
植物生态学报 ›› 2011, Vol. 35 ›› Issue (8): 864-871.DOI: 10.3724/SP.J.1258.2011.00864 cstr: 32100.14.SP.J.1258.2011.00864
刘富俊1, 黎云祥1, 廖咏梅1,*( ), 陈劲松2,*(), 权秋梅1, 龚新越1
), 陈劲松2,*(), 权秋梅1, 龚新越1
收稿日期:2011-04-26
接受日期:2011-06-01
出版日期:2011-04-26
发布日期:2011-07-28
作者简介: 共同通讯作者 (E-mail: lym3326@126.com; cjs74@163.com)
LIU Fu-Jun1, LI Yun-Xiang1, LIAO Yong-Mei1,*(), CHEN Jin-Song2,*(), QUAN Qiu-Mei1, GONG Xin-Yue1
Received:2011-04-26
Accepted:2011-06-01
Online:2011-04-26
Published:2011-07-28
摘要:
采用盆栽试验研究了异质性重金属镉胁迫下, 克隆整合对匍匐茎草本植物积雪草(Centella asiatica)生长的影响。将远端分株(相对年幼的分株)分别置于对照和镉胁迫处理下, 并对远端分株与近端分株(相对年长的分株)之间的匍匐茎进行切断或保持连接处理。研究结果显示: 镉胁迫处理显著降低了积雪草远端分株的净光合速率(Pn)、最大光量子产量(Fv/Fm)、叶绿素含量、叶面积、分株数和生物量; 克隆整合缓解了镉胁迫对远端分株生长的不利影响; 克隆整合不仅未导致相连近端分株的损耗, 而且相连近端分株的光合效率也没有表现出补偿性增加; 克隆整合降低了远端受胁迫分株的根冠比, 从而使之减少了对土壤中重金属镉的吸收; 匍匐茎切断和镉胁迫处理对近端分株、远端分株的叶柄长没有显著的影响。结果表明: 克隆整合提高了积雪草遭受镉胁迫的远端分株的生长, 改变了其生物量分配格局, 并有助于整个克隆片段在异质性重金属胁迫下的生长。该研究对于丰富和发展异质性环境胁迫下克隆整合的生态适应对策具有重要意义。
刘富俊, 黎云祥, 廖咏梅, 陈劲松, 权秋梅, 龚新越. 异质性重金属镉胁迫下克隆整合对匍匐茎草本植物积雪草生长的影响. 植物生态学报, 2011, 35(8): 864-871. DOI: 10.3724/SP.J.1258.2011.00864
LIU Fu-Jun, LI Yun-Xiang, LIAO Yong-Mei, CHEN Jin-Song, QUAN Qiu-Mei, GONG Xin-Yue. Effects of clonal integration on growth of stoloniferous herb Centella asiaticasuffering from heterogeneous heavy metal Cd<sup>2+</sup>stress. Chinese Journal of Plant Ecology, 2011, 35(8): 864-871. DOI: 10.3724/SP.J.1258.2011.00864
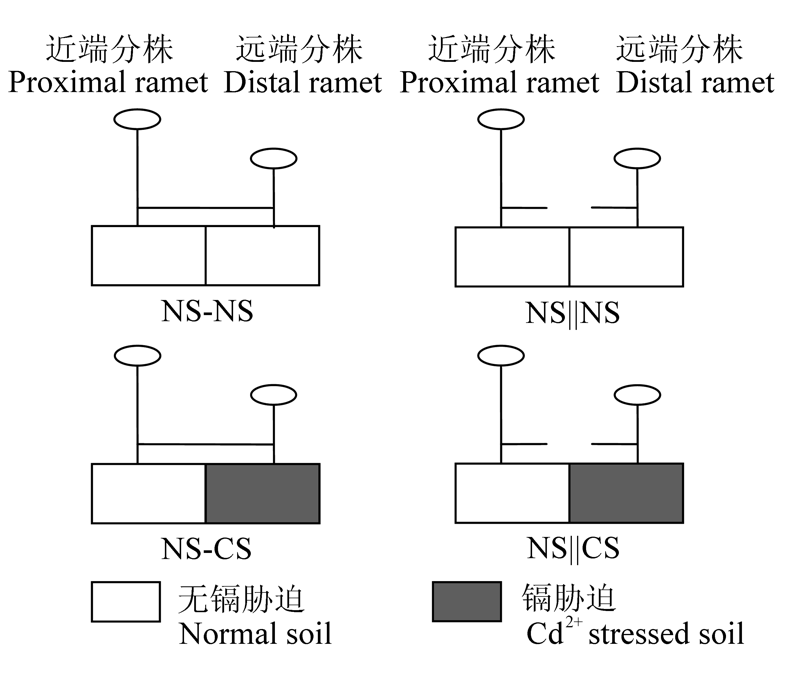
图1 试验设计图解。NS-CS, 匍匐茎保持连接且对远端分株实施镉胁迫; NS||CS, 匍匐茎切断且对远端分株实施镉胁迫; NS-NS, 匍匐茎保持连接且对远端分株不实施镉胁迫; NS||NS, 匍匐茎切断且对远端分株不实施镉胁迫。
Fig. 1 Schematic representation of the experimental design. NS-CS, stolon connections left intact and distal ramets are assigned to Cd2+ stressed soil; NS||CS, stolon connections are severed and distal ramets are assigned to Cd2+ stressed soil; NS-NS, stolon connections are left intact and distal ramets are assigned to normal soil; NS||NS, stolon connections are severed and distal ramets are assigned to normal soil.
| 叶柄长 Petiole length | 叶面积 Leaf area | 分株数 Number of ramets | 生物量 Biomass | 根冠比 Root to shoot ratio | |
|---|---|---|---|---|---|
| 近端分株 Proximal ramets, df(1,12) | |||||
| S | 0.14ns | 0.01ns | 25.00** | 0.56ns | 1.50ns |
| C | 0.01ns | 0.02ns | 1.00ns | 0.16ns | 2.27ns |
| S × C | 0.04ns | 0.08ns | 1.00ns | 1.55ns | 2.39ns |
| 远端分株 Distal ramets, df(1,12) | |||||
| S | 0.40ns | 1.84ns | 6.25* | 0.96ns | 1.83ns |
| C | 3.46ns | 11.01** | 20.25** | 39.38** | 9.08* |
| S × C | 0.08ns | 1.45ns | 2.25ns | 5.25* | 3.44ns |
| 克隆片段 Whole clonal fragments, df(1,16) | |||||
| S | - | 1.58ns | 0.00ns | 1.74ns | - |
| C | - | 21.66** | 10.67* | 34.70** | - |
| S × C | - | 3.20ns | 0.67ns | 4.95* | - |
表1 匍匐茎切断(S)和镉胁迫(C)及其交互作用(S × C)对积雪草近端、远端分株和(或)整个克隆片段叶柄长、叶面积、分株数、生物量和根冠比的双因素方差分析
Table 1 Results of two-way analysis of variance on the effects of stolen severing (S), Cd2+ stress (C) and their interaction (S × C) on petiole length, leaf area, number of ramets, biomass and root to shoot ratio of the proximal, distal ramets and/or the whole clonal fragments of Centella asiatica
| 叶柄长 Petiole length | 叶面积 Leaf area | 分株数 Number of ramets | 生物量 Biomass | 根冠比 Root to shoot ratio | |
|---|---|---|---|---|---|
| 近端分株 Proximal ramets, df(1,12) | |||||
| S | 0.14ns | 0.01ns | 25.00** | 0.56ns | 1.50ns |
| C | 0.01ns | 0.02ns | 1.00ns | 0.16ns | 2.27ns |
| S × C | 0.04ns | 0.08ns | 1.00ns | 1.55ns | 2.39ns |
| 远端分株 Distal ramets, df(1,12) | |||||
| S | 0.40ns | 1.84ns | 6.25* | 0.96ns | 1.83ns |
| C | 3.46ns | 11.01** | 20.25** | 39.38** | 9.08* |
| S × C | 0.08ns | 1.45ns | 2.25ns | 5.25* | 3.44ns |
| 克隆片段 Whole clonal fragments, df(1,16) | |||||
| S | - | 1.58ns | 0.00ns | 1.74ns | - |
| C | - | 21.66** | 10.67* | 34.70** | - |
| S × C | - | 3.20ns | 0.67ns | 4.95* | - |
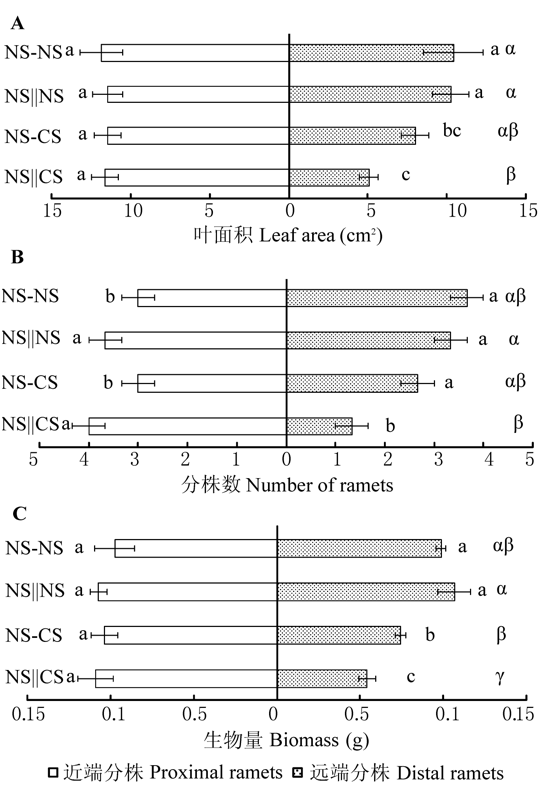
图2 近端、远端分株和整个克隆片段的叶面积(A)、分株数(B)和生物量(C) (平均值±标准误差)。对于近端和远端分株, 相同的小写字母表示各处理间差异不显著(p = 0.05); 对于整个克隆片段, 相同的希腊字母表示各处理间差异不显著 (p = 0.05)。NS-CS, 匍匐茎保持连接且对远端分株实施镉胁迫; NS||CS, 匍匐茎切断且对远端分株实施镉胁迫; NS-NS, 匍匐茎保持连接且对远端分株不实施镉胁迫; NS||NS, 匍匐茎切断且对远端分株不实施镉胁迫。
Fig. 2 Leaf area (A), number of ramets (B) and biomass (C) of the proximal, distal ramets and the whole clonal fragments (mean ± SE). For the proximal and distal ramets, the same lowercase letters are not significantly different at p = 0.05, for the whole clonal fragments, the same Greek letters are not significantly different at p = 0.05. NS-CS, stolon connections left intact and distal ramets are assigned to Cd 2+ stressed soil; NS||CS, stolon connections are severed and distal ramets are assigned to Cd2+ stressed soil; NS-NS, stolon connections are left intact and distal ramets are assigned to normal soil; NS||NS, stolon connections are severed and distal ramets are assigned to normal soil.
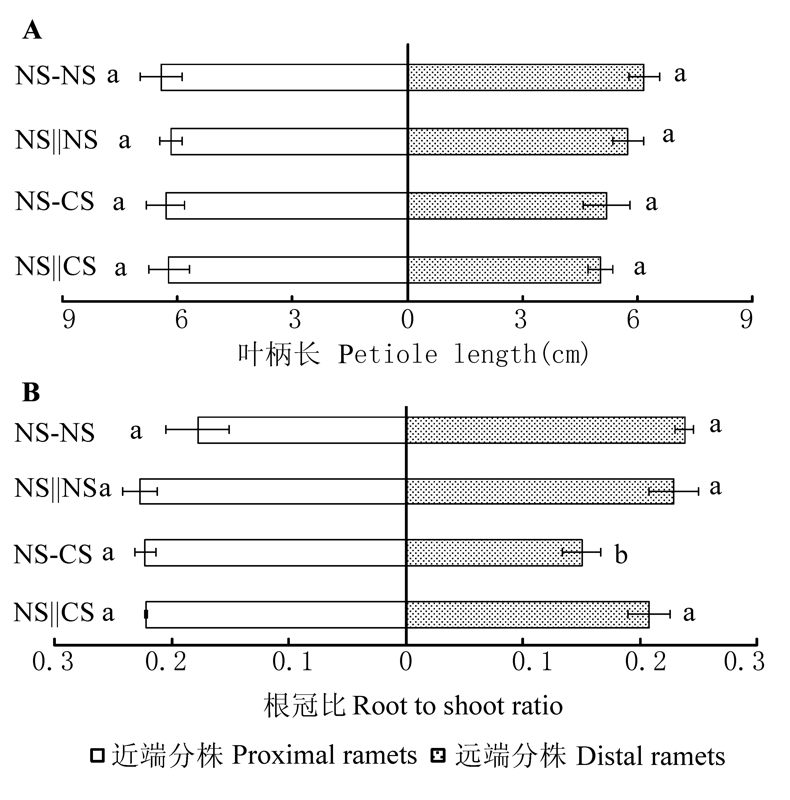
图3 近端、远端分株的叶柄长(A)和根冠比(B) (平均值±标准误差)。相同小写字母表示各处理间差异不显著(p = 0.05)。NS-CS, 匍匐茎保持连接且对远端分株实施镉胁迫; NS||CS, 匍匐茎切断且对远端分株实施镉胁迫; NS-NS, 匍匐茎保持连接且对远端分株不实施镉胁迫; NS||NS, 匍匐茎切断且对远端分株不实施镉胁迫。
Fig. 3 Petiole length (A) and root to shoot ratio (B) of the proximal and distal ramets (mean ± SE). The bars with the same lowercase letters are not significantly different at p = 0.05. NS-CS, stolon connections left intact and distal ramets are assigned to Cd 2+ stressed soil; NS||CS, stolon connections are severed and distal ramets are assigned to Cd2+ stressed soil; NS-NS, stolon connections are left intact and distal ramets are assigned to normal soil; NS||NS, stolon connections are severed and distal ramets are assigned to normal soil.
| 近端分株 Proximal ramets | 远端分株 Distal ramets | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Pn | Pn | Fv/Fm | SPAD | ||||||||
| df | F | df | F | df | F | df | F | ||||
| 组间效应 Between-subject effects | |||||||||||
| S | 1 | 0.03ns | 1 | 6.49* | 1 | 5.71* | 1 | 5.18* | |||
| C | 1 | 0.54ns | 1 | 66.00** | 1 | 5.89* | 1 | 35.29** | |||
| S × C | 1 | 0.39ns | 1 | 10.96 * | 1 | 2.38ns | 1 | 1.80ns | |||
| 误差 Error | 8 | 8 | 8 | 12 | |||||||
| 组内效应 Within-subject effects | |||||||||||
| 次数 Time | 3 | 0.41ns | 3 | 0.24ns | 2 | 0.84ns | 3 | 5.58* | |||
| 切断×次数 S × time | 3 | 0.17ns | 3 | 0.17ns | 2 | 2.20ns | 3 | 0.48 ns | |||
| 镉胁迫×次数 C × time | 3 | 0.14ns | 3 | 2.75ns | 2 | 5.17ns | 3 | 5.66* | |||
| 切断×镉胁迫×次数 S × C × time | 3 | 1.68ns | 3 | 0.14ns | 2 | 1.29ns | 3 | 0.23ns | |||
| 误差 Error | 24 | 24 | 16 | 36 | |||||||
表2 积雪草远端分株净光合速率(Pn)、叶绿素荧光(最大光量子产量, Fv/Fm)和叶绿素含量(SPAD values)以及近端分株的Pn的双因素重复测量方差分析, 以匍匐茎切断(S)和镉胁迫(C)作为组间因素
Table 2 Results of Two-way repeated-measure analysis of variance, with stolen severing (S) and Cd2+ stress (C) as between-subject effects, for differences in net photosynthesis rate (Pn), chlorophyll fluorescence (maximum quantum yield of photosystem II, Fv/Fm) and chlorophyll contents (SPAD values) between distal ramets and Pn between proximal ramets of Centella asiatica
| 近端分株 Proximal ramets | 远端分株 Distal ramets | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Pn | Pn | Fv/Fm | SPAD | ||||||||
| df | F | df | F | df | F | df | F | ||||
| 组间效应 Between-subject effects | |||||||||||
| S | 1 | 0.03ns | 1 | 6.49* | 1 | 5.71* | 1 | 5.18* | |||
| C | 1 | 0.54ns | 1 | 66.00** | 1 | 5.89* | 1 | 35.29** | |||
| S × C | 1 | 0.39ns | 1 | 10.96 * | 1 | 2.38ns | 1 | 1.80ns | |||
| 误差 Error | 8 | 8 | 8 | 12 | |||||||
| 组内效应 Within-subject effects | |||||||||||
| 次数 Time | 3 | 0.41ns | 3 | 0.24ns | 2 | 0.84ns | 3 | 5.58* | |||
| 切断×次数 S × time | 3 | 0.17ns | 3 | 0.17ns | 2 | 2.20ns | 3 | 0.48 ns | |||
| 镉胁迫×次数 C × time | 3 | 0.14ns | 3 | 2.75ns | 2 | 5.17ns | 3 | 5.66* | |||
| 切断×镉胁迫×次数 S × C × time | 3 | 1.68ns | 3 | 0.14ns | 2 | 1.29ns | 3 | 0.23ns | |||
| 误差 Error | 24 | 24 | 16 | 36 | |||||||
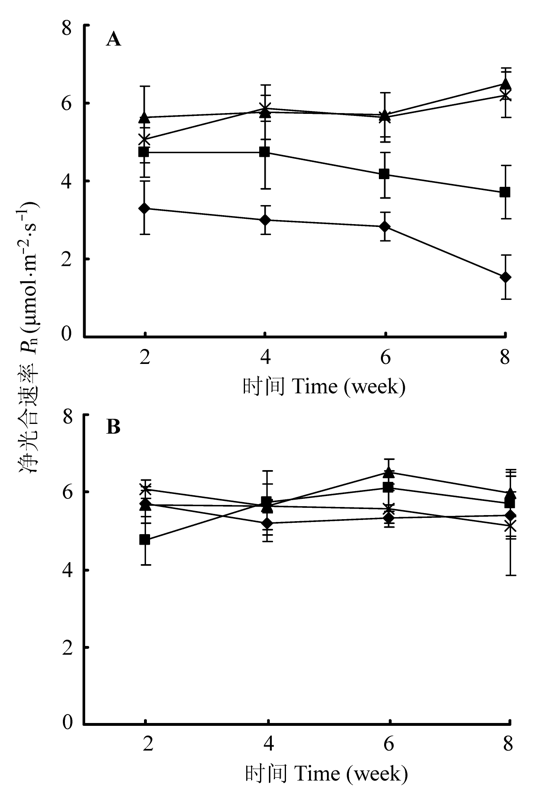
图4 远端分株(A)和近端分株(B)的净光合速率(Pn)随时间的变化(平均值±标准误差)。◆、■、▲和×分别表示匍匐茎切断且对远端分株实施镉胁迫处理、匍匐茎保持连接且对远端分株实施镉胁迫处理、匍匐茎切断且对远端分株不实施镉胁迫处理和匍匐茎保持连接且对远端分株不实施镉胁迫处理。
Fig. 4 Variations of net photosynthetic rate (Pn) of distal ramets (A) and proximal ramets (B) with time (mean ± SE). ◆, ■, ▲ and × represent stolon connections are severed and distal ramets are assigned to Cd 2+ stressed soil, stolon connections left intact and distal ramets are assigned to Cd2+ stressed soil, stolon connections are severed and distal ramets are assigned to normal soil, and stolon connections are left intact and distal ramets are assigned to normal soil treatments, respectively.
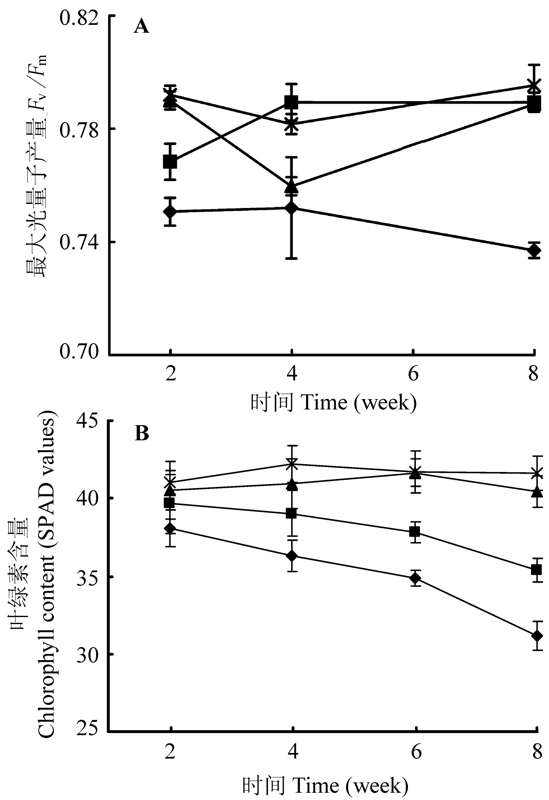
图5 远端分株的光系统II最大光量子产量(A)和叶绿素含量(B)随时间的变化(平均值±标准误差)。◆、■、▲和×分别表示匍匐茎切断且对远端分株实施镉胁迫处理、匍匐茎保持连接且对远端分株实施镉胁迫处理、匍匐茎切断且对远端分株不实施镉胁迫处理和匍匐茎保持连接且对远端分株不实施镉胁迫处理。远端分株的第6周测定的最大光量子产量数据不可用。
Fig. 5 Variations of maximum quantum yields of photosystem II (A) and chlorophyll contents (B) of distal ramets with time (mean ± SE). ◆, ■, ▲ and × represent stolon connections are severed and distal ramets are assigned to Cd 2+ stressed soil, stolon connections left intact and distal ramets are assigned to Cd2+ stressed soil, stolon connections are severed and distal ramets are assigned to normal soil, and stolon connections are left intact and distal ramets are assigned to normal soil treatments, respectively. Data on maximum quantum yields of photosystem II in distal ramets is not available at sixth week.
| [1] | Bell G, Lechowicz MJ (1994). Spatial heterogeneity at small scales and how plants respond to it. In: Caldwell MM, Pearcy RW eds. Exploitation of Environmental Heterogeneity by Plants: Ecophysiological Processes Above- and Belowground. Academic Press, San Diego.391-414. |
| [2] |
Björkman O, Demmig B (1987). 77 K among vascular plants of diverse origins. Planta, 170,489-504.
URL PMID |
| [3] | Bolhàr-Nordenkampf HR, Long SP, Baker NR, Öquist G, Scheiber U, Lechner EG (1989). Chlorophyll fluorescence as a probe of the photosynthetic competence of leaves in the field: a review of current instrumentation. Functional Ecology, 3,497-514. |
| [4] | Chen JS, Lei NF, Dong M (2010). Clonal integration improves the tolerance of Carex praeclara to sand burial by compensatory response . Acta Oecologica, 36,23-28. |
| [5] | Delectis Florae Reipublicae Popularis Sinicae Agendae Academiae Sinicae Edita (中国科学院中国植物志编辑委员会) (1979). Flora Reipublicae Popularis Sinicae (中国植物志), 55 (1). Science Press, Beijing.31-33. (in Chinese) |
| [6] | Hartnett DC, Bazzaz FA (1983). Physiological integration among intraclonal ramets in Solidago canadensis. Ecology, 64,779-788. |
| [7] | He ZS, He WM, Yu FH, Shi PL, Zhang XZ, He YT, Zhong ZM, Dong M (2007). Do clonal growth form and habitat origin affect resource-induced plasticity in Tibetan alpine herbs? Flora, 202,408-416. |
| [8] | Jiang LF (姜丽芬), Shi FC (石福臣), Wang HT (王化田), Zu YG (祖元刚) (2005). Application tryout of chlorophyll meter SPAD-502 in forestry. Chinese Journal of Ecology (生态学杂志), 24,1543-1548. (in Chinese with English abstract) |
| [9] | Johnson GN, Scholes JD, Young AJ, Horton P (1993). The dissipation of excess excitation energy in British plant species. Plant, Cell & Environment, 16,673-679. |
| [10] | Jónsdóttir IS, Callaghan TV (1989). Localized defoliation stress and the movement of 14C-photoassimilates between tillers of Carex bigelowii. Oikos , 54,211-219. |
| [11] | Krupa Z (1988). Cadmium-induced changes in the composition and structure of the light-harvesting chlorophyll a/b protein complex II in radish cotyledons. Physiologia Plantarum, 73,518-524. |
| [12] | Liu FH, Ye XH, Yu FH, Dong M (2006). Clonal integration modifies responses of Hedysarum laeve to local sand burial in Mu Us sandland . Journal of Plant Ecology, 30,278-285. |
| [13] |
Markwell J, Osterman JC, Mitchell JL (1995). Calibration of the Minolta SPAD-502 leaf chlorophyll meter. Photosynthesis Research, 46,467-472.
DOI URL PMID |
| [14] | Oborny B, Cain ML (1997). Models of spatial spread and foraging in clonal plants. In: de Kroon H, van GroenendaelJ eds . The Ecology and Evolution of Clonal Plants. Backhuys Publishers, Leiden.155-183. |
| [15] | Oborny B, Podani J (1996). The Role of Clonality in Plant Communities. Opulus Press, Uppsala. |
| [16] | Prasad MNV (1995). Cadmium toxicity and tolerance in vascular plants. Environmental and Experimental Botany, 35,525-545. |
| [17] | Roiloa SR, Retuerto R (2006). Physiological integration ameliorates effects of serpentine soils in the clonal herb Fragaria vesca. Physiologia Plantarum, 128,662-676. |
| [18] | Roiloa SR, Retuerto R (2007). Responses of the clonal Fragaria vesca to microtopographic heterogeneity under different water and light conditions . Environmental and Experimental Botany, 61,1-9. |
| [19] | Shumway SW (1995). Physiological integration among clonal ramets during invasion of disturbance patches in a New England salt marsh. Annals of Botany, 76,225-233. |
| [20] |
Stuefer JF, Hutchings MJ (1994). Environmental heterogeneity and clonal growth: a study of the capacity for reciprocal translocation in Glechoma hederacea L . Oecologia, 100,302-308.
URL PMID |
| [21] | Suzuki N, Koizumi N, Sano H (2001). Screening of cadmium-responsive genes in Arabidopsis thaliana . Plant, Cell & Environment, 24,1177-1188. |
| [22] |
van Kleunen M, Fischer M, Schmid B (2000). Clonal integration in Ranunculus reptans: by-product or adaptation ? Journal of Evolutionary Biology, 13,237-248.
DOI URL |
| [23] | van Kleunen M, Stuefer JF (1999). Quantifying the effects of reciprocal assimilate and water translocation in a clonal plant by the use of steam-girdling. Oikos, 85,135-145. |
| [24] | Wang H (汪洪), Zhao SC (赵士诚), Xia WJ (夏文建), Wang XB (王秀斌), Fan HL (范洪黎), Zhou W (周卫) (2008). Effect of cadmium stress on photosynthesis, lipid peroxidation and antioxidant enzyme activities in maize ( Zea mays L.) seedings . Plant Nutrition and Fertilizer Science (植物营养与肥料学报), 14,36-42. (in Chinese with English abstract) |
| [25] |
Wang N, Yu FH, Li PX, He WM, Liu FH, Liu JM, Dong M (2008). Clonal integration affects growth, photosynthetic efficiency and biomass allocation, but not the competitive ability, of the alien invasive Alternanthera philoxeroides under severe stress . Annals of Botany, 101,671-678.
DOI URL PMID |
| [26] | Yang HL (杨慧玲), Xue RL (薛瑞丽), Ye YZ (叶永忠), Wang HQ (王会勤), Dong M (董鸣), Huang ZY (黄振英) (2009). Effects of clonal integration on growth and survival of Bromus inermis (Poaceae) in heterogeneous saline environments of the Otindag Sandland. Acta Ecologica Sinica (生态学报) , 29,2827-2834. (in Chinese with English abstract) |
| [27] | Yu FH, Chen YF, Dong M (2002). Clonal integration enhances survival and performance of Potentilla anserina, suffering from partial sand burial on Ordos plateau, China . Evolutionary Ecology, 15,303-318. |
| [28] | Yu FH, Dong M, Krüsi B (2004). Clonal integration helps Psammochloa villosa survive sand burial in an inland dune . New Phytologist, 162,697-704. |
| [29] | Zhang XY (张想英), Fan DY (樊大勇), Xie ZQ (谢宗强), Xiong GM (熊高明), Li ZJ (李兆佳) (2010). Clonal integration enhances performance of Cynodon dactylon subjected to submergence . Chinese Journal of Plant Ecology (植物生态学报), 34,1075-1083. (in Chinese with English abstract) |
| [1] | 戴丽君, 向玲艺, 蹇陈, 王晓锋. 三峡回水扰动增强入库小流域河岸带典型草本植物功能性状的局域分化[J]. 植物生态学报, 2026, 50(2): 442-460. |
| [2] | 张箫荻, 王晓霞, 章毓文, 侯靖雨, 石骁鹏, 和璐璐, 刘亚栋, 薛柳, 何宝华, 段劼. 北京山区三种林下灌木水力结构、叶片功能性状及其环境适应策略[J]. 植物生态学报, 2025, 49(7): 1128-1143. |
| [3] | 李姝雯, 汤璐瑶, 张博纳, 叶琳峰, 童金莲, 谢江波, 李彦, 王忠媛. 降水梯度带榆树枝叶协作关系的区域分异规律[J]. 植物生态学报, 2025, 49(2): 282-294. |
| [4] | 童金莲, 张博纳, 汤璐瑶, 叶琳峰, 李姝雯, 谢江波, 李彦, 王忠媛. C4植物狗尾草功能性状网络沿降水梯度带的区域分异规律[J]. 植物生态学报, 2025, 49(11): 1817-1832. |
| [5] | 陈雪纯, 刘虹, 朱少琦, 孙铭遥, 宇振荣, 王庆刚. 漓江流域不同弃耕年限下4种常见草本植物功能性状种内变化及其影响因素[J]. 植物生态学报, 2023, 47(4): 559-570. |
| [6] | 王雪梅, 闫帮国, 史亮涛, 刘刚才. 车桑子幼苗生物量分配与叶性状对氮磷浓度的响应差异[J]. 植物生态学报, 2020, 44(12): 1247-1261. |
| [7] | 李建军, 刘恋, 陈迪马, 许丰伟, 程军回, 白永飞. 底座入土深度和面积对典型草原土壤呼吸测定结果的影响[J]. 植物生态学报, 2019, 43(2): 152-164. |
| [8] | 周玮, 李洪波, 曾辉. 西藏高寒草原群落植物根系属性在降水梯度下的变异格局[J]. 植物生态学报, 2018, 42(11): 1094-1102. |
| [9] | 颉洪涛, 虞木奎, 成向荣. 光照强度变化对5种耐阴植物氮磷养分含量、分配以及限制状况的影响[J]. 植物生态学报, 2017, 41(5): 559-569. |
| [10] | 张峰, 周广胜. 玉米农田冠层光合参数的多光谱遥感反演[J]. 植物生态学报, 2014, 38(7): 710-719. |
| [11] | 郭京衡, 曾凡江, 李尝君, 张波. 塔克拉玛干沙漠南缘三种防护林植物根系构型及其生态适应策略[J]. 植物生态学报, 2014, 38(1): 36-44. |
| [12] | 魏宇航,周晓波,陈劲松,谌利民,李娇,刘庆. 模拟采食干扰下克隆整合对两种箭竹分株种群更新的影响[J]. 植物生态学报, 2013, 37(8): 699-708. |
| [13] | 叶学华,胡宇坤,刘志兰,高树琴,董鸣. 水分异质性影响两种根茎型克隆植物赖草和假苇拂子茅的水分存储能力[J]. 植物生态学报, 2013, 37(5): 427-435. |
| [14] | 丁继军,潘远智,李丽,刘柿良,崔明峰,高佩刚. 外源谷胱甘肽对石竹幼苗镉毒害的缓解效应[J]. 植物生态学报, 2013, 37(10): 950-960. |
| [15] | 刘会良, 张永宽, 张道远, 尹林克, 张元明. 不同居群准噶尔无叶豆果实和种子特性及种子萌发差异[J]. 植物生态学报, 2012, 36(8): 802-811. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
Copyright © 2026 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19
![]()