
植物生态学报 ›› 2016, Vol. 40 ›› Issue (11): 1154-1163.DOI: 10.17521/cjpe.2016.0157
所属专题: 凋落物
宋彦君1,2,3, 田文斌4, 刘翔宇1,2,3, 尹芳1,2,3, 程浚洋1,2,3, 朱丹妮1,2,3, AliARSHAD1,2,3, 阎恩荣1,2,3,*( )
)
收稿日期:2016-05-05
接受日期:2016-09-21
出版日期:2016-11-10
发布日期:2016-11-25
通讯作者:
阎恩荣
基金资助:
Yan-Jun SONG1,2,3, Wen-Bin TIAN4, Xiang-Yu LIU1,2,3, Fang YIN1,2,3, Jun-Yang CHENG1,2,3, Dan-Ni ZHU1,2,3, ARSHAD Ali1,2,3, En-Rong YAN1,2,3,*()
Received:2016-05-05
Accepted:2016-09-21
Online:2016-11-10
Published:2016-11-25
Contact:
En-Rong YAN
摘要:
森林凋落物动态是森林生态系统过程中的重要组成部分, 探索森林凋落物动态特征与微气候间的关系, 对深入了解生态系统变化过程运行机理具有重要意义。该研究以浙江普陀山岛典型森林类型枫香(Liquidambar formosana)林、天竺桂(Cinnamomum japonicum)+红楠(Machilus thunbergii)林、马尾松(Pinus massoniana)林、青冈(Cyclobalanopsis glauca)林、台湾蚊母树(Distylium gracile)林为研究对象, 探究森林凋落量与微气候的关系。结果表明: 1)森林年凋落量介于3.45-5.36 t·hm-2·a-1, 年凋落量各组分比例表现出不同的组成特征, 森林类型与森林月凋落量无关。2)森林月凋落量动态模式主要呈双峰型和三峰型, 凋落峰值和风速峰值趋势相一致, 主要集中在台风干扰较大的4、7、12月份。3)冗余分析结果表明, 影响不同组分月凋落量的微气候因子不同: 月总凋落量、叶凋落量、果凋落量和碎屑凋落量的主要控制因素均为空气温度, 随空气温度的升高而增大; 枝凋落量的主要影响因素是森林上层风速, 同样起着显著的正向作用; 花凋落量与空气湿度之间呈负相关关系。森林年凋落量各组分所占比例存在差异, 森林月凋落量和森林类型无关, 主要与空气温度、空气湿度、森林上层风速等微气候因子有关。
宋彦君, 田文斌, 刘翔宇, 尹芳, 程浚洋, 朱丹妮, AliARSHAD, 阎恩荣. 浙江普陀山岛森林凋落物动态与微气候的关联性. 植物生态学报, 2016, 40(11): 1154-1163. DOI: 10.17521/cjpe.2016.0157
Yan-Jun SONG, Wen-Bin TIAN, Xiang-Yu LIU, Fang YIN, Jun-Yang CHENG, Dan-Ni ZHU, ARSHAD Ali, En-Rong YAN. Associations between litterfall dynamics and micro-climate in forests of Putuoshan Island, Zhejiang, China. Chinese Journal of Plant Ecology, 2016, 40(11): 1154-1163. DOI: 10.17521/cjpe.2016.0157
| 森林类型 Forest type | 年龄 Age (a) | 海拔 Altitude (m) | 坡度 Slope | 坡向 Aspect | 群落高度 Community height (m) | 盖度 Coverage (%) | 优势树种 Dominant tree species |
|---|---|---|---|---|---|---|---|
| 枫香林 Liquidambar formosana forest | 50 | 65 | 19° | 东南 SE | 13.5 | 70 | 枫香 Liquidambar formosana 麻栎 Quercus acutissima 樟树 Cinnamomum camphora |
| 天竺桂+红楠林 Cinnamomum japonicum+ Machilus thunbergii forest | 80 | 70 | 15° | 西 W | 9.5 | 100 | 红楠 Machilus thunbergii 天竺桂 Cinnamomum japonicum |
| 马尾松林 Pinus massoniana forest | 40 | 50 | 14° | 东北 NE | 13.5 | 80 | 马尾松 Pinus massoniana 石栎 Lithocarpus glabra 青冈 Cyclobalanopsis glauca |
| 青冈林 Cyclobalanopsis glauca forest | 60 | 153 | 15° | 东北 NE | 8.5 | 90 | 青冈 Cyclobalanopsis glauca 石栎 Lithocarpus glaber |
| 台湾蚊母树林 Distylium gracile forest | 160 | 241 | 8° | 东北 NE | 9.6 | 100 | 台湾蚊母树 Distylium gracile 铁冬青 Ilex rotunda 红楠 Machilus thunbergii 红山茶 Camellia longicaudata |
表1 浙江普陀山岛5种森林类型的样地特征
Table 1 Characteristics of the five forest types in Putuoshan Island, Zhejiang, East China
| 森林类型 Forest type | 年龄 Age (a) | 海拔 Altitude (m) | 坡度 Slope | 坡向 Aspect | 群落高度 Community height (m) | 盖度 Coverage (%) | 优势树种 Dominant tree species |
|---|---|---|---|---|---|---|---|
| 枫香林 Liquidambar formosana forest | 50 | 65 | 19° | 东南 SE | 13.5 | 70 | 枫香 Liquidambar formosana 麻栎 Quercus acutissima 樟树 Cinnamomum camphora |
| 天竺桂+红楠林 Cinnamomum japonicum+ Machilus thunbergii forest | 80 | 70 | 15° | 西 W | 9.5 | 100 | 红楠 Machilus thunbergii 天竺桂 Cinnamomum japonicum |
| 马尾松林 Pinus massoniana forest | 40 | 50 | 14° | 东北 NE | 13.5 | 80 | 马尾松 Pinus massoniana 石栎 Lithocarpus glabra 青冈 Cyclobalanopsis glauca |
| 青冈林 Cyclobalanopsis glauca forest | 60 | 153 | 15° | 东北 NE | 8.5 | 90 | 青冈 Cyclobalanopsis glauca 石栎 Lithocarpus glaber |
| 台湾蚊母树林 Distylium gracile forest | 160 | 241 | 8° | 东北 NE | 9.6 | 100 | 台湾蚊母树 Distylium gracile 铁冬青 Ilex rotunda 红楠 Machilus thunbergii 红山茶 Camellia longicaudata |
| 群落类型 Community type | 组分凋落量及占总凋落量百分比 Component production and percentage over the total | ||||
|---|---|---|---|---|---|
| 叶 Leaf | 枝 Twig | 花 Flower | 果 Fruit | 残体 Miscellaneous | |
| 枫香林 Liquidambar formosana forest | 2.73 (68.23) | 0.35 (8.62) | 0.19 (4.83) | 0.56 (14.05) | 0.17 (4.27) |
| 天竺桂+红楠林 Cinnamomum japonicum + Machilus thunbergii forest | 2.96 (77.57) | 0.28 (7.38) | 0.19 (4.95) | 0.18 (4.62) | 0.21 (5.47) |
| 马尾松林 Pinus massoniana forest | 3.09 (78.50) | 0.15 (3.72) | 0.17 (4.21) | 0.18 (4.49) | 0.36 (9.08) |
| 青冈林 Cyclobalanopsis glauca forest | 3.22 (82.85) | 0.21 (5.48) | 0.08 (2.10) | 0.08 (2.17) | 0.29 (7.40) |
| 台湾蚊母树林 Distylium gracile forest | 3.81 (71.08) | 0.98 (18.21) | 0.06 (1.08) | 0.15 (2.78) | 0.37 (6.85) |
表2 浙江普陀山岛5种森林凋落物各组分年凋落量(t·hm-2·a-1)及其占总凋落量的比例(括号中的数字为百分比, %)
Table 2 Characteristics of annual productions (t·hm-2·a-1) of different litterfall components of the five forest types in Putuoshan Island, Zhejiang, East China (Data in parentheses are percentages)
| 群落类型 Community type | 组分凋落量及占总凋落量百分比 Component production and percentage over the total | ||||
|---|---|---|---|---|---|
| 叶 Leaf | 枝 Twig | 花 Flower | 果 Fruit | 残体 Miscellaneous | |
| 枫香林 Liquidambar formosana forest | 2.73 (68.23) | 0.35 (8.62) | 0.19 (4.83) | 0.56 (14.05) | 0.17 (4.27) |
| 天竺桂+红楠林 Cinnamomum japonicum + Machilus thunbergii forest | 2.96 (77.57) | 0.28 (7.38) | 0.19 (4.95) | 0.18 (4.62) | 0.21 (5.47) |
| 马尾松林 Pinus massoniana forest | 3.09 (78.50) | 0.15 (3.72) | 0.17 (4.21) | 0.18 (4.49) | 0.36 (9.08) |
| 青冈林 Cyclobalanopsis glauca forest | 3.22 (82.85) | 0.21 (5.48) | 0.08 (2.10) | 0.08 (2.17) | 0.29 (7.40) |
| 台湾蚊母树林 Distylium gracile forest | 3.81 (71.08) | 0.98 (18.21) | 0.06 (1.08) | 0.15 (2.78) | 0.37 (6.85) |
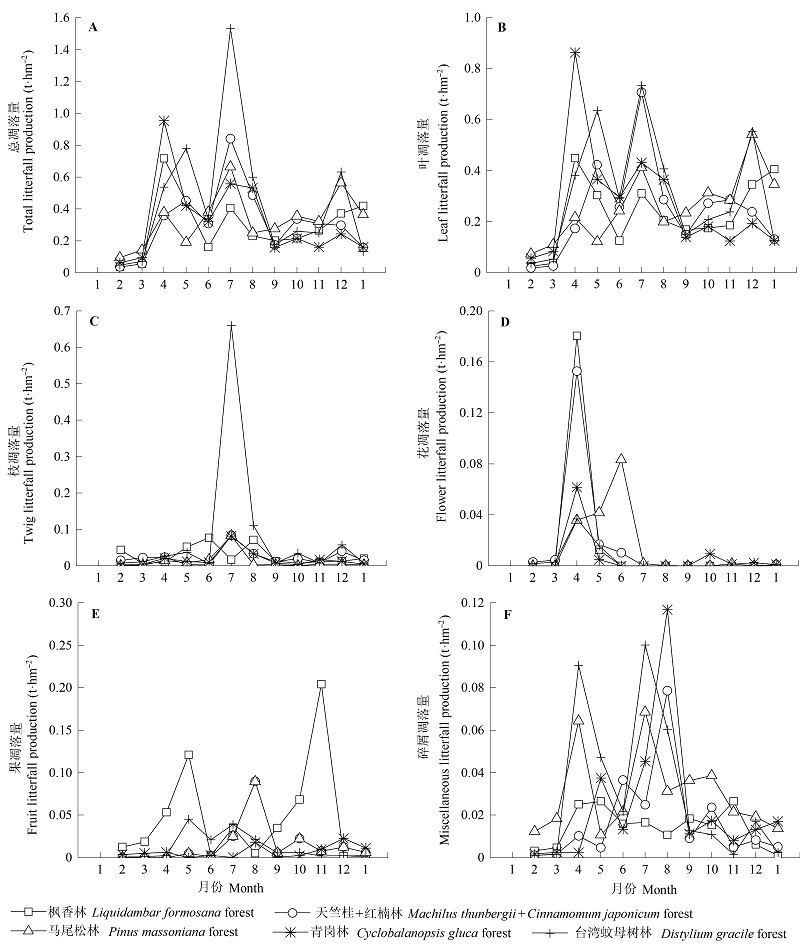
图1 浙江普陀山岛5种森林类型凋落物季节动态。
Fig. 1 Seasonal dynamics of litterfall production of the five forest types in Putuoshan Island, Zhejiang.
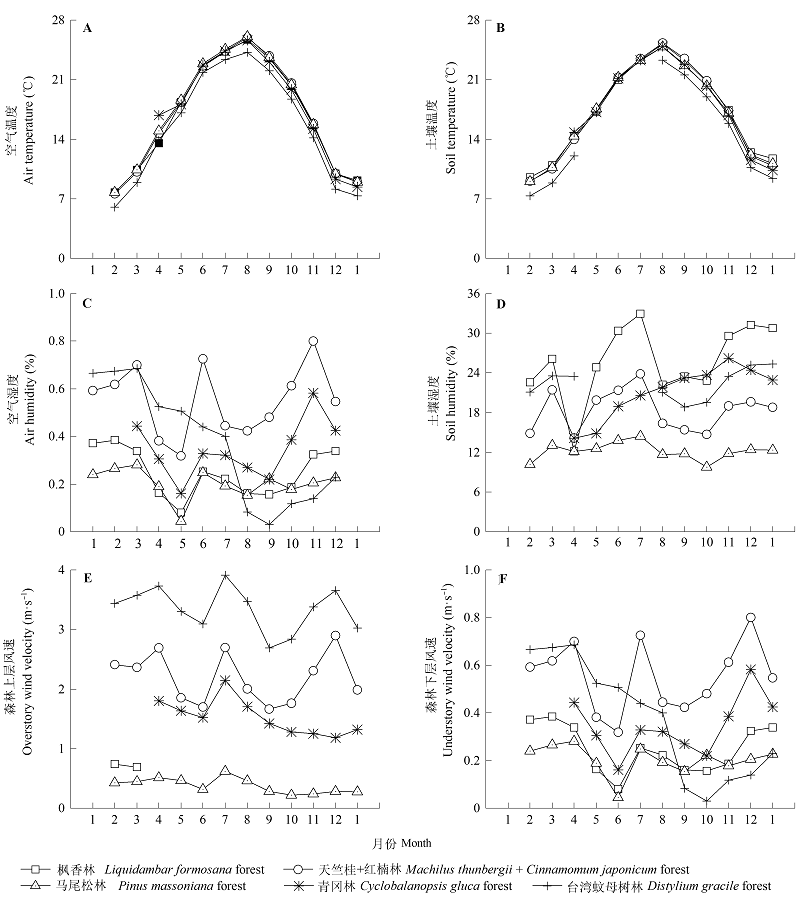
图2 浙江普陀山5种森林类型微气候月动态。
Fig. 2 Monthly dynamics of micro-climatic factors across the five forest types in Putuoshan Island, Zhejiang.
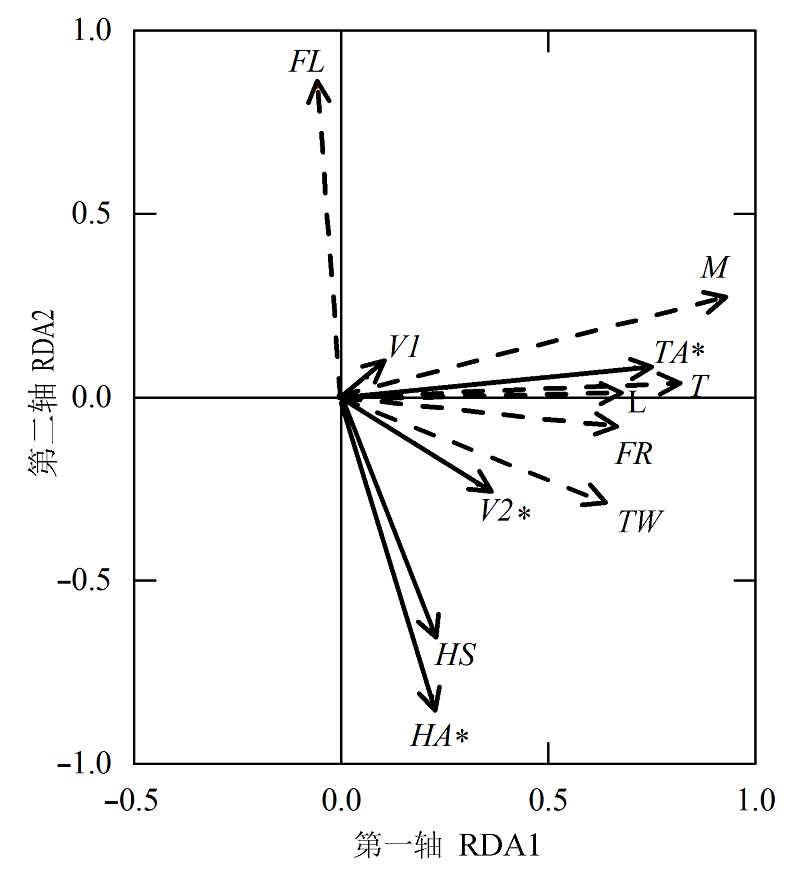
图3 浙江普陀山岛森林月总凋落量及各组分与微气候因子的冗余分析。实线向量为微气候因子, 虚线向量为森林月总凋落量及各组分。FL, FR, HA, HS, L, M, T, TA, TW, V1, V2分别代表花、果、空气温度、空气湿度、土壤湿度、叶、碎屑、总凋落量、枝、森林下层风速和森林上层风速。*, p < 0.05。
Fig. 3 Redundancy analyses (RDAs) calculated from monthly litterfall production (dash line vectors) in Putuoshan Island of Zhejiang and the monthly air temperature, air humidity, overstory wind velocity (bold line vectors) over one year. FL, flower litterfall production; FR, fruit litterfall production; HA, air humidity; HS, soil humidity; L, leaf litterfall production; M, miscellaneous; T, total litterfall production; TA, air temperature; TW, twig litterfall production; V1, understory wind velocity; V2, overstory wind velocity. *, p < 0.05.
| [1] | Berg B, Meentemeyer V (2001). Litter fall in some European coniferous forests as dependent on climate: A synthesis. Canadian Journal of Forest Research, 31, 292-301. |
| [2] | Borchert R (1996). Phenology and flowering periodicity of neotropical dry forest species: Evidence from herbarium collections.Journal of Tropical Ecology, 12, 65-80. |
| [3] | Brando PM, Nepstad DC, Davidson EA, Trumbore SE, Ray D, Camargo P (2008). Drought effects on litterfall, wood production and belowground carbon cycling in an Amazon forest: Results of a through fall reduction experiment.Philosophical Transactions of the Royal Society of London, 363, 1839-1848. |
| [4] | Chi Y, Shi HH, Guo Z, Ding DW (2015). Connotation, features and causes of island ecological vulnerability.Acta Oceanologica Sinica, 37, 93-105. (in Chinese with English abstract)[池源, 石洪华, 郭振, 丁德文 (2015). 海岛生态脆弱性的内涵、特征及成因探析. 海洋学报, 37, 93-105.] |
| [5] | Coley PD, Barone JA (1996). Herbivory and plant defenses in tropical forests. Annual Review of Ecology and Systemat- ics, 27, 305-335. |
| [6] | Dale JE (1982). The Growth of Leaves. Edward Arnold Limited, London. 31-41. |
| [7] | Friend A, Stevens A, Knox R, Cannell M (1997). A process- based, terrestrial biosphere model of ecosystem dynamics (Hybrid v3.0).Ecological Modelling, 95, 249-287. |
| [8] | Gosz JR, Likens GE, Bormann FH (1972). Nutrient content of litterfall on the Hubbard Brook experimental forest, New Hampshire.Ecology, 1972, 769-784. |
| [9] | Gross J (2003). Variance inflation factors.R News, 3, 13-15. |
| [10] | Guo J, Yu LH, Fang X, Xiang WH, Deng XW, Lu X (2015). Litter production and turnover in four types of subtropical forests in China.Acta Ecologica Sinica, 35, 4668-4677. (in Chinese with English abstract)[郭婧, 喻林华, 方晰, 项文化, 邓湘雯, 路翔 (2015). 中亚热带4种森林凋落物量、组成、动态及其周转期. 生态学报, 35, 4668-4677.] |
| [11] | Hoque MM, Mustafa Kamal AH, Idris MH, Haruna Ahmed O, Rafiqul Hoque ATM, Masum Billah M (2015). Litterfall production in a tropical mangrove of Sarawak, Malaysia.Zoology and Ecology, 25, 157-165. |
| [12] | Legendre P, Oksanen J, ter Braak CJF (2011). Testing the significance of canonical axes in redundancy analysis.Methods in Ecology and Evolution, 2, 269-277. |
| [13] | Lin KC, Hamburg SP, Tang S, Hsia YJ, Lin TC (2011). Typhoon effects on litterfall in a subtropical forest.Canadian Journal of Forest Research, 33, 2184-2192. |
| [14] | Liski J, Nissinen A, Erhard M, Taskinen O (2003). Climatic effects on litter decomposition from arctic tundra to tropical rainforest.Global Change Biology, 9, 575-584. |
| [15] | Liu CJ, Westman CJ, Berg B, Kutsch W, Wang GZ, Man RZ, Ilvesniemi H (2004). Variation in litterfall-climate relationships between coniferous and broadleaf forests in Eurasia.Global Ecology and Biogeography, 13, 105-114. |
| [16] | Lovett GM, Christenson LM, Groffman PM, Jones CG, Hart JE, Mitchell MJ (2010). Insect defoliation and nitrogen cycling in forests.Bioscience, 52, 335-341. |
| [17] | Ma WJ, Zhao YT, Zhang QQ, Arshad A, Shi QR, Yan ER (2014). C:N:P stoichiometry in forest floor litter of ever- green broad-leaved forests at different successional stages in Tiantong, Zhejiang, eastern China.Chinese Journal of Plant Ecology, 38, 833-842. (in Chinese with English abstract)[马文济, 赵延涛, 张晴晴, Ali Arshad, 史青茹, 阎恩荣 (2014). 浙江天童常绿阔叶林不同演替阶段地表凋落物的C:N:P化学计量特征. 植物生态学报, 38, 833-842.] |
| [18] | Ndakara OE (2011). Litterfall and nutrient returns isolated stands of xPersea gratissima(Avocado pear) in the rainforest zone of southern Nigeria. Ethiopian Journal of Environmental Studies and Management, 4, 42-50. |
| [19] | Ning XB, Xiang WH, Wang GJ, Fang X, Yan WD, Deng XW (2009). Litterfall production and dynamic for twenty years of a successive replanting Cunninghamia lanceolata plantation at Huitong, Hunan. Acta Ecologica Sinica, 29, 5122-5129. (in Chinese with English abstract)[宁晓波, 项文化, 王光军, 方晰, 闫文德, 邓湘雯 (2009). 湖南会同连作杉木林凋落物量20年动态特征. 生态学报, 29, 5122-5129.] |
| [20] | Oksanen J, Kindt R, Legendre P, O’Hara B, Stevens MHH, Oksanen MJ, Suggests MASS (2007). The vegan Package.Community Ecology Package, 10, 631-637. |
| [21] | Olena P, Nedret B (2007). Impact of deciduous tree species on litterfall quality, decomposition rates and nutrient circulation in pine stands.Forest Ecology and Manage- ment, 253, 11-18. |
| [22] | Qian YF, Yin LT, Hu JF, Zhang C, Yu SQ, Shen YL, Peng DQ (2012). Niche characteristics of main plant species in Putuo Mountain, Zhejiang Province of East China. Chinese Journal of Ecology, 31, 561-568. (in Chinese with English abstract)[钱逸凡, 伊力塔, 胡军飞, 张超, 余树全, 沈摇露, 彭东琴 (2012). 普陀山主要植物种生态位特征. 生态学杂志, 31, 561-568.] |
| [23] | Running SW, Hunt ER Jr.(1993). Generalization of a forest ecosystem process model for other biomes, Biome-BGC, and an application for global-scale models. In: Ehleringer JR, Field C eds. Scaling Physiological Processes: Leaf to Globe. Academic Press, San Diego, USA. |
| [24] | Scherer-Lorenzen M, Bonilla JL, Potvin C (2007). Tree species richness affects litter production and decomposition rates in a tropical biodiversity experiment.Oikos, 116, 2108-2124. |
| [25] | Sharma S, Hoque ATMR, Analuddin K, Hagihara A (2012). Litterfall dynamics in an overcrowded mangrove Kandelia obovata(S., L.) Yong stand over five years. Estuarine Coastal and Shelf Science, 98, 31-41. |
| [26] | Thuille A, Schulze ED (2006). Carbon dynamics in success- sional and afforested spruce stands in Thuringia and the Alps. Global Change Biology, 12, 325-342. |
| [27] | Valentini CMA, Sanches L, de Paula SR, Vourlitis GL, Nogueira JD, Pinto OB, Lobo FD (2008). Soil respiration and aboveground litter dynamics of a tropical transitional forest in northwest Mato Grosso, Brazil. Journal of Geophysical Research: Biogeosciences, 113, G00B10. doi: 10.1029/2007JG000619. |
| [28] | Wang HC, Lin KC, Huang CY (2016). Temporal and spatial patterns of remotely sensed litterfall in tropical and subtropical forests of Taiwan.Journal of Geophysical Research Biogeosciences, 121, 509-522. |
| [29] | Wen YG, Wei BE, Li JJ (1989). A study on the litter production and dynamics of subtropical forest.Scientia Silvae Sinicae, 25, 542-547. (in Chinese with English abstract)[温远光, 韦炳二, 黎洁娟 (1989). 亚热带森林凋落物产量及动态的研究. 林业科学, 25, 542-548.] |
| [30] | Weng H, Li ZA, Tu MZ, Yao WH (1993). The production and nutrient contents of litter in forests of Dinghushan mountain.Acta Phytoecologica Sinica, 17, 299-304. (in Chinese with English abstract)[翁轰, 李志安, 屠梦照, 姚文华 (1993). 鼎湖山森林凋落物量及营养元素含量研究. 植物生态学报, 17, 299-304.] |
| [31] | Xia YJ, Tang JQ, Zhang GF, Huang C, Meng FQ, Sun SC (2013). First and second sets of shoots in five evergreen woody species from Tiantong National Forest Park of Zhejiang, China.Chinese Journal of Plant Ecology, 37, 220-229. (in Chinese with English abstract)[夏洋洁, 唐坚强, 张光富, 黄超, 蒙凤群, 孙书存 (2013). 浙江天童国家森林公园5种常绿阔叶植物的一次和二次抽枝进程. 植物生态学报, 37, 220-229.] |
| [32] | Xu WM, Yan WD, Li JB, Zhao J, Wang GJ (2013). Amount and dynamic characteristics of litterfall in four forest types in subtropical China.Acta Ecologica Sinica, 33, 7570-7575. (in Chinese with English abstract)[徐旺明, 闫文德, 李洁冰, 赵晶, 王光军 (2013). 亚热带4种森林凋落物量及其动态特征. 生态学报, 33, 7570-7575.] |
| [33] | Xu XN, Hirata E, Shibata H (2004). Effect of typhoon disturbance on fine litterfall and related nutrient input in a subtropical forest on Okinawa Island, Japan.Basic and Applied Ecology, 5, 271-282. |
| [34] | Yan ER, Wang XH, Zhou W (2008). Characteristics of litterfall in relation to soil nutrients in mature and degraded evergreen broad-leaved forests of Tiantong, east China.Journal of Plant Ecology (Chinese Version), 32, 1-12. (in Chinese with English abstract)[阎恩荣, 王希华, 周武 (2008). 天童常绿阔叶林不同退化群落的凋落物特征及与土壤养分动态的关系. 植物生态学报, 32, 1-12.] |
| [35] | Yuan ZQ, Li BH, Bai XJ, Lin F, Shi S, Ye J, Wang XG, Hao ZQ (2010). Composition and seasonal dynamics of litterfalls in a broad-leaved Korean pine (Pinus koraiensis) mixed forest in Changbai Mountains, Northeast China. Chinese Journal of Applied Ecology, 21, 2171-2178. (in Chinese with English abstract)[原作强, 李步杭, 白雪娇, 蔺菲, 师帅, 叶吉, 王绪高, 郝占庆 (2010). 长白山阔叶红松林凋落物组成及其季节动态. 应用生态学报, 21, 2171-2178.] |
| [36] | Zhang HC, Yuan WP, Dong WJ, Liu SG (2014). Seasonal patterns of litterfall in forest ecosystem worldwide.Ecological Complexity, 20, 240-247. |
| [37] | Zhang QQ, Zhou LL, Zhao YT, Xu MS, Yan ER (2016). Litter dynamics of plants in a successional series of evergreen broad-leaved forests in Tiantong region, Zhejiang Province.Chinese Journal of Ecology, 35, 290-299. (in Chinese with English abstract)[张晴晴, 周刘丽, 赵延涛, 许洺山, 阎恩荣 (2016). 浙江天童常绿阔叶林演替系列植物叶片的凋落节律. 生态学杂志, 35, 290-299.] |
| [38] | Zhang XP, Wang XP, Zhu B, Zong ZJ, Peng CH, Fang JY (2008). Litterfall production in relation to environmental factors in northeast China’s forests.Journal of Plant Ecology (Chinese Version), 32, 1031-1040. (in Chinese with English abstract)[张新平, 王襄平, 朱彪, 宗占江, 彭长辉, 方精云 (2008). 我国东北主要森林类型的凋落物产量及其影响因素. 植物生态学报, 32, 1031-1040.] |
| [39] | Zhao CL, Ou DY, Hu JF, Zhou WP, Zhao Y (2009). Resource evaluation and protection countermeasure of ancient and famous trees in Putuo Mountain.Journal of Zhejiang Ocean University (Natural Science), 28, 200-204. (in Chinese with English abstract)[赵慈良, 欧丹燕, 胡军飞, 周伟平, 赵颖 (2009). 普陀山古树名木资源评价与保护对策. 浙江海洋学院学报: 自然科学版, 28, 200-204.] |
| [40] | Zheng Z, Li YR, Liu HM, Feng ZL, Gan JM, Kong WJ (2005). Litterfall of tropical rainforests at different altitudes, Xishuangbanna, Southwest China.Acta Phytoecologica Sinica, 29, 884-893. (in Chinese with English abstract)[郑征, 李佑荣, 刘宏茂, 冯志立, 甘建民, 孔维静 (2005). 西双版纳不同海拔热带雨林凋落量变化研究. 植物生态学报, 29, 884-893.] |
| [41] | Zou B, Li ZA, Ding YZ, Tan WN (2006). Litterfall of common plantations in south subtropical China.Acta Ecologica Sinica, 26, 715-721. (in Chinese with English abstract)[邹碧, 李志安, 丁永祯, 谭万能 (2006). 南亚热带4种人工林凋落物动态特征. 生态学报, 26, 715-721.] |
| [1] | 郑宁, 李素英, 王鑫厅, 吕世海, 赵鹏程, 臧琛, 许玉珑, 何静, 秦文昊, 高恒睿. 基于环境因子对叶绿素影响的典型草原植物生活型优势研究[J]. 植物生态学报, 2022, 46(8): 951-960. |
| [2] | 郝建锋, 周润惠, 姚小兰, 喻静, 陈聪琳, 向琳, 王姚瑶, 苏天成, 齐锦秋. 二代野猪放牧对夹金山针阔混交林物种多样性与土壤理化性质的影响[J]. 植物生态学报, 2022, 46(2): 197-207. |
| [3] | 张央, 安明态, 武建勇, 刘锋, 汪伟. 中国兜兰属宽瓣亚属植物地理分布格局及其主导气候因子[J]. 植物生态学报, 2022, 46(1): 40-50. |
| [4] | 陈林, 王磊, 杨新国, 宋乃平, 李月飞, 苏莹, 卞莹莹, 祝忠有, 孟文婷. 荒漠草原猪毛蒿种群繁殖特征的土壤驱动因子分析[J]. 植物生态学报, 2019, 43(1): 65-76. |
| [5] | 梁士楚, 刘润红, 荣春艳, 常斌, 姜勇. 漓江河岸带植物功能性状变异与关联[J]. 植物生态学报, 2019, 43(1): 16-26. |
| [6] | 李静鹏, 郑志荣, 赵念席, 高玉葆. 刈割、围封、放牧三种利用方式下草原生态系统的多功能性与植物物种多样性之间的关系[J]. 植物生态学报, 2016, 40(8): 735-747. |
| [7] | 于健, 徐倩倩, 刘文慧, 罗春旺, 杨君珑, 李俊清, 刘琪璟. 长白山东坡不同海拔长白落叶松径向生长对气候变化的响应[J]. 植物生态学报, 2016, 40(1): 24-35. |
| [8] | 王晓春, 宋来萍, 张远东. 大兴安岭北部樟子松树木生长与气候因子的关系[J]. 植物生态学报, 2011, 35(3): 294-302. |
| [9] | 王瑞霞, 何明高, 宋松泉. 培养基与光照对沼兰种子非共生萌发的影响[J]. 植物生态学报, 2010, 34(4): 438-443. |
| [10] | 春敏莉, 谢宗强, 赵常明, 樊大勇, 徐新武, 平亮. 神农架巴山冷杉天然林凋落量及养分特征[J]. 植物生态学报, 2009, 33(3): 492-498. |
| [11] | Pierre Legendre. β-多样性的研究:应用多元回归和典范分析研究生态方差的分解[J]. 植物生态学报, 2007, 31(5): 976-981. |
| [12] | 于明坚, 陈启常, 李铭红, 常杰, 潘晓东, 陈增鸿, 邵剑波. 浙江建德青冈常绿阔叶林凋落量研究[J]. 植物生态学报, 1996, 20(2): 144-150. |
| [13] | 李景文, 刘传照, 任淑文, 詹鸿振. 天然枫桦红松林凋落量动态及养分归还量[J]. 植物生态学报, 1989, 13(1): 42-48. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
Copyright © 2022 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19