |
| 凸岸 Convex | 261 | 0.394 | -6.912** | -1.161 | -0.084 | -0.606 | -0.161 | -0.861 | R | ||
| L2 | 1 × 1 | 凹岸 Concave | 57 | 3.735 | 14.473** | 6.046 | 3.188 | 2.735 | 7.046 | 0.165 | C |
| 凸岸 Convex | 89 | 3.019 | 13.391** | 2.541 | 2.813 | 2.019 | 3.541 | 0.394 | C | ||
| 2 × 2 | 凹岸 Concave | 57 | 1.645 | 3.415** | 1.812 | 1.002 | 0.645 | 2.812 | 0.552 | C | |
| 凸岸 Convex | 89 | 1.982 | 6.512** | 1.412 | 1.677 | 0.982 | 2.412 | 0.708 | C | ||
| 5 × 5 | 凹岸 Concave | 57 | 0.803 | -1.044 | -0.692 | 0.088 | -0.197 | 0.308 | -1.445 | R | |
| 凸岸 Convex | 89 | 0.988 | -0.082 | -0.028 | 0.433 | -0.012 | 0.972 | -36.098 | R | ||
| D1 | 1 × 1 | 凸岸 Convex | 39 | 2.697 | 7.395** | 6.264 | 1.967 | 1.697 | 7.264 | 0.160 | C |
| 2 × 2 | 凸岸 Convex | 39 | 0.704 | -1.291 | -1.093 | -0.025 | -0.296 | -0.093 | -0.915 | R | |
| 5 × 5 | 凸岸 Convex | 39 | 0.307 | -3.021** | -5.332 | -0.563 | -0.693 | -4.332 | -0.188 | R | |
| D2 | 1 × 1 | 凹岸 Concave | 6 | 1.238 | 0.376 | 1.984 | 0.358 | 0.238 | 2.984 | 0.504 | C |
| 凸岸 Convex | 46 | 3.320 | 11.006** | 2.522 | 3.240 | 2.320 | 3.522 | 0.396 | C | ||
| 2 × 2 | 凹岸 Concave | 6 | 0.265 | -1.163 | -8.824 | -0.652 | -0.735 | -7.824 | -0.113 | R | |
| 凸岸 Convex | 46 | 1.050 | 0.237 | 0.078 | 0.689 | 0.050 | 1.078 | 12.811 | C | ||
| 5 × 5 | 凹岸 Concave | 6 | 0.027 | -1.539 | -8.111 | -0.853 | -0.973 | -7.111 | -0.123 | R | |
| 凸岸 Convex | 46 | 0.170 | -3.935** | -0.902 | 0.090 | -0.830 | 0.098 | -1.109 | R | ||
| J1 | 1 × 1 | 凸岸 Convex | 72 | 3.880 | 17.158** | 6.400 | 3.330 | 2.880 | 7.400 | 0.156 | C |
| 2 × 2 | 凸岸 Convex | 72 | 1.768 | 4.575** | 1.706 | 1.218 | 0.768 | 2.706 | 0.586 | C | |
| 5 × 5 | 凸岸 Convex | 72 | 0.678 | -1.918 | -0.894 | 0.038 | -0.322 | 0.106 | -1.118 | R | |
| J2 | 1 × 1 | 凸岸 Convex | 50 | 2.482 | 7.338** | 5.930 | 1.732 | 1.482 | 6.930 | 0.169 | C |
| 2 × 2 | 凸岸 Convex | 50 | 0.936 | -0.315 | -0.320 | 0.135 | -0.064 | 0.680 | -3.122 | R | |
| 5 × 5 | 凸岸 Convex | 50 | 0.173 | -4.092** | -3.307 | -0.577 | -0.827 | -2.307 | -0.302 | R |
| 样带 Transect | 岸型 Bank type | 上游/下游 Upstream/downstream | 扩散系数 C | t检验 t test | Cassie指数 Ca | 平均拥挤度 m* | 丛生指数 I | 聚块性指数 m*/m | 负二项指数 K | 分布型 Pattern |
|---|---|---|---|---|---|---|---|---|---|---|
| L1 | 凹岸 Concave | 上游 Upstream | 0.410 | -0.590 | -39.342 | -0.575 | -0.590 | -38.342 | -0.025 | R |
| 下游 Downstream | 0.702 | -1.096 | -3.196 | -0.205 | -0.298 | -2.196 | -0.313 | R | ||
| 凸岸 Convex | 上游 Upstream | 1.346 | 2.656** | 0.581 | 0.941 | 0.346 | 1.581 | 1.721 | C | |
| 下游 Downstream | 1.729 | 6.124** | 1.541 | 1.203 | 0.729 | 2.541 | 0.649 | C | ||
| L2 | 凹岸 Concave | 上游 Upstream | 0.447 | -0.874 | -7.368 | -0.478 | -0.553 | -6.368 | -0.136 | R |
| 下游 Downstream | 1.574 | 2.868** | 0.900 | 1.211 | 0.574 | 1.900 | 1.112 | C | ||
| 凸岸 Convex | 上游 Upstream | 1.154 | 0.409 | 0.659 | 0.389 | 0.154 | 1.659 | 1.518 | C | |
| 下游 Downstream | 1.837 | 5.056** | 0.724 | 1.993 | 0.837 | 1.724 | 1.382 | C | ||
| D1 | 凸岸 Convex | 上游 Upstream | 0.493 | -1.389 | -2.283 | -0.285 | -0.507 | -1.283 | -0.438 | R |
| 下游 Downstream | 0.759 | -0.780 | -0.788 | 0.065 | -0.241 | 0.212 | -1.270 | R | ||
| D2 | 凹岸 Concave | 上游 Upstream | 0.297 | -0.861 | -6.328 | -0.592 | -0.703 | -5.328 | -0.158 | R |
| 下游 Downstream | 0.219 | -0.552 | -14.063 | -0.726 | -0.781 | -13.063 | -0.071 | R | ||
| 凸岸 Convex | 上游 Upstream | 0.730 | -0.738 | -0.606 | 0.175 | -0.270 | 0.394 | -1.649 | R | |
| 下游 Downstream | 1.219 | 0.833 | 0.263 | 1.052 | 0.219 | 1.263 | 3.810 | C | ||
| J1 | 凸岸 Convex | 上游 Upstream | 0.861 | -0.340 | -0.853 | 0.024 | -0.139 | 0.147 | -1.172 | R |
| 下游 Downstream | 1.572 | 3.028** | 0.803 | 1.285 | 0.572 | 1.803 | 1.245 | C | ||
| J2 | 凸岸 Convex | 上游 Upstream | 0.483 | -0.896 | -7.091 | -0.444 | -0.517 | -6.091 | -0.141 | R |
| 下游 Downstream | 0.964 | -0.164 | -0.130 | 0.240 | -0.036 | 0.870 | -7.699 | R |
表3 弯道上下游海南杜鹃空间分布格局(尺度2 m × 2 m)
Table 3 Distribution patterns of Rhododendron hainanense on upstream and downstream of stream bends evaluated as the scale of 2 m × 2 m
| 样带 Transect | 岸型 Bank type | 上游/下游 Upstream/downstream | 扩散系数 C | t检验 t test | Cassie指数 Ca | 平均拥挤度 m* | 丛生指数 I | 聚块性指数 m*/m | 负二项指数 K | 分布型 Pattern |
|---|---|---|---|---|---|---|---|---|---|---|
| L1 | 凹岸 Concave | 上游 Upstream | 0.410 | -0.590 | -39.342 | -0.575 | -0.590 | -38.342 | -0.025 | R |
| 下游 Downstream | 0.702 | -1.096 | -3.196 | -0.205 | -0.298 | -2.196 | -0.313 | R | ||
| 凸岸 Convex | 上游 Upstream | 1.346 | 2.656** | 0.581 | 0.941 | 0.346 | 1.581 | 1.721 | C | |
| 下游 Downstream | 1.729 | 6.124** | 1.541 | 1.203 | 0.729 | 2.541 | 0.649 | C | ||
| L2 | 凹岸 Concave | 上游 Upstream | 0.447 | -0.874 | -7.368 | -0.478 | -0.553 | -6.368 | -0.136 | R |
| 下游 Downstream | 1.574 | 2.868** | 0.900 | 1.211 | 0.574 | 1.900 | 1.112 | C | ||
| 凸岸 Convex | 上游 Upstream | 1.154 | 0.409 | 0.659 | 0.389 | 0.154 | 1.659 | 1.518 | C | |
| 下游 Downstream | 1.837 | 5.056** | 0.724 | 1.993 | 0.837 | 1.724 | 1.382 | C | ||
| D1 | 凸岸 Convex | 上游 Upstream | 0.493 | -1.389 | -2.283 | -0.285 | -0.507 | -1.283 | -0.438 | R |
| 下游 Downstream | 0.759 | -0.780 | -0.788 | 0.065 | -0.241 | 0.212 | -1.270 | R | ||
| D2 | 凹岸 Concave | 上游 Upstream | 0.297 | -0.861 | -6.328 | -0.592 | -0.703 | -5.328 | -0.158 | R |
| 下游 Downstream | 0.219 | -0.552 | -14.063 | -0.726 | -0.781 | -13.063 | -0.071 | R | ||
| 凸岸 Convex | 上游 Upstream | 0.730 | -0.738 | -0.606 | 0.175 | -0.270 | 0.394 | -1.649 | R | |
| 下游 Downstream | 1.219 | 0.833 | 0.263 | 1.052 | 0.219 | 1.263 | 3.810 | C | ||
| J1 | 凸岸 Convex | 上游 Upstream | 0.861 | -0.340 | -0.853 | 0.024 | -0.139 | 0.147 | -1.172 | R |
| 下游 Downstream | 1.572 | 3.028** | 0.803 | 1.285 | 0.572 | 1.803 | 1.245 | C | ||
| J2 | 凸岸 Convex | 上游 Upstream | 0.483 | -0.896 | -7.091 | -0.444 | -0.517 | -6.091 | -0.141 | R |
| 下游 Downstream | 0.964 | -0.164 | -0.130 | 0.240 | -0.036 | 0.870 | -7.699 | R |
| 样带 Transect | 岸型 Bank type | 龄级 Age class | 扩散系数 C | t检验 t test | Cassie指数 Ca | 平均拥挤度 m* | 丛生指数 I | 聚块性指数 m*/m | 负二项指数 K | 分布型 Pattern |
|---|---|---|---|---|---|---|---|---|---|---|
| L1 | 凸岸 Convex | Ⅰ | 1.260 | 2.142* | 0.948 | 0.534 | 0.260 | 1.948 | 1.055 | C |
| Ⅱ | 0.705 | -1.723 | -2.141 | -0.157 | -0.295 | -1.141 | -0.467 | R | ||
| Ⅲ | 0.480 | -2.112* | -7.648 | -0.452 | -0.520 | -6.648 | -0.131 | R | ||
| Ⅳ | 0.498 | -1.588 | -11.960 | -0.460 | -0.502 | -10.960 | -0.084 | R | ||
| L2 | 凸岸 Convex | Ⅰ | 0.670 | -1.379 | -1.172 | -0.048 | -0.330 | -0.172 | -0.853 | R |
| Ⅱ | 0.928 | -0.274 | -0.307 | 0.162 | -0.072 | 0.693 | -3.253 | R | ||
| Ⅲ | 0.590 | -1.004 | -4.037 | -0.308 | -0.410 | -3.037 | -0.248 | R | ||
| Ⅳ | 0.451 | -1.228 | -6.390 | -0.463 | -0.549 | -5.390 | -0.157 | R | ||
| D1 | 凸岸 Convex | Ⅰ | 0.616 | -1.486 | -1.782 | -0.168 | -0.384 | -0.782 | -0.561 | R |
| Ⅱ | 0.386 | -0.971 | -14.743 | -0.573 | -0.614 | -13.743 | -0.068 | R | ||
| Ⅲ | 0.243 | -0.535 | -54.514 | -0.743 | -0.757 | -53.514 | -0.018 | R | ||
| Ⅳ | - | - | - | - | - | - | - | - | ||
| D2 | 凸岸 Convex | Ⅰ | 1.156 | 0.623 | 0.283 | 0.706 | 0.156 | 1.283 | 3.529 | C |
| Ⅱ | 0.259 | -1.386 | -5.558 | -0.608 | -0.741 | -4.558 | -0.180 | R | ||
| Ⅲ | 0.330 | -0.820 | -10.045 | -0.603 | -0.670 | -9.045 | -0.100 | R | ||
| Ⅳ | 0.250 | - | -45.000 | -0.733 | -0.750 | -44.000 | -0.022 | R | ||
| J1 | 凸岸 Convex | Ⅰ | 1.643 | 3.582** | 1.634 | 1.037 | 0.643 | 2.634 | 0.612 | C |
| Ⅱ | 0.370 | -1.261 | -11.206 | -0.574 | -0.630 | -10.206 | -0.089 | R | ||
| Ⅲ | - | - | - | - | - | - | - | - | ||
| Ⅳ | - | - | - | - | - | - | - | - | ||
| J2 | 凸岸 Convex | Ⅰ | 0.839 | -0.646 | -1.174 | -0.024 | -0.161 | -0.174 | -0.852 | R |
| Ⅱ | 0.733 | -0.706 | -4.271 | -0.204 | -0.267 | -3.271 | -0.234 | R | ||
| Ⅲ | 0.250 | - | -180.000 | -0.746 | -0.750 | -179.000 | -0.006 | R | ||
| Ⅳ | 0.250 | - | -180.000 | -0.746 | -0.750 | -179.000 | -0.006 | R |
表4 不同龄级海南杜鹃空间分布格局(尺度2 m × 2 m)
Table 4 Distribution patterns of different age classes of Rhododendron hainanense evaluated at the scale of 2 m × 2 m
| 样带 Transect | 岸型 Bank type | 龄级 Age class | 扩散系数 C | t检验 t test | Cassie指数 Ca | 平均拥挤度 m* | 丛生指数 I | 聚块性指数 m*/m | 负二项指数 K | 分布型 Pattern |
|---|---|---|---|---|---|---|---|---|---|---|
| L1 | 凸岸 Convex | Ⅰ | 1.260 | 2.142* | 0.948 | 0.534 | 0.260 | 1.948 | 1.055 | C |
| Ⅱ | 0.705 | -1.723 | -2.141 | -0.157 | -0.295 | -1.141 | -0.467 | R | ||
| Ⅲ | 0.480 | -2.112* | -7.648 | -0.452 | -0.520 | -6.648 | -0.131 | R | ||
| Ⅳ | 0.498 | -1.588 | -11.960 | -0.460 | -0.502 | -10.960 | -0.084 | R | ||
| L2 | 凸岸 Convex | Ⅰ | 0.670 | -1.379 | -1.172 | -0.048 | -0.330 | -0.172 | -0.853 | R |
| Ⅱ | 0.928 | -0.274 | -0.307 | 0.162 | -0.072 | 0.693 | -3.253 | R | ||
| Ⅲ | 0.590 | -1.004 | -4.037 | -0.308 | -0.410 | -3.037 | -0.248 | R | ||
| Ⅳ | 0.451 | -1.228 | -6.390 | -0.463 | -0.549 | -5.390 | -0.157 | R | ||
| D1 | 凸岸 Convex | Ⅰ | 0.616 | -1.486 | -1.782 | -0.168 | -0.384 | -0.782 | -0.561 | R |
| Ⅱ | 0.386 | -0.971 | -14.743 | -0.573 | -0.614 | -13.743 | -0.068 | R | ||
| Ⅲ | 0.243 | -0.535 | -54.514 | -0.743 | -0.757 | -53.514 | -0.018 | R | ||
| Ⅳ | - | - | - | - | - | - | - | - | ||
| D2 | 凸岸 Convex | Ⅰ | 1.156 | 0.623 | 0.283 | 0.706 | 0.156 | 1.283 | 3.529 | C |
| Ⅱ | 0.259 | -1.386 | -5.558 | -0.608 | -0.741 | -4.558 | -0.180 | R | ||
| Ⅲ | 0.330 | -0.820 | -10.045 | -0.603 | -0.670 | -9.045 | -0.100 | R | ||
| Ⅳ | 0.250 | - | -45.000 | -0.733 | -0.750 | -44.000 | -0.022 | R | ||
| J1 | 凸岸 Convex | Ⅰ | 1.643 | 3.582** | 1.634 | 1.037 | 0.643 | 2.634 | 0.612 | C |
| Ⅱ | 0.370 | -1.261 | -11.206 | -0.574 | -0.630 | -10.206 | -0.089 | R | ||
| Ⅲ | - | - | - | - | - | - | - | - | ||
| Ⅳ | - | - | - | - | - | - | - | - | ||
| J2 | 凸岸 Convex | Ⅰ | 0.839 | -0.646 | -1.174 | -0.024 | -0.161 | -0.174 | -0.852 | R |
| Ⅱ | 0.733 | -0.706 | -4.271 | -0.204 | -0.267 | -3.271 | -0.234 | R | ||
| Ⅲ | 0.250 | - | -180.000 | -0.746 | -0.750 | -179.000 | -0.006 | R | ||
| Ⅳ | 0.250 | - | -180.000 | -0.746 | -0.750 | -179.000 | -0.006 | R |
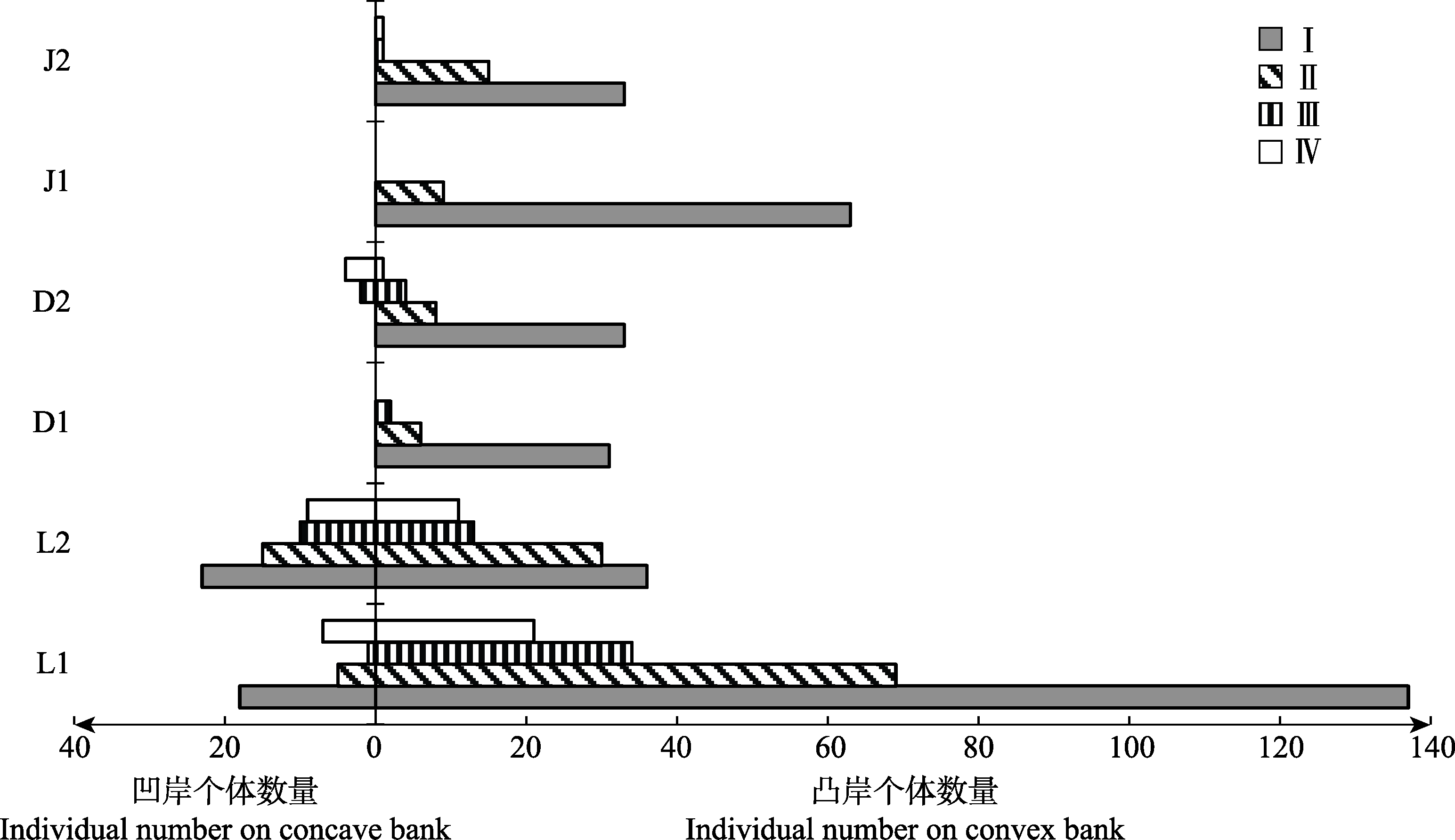
图2 溪流弯道两侧海南杜鹃种群的年龄结构。D1, 吊罗山1号样带; D2, 吊罗山2号样带; J1, 尖峰岭1号样带; J2, 尖峰岭2号样带; L1, 黎母山1号样带; L2, 黎母山2号样带。I, 幼苗; II, 幼树; III, 小树; IV, 大树。
Fig. 2 Age structures of the Rhododendron hainanense populations on both sides of stream bends. D1, No. 1 transect of Mt. Diaoluo; D2, No. 2 transect of Mt. Diaoluo; J1, No. 1 transect of Mt. Jianfeng; J2, No. 2 transect of Mt. Jianfeng; L1, No. 1 transect of Mt. Limu; L2, No. 2 transect of Mt. Limu. I, seedling; II, sapling; III, undershrub; IV, large shrub.
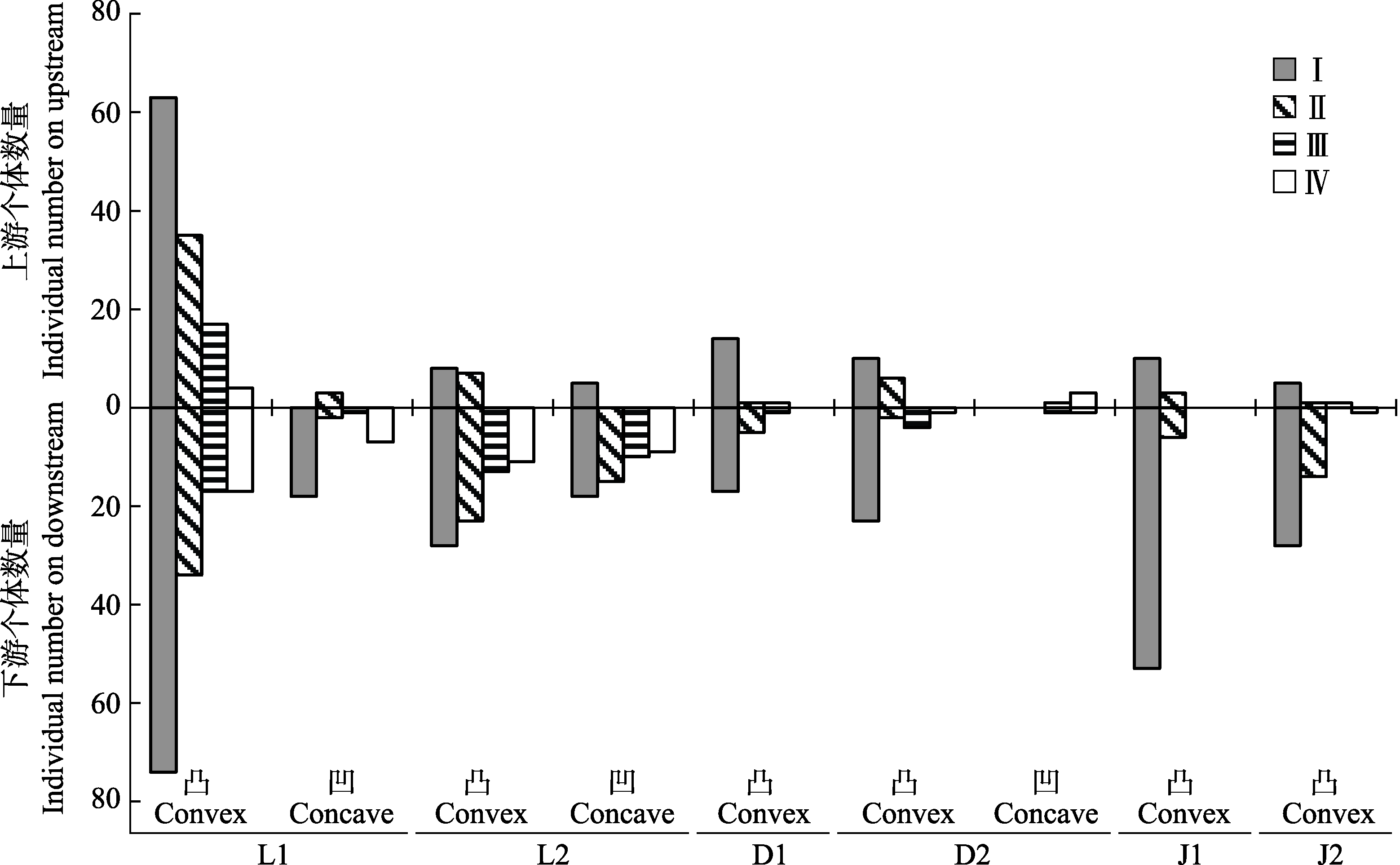
图3 溪流弯道上下游海南杜鹃种群的年龄结构。D1, 吊罗山1号样带; D2, 吊罗山2号样带; J1, 尖峰岭1号样带; J2, 尖峰岭2号样带; L1, 黎母山1号样带; L2, 黎母山2号样带。I, 幼苗; II, 幼树; III, 小树; IV, 大树。
Fig. 3 Age structures of the Rhododendron hainanense populations on upstream and downstream of stream bends. D1, No. 1 transect of Mt. Diaoluo; D2, No. 2 transect of Mt. Diaoluo; J1, No. 1 transect of Mt. Jianfeng; J2, No. 2 transect of Mt. Jianfeng; L1, No. 1 transect of Mt. Limu; L2, No. 2 transect of Mt. Limu. I, seedling; II, sapling; III, undershrub; IV, large shrub.
| [1] |
Balke T, Webb EL, van den Elzen E, Galli D, Herman PMJ, Bouma TJ ( 2013). Seedling establishment in a dynamic sedimentary environment: A conceptual framework using mangroves. Journal of Applied Ecology, 50, 740-747.
DOI URL PMID |
| [2] |
Bendix J, Hupp CR ( 2000). Hydrological and geomorphic impacts on riparian plant communities. Hydrological Processes, 14, 2977-2990.
DOI URL |
| [3] |
Bertoldi W, Drake NA, Gurnell AM ( 2011). Interactions between river flows and colonizing vegetation on a braided river: Exploring spatial and temporal dynamics in riparian vegetation cover using satellite data. Earth Surface Processes and Landforms, 36, 1474-1486.
DOI URL |
| [4] |
Bhuiyan F, Hey RD, Wormleaton PR ( 2010). Bank-attached vanes for bank erosion control and restoration of river meanders. Journal of Hydraulic Engineering, 136, 583-596.
DOI URL |
| [5] |
Burkart M ( 2001). River corridor plants (Stromtalpflanzen) in Central European lowland: A review of a poorly understood plant distribution pattern. Global Ecology and Biogeography, 10, 449-468.
DOI URL |
| [6] |
Burt T, Pinay G, Sabater S ( 2010). What do we still need to know about the ecohydrology of riparian zones? Ecohydrology, 3, 373-377.
DOI URL |
| [7] |
Camporeale C, Perucca E, Ridolfi L, Gurnell AM ( 2013). Modeling the interaction between river morphodynamics and riparian vegetation. Reviews of Geophysics, 51, 379-414.
DOI URL |
| [8] |
Catford JA, Jansson R ( 2014). Drowned, buried and carried away: Effects of plant traits on the distribution of native and alien species in riparian ecosystems. New Phytologist, 204, 19-36.
DOI URL PMID |
| [9] |
Corenblit D, Tabacchi E, Steiger J, Gurnell AM ( 2007). Reciprocal interactions and adjustments between fluvial landforms and vegetation dynamics in river corridors: A review of complementary approaches. Earth-Science Reviews, 84, 56-86.
DOI URL |
| [10] | Crosato A ( 2008). Analysis and Modelling of River Meandering. IOS Press, Amsterdam. |
| [11] |
Curran JC, Hession WC ( 2013). Vegetative impacts on hydraulics and sediment processes across the fluvial system. Journal of Hydrology, 505, 364-376.
DOI URL |
| [12] |
Da LJ, Yang YC, Song YC ( 2004). Population structure and regeneration types of dominant species in an evergreen broadleaved forest in Tiantong National Forest Park, Zhejiang Province, eastern China. Acta Phytoecologica Sinica, 28, 376-384.
DOI URL |
|
[ 达良俊, 杨永川, 宋永昌 ( 2004). 浙江天童国家森林公园常绿阔叶林主要组成种的种群结构及更新类型. 植物生态学报, 28, 376-384.]
DOI URL |
|
| [13] |
Drezner TD, Fall PL, Stromberg JC ( 2001). Plant distribution and dispersal mechanisms at the Hassayampa River Preserve, Arizona, USA. Global Ecology and Biogeography, 10, 205-217.
DOI URL |
| [14] | Fang MY, Fang RZ, He MY, Hu LZ, Yang HB, Chamberlain DF ( 2005). Flora of China. Vol. 14. Science Press and Missouri Botanical Garden Press, Beijing. 438. |
| [15] |
Garssen AG, Baattrup-Pedersen A, Voesenek LACJ, Verhoeven JTA, Soons MB ( 2015). Riparian plant community responses to increased flooding: A meta-analysis. Global Change Biology, 21, 2881-2890.
DOI URL PMID |
| [16] |
Greet J, Webb JA, Cousens RD ( 2011). The importance of seasonal flow timing for riparian vegetation dynamics: A systematic review using causal criteria analysis. Freshwater Biology, 56, 1231-1247.
DOI URL |
| [17] |
Gurnell AM, Bertoldi W, Corenblit D ( 2012). Changing river channels: The roles of hydrological processes, plants and pioneer fluvial landforms in humid temperate, mixed load, gravel bed rivers. Earth-Science Reviews, 111, 129-141.
DOI URL |
| [18] | Han L, Wang HZ, Yu J ( 2013). Research progress and prospects on riparian zone ecology. Chinese Journal of Ecology, 22, 879-886. |
| [ 韩路, 王海珍, 于军 ( 2013). 河岸带生态学研究进展与展望. 生态环境学报, 22, 879-886.] | |
| [19] |
He D, Wei XZ, Li LF, Jiang MX, Yang JY, Yu J ( 2009). Population structure and dynamics of Cercidiphyllum japonicum in riparian zones of the Shennongjia Mountainous Region, central China. Chinese Journal of Plant Ecology, 33, 469-481.
DOI URL |
|
[ 何东, 魏新增, 李连发, 江明喜, 杨敬元, 喻杰 ( 2009). 神农架山地河岸带连香树的种群结构与动态. 植物生态学报, 33, 469-481.]
DOI URL |
|
| [20] |
He HB, Zhang HJ, Jia GX ( 2006). Population structure and spatial distribution pattern of Ammopiptanthus mongolicusin in Dengkou County, Inner Mongolia Autonomous Region. Scientia Silvae Sinicae, 42(10), 13-18.
DOI URL |
|
[ 何恒斌, 张惠娟, 贾桂霞 ( 2006). 磴口县沙冬青种群结构和空间分布格局的研究. 林业科学, 42(10), 13-18.]
DOI URL |
|
| [21] |
Horvath TG ( 2004). Retention of particulate matter by macrophytes in a first-order stream. Aquatic Botany, 78, 27-36.
DOI URL |
| [22] |
Huang XB, Li SF, Su JR, Su L ( 2015). Influences of selective cutting on the structure and spatial distribution patterns of a natural population of Pinus kesiya var. langbianensis. Acta Ecologica Sinica, 35, 8241-8250.
DOI URL |
|
[ 黄小波, 李帅锋, 苏建荣, 苏磊 ( 2015). 择伐对思茅松自然种群结构和空间分布格局的影响. 生态学报, 35, 8241-8250.]
DOI URL |
|
| [23] |
Kang HJ, Liu P, Chen ZL, Liao CC, Li CH, Chen WX, Lei ZP ( 2007). Size-class structure and distribution pattern of Emmenopterys henryi in different habitats. Scientia Silvae Sinicae, 43(12), 22-27.
DOI URL |
|
[ 康华靖, 刘鹏, 陈子林, 廖承川, 李成惠, 陈卫新, 雷祖培 ( 2007). 不同生境香果树种群的径级结构与分布格局. 林业科学, 43(12), 22-27.]
DOI URL |
|
| [24] | Li XL, Sun ZY, Li JY, Xu Y, Luo J ( 2013). Population structure and spatial distribution pattern of Camellia azalea in E'huangzhang Nature Reserve of Guangdong, China. Chinese Journal of Applied Ecology, 24, 2115-2121. |
| [ 李辛雷, 孙振元, 李纪元, 徐翊, 罗建 ( 2013). 广东鹅凰嶂自然保护区杜鹃红山茶种群结构与空间分布格局. 应用生态学报, 24, 2115-2121.] | |
| [25] |
Liu YQ ( 2003). Development of research on essential characteristics of meandering rivers. Pearl River, ( 2), 1-4.
DOI URL |
|
[ 刘月琴 ( 2003). 弯曲型河流基本特性研究进展. 人民珠江, ( 2), 1-4.]
DOI URL |
|
| [26] |
Merritt DM, Nilsson C, Jansson R ( 2010 a). Consequences of propagule dispersal and river fragmentation for riparian plant community diversity and turnover. Ecological Monographs, 80, 609-626.
DOI URL |
| [27] |
Merritt DM, Scott ML, LeRoy Poff N, Auble GT, Lytle DA ( 2010 b). Theory, methods and tools for determining environmental flows for riparian vegetation: Riparian vegetation- flow response guilds. Freshwater Biology, 55, 206-225.
DOI URL |
| [28] |
Millar RG ( 2000). Influence of bank vegetation on alluvial channel patterns. Water Resources Research, 36, 1109-1118.
DOI URL |
| [29] |
Naiman RJ, Decamps H ( 1997). The ecology of interfaces: Riparian zones. Annual Review of Ecology and Systematics, 28, 621-658.
DOI URL |
| [30] |
Piégay H, Darby SE, Mosselman E, Surian N ( 2005). A review of techniques available for delimiting the erodible river corridor: A sustainable approach to managing bank erosion. River Research and Applications, 21, 773-789.
DOI URL |
| [31] |
Richardson DM, Holmes PM, Esler KJ, Galatowitsch SM, Stromberg JC, Kirkman SP, Pysek P, Hobbs RJ ( 2007). Riparian vegetation: Degradation, alien plant invasions, and restoration prospects. Diversity and Distributions, 13, 126-139.
DOI URL |
| [32] |
Shi YH, Li SP, Liang WH, Song XQ, Tan JH ( 2010). Germplasm resourses of Rhododendron in Hainan. Chinese Journal of Tropical Crops, 31, 551-555.
DOI URL |
|
[ 史佑海, 李绍鹏, 梁伟红, 宋希强, 谭金红 ( 2010). 海南野生杜鹃花属植物种质资源调查研究. 热带作物学报, 31, 551-555.]
DOI URL |
|
| [33] |
Solari L, van Oorschot M, Belletti B, Hendriks D, Rinaldi M, Vargas-Luna A ( 2016). Advances on modelling riparian vegetation—Hydromorphology interactions. River Research and Applications, 32, 164-178.
DOI URL |
| [34] |
Tal M, Paola C ( 2010). Effects of vegetation on channel morphodynamics: Results and insights from laboratory experiments. Earth Surface Processes and Landforms, 35, 1014-1028.
DOI URL |
| [35] | Tu Y ( 2017). Experimental study of effects of emergent rigid vegetation on flow and sediment movement in the curved flume. PhD dissertation, China Institute of Water Resources & Hydropower Research, Beijing. (in Chinese). |
| [ 凃洋 ( 2017). 挺水植被对弯道水流和泥沙运动影响试验研究. 博士学位论文, 中国水利水电科学研究院, 北京.] | |
| [36] |
Wei XZ, Huang HD, Jiang MX, Yang JY ( 2008). Quantitative characteristics and spatial distribution patterns of Euptelea pleiospermum populations in riparian zones of the Shennongjia area, central China. Journal of Plant Ecology (Chinese Version), 32, 825-837.
DOI URL |
|
[ 魏新增, 黄汉东, 江明喜, 杨敬元 ( 2008). 神农架地区河岸带中领春木种群数量特征与空间分布格局. 植物生态学报, 32, 825-837.]
DOI URL |
|
| [37] | Zhang PJ, Qing H, Zhang L, Xu YD, Mu L, Ye RH, Qiu X, Chang H, Shen HH, Yang J ( 2017). Population structure and spatial pattern of Caragana tibetica communities in Nei Mongol shrub-encroached grassland. Chinese Journal of Plant Ecology, 41, 165-174. |
| [ 张璞进, 清华, 张雷, 徐延达, 木兰, 晔薷罕, 邱晓, 常虹, 沈海花, 杨劼 ( 2017). 内蒙古灌丛化草原毛刺锦鸡儿种群结构和空间分布格局. 植物生态学报, 41, 165-174.] | |
| [38] | Zhang WH, Wang YP, Kang YX, Liu XJ ( 2005). Spatial distribution pattern of Larix chinensis population in Taibai Mt. Chinese Journal of Applied Ecology, 16, 207-212. |
| [ 张文辉, 王延平, 康永祥, 刘祥君 ( 2005). 太白山太白红杉种群空间分布格局研究. 应用生态学报, 16, 07-212.] | |
| [39] |
Zhao ZS, Zheng YQ, Liang JY, Han ZJ, Li ZJ ( 2016). Spatial distribution pattern of Populus euphratica and P. pruinosa clonal ramets in Tarim River Basin, China. Chinese Journal of Applied Ecology, 27, 403-411.
DOI URL |
|
[ 赵正帅, 郑亚琼, 梁继业, 韩占江, 李志军 ( 2016). 塔里木河流域胡杨和灰叶胡杨克隆分株空间分布格局. 应用生态学报, 27, 403-411.]
DOI URL |
|
| [40] | Zhen JH, Liu GH ( 2008). Population structure characteristics of Tetraena mongolica in different habitats. Acta Ecologica Sinica, 28, 1829-1841. |
| [ 甄江红, 刘果厚 ( 2008). 不同生境条件下四合木(Tetraena mongolica Maxim.)种群结构特征. 生态学报, 28, 1829-1841.] |
| [1] | 王雨婷, 刘旭婧, 唐驰飞, 陈玮钰, 王美娟, 向松竹, 刘梅, 杨林森, 傅强, 晏召贵, 孟红杰. 神农架极小种群植物庙台槭群落特征及种群动态[J]. 植物生态学报, 2024, 48(1): 80-91. |
| [2] | 李晓田, 王铁娟, 韩文娟, 张丽, 张慧, 刘晓婷, 刘雅洁. 东阿拉善珍稀濒危植物绵刺种群结构与点格局分析[J]. 植物生态学报, 2023, 47(4): 506-514. |
| [3] | 林春惠, 顾惠怡, 叶钦良, 张志坚, 钟智明, 易绮斐. 珍稀濒危植物大苞山茶种群结构与动态特征[J]. 植物生态学报, 2023, 47(12): 1684-1692. |
| [4] | 蔚亮, 李均力, 包安明, 白洁, 黄粤, 刘铁, 沈占锋. 塔里木河下游湿地面积时序变化及对生态输水的响应[J]. 植物生态学报, 2020, 44(6): 616-627. |
| [5] | 赵阳, 刘锦乾, 陈学龙, 杨萌萌, 曹家豪, 齐瑞, 曹秀文. 洮河上游紫果云杉种群结构特征[J]. 植物生态学报, 2020, 44(3): 266-276. |
| [6] | 魏雪莹, 叶育石, 林喜珀, 崔煜文, 曾飞燕, 王发国. 极小种群植物猪血木的种群现状及保护对策[J]. 植物生态学报, 2020, 44(12): 1236-1246. |
| [7] | 拓锋, 刘贤德, 刘润红, 赵维俊, 敬文茂, 马剑, 武秀荣, 赵晶忠, 马雪娥. 祁连山大野口流域青海云杉种群空间格局及其关联性[J]. 植物生态学报, 2020, 44(11): 1172-1183. |
| [8] | 梁士楚, 刘润红, 荣春艳, 常斌, 姜勇. 漓江河岸带植物功能性状变异与关联[J]. 植物生态学报, 2019, 43(1): 16-26. |
| [9] | 李亚飞, 于静洁, 陆凯, 王平, 张一驰, 杜朝阳. 额济纳三角洲胡杨和多枝柽柳水分来源解析[J]. 植物生态学报, 2017, 41(5): 519-528. |
| [10] | 张璞进, 清华, 张雷, 徐延达, 木兰, 晔薷罕, 邱晓, 常虹, 沈海花, 杨劼. 内蒙古灌丛化草原毛刺锦鸡儿种群结构和空间分布格局[J]. 植物生态学报, 2017, 41(2): 165-174. |
| [11] | 李艳朋, 许涵, 李意德, 骆土寿, 陈德祥, 周璋, 林明献, 杨怀. 海南尖峰岭热带山地雨林物种多样性空间分布格局的尺度效应[J]. 植物生态学报, 2016, 40(9): 861-870. |
| [12] | 岳楷, 杨万勤, 彭艳, 黄春萍, 张川, 吴福忠. 高寒森林溪流对凋落叶分解过程中木质素降解的影响[J]. 植物生态学报, 2016, 40(9): 893-901. |
| [13] | 吴毅, 刘文耀, 宋亮, 陈曦, 卢华正, 李苏, 石贤萌. 基于林冠塔吊的附生植物生态学研究进展[J]. 植物生态学报, 2016, 40(5): 508-522. |
| [14] | 朱绪超, 袁国富, 邵明安, 易小波, 杜涛. 塔里木河下游河岸带植被的空间结构特征[J]. 植物生态学报, 2015, 39(11): 1053-1061. |
| [15] | 马松梅, 聂迎彬, 耿庆龙, 王荣学. 气候变化对蒙古扁桃适宜分布范围和空间格局的影响[J]. 植物生态学报, 2014, 38(3): 262-269. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
Copyright © 2022 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19